
Root-System Architectures of Two Cuban Rice Cultivars with Salt Stress at Early Development Stages


 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Seed Pre-Treatment
2.2. Roll Experiment
2.3. Mini-Rhizotron Experiment
2.4. Root Image Acquisition and Analysis
2.5. Plant P Concentration
2.6. Statistical Analyses
3. Results
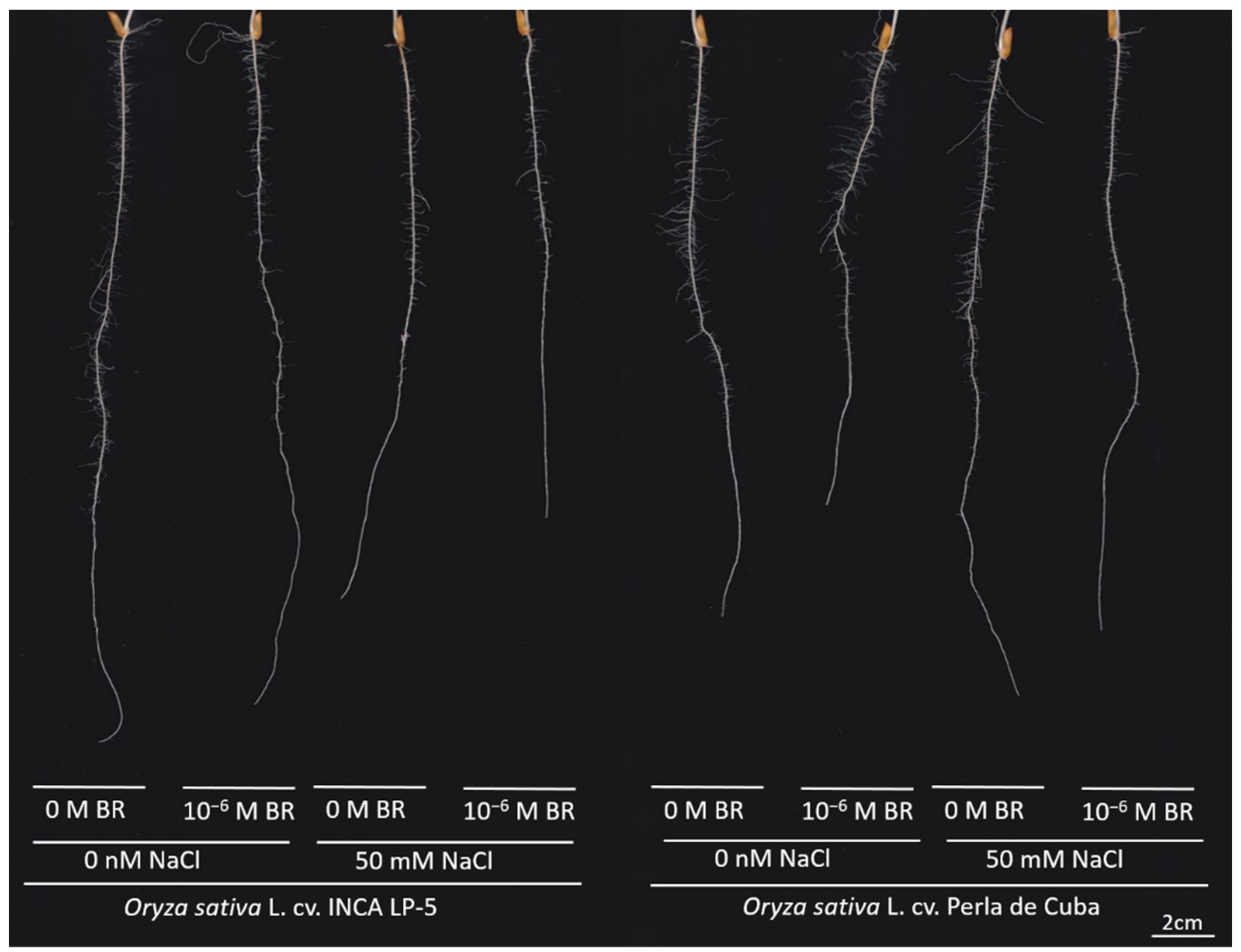
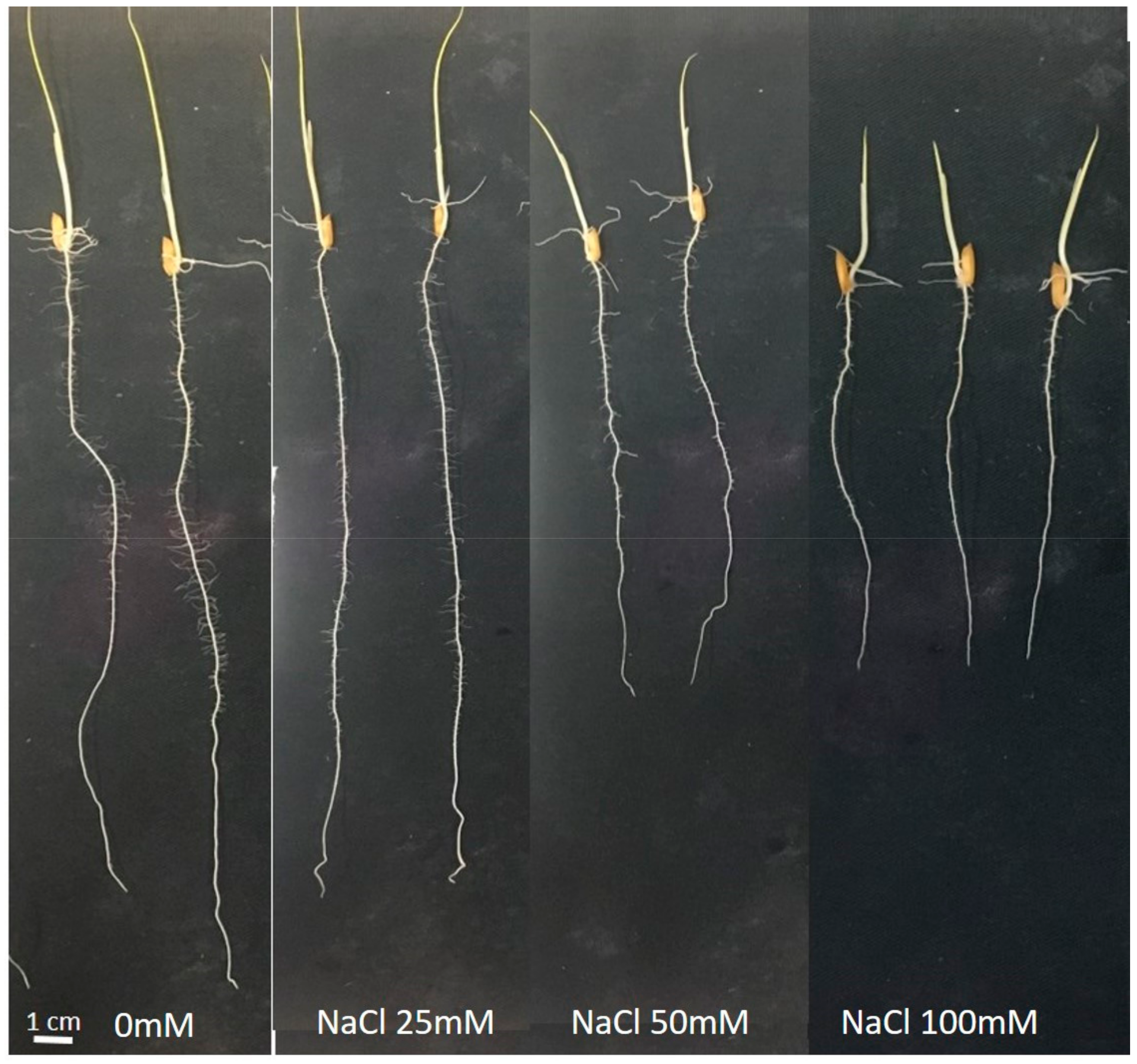
3.1. Roll Experiment
3.2. Mini-Rhizotron Experiment–Biomass and P Concentration
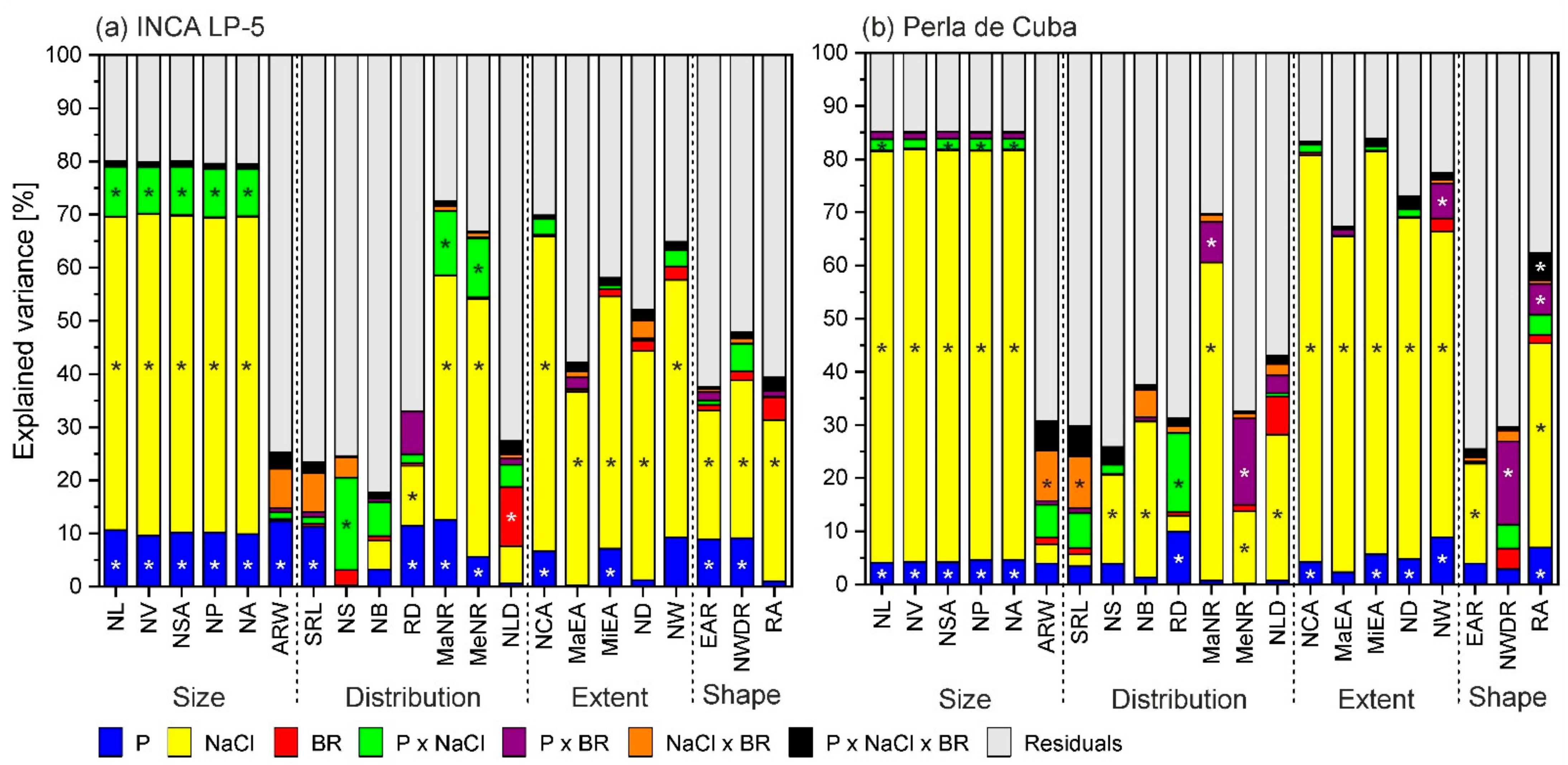
3.3. Root-System Traits in Mini-Rhizotrons
4. Discussion
4.1. Strong Effect of Salt Stress on Rice Growth and Root Architecture
4.2. Differences between the Genotypes
4.3. The Role of P and BR Interrelated to Salt Stress
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 mM NaCl | 50 mM NaCl | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 10 ppm P | 1 ppm P | 10 ppm P | 1 ppm P | |||||||
| 0 M BR | 10−6 M BR | 0 M BR | 10−6 M BR | 0 M BR | 10−6 M BR | 0 M BR | 10−6 M BR | Significance | ||
| NL | INCA LP-5 | 756 ± 241 | 779 ± 33.6 | 466 ± 134 | 388 ± 148 | 224 ± 135 | 152 ± 91.4 | 150 ± 67.3 | 204 ± 117 | P ***, NaCl ***, P × NaCl *** |
| [cm] | Perla de Cuba | 420 ± 34.3 | 378 ± 115 | 286 ± 51.1 | 310 ± 73.1 | 128 ± 18.6 | 74.5 ± 35.4 | 74.5 ± 60.6 | 94.4 ± 60.2 | P **, NaCl ***, P × NaCl * |
| NV | INCA LP-5 | 0.21 ± 0.06 | 0.21 ± 0.01 | 0.13 ± 0.03 | 0.11 ± 0.04 | 0.06 ± 0.04 | 0.04 ± 0.03 | 0.04 ± 0.02 | 0.06 ± 0.03 | P ***, NaCl ***, P × NaCl ** |
| [cm3] | Perla de Cuba | 0.11 ± 0.01 | 0.11 ± 0.03 | 0.08 ± 0.02 | 0.09 ± 0.02 | 0.04 ± 0.01 | 0.02 ± 0.01 | 0.02 ± 0.02 | 0.03 ± 0.02 | P **, NaCl *** |
| NSA | INCA LP-5 | 43.3 ± 13.6 | 44.6 ± 2.13 | 26.8 ± 7.32 | 22.8 ± 8.61 | 13.0 ± 7.88 | 8.77 ± 5.35 | 8.81 ± 4.01 | 11.8 ± 6.82 | P ***, NaCl ***, P × NaCl *** |
| [cm2] | Perla de Cuba | 23.9 ± 2.06 | 21.9 ± 6.33 | 16.6 ± 3.09 | 17.8 ± 4.25 | 7.59 ± 1.19 | 4.31 ± 2.07 | 4.32 ± 3.54 | 5.42 ± 3.48 | P **, NaCl ***, P × NaCl * |
| NP | INCA LP-5 | 1084 ± 346 | 1111 ± 56.1 | 665 ± 179 | 575 ± 214 | 327 ± 203 | 221 ± 136 | 224 ± 100 | 297 ± 173 | P ***, NaCl ***, P × NaCl *** |
| [cm] | Perla de Cuba | 595 ± 57.5 | 554 ± 155 | 413 ± 79.0 | 440 ± 103 | 195 ± 32.9 | 110 ± 51.7 | 111 ± 90.3 | 136 ± 86.5 | P **, NaCl ***, P × NaCl * |
| NA | INCA LP-5 | 9.49 ± 3.00 | 9.72 ± 0.53 | 5.86 ± 1.51 | 5.12 ± 1.90 | 2.90 ± 1.80 | 1.94 ± 1.21 | 1.99 ± 0.91 | 2.62 ± 1.52 | P ***, NaCl ***, P × NaCl ** |
| [cm2] | Perla de Cuba | 5.19 ± 0.51 | 4.89 ± 1.32 | 3.65 ± 0.72 | 3.87 ± 0.92 | 1.74 ± 0.31 | 0.96 ± 0.46 | 0.97 ± 0.80 | 1.19 ± 0.77 | P **, NaCl ***, P × NaCl * |
| ARW | INCA LP-5 | 0.18 ± 0.00 | 0.18 ± 0.00 | 0.18 ± 0.00 | 0.19 ± 0.00 | 0.18 ± 0.00 | 0.18 ± 0.01 | 0.19 ± 0.00 | 0.18 ± 0.00 | P * |
| [mm] | Perla de Cuba | 0.18 ± 0.00 | 0.19 ± 0.00 | 0.18 ± 0.00 | 0.18 ± 0.00 | 0.19 ± 0.01 | 0.18 ± 0.00 | 0.18 ± 0.00 | 0.18 ± 0.00 | NaCl × BR * |
| SRL | INCA LP-5 | 3670 ± 144 | 3687 ± 75.7 | 3595 ± 177 | 3430 ± 121 | 3581 ± 153 | 3689 ± 307 | 3481 ± 168 | 3623 ± 216 | P * |
| [cm cm3] | Perla de Cuba | 3717 ± 57.8 | 3542 ± 184 | 3585 ± 90.3 | 3628 ± 92.5 | 3405 ± 232 | 3590 ± 134 | 3597 ± 148 | 3687 ± 196 | NaClxBR * |
| NS | INCA LP-5 | 8.66 ± 1.09 | 8.79 ± 1.31 | 7.39 ± 1.42 | 7.09 ± 1.27 | 6.64 ± 1.46 | 7.71 ± 2.20 | 7.81 ± 2.29 | 9.17 ± 1.54 | PxNaCl * |
| [%] | Perla de Cuba | 9.70 ± 1.51 | 9.31 ± 1.36 | 9.35 ± 2.33 | 10.2 ± 1.53 | 10.2 ± 1.47 | 11.2 ± 2.29 | 12.5 ± 3.26 | 11.7 ± 1.94 | NaCl * |
| NB | INCA LP-5 | 2.15 ± 0.53 | 2.27 ± 0.13 | 2.22 ± 0.52 | 2.38 ± 1.19 | 2.65 ± 0.54 | 2.97 ± 0.76 | 2.35 ± 0.37 | 2.21 ± 0.34 | ns |
| [n n−1] | Perla de Cuba | 3.03 ± 1.45 | 4.00 ± 0.91 | 3.20 ± 1.10 | 3.32 ± 1.11 | 2.54 ± 0.98 | 2.05 ± 0.58 | 2.27 ± 1.01 | 1.82 ± 0.39 | NaCl *** |
| RD | INCA LP-5 | 15.3 ± 2.71 | 16.8 ± 2.60 | 14.4 ± 1.68 | 13.5 ± 1.58 | 13.1 ± 2.10 | 14.8 ± 2.22 | 13.5 ± 1.61 | 12.5 ± 2.06 | P *, NaCl * |
