
Non-Uniform Distribution of Cryoprotecting Agents in Rice Culture Cells Measured by CARS Microscopy

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
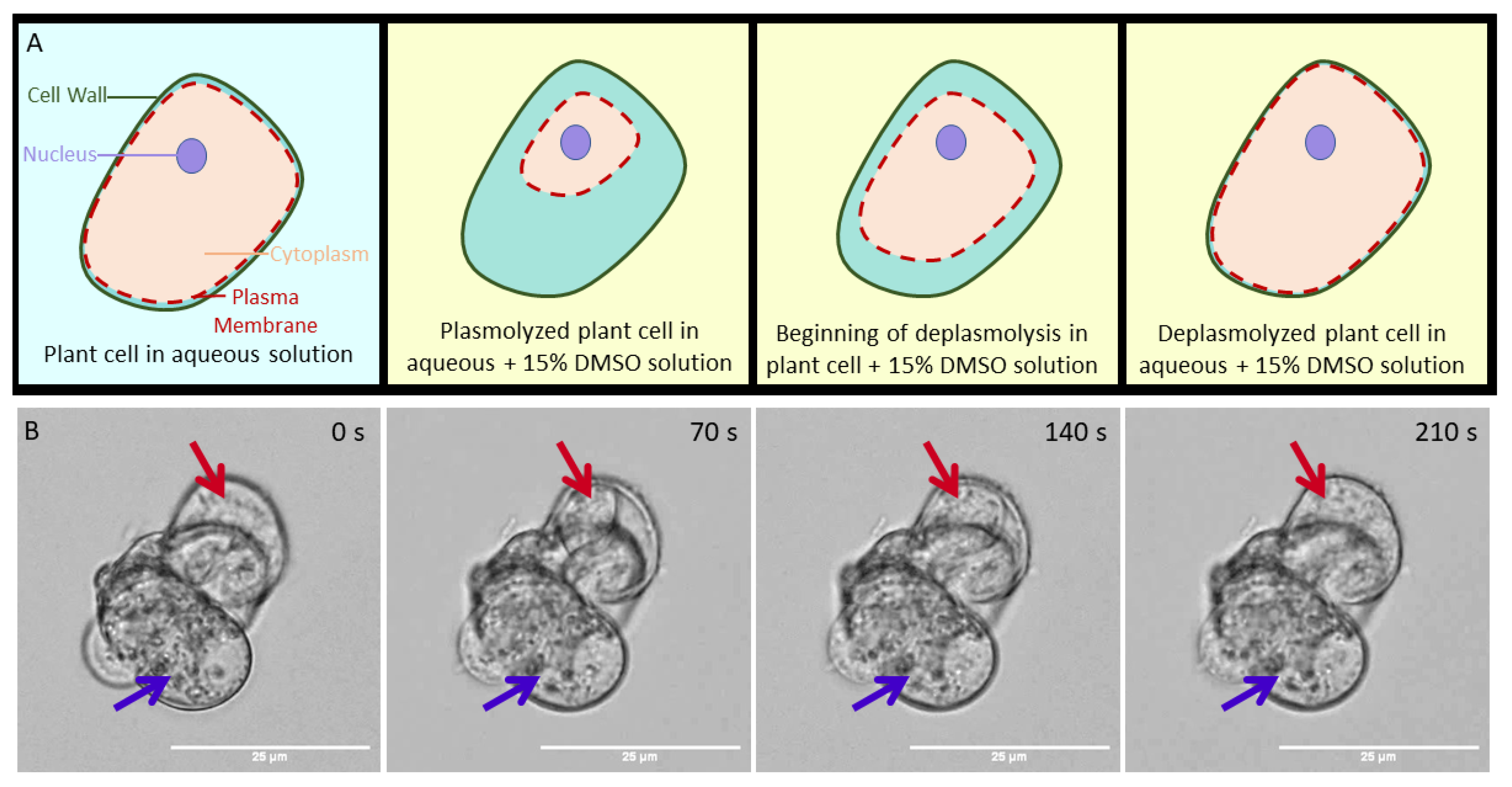
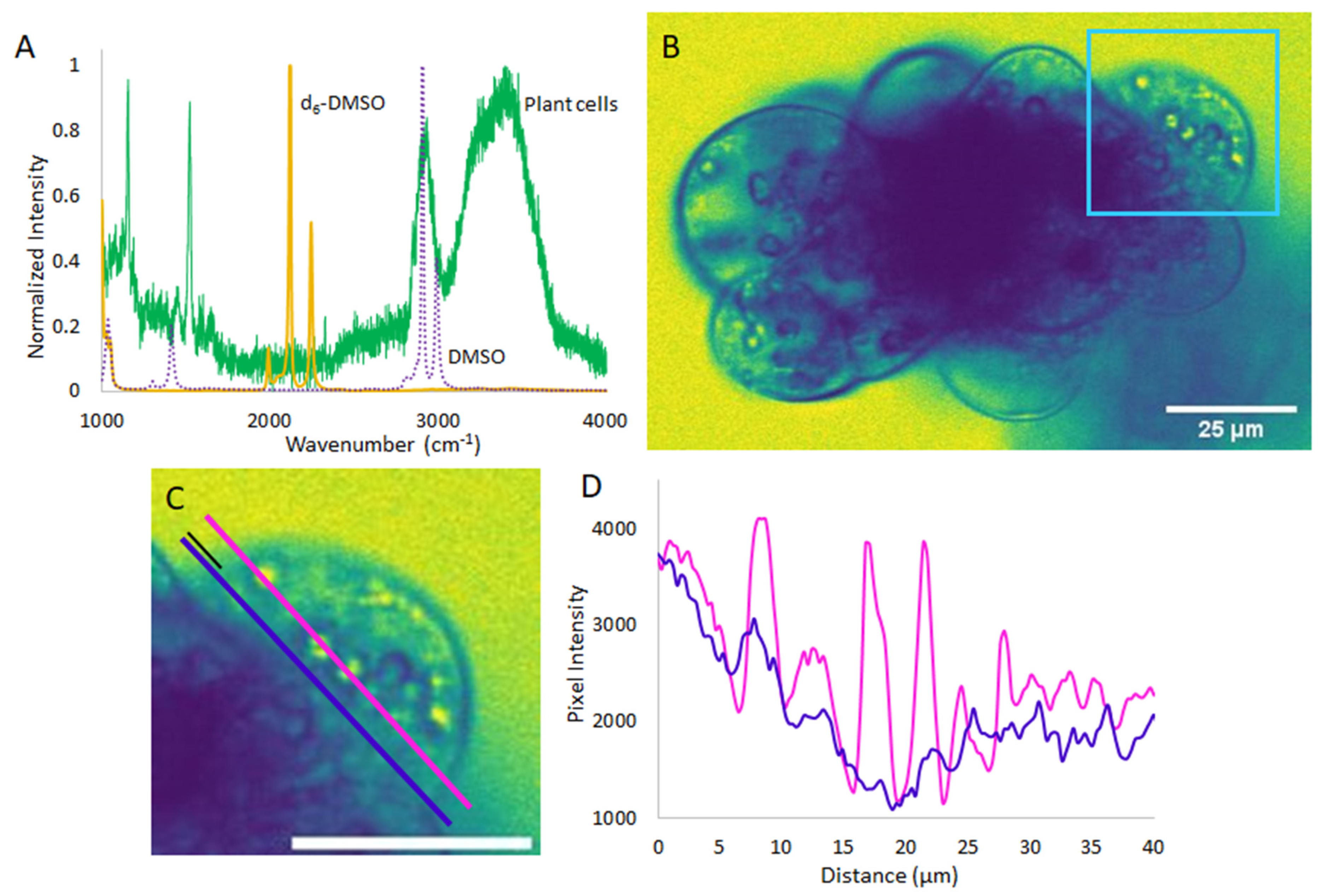
2. Results and Discussion
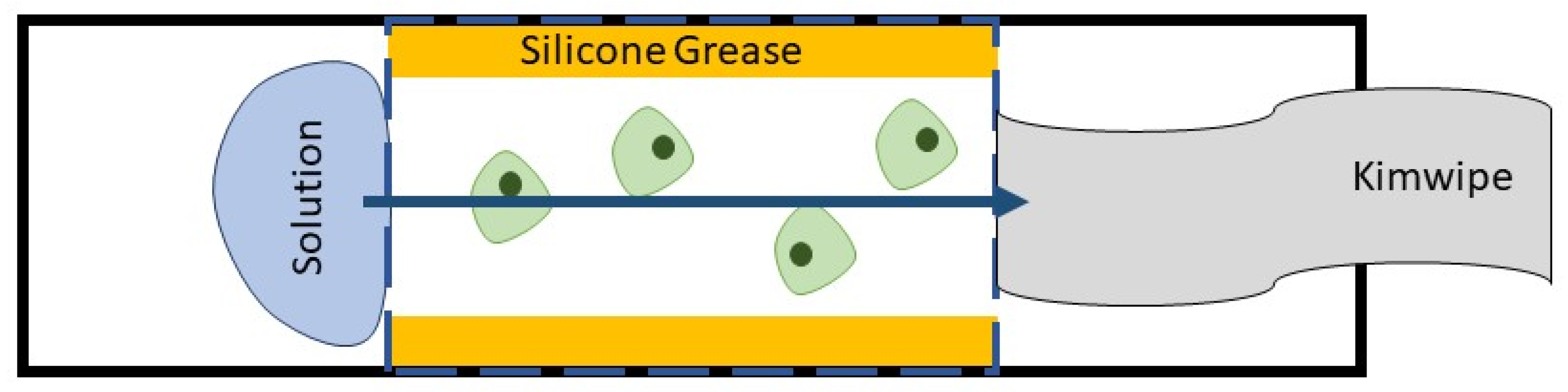
3. Materials and Methods
3.1. Growth and Maintenance of Oryza sativa (Asian Rice) Cells
3.1.1. Rewarming Cells and Initial Plating
3.1.2. Cell Culture
3.2. Cryoprotectant Solutions
3.3. Cell Imaging
3.4. IKI Staining
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coriell, L.L.; Greene, A.E.; Silver, R.K. Historical development of cell and tissue culture freezing. Cryobiology 1964, 1, 72–79. [Google Scholar] [CrossRef]
- Fuller, B.J. Cryoprotectants: The essential antifreezes to protect life in the frozen state. Cryo Lett. 2004, 25, 375–388. [Google Scholar]
- Holt, W.V. Cryobiology, wildlife conservation and reality. CryoLetters 2008, 29, 43–52. [Google Scholar] [PubMed]
- Global Strategy for Plant Conservation. The Targets 2011–2020. Conversion on Biological Diversity. 2011. Available online: https://www.cbd.int/gspc/targets.shtml (accessed on 15 October 2020).