| [n cm−1] | Perla de Cuba | 14.4 ± 2.26 | 14.2 ± 1.47 | 11.9 ± 1.30 | 10.4 ± 1.75 | 11.9 ± 2.77 | 11.7 ± 1.07 | 11.8 ± 2.59 | 12.3 ± 2.68 | P *, P *, NaCl * |
| MaNR | INCA LP-5 | 49.6 ± 13.5 | 48.6 ± 10.8 | 31.2 ± 9.76 | 24.2 ± 6.18 | 18.4 ± 9.74 | 17.2 ± 6.94 | 15.2 ± 4.76 | 20.0 ± 5.48 | P ***, NaCl ***, P × NaCl *** |
| [n] | Perla de Cuba | 46.6 ± 5.18 | 42.6 ± 13.9 | 37.2 ± 8.26 | 47.8 ± 11.4 | 28.4 ± 7.13 | 15.6 ± 3.21 | 16.4 ± 9.18 | 21.8 ± 10.4 | NaCl ***, P × BR ** |
| MeNR | INCA LP-5 | 23.6 ± 7.50 | 21.4 ± 4.62 | 14.8 ± 5.93 | 12.4 ± 7.40 | 7.00 ± 3.54 | 5.80 ± 2.39 | 6.80 ± 2.95 | 9.00 ± 1.83 | P *, NaCl ***, P × NaCl ** |
| [n] | Perla de Cuba | 18.8 ± 10.1 | 10.6 ± 2.88 | 12.6 ± 4.93 | 15.8 ± 6.30 | 12.2 ± 4.32 | 8.00 ± 2.55 | 7.80 ± 4.82 | 11.8 ± 5.22 | NaCl *, P × BR ** |
| NLD | INCA LP-5 | 0.91 ± 0.46 | 1.17 ± 0.46 | 1.06 ± 0.46 | 1.24 ± 0.50 | 1.38 ± 0.55 | 1.52 ± 0.07 | 0.90 ± 0.31 | 1.51 ± 0.34 | BR * |
| [n n−1] | Perla de Cuba | 1.14 ± 0.92 | 0.72 ± 0.30 | 1.22 ± 0.59 | 0.66 ± 0.53 | 0.11 ± 0.14 | 0.33 ± 0.40 | 0.67 ± 0.43 | 0.16 ± 0.25 | NaCl *** |
| NCA | INCA LP-5 | 111 ± 35.2 | 113 ± 19.2 | 82.3 ± 27.8 | 73.1 ± 26.4 | 43.2 ± 22.6 | 30.2 ± 22.2 | 28.3 ± 13.5 | 31.4 ± 23.1 | P *, NaCl *** |
| [cm2] | Perla de Cuba | 54.1 ± 5.55 | 52.3 ± 11.3 | 41.1 ± 13.6 | 39.0 ± 13.6 | 17.1 ± 2.46 | 9.18 ± 4.78 | 9.63 ± 9.50 | 9.81 ± 5.68 | P **, NaCl *** |
| MaEA | INCA LP-5 | 13.6 ± 1.55 | 13.7 ± 1.96 | 13.4 ± 1.62 | 13.8 ± 3.28 | 11.5 ± 2.78 | 8.72 ± 3.22 | 9.19 ± 2.42 | 9.91 ± 3.93 | NaCl *** |
| [cm] | Perla de Cuba | 8.74 ± 1.36 | 8.57 ± 1.52 | 7.77 ± 1.54 | 7.91 ± 1.79 | 5.40 ± 0.58 | 4.27 ± 1.66 | 3.98 ± 1.73 | 4.50 ± 1.39 | NaCl *** |
| MiEA | INCA LP-5 | 7.40 ± 2.20 | 7.35 ± 1.40 | 6.44 ± 1.92 | 4.97 ± 1.18 | 3.98 ± 2.13 | 3.45 ± 2.01 | 2.93 ± 1.16 | 2.78 ± 1.10 | P *, NaCl *** |
| [cm] | Perla de Cuba | 5.77 ± 0.26 | 5.85 ± 0.83 | 4.83 ± 1.07 | 4.53 ± 0.62 | 2.86 ± 0.32 | 2.17 ± 0.78 | 1.84 ± 1.07 | 2.19 ± 0.78 | P **, NaCl *** |
| ND | INCA LP-5 | 16.0 ± 1.49 | 17.0 ± 2.64 | 16.1 ± 2.39 | 15.8 ± 2.69 | 13.8 ± 2.96 | 9.85 ± 4.09 | 11.2 ± 2.71 | 10.2 ± 3.87 | NaCl *** |
| [cm] | Perla de Cuba | 11.7 ± 2.07 | 13.0 ± 1.15 | 10.5 ± 2.34 | 9.41 ± 2.47 | 6.25 ± 0.74 | 5.64 ± 2.59 | 4.59 ± 2.14 | 5.88 ± 1.84 | P *, NaCl *** |
| NW | INCA LP-5 | 10.7 ± 2.30 | 10.4 ± 2.95 | 8.40 ± 3.09 | 6.11 ± 1.47 | 5.32 ± 2.27 | 3.81 ± 2.45 | 3.84 ± 1.39 | 3.54 ± 1.33 | P *, NaCl *** |
| [cm] | Perla de Cuba | 8.28 ± 1.11 | 7.25 ± 1.23 | 6.23 ± 1.10 | 6.56 ± 1.33 | 5.66 ± 1.11 | 2.76 ± 1.10 | 2.43 ± 1.68 | 3.06 ± 1.31 | P **, NaCl ***, P × BR ** |
| EAR | INCA LP-5 | 0.55 ± 0.16 | 0.55 ± 0.19 | 0.49 ± 0.16 | 0.37 ± 0.07 | 0.36 ± 0.21 | 0.37 ± 0.13 | 0.31 ± 0.08 | 0.28 ± 0.05 | NaCl ** |
| [cm cm−1] | Perla de Cuba | 0.68 ± 0.12 | 0.70 ± 0.15 | 0.64 ± 0.16 | 0.59 ± 0.10 | 0.54 ± 0.09 | 0.54 ± 0.16 | 0.44 ± 0.13 | 0.53 ± 0.25 | NaCl ** |
| NWDR | INCA LP-5 | 0.67 ± 0.15 | 0.64 ± 0.27 | 0.53 ± 0.18 | 0.39 ± 0.08 | 0.39 ± 0.15 | 0.35 ± 0.13 | 0.33 ± 0.07 | 0.35 ± 0.02 | P *, NaCl *** |
| [cm cm−1] | Perla de Cuba | 0.73 ± 0.14 | 0.56 ± 0.09 | 0.60 ± 0.09 | 0.72 ± 0.19 | 0.91 ± 0.18 | 0.54 ± 0.20 | 0.52 ± 0.32 | 0.58 ± 0.35 | PxBR * |
| RA | INCA LP-5 | 109 ± 31.0 | 95.1 ± 11.6 | 103 ± 29.5 | 82.7 ± 10.5 | 76.6 ± 19.1 | 51.6 ± 49.9 | 57.3 ± 32.3 | 64.0 ± 7.16 | NaCl *** |
| [°] | Perla de Cuba | 112 ± 13.3 | 107 ± 14.3 | 106 ± 13.7 | 103 ± 17.4 | 103 ± 15.4 | 50.6 ± 37.2 | 31.3 ± 45.3 | 51.7 ± 32.7 | NaCl ***, P *, P × BR *, P × NaCl × BR * |


References
- Hussain, S.; Zhang, J.; Zhong, C.; Zhu, L.-F.; Cao, X.-C.; Yu, S.-M.; Bohr, J.A.; Hu, J.-J.; Jin, Q.-Y. Effects of salt stress on rice growth, development characteristics, and the regulating ways: A review. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef]
- Hoang, T.M.L.; Tran, T.N.; Nguyen, T.K.T.; Williams, B.; Wurm, P.; Bellairs, S.; Mundree, S. Improvement of Salinity Stress Tolerance in Rice: Challenges and Opportunities. Agronomy 2016, 6, 54. [Google Scholar] [CrossRef]
- Oficina Nacional de Estadística e Información (ONEI) Medio Ambiente. Anuario Estadístico de Cuba; La Habana, Cuba, 2019; pp. 7–62. Available online: http://www.onei.gob.cu/ (accessed on 18 May 2021).
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High Salinity Induces Different Oxidative Stress and Antioxidant Responses in Maize Seedlings Organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef]
- Akram, S.; Siddiqui, N.; Hussain, B.M.N.; Al Bari, A.; Mostofa, M.G.; Hossain, M.A.; Tran, L.-S.P. Exogenous Glutathione Modulates Salinity Tolerance of Soybean [Glycine max (L.) Merrill] at Reproductive Stage. J. Plant Growth Regul. 2017, 36, 877–888. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced Senescence in Leaves of Rice (Oryza sativa L.) Cultivars Differing in Salinity Resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Ganie, S.A.; Molla, K.A.; Henry, R.; Bhat, K.V.; Mondal, T.K. Advances in understanding salt tolerance in rice. Theor. Appl. Genet. 2019, 132, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Gregorio, G.; Senadhira, D.; Mendoza, R.; Manigbas, N.; Roxas, J.; Guerta, C. Progress in breeding for salinity tolerance and associated abiotic stresses in rice. Field Crop. Res. 2002, 76, 91–101. [Google Scholar] [CrossRef]
- Rahman, A.; Nahar, K.; Al Mahmud, J.; Hasanuzzaman, M.; Hossain, S.; Fujita, M. Salt Stress Tolerance in Rice: Emerging Role of Exogenous Phytoprotectants. Adv. Int. Rice Res. 2017, 15, 139–174. [Google Scholar] [CrossRef]
- Kumar, V.; Khare, T. Differential growth and yield responses of salt-tolerant and susceptible rice cultivars to individual (Na+ and Cl−) and additive stress effects of NaCl. Acta Physiol. Plant. 2016, 38, 170. [Google Scholar] [CrossRef]
- Ologundudu, A.F.; Adelusi, A.A.; Akinwale, R.O. Effect of Salt Stress on Germination and Growth Parameters of Rice (Oryza sativa L.). Not. Sci. Biol. 2014, 6, 6. [Google Scholar] [CrossRef]
- Rahman, A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium Supplementation Improves Na+/K+ Ratio, Antioxidant Defense and Glyoxalase Systems in Salt-Stressed Rice Seedlings. Front. Plant Sci. 2016, 7, 609. [Google Scholar] [CrossRef] [PubMed]
- Naidu, R.; Rengasamy, P. Ion interactions and constraints to plant nutrition in Australian sodic soils. Soil Res. 1993, 31, 801–819. [Google Scholar] [CrossRef]
- Russell, E.J.; Wild, A.; Russell, E.J. Russell’s Soil Conditions and Plant Growth; Longman Scientific & Technical; John Wiley & Sons: Harlow, UK, 1988; ISBN 978-0-470-20796-3. [Google Scholar]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New Insights on Plant Salt Tolerance Mechanisms and Their Potential Use for Breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef]
- Van Nguyen, L.; Bertero, D.; Nguyen, L.V. Genetic variation in root development responses to salt stresses of quinoa. J. Agron. Crop. Sci. 2020, 206, 538–547. [Google Scholar] [CrossRef]
- Ben Abdallah, H.; Mai, H.-J.; Álvarez-Fernández, A.; Abadía, J.; Bauer, P. Natural variation reveals contrasting abilities to cope with alkaline and saline soil among different Medicago truncatula genotypes. Plant Soil 2017, 418, 45–60. [Google Scholar] [CrossRef]
- Wang, H.; Liang, L.; Liu, S.; An, T.; Fang, Y.; Xu, B.; Zhang, S.; Deng, X.; Palta, J.A.; Siddique, K.H.M.; et al. Maize genotypes with deep root systems tolerate salt stress better than those with shallow root systems during early growth. J. Agron. Crop. Sci. 2020, 206, 711–721. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, B.; Li, J.; Song, Z.; Lu, B.; Chi, M.; Yang, B.; Liu, J.; Lam, Y.-W.; Li, J.; et al. Salt-response analysis in two rice cultivars at seedling stage. Acta Physiol. Plant. 2017, 39, 1–9. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- Tang, H.; Niu, L.; Wei, J.; Chen, X.; Chen, Y. Phosphorus Limitation Improved Salt Tolerance in Maize Through Tissue Mass Density Increase, Osmolytes Accumulation, and Na+ Uptake Inhibition. Front. Plant Sci. 2019, 10, 856. [Google Scholar] [CrossRef]