- Sharrock, S. Plant Conservation Report 2020: A Review of Progress in Implementation of the Global Strategy for Plant Conservation 2011–2020; Secretariate of the Convention on Biological Diversity: Montreal, QB, Canada, 2020. [Google Scholar]
- Day, J.G.; Stacey, G. (Eds.) Cryopreservation and Freeze-Drying Protocols, 3rd ed.; Humana Press: New York, NY, USA, 2015. [Google Scholar]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Rep. 1990, 9, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, S.; Sakai, A.; Amano, Y.; Matsuzawa, T. Cryopreservation of asparagus (Asparagus officinalis L.) embryogenic suspension cells and subsequent plant regeneration by vitrification. Plant Sci. 1993, 91, 67–73. [Google Scholar] [CrossRef]
- Volk, G.M.; Caspersen, A.M. Cryoprotectants and components induce plasmolytic responses in sweet potato (Ipomoea batatas (L.) Lam.) suspension cells. Vitr. Cell. Dev. Biol. Anim. 2017, 53, 363–371. [Google Scholar] [CrossRef]
- Volk, G.M.; Caspersen, A.M. Plasmolysis and recovery of different cell types in cryoprotected shoot tips of Mentha × piperita. Protoplasma 2007, 231, 215–226. [Google Scholar] [CrossRef]
- Salma, M.; Engelmann-Sylvestre, I.; Collin, M.; Escoute, J.; Lartaud, M.; Yi, J.-Y.; Kim, H.-H.; Verdeil, J.-L.; Engelmann, F. Effect of the successive steps of a cryopreservation protocol on the structural integrity of Rubia akane Nakai hairy roots. Protoplasma 2013, 251, 649–659. [Google Scholar] [CrossRef]
- Kratochvílová, I.; Kopečná, O.; Bačíková, A.; Pagáčová, E.; Falková, I.; Follett, S.E.; Elliott, K.; Varga, K.; Golan, M.; Falk, M. Changes in Cryopreserved Cell Nuclei Serve as Indicators of Processes during Freezing and Thawing. Langmuir 2018, 35, 7496–7508. [Google Scholar] [CrossRef]
- Zilli, L.; Schiavone, R.; Zonno, V.; Storelli, C.; Vilella, S. Evaluation of DNA damage in Dicentrarchus labrax sperm following cryopreservation. Cryobiology 2003, 47, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Verheijen, M.; Lienhard, M.; Schrooders, Y.; Clayton, O.; Nudischer, R.; Boerno, S.; Timmermann, B.; Selevsek, N.; Schlapbach, R.; Gmuender, H.; et al. DMSO induces drastic changes in human cellular processes and epigenetic landscape in vitro. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Kaczmarczyk, A.; Rutten, T.; Melzer, M.; Keller, E.R.J. Ultrastructural changes associated with cryopreservation of potato (Solanum tuberosum L.) shoot tips. CryoLetters 2008, 29, 145–156. [Google Scholar] [PubMed]
- Halmagyi, A.; Coste, A.; Tripon, S.; Crăciun, C. Low temperature induced ultrastructural alterations in tomato (Lycopersicon esculentum Mill.) shoot apex cells. Sci. Hortic. 2017, 222, 22–31. [Google Scholar] [CrossRef]
- Shi, J.; Wong, T.T.W.; He, Y.; Li, L.; Zhang, R.; Yung, C.S.; Hwang, J.; Maslov, K.; Wang, L.V. High-resolution, high-contrast mid-infrared imaging of fresh biological samples with ultraviolet-localized photoacoustic microscopy. Nat. Photon. 2019, 13, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.-X.; Xie, X.S. Vibrational spectroscopic imaging of living systems: An emerging platform for biology and medicine. Science 2015, 350, aaa8870. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, P.; Slipchenko, M.N.; Cheng, J.-X. Fast Vibrational Imaging of Single Cells and Tissues by Stimulated Raman Scattering Microscopy. Accounts Chem. Res. 2014, 47, 2282–2290. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.L.; Xie, X.S. Coherent Anti-Stokes Raman Scattering Microscopy: Chemical Imaging for Biology and Medicine. Annu. Rev. Anal. Chem. 2008, 1, 883–909. [Google Scholar] [CrossRef]
- Lee, H.J.; Cheng, J.-X. Imaging chemistry inside living cells by stimulated Raman scattering microscopy. Methods 2017, 128, 119–128. [Google Scholar] [CrossRef]
- Zhang, C.; Cheng, J.-X. Perspective: Coherent Raman scattering microscopy, the future is bright. APL Photon. 2018, 3, 090901. [Google Scholar] [CrossRef]