- de Dorlodot, S.; Forster, B.; Pagès, L.; Price, A.; Tuberosa, R.; Draye, X. Root system architecture: Opportunities and constraints for genetic improvement of crops. Trends Plant Sci. 2007, 12, 474–481. [Google Scholar] [CrossRef]
- Jung, J.K.H.M.; McCouch, S.R.M. Getting to the roots of it: Genetic and hormonal control of root architecture. Front. Plant Sci. 2013, 4, 186. [Google Scholar] [CrossRef]
- Krishnamurthy, P.; Ranathunge, K.; Nayak, S.; Schreiber, L.; Mathew, M.K. Root apoplastic barriers block Na+ transport to shoots in rice (Oryza sativa L.). J. Exp. Bot. 2011, 62, 4215–4228. [Google Scholar] [CrossRef]
- Toyofuku, K.; Matsunami, M.; Ogawa, A. Genotypic Variation in Osmotic Stress Tolerance Among Rice Cultivars and Its Association with L-Type Lateral Root Development. Plant Prod. Sci. 2015, 18, 246–253. [Google Scholar] [CrossRef]
- Vejchasarn, P.; Lynch, J.P.; Brown, K.M. Genetic Variability in Phosphorus Responses of Rice Root Phenotypes. Rice 2016, 9, 29. [Google Scholar] [CrossRef]
- Lynch, J.P.; Brown, K.M. Topsoil Foraging—An Architectural Adaptation of Plants to Low Phosphorus Availability. Plant Soil 2001, 237, 225–237. [Google Scholar] [CrossRef]
- Sima, N.A.K.K.; Ahmad, S.T.; Alitabar, R.A.; Mottaghi, A.; Pessarakli, M. Interactive Effects of Salinity and Phosphorus Nutrition on Physiological Responses of Two Barley Species. J. Plant Nutr. 2012, 35, 1411–1428. [Google Scholar] [CrossRef]
- Zribi, O.T.; Abdelly, C.; Debez, A. Interactive effects of salinity and phosphorus availability on growth, water relations, nutritional status and photosynthetic activity of barley (Hordeum vulgare L.). Plant Biol. 2011, 13, 872–880. [Google Scholar] [CrossRef]
- Kawa, D.; Julkowska, M.M.; Sommerfeld, H.M.; Ter Horst, A.; Haring, M.A.; Testerink, C. Phosphate-dependent root system architecture responses to salt stress. Plant Physiol. 2016, 172, 690–706. [Google Scholar] [CrossRef]
- Divi, U.K.; Krishna, P. Brassinosteroid: A biotechnological target for enhancing crop yield and stress tolerance. New Biotechnol. 2009, 26, 131–136. [Google Scholar] [CrossRef]
- Talaat, N.B.; Shawky, B.T. 24-Epibrassinolide alleviates salt-induced inhibition of productivity by increasing nutrients and compatible solutes accumulation and enhancing antioxidant system in wheat (Triticum aestivum L.). Acta Physiol. Plant. 2013, 35, 729–740. [Google Scholar] [CrossRef]
- González-García, M.-P.; Vilarrasa-Blasi, J.; Zhiponova, M.; Divol, F.; Mora-García, S.; Russinova, E.; Caño-Delgado, A.I. Brassinosteroids control meristem size by promoting cell cycle progression in Arabidopsis roots. Development 2011, 138, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Kim, Y.; Pham, G.; Kim, J.W.; Song, J.-H.; Lee, Y.; Hwang, Y.-S.; Roux, S.J.; Kim, S.-H. Brassinazole resistant 1 (BZR1)-dependent brassinosteroid signalling pathway leads to ectopic activation of quiescent cell division and suppresses columella stem cell differentiation. J. Exp. Bot. 2015, 66, 4835–4849. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Tian, H.; Zhang, F.; Liu, J.; Lu, S.; Bai, M.; Li, C.; Ding, Z. Brassinosteroids regulate root growth by controlling reactive oxygen species homeostasis and dual effect on ethylene synthesis in Arabidopsis. PLoS Genet. 2018, 14, e1007144. [Google Scholar] [CrossRef] [PubMed]
- Müssig, C.; Shin, G.-H.; Altmann, T. Brassinosteroids Promote Root Growth in Arabidopsis. Plant Physiol. 2003, 133, 1261–1271. [Google Scholar] [CrossRef]
- Bao, F.; Shen, J.; Brady, S.R.; Muday, G.K.; Asami, T.; Yang, Z. Brassinosteroids Interact with Auxin to Promote Lateral Root Development in Arabidopsis. Plant Physiol. 2004, 134, 1624–1631. [Google Scholar] [CrossRef]
- Anuradha, S.; Rao, S.S.R. Application of brassinosteroids to rice seeds (Oryza sativa L.) reduced the impact of salt stress on growth, prevented photosynthetic pigment loss and increased nitrate reductase activity. Plant Growth Regul. 2003, 40, 29–32. [Google Scholar] [CrossRef]
- Anuradha, S.; Rao, S.S.R. Effect of brassinosteroids on salinity stress induced inhibition of seed germination and seedling growth of rice (Oryza sativa L.). Plant Growth Regul. 2001, 33, 151–153. [Google Scholar] [CrossRef]
- Özdemir, F.; Bor, M.; Demiral, T.; Türkan, I. Effects of 24-epibrassinolide on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress. Plant Growth Regul. 2004, 42, 203–211. [Google Scholar] [CrossRef]
- Wei, Z.; Li, J. Brassinosteroids Regulate Root Growth, Development, and Symbiosis. Mol. Plant 2016, 9, 86–100. [Google Scholar] [CrossRef]
- Guimarães, P.H.R.; De Lima, I.P.; De Castro, A.P.; Lanna, A.C.; Melo, P.G.S.; De Raïssac, M. Phenotyping Root Systems in a Set of Japonica Rice Accessions: Can Structural Traits Predict the Response to Drought? Rice 2020, 13, 1–19. [Google Scholar] [CrossRef]
- Shrestha, R.; Al-Shugeairy, Z.; Al-Ogaidi, F.; Munasinghe, M.; Radermacher, M.; Vandenhirtz, J.; Price, A.H. Comparing simple root phenotyping methods on a core set of rice genotypes. Plant Biol. 2013, 16, 632–642. [Google Scholar] [CrossRef]
- Ali, N.; Yeasmin, L.; Gantait, S.; Goswami, R.; Chakraborty, S. Screening of rice landraces for salinity tolerance at seedling stage through morphological and molecular markers. Physiol. Mol. Biol. Plants 2014, 20, 411–423. [Google Scholar] [CrossRef]
- Rasel, M.; Tahjib-Ul-Arif, M.; Hossain, M.A.; Hassan, L.; Farzana, S.; Brestic, M. Screening of Salt-Tolerant Rice Landraces by Seedling Stage Phenotyping and Dissecting Biochemical Determinants of Tolerance Mechanism. J. Plant Growth Regul. 2020, 1–16. [Google Scholar] [CrossRef]
- Pérez, N.J.; González, M.C.; Cristo, E.; Díaz, S.H.; Díaz, E.C.; Blanco, G. Cultivares Cubanos de Arroz. Inst. Nac. Cienc. Agríc. INCA 2015, 3–24. [Google Scholar]
- Pérez, N.; Castro, R.I. A New Short Cycle Rice Variety: INCA LP-5. Cultiv. Trop. 2000, 21, 55. [Google Scholar]
- Parra-Londono, S.; Kavka, M.; Samans, B.; Snowdon, R.; Wieckhorst, S.; Uptmoor, R. Sorghum root-system classification in contrasting P environments reveals three main rooting types and root-architecture-related marker–trait associations. Ann. Bot. 2018, 121, 267–280. [Google Scholar] [CrossRef]
- Yoshida, S.; Forno, D.A.; Cock, J.H.; Gomez, K.A. Laboratory Manual for Physiological Studies of Rice, 3rd ed.; The International Rice Research Institute: Los Baños, CA, USA, 1976; ISBN 978-971-10-4035-2. [Google Scholar]
- Galkovskyi, T.; Mileyko, Y.; Bucksch, A.; Moore, B.; Symonova, O.; Price, C.A.; Topp, C.N.; Iyer-Pascuzzi, A.S.; Zurek, P.R.; Fang, S.; et al. GiA Roots: Software for the high throughput analysis of plant root system architecture. BMC Plant Biol. 2012, 12, 116. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Lobet, G.; Pagès, L.; Draye, X. A Novel Image-Analysis Toolbox Enabling Quantitative Analysis of Root System Architecture. Plant Physiol. 2011, 157, 29–39. [Google Scholar] [CrossRef]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis. Part 2. In Chemical and Microbiological Properties, 2nd ed.; Soil Science Society of America: Madison, WI, USA, 1982. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; ISBN 3-900051-07-01. [Google Scholar]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Nakazawa, M. Fmsb: Functions for Medical Statistics Book with Some Demographic Data. R Package Version 0.7.0. 2019. Available online: https://rdrr.io/cran/fmsb/ (accessed on 18 May 2021).