- Potma, E.O.; De Boeij, W.P.; Van Haastert, P.J.; Wiersma, D.A. Real-time visualization of intracellular hydrodynamics in single living cells. Proc. Natl. Acad. Sci. USA 2001, 98, 1577–1582. [Google Scholar] [CrossRef] [PubMed]
- Mokrousova, V.; Okotrub, K.; Amstislavsky, S.; Surovtsev, N. Raman spectroscopy evidence of lipid separation in domestic cat oocytes during freezing. Cryobiology 2020, 95, 177–182. [Google Scholar] [CrossRef]
- Dong, J.; Malsam, J.; Bischof, J.C.; Hubel, A.; Aksan, A. Spatial Distribution of the State of Water in Frozen Mammalian Cells. Biophys. J. 2010, 99, 2453–2459. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Li, R.; Hubel, A. Interfacial Interactions of Sucrose during Cryopreservation Detected by Raman Spectroscopy. Langmuir 2019, 35, 7388–7395. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Yap, Y.R.; Pollock, K.; Hubel, A. Characterizing Intracellular Ice Formation of Lymphoblasts Using Low-Temperature Raman Spectroscopy. Biophys. J. 2017, 112, 2653–2663. [Google Scholar] [CrossRef] [PubMed]
- Karpegina, Y.; Okotrub, K.; Brusentsev, E.; Amstislavsky, S.; Surovtsev, N. Cryoprotectant redistribution along the frozen straw probed by Raman spectroscopy. Cryobiology 2016, 72, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Vasudevan, V.; Mushrif, S.H. Insights into the solvation of glucose in water, dimethyl sulfoxide (DMSO), tetrahydrofuran (THF) and N,N-dimethylformamide (DMF) and its possible implications on the conversion of glucose to platform chemicals. RSC Adv. 2015, 5, 20756–20763. [Google Scholar] [CrossRef]
- Tusch, M.; Krüger, J.; Fels, G. Structural Stability of V-Amylose Helices in Water-DMSO Mixtures Analyzed by Molecular Dynamics. J. Chem. Theory Comput. 2011, 7, 2919–2928. [Google Scholar] [CrossRef]
- Galvao, J.; Davis, B.; Tilley, M.; Normando, E.; Duchen, M.R.; Cordeiro, M.F. Unexpected low-dose toxicity of the universal solvent DMSO. FASEB J. 2014, 28, 1317–1330. [Google Scholar] [CrossRef]
- Cheng, C.-Y.; Song, J.; Pas, J.; Meijer, L.H.; Han, S. DMSO Induces Dehydration near Lipid Membrane Surfaces. Biophys. J. 2015, 109, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Gurtovenko, A.A.; Anwar, J. Modulating the Structure and Properties of Cell Membranes: The Molecular Mechanism of Action of Dimethyl Sulfoxide. J. Phys. Chem. B 2007, 111, 10453–10460. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, G.W.; Sharpe, F.T., Jr. Lysine in seed protein from S-aminoethyl-L-cysteine resistant anther-derived tissue cultures of rice. In Vitro 1981, 17, 345–352. [Google Scholar] [CrossRef]
- Finkle, B.J.; Ulrich, J.M. Cryoprotectant removal temperature as a factor in the survival of frozen rice and sugarcane cells. Cryobiology 1982, 19, 329–335. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samuels, F.M.D.; Stich, D.G.; Bonnart, R.; Volk, G.M.; Levinger, N.E. Non-Uniform Distribution of Cryoprotecting Agents in Rice Culture Cells Measured by CARS Microscopy. Plants 2021, 10, 589. https://doi.org/10.3390/plants10030589
Samuels FMD, Stich DG, Bonnart R, Volk GM, Levinger NE. Non-Uniform Distribution of Cryoprotecting Agents in Rice Culture Cells Measured by CARS Microscopy. Plants. 2021; 10(3):589. https://doi.org/10.3390/plants10030589
Chicago/Turabian StyleSamuels, Fionna M. D., Dominik G. Stich, Remi Bonnart, Gayle M. Volk, and Nancy E. Levinger. 2021. "Non-Uniform Distribution of Cryoprotecting Agents in Rice Culture Cells Measured by CARS Microscopy" Plants 10, no. 3: 589. https://doi.org/10.3390/plants10030589
APA StyleSamuels, F. M. D., Stich, D. G., Bonnart, R., Volk, G. M., & Levinger, N. E. (2021). Non-Uniform Distribution of Cryoprotecting Agents in Rice Culture Cells Measured by CARS Microscopy. Plants, 10(3), 589. https://doi.org/10.3390/plants10030589