- Topp, C.N.; Iyer-Pascuzzi, A.; Anderson, J.; Lee, C.-R.; Zurek, P.R.; Symonova, O.; Zheng, Y.; Bucksch, A.; Mileyko, Y.; Galkovskyi, T.; et al. 3D phenotyping and quantitative trait locus mapping identify core regions of the rice genome controlling root architecture. Proc. Natl. Acad. Sci. USA 2013, 110, E1695–E1704. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-S.; Choi, W.-Y.; Ko, J.-C.; Kim, T.-S.; Gregorio, G.B. Salinity tolerance of japonica and indica rice (Oryza sativa L.) at the seedling stage. Planta 2003, 216, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Shannon, M.C.; Lesch, S.M. Timing of salinity stress affects rice growth and yield components. Agric. Water Manag. 2001, 48, 191–206. [Google Scholar] [CrossRef]
- Hakim, M.A.; Juraimi, A.S.; Begum, M.; Hanafi, M.M.; Ismail, M.R.; Selamat, A. Effect of salt stress on germination and early seedling growth of rice (Oryza sativa L.). Afr. J. Biotechnol. 2010, 9, 1911–1918. [Google Scholar] [CrossRef]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Rewald, B.; Shelef, O.; Ephrath, J.E.; Rachmilevitch, S. Adaptive Plasticity of Salt-Stressed Root Systems. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M., Prasad, M., Eds.; Springer Science and Business Media LLC: New York, NY, USA, 2013; pp. 169–201. [Google Scholar] [CrossRef]
- Samarajeewa, P.K.; Barrero, R.A.; Umeda-Hara, C.; Kawai, M.; Uchimiya, H. Cortical cell death, cell proliferation, macromolecular movements and rTip1 expression pattern in roots of rice (Oryza sativa L.) under NaCl stress. Planta 1999, 207, 354–361. [Google Scholar] [CrossRef]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Kano, M.; Inukai, Y.; Kitano, H.; Yamauchi, A. Root plasticity as the key root trait for adaptation to various intensities of drought stress in rice. Plant Soil 2011, 342, 117–128. [Google Scholar] [CrossRef]
- Albacete, A.; Ghanem, M.E.; Martínez-Andújar, C.; Acosta, M.; Sánchez-Bravo, J.; Martinez, V.; Lutts, S.; Dodd, I.C.; Pérez-Alfocea, F. Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 2008, 59, 4119–4131. [Google Scholar] [CrossRef]
- Maggio, A.; Hasegawa, P.M.; Bressan, R.A.; Consiglio, M.F.; Joly, R.J. Review: Unravelling the functional relationship between root anatomy and stress tolerance. Funct. Plant Biol. 2001, 28, 999. [Google Scholar] [CrossRef]
- Nibau, C.; Gibbs, D.J.; Coates, J.C. Branching out in new directions: The control of root architecture by lateral root formation. New Phytol. 2008, 179, 595–614. [Google Scholar] [CrossRef]
- Bernstein, N.; Kafkafi, U. Root Growth Under Salinity Stress. Plant Roots 2002, 787–805. [Google Scholar] [CrossRef]
- Waziri, A.; Kumar, P.; Purty, R.S. Saltol QTL and Their Role in Salinity Tolerance in Rice. Austin. J. Biotechnol. Bioeng. 2016, 3, 1–5. [Google Scholar]
- Bargaz, A.; Nassar, R.M.A.; Rady, M.M.; Gaballah, M.S.; Thompson, S.M.; Brestic, M.; Schmidhalter, U.; Abdelhamid, M.T. Improved salinity tolerance by phosphorus fertilizer in two Phaseolus vulgaris recombinant inbred lines contrasting in their P-efficiency. J. Agron. Crop. Sci. 2016, 202, 497–507. [Google Scholar] [CrossRef]
- Li, J.; Xie, Y.; Dai, A.; Liu, L.; Li, Z. Root and shoot traits responses to phosphorus deficiency and QTL analysis at seedling stage using introgression lines of rice. J. Genet. Genom. 2009, 36, 173–183. [Google Scholar] [CrossRef]
- Shimizu, A.; Yanagihara, S.; Kawasaki, S.; Ikehashi, H. Phosphorus deficiency-induced root elongation and its QTL in rice (Oryza sativa L.). Theor. Appl. Genet. 2004, 109, 1361–1368. [Google Scholar] [CrossRef]
- Lambers, H.; Finnegan, P.M.; Laliberté, E.; Pearse, S.J.; Ryan, M.H.; Shane, M.W.; Veneklaas, E.J. Phosphorus Nutrition of Proteaceae in Severely Phosphorus-Impoverished Soils: Are There Lessons to Be Learned for Future Crops? Plant Physiol. 2011, 156, 1058–1066. [Google Scholar] [CrossRef]
- Jin, J.; Tang, C.; Armstrong, R.; Sale, P. Phosphorus supply enhances the response of legumes to elevated CO2 (FACE) in a phosphorus-deficient vertisol. Plant Soil 2012, 358, 91–104. [Google Scholar] [CrossRef]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2012, 112, 391–408. [Google Scholar] [CrossRef]
- Arora, N.; Bhardwaj, R.; Sharma, P.; Arora, H.K. Effects of 28-homobrassinolide on growth, lipid peroxidation and antioxidative enzyme activities in seedlings of Zea mays L. under salinity stress. Acta Physiol. Plant. 2008, 30, 833–839. [Google Scholar] [CrossRef]
- Vázquez-Glaría, A.; Duvergel-Terry, A.; Ortega-Rodés, P.; Ortega, E.; Coll, Y.; Loiret, F.G. El análogo de brasinoesteroides DI-31 varía la arquitectura de la raíz en los cultivares de interés comercial INCA LP-5 y Perla de Cuba de Oryza sativa L. (Poaceae)/The brassinosteroid analog DI-31 modify the root architecture of commercial cultivars INCA LP-5 and Perla de Cuba of Oryza sativa L. (Poaceae). Rev. Cuba. Cienc. Biológicas 2019, 7, 1–8. [Google Scholar]





| Category | Root trait | Abbreviation | Unit |
|---|---|---|---|
| Size | Network length | NL | cm |
| Network volume | NV | cm3 | |
| Network surface area | NSA | cm2 | |
| Network perimeter | NP | cm | |
| Network area | NA | cm2 | |
| Average root width | ARW | mm | |
| Distribution | Specific root length | SRL | cm cm−3 |
| Network solidity | NS | % | |
| Network bushiness | NB | n n−1 | |
| Lateral root density | RD | n cm−1 | |
| Maximum number of roots | MaNR | n | |
| Median number of roots | MeNR | n | |
| Network length distribution | NLD | n n−1 | |
| Extent | Network convex area | NCA | cm2 |
| Major ellipse axis | MaEA | cm | |
| Minor ellipse axis | MiEA | cm | |
| Network depth | ND | cm | |
| Network width | NW | cm | |
| Shape | Ellipse axis ratio | EAR | cm cm−1 |
| Network width-to-depth ratio | NWDR | cm cm−1 | |
| Root angle | RA | ° |
| 0 mM NaCl | 50 mM NaCl | |||||
|---|---|---|---|---|---|---|
| 0 M BR | 10−6 M BR | 0 M BR | 10−6 M BR | Significances | ||
| Shoot fresh weight | INCA LP-5 | 77.7 ± 5.39 | 72.9 ± 7.21 | 47.2 ± 8.23 | 53.7 ± 96.4 | NaCl *** |
| [mg] | Perla de Cuba | 51.1 ± 10.6 ** | 58.9 ± 4.23 ** | 52.3 ± 15 0 | 58.9 ± 5.31 | |
| Root fresh weight | INCA LP-5 | 21.1 ± 2.26 | 19.5 ± 1.26 | 11.5 ± 4.40 | 12.6 ± 3.28 | NaCl *** |
| [mg] | Perla de Cuba | 16.1 ± 6.11 | 14.8 ± 5.23 | 16.3 ± 7.52 | 13.8 ± 4.06 | |
| Root-to-shoot ratio | INCA LP-5 | 0.27 ± 0.03 | 0.27 ± 0.04 | 0.24 ± 0.05 | 0.23 ± 0.04 | |
| [mg mg−1] | Perla de Cuba | 0.31 ± 0.10 | 0.25 ± 0.09 | 0.30 ± 0.07 | 0.23 ± 0.06 | |
| Primary root length | INCA LP-5 | 19.0 ± 3.47 | 19.7 ± 4.01 | 13.3 ± 6.65 | 12.5 ± 6.52 | NaCl *** |
| [cm] | Perla de Cuba | 13.5 ± 4.46 *** | 9.67 ± 4.06 *** | 15.8 ± 7.51 | 12.8 ± 5.86 | BR * |
| Lateral root density | INCA LP-5 | 14.4 ± 2.06 | 11.8 ± 1.53 | 10.8 ± 3.95 | 11.7 ± 2.02 | NaCl **, BR * |
| [cm−1] | Perla de Cuba | 14.5 ± 1.96 | 14.2 ± 1.51 *** | 12.2 ± 2.46 | 11.5 ± 2.03 | NaCl *** |
| Network length | INCA LP-5 | 80.6 ± 25.6 | 75.7 ± 19.4 | 41.1 ± 33.5 | 33.5 ± 18.8 | NaCl *** |
| [cm] | Perla de Cuba | 61.5 ± 25.9 | 42.1 ± 20.0 *** | 51.0 ± 28.2 | 34.9 ± 18.1 | BR ** |
| 0 mM NaCl | 50 mM NaCl | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 10 ppm P | 1 ppm P | 10 ppm P | 1 ppm P | |||||||
| 0 M BR | 10−6 M BR | 0 M BR | 10−6 M BR | 0 M BR | 10−6 M BR | 0 M BR | 10−6 M BR | Significance | ||
| NL | INCA LP-5 | 756 ± 241 | 779 ± 33.6 | 466 ± 134 | 388 ± 148 | 224 ± 135 | 152 ± 91.4 | 150 ± 67.3 | 204 ± 117 | P ***, NaCl ***, P × NaCl *** |
| [cm] | Perla de Cuba | 420 ± 34.3 * | 378 ± 115 *** | 286 ± 51.1 * | 310 ± 73.1 | 128 ± 18.6 | 74.5 ± 35.4 | 74.5 ± 60.6 | 94.4 ± 60.2 * | P **, NaCl ***, P × NaCl * |
| ARW | INCA LP-5 | 0.18 ± 0.00 | 0.18 ± 0.00 | 0.18 ± 0.00 | 0.19 ± 0.00 | 0.18 ± 0.00 | 0.18 ± 0.01 | 0.19 ± 0.00 | 0.18 ± 0.00 | P * |
| [mm] | Perla de Cuba | 0.18 ± 0.00 | 0.19 ± 0.00 | 0.18 ± 0.00 | 0.18 ± 0.00 * | 0.19 ± 0.01 | 0.18 ± 0.00 | 0.18 ± 0.00 | 0.18 ± 0.00 | NaCl × BR * |
| NS | INCA LP-5 | 8.66 ± 1.09 | 8.79 ± 1.31 | 7.39 ± 1.42 | 7.09 ± 1.27 | 6.64 ± 1.46 | 7.71 ± 2.20 | 7.81 ± 2.29 | 9.17 ± 1.54 | P × NaCl * |
| [%] | Perla de Cuba | 9.70 ± 1.51 | 9.31 ± 1.36 | 9.35 ± 2.33 | 10.2 ± 1.53 ** | 10.2 ± 1.47 ** | 11.2 ± 2.29 * | 12.5 ± 3.26 * | 11.7 ± 1.94 | NaCl * |
| NB | INCA LP-5 | 2.15 ± 0.53 | 2.27 ± 0.13 | 2.22 ± 0.52 | 2.38 ± 1.19 | 2.65 ± 0.54 | 2.97 ± 0.76 | 2.35 ± 0.37 | 2.21 ± 0.34 | ns |
| [n n−1] | Perla de Cuba | 3.03 ± 1.45 | 4.00 ± 0.91 ** | 3.20 ± 1.10 | 3.32 ± 1.11 | 2.54 ± 0.98 | 2.05 ± 0.58 | 2.27 ± 1.01 | 1.82 ± 0.39 | NaCl *** |
| RD | INCA LP-5 | 15.3 ± 2.71 | 16.8 ± 2.60 | 14.4 ± 1.68 | 13.5 ± 1.58 | 13.1 ± 2.10 | 14.8 ± 2.22 | 13.5 ± 1.61 | 12.5 ± 2.06 | P *, NaCl * |
| [n cm−1] | Perla de Cuba | 14.4 ± 2.26 | 14.2 ± 1.47 | 11.9 ± 1.30 * | 10.4 ± 1.75 * | 11.9 ± 2.77 | 11.7 ± 1.07 * | 11.8 ± 2.59 | 12.3 ± 2.68 | P *, P *NaCl * |
| MaNR | INCA LP-5 | 49.6 ± 13.5 | 48.6 ± 10.8 | 31.2 ± 9.76 | 24.2 ± 6.18 | 18.4 ± 9.74 | 17.2 ± 6.94 | 15.2 ± 4.76 | 20.0 ± 5.48 | P ***, NaCl ***, P × NaCl *** |
| [n] | Perla de Cuba | 46.6 ± 5.18 | 42.6 ± 13.9 | 37.2 ± 8.26 | 47.8 ± 11.4 ** | 28.4 ± 7.13 | 15.6 ± 3.21 | 16.4 ± 9.18 | 21.8 ± 10.4 | NaCl ***, PxBR ** |
| NLD | INCA LP-5 | 0.91 ± 0.46 | 1.17 ± 0.46 | 1.06 ± 0.46 | 1.24 ± 0.50 | 1.38 ± 0.55 | 1.52 ± 0.07 | 0.90 ± 0.31 | 1.51 ± 0.34 | BR * |
| [n n−1] | Perla de Cuba | 1.14 ± 0.92 | 0.72 ± 0.30 | 1.22 ± 0.59 | 0.66 ± 0.53 | 0.11 ± 0.14 ** | 0.33 ± 0.40 *** | 0.67 ± 0.43 | 0.16 ± 0.25 *** | NaCl *** |
| NCA | INCA LP-5 | 111 ± 35.2 | 113 ± 19.2 | 82.3 ± 27.8 | 73.1 ± 26.4 | 43.2 ± 22.6 | 30.2 ± 22.2 | 28.3 ± 13.5 | 31.4 ± 23.1 | P *, NaCl *** |
| [cm2] | Perla de Cuba | 54.1 ± 5.55 ** | 52.3 ± 11.3 *** | 41.1 ± 13.6 * | 39.0 ± 13.6 * | 17.1 ± 2.46 * | 9.18 ± 4.78 | 9.63 ± 9.50 * | 9.81 ± 5.68 | P **, NaCl *** |
| ND | INCA LP-5 | 16.0 ± 1.49 | 17.0 ± 2.64 | 16.1 ± 2.39 | 15.8 ± 2.69 | 13.8 ± 2.96 | 9.85 ± 4.09 | 11.2 ± 2.71 | 10.2 ± 3.87 | NaCl *** |
| [cm] | Perla de Cuba | 11.7 ± 2.07 ** | 13.0 ± 1.15 * | 10.5 ± 2.34 ** | 9.41 ± 2.47 ** | 6.25 ± 0.74 ** | 5.64 ± 2.59 | 4.59 ± 2.14 ** | 5.88 ± 1.84 | P *, NaCl *** |
| NW | INCA LP-5 | 10.7 ± 2.30 | 10.4 ± 2.95 | 8.40 ± 3.09 | 6.11 ± 1.47 | 5.32 ± 2.27 | 3.81 ± 2.45 | 3.84 ± 1.39 | 3.54 ± 1.33 | P *, NaCl *** |
| [cm] | Perla de Cuba | 8.28 ± 1.11 | 7.25 ± 1.23 | 6.23 ± 1.10 | 6.56 ± 1.33 | 5.66 ± 1.11 | 2.76 ± 1.10 | 2.43 ± 1.68 | 3.06 ± 1.31 | P **, NaCl ***, P × BR ** |
| NWDR | INCA LP-5 | 0.67 ± 0.15 | 0.64 ± 0.27 | 0.53 ± 0.18 | 0.39 ± 0.08 | 0.39 ± 0.15 | 0.35 ± 0.13 | 0.33 ± 0.07 | 0.35 ± 0.02 | P *, NaCl *** |
| [cm cm−1] | Perla de Cuba | 0.73 ± 0.14 | 0.56 ± 0.09 | 0.60 ± 0.09 | 0.72 ± 0.19 ** | 0.91 ± 0.18 ** | 0.54 ± 0.20 | 0.52 ± 0.32 | 0.58 ± 0.35 | P × BR * |
| RA | INCA LP-5 | 109 ± 31.0 | 95.1 ± 11.6 | 103 ± 29.5 | 82.7 ± 10.5 | 76.6 ± 19.1 | 51.6 ± 49.9 | 57.3 ± 32.3 | 64.0 ± 7.16 | NaCl *** |
| [°] | Perla de Cuba | 112 ± 13.3 | 107 ± 14.3 | 106 ± 13.7 | 103 ± 17.4 | 103 ± 15.4 * | 50.6 ± 37.2 | 31.3 ± 45.3 | 51.7 ± 32.7 | NaCl ***, P *, P × BR *, P × NaCl × BR * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez-Glaría, A.; Eichler-Löbermann, B.; Loiret, F.G.; Ortega, E.; Kavka, M. Root-System Architectures of Two Cuban Rice Cultivars with Salt Stress at Early Development Stages. Plants 2021, 10, 1194. https://doi.org/10.3390/plants10061194
Vázquez-Glaría A, Eichler-Löbermann B, Loiret FG, Ortega E, Kavka M. Root-System Architectures of Two Cuban Rice Cultivars with Salt Stress at Early Development Stages. Plants. 2021; 10(6):1194. https://doi.org/10.3390/plants10061194
Chicago/Turabian StyleVázquez-Glaría, Alenna, Bettina Eichler-Löbermann, F. G. Loiret, Eduardo Ortega, and Mareike Kavka. 2021. "Root-System Architectures of Two Cuban Rice Cultivars with Salt Stress at Early Development Stages" Plants 10, no. 6: 1194. https://doi.org/10.3390/plants10061194
APA StyleVázquez-Glaría, A., Eichler-Löbermann, B., Loiret, F. G., Ortega, E., & Kavka, M. (2021). Root-System Architectures of Two Cuban Rice Cultivars with Salt Stress at Early Development Stages. Plants, 10(6), 1194. https://doi.org/10.3390/plants10061194