
Phenology and Floret Development as Affected by the Interaction between Eps-7D and Ppd-D1

 ,
, 
Abstract
1. Introduction
2. Results
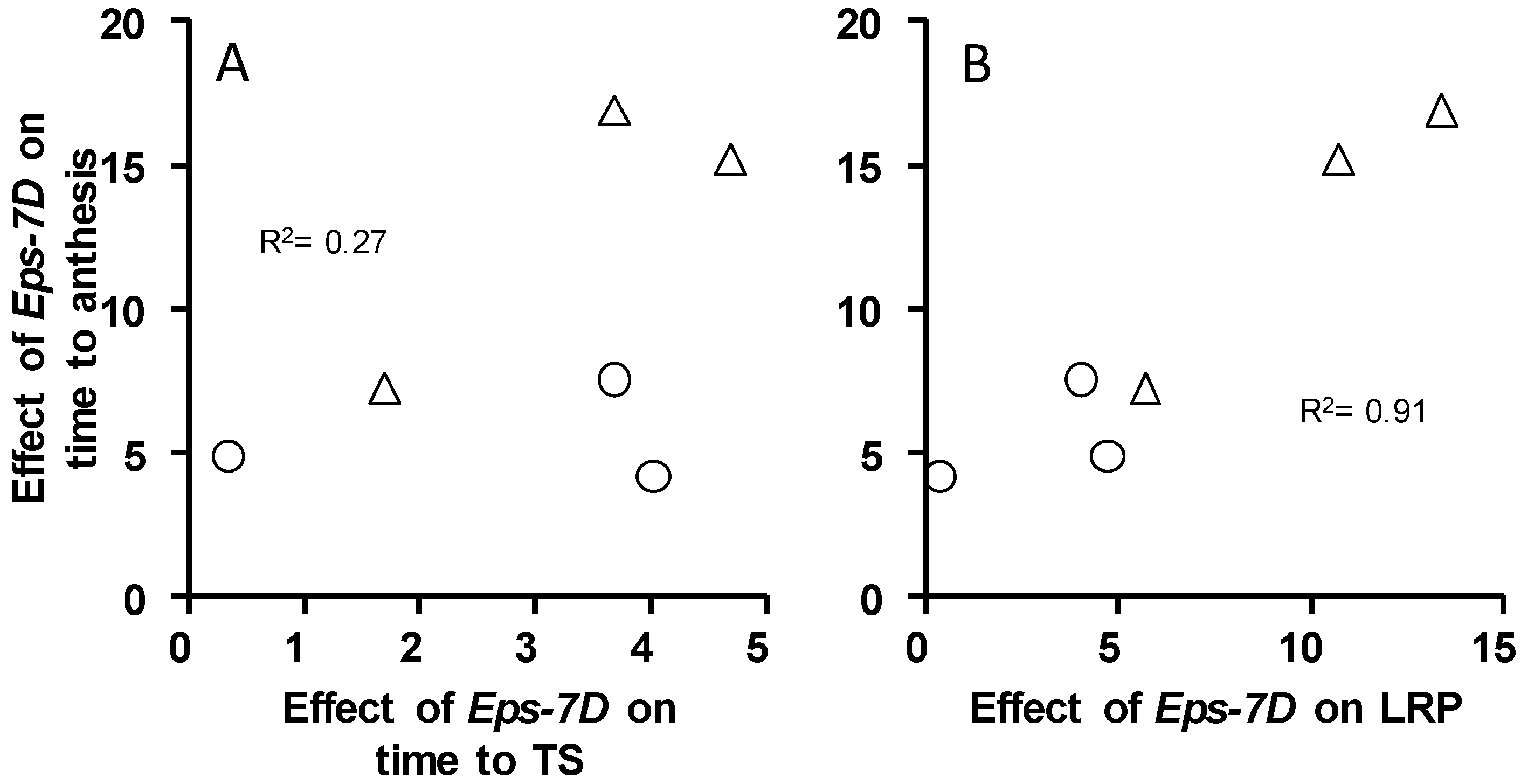
2.1. Phenology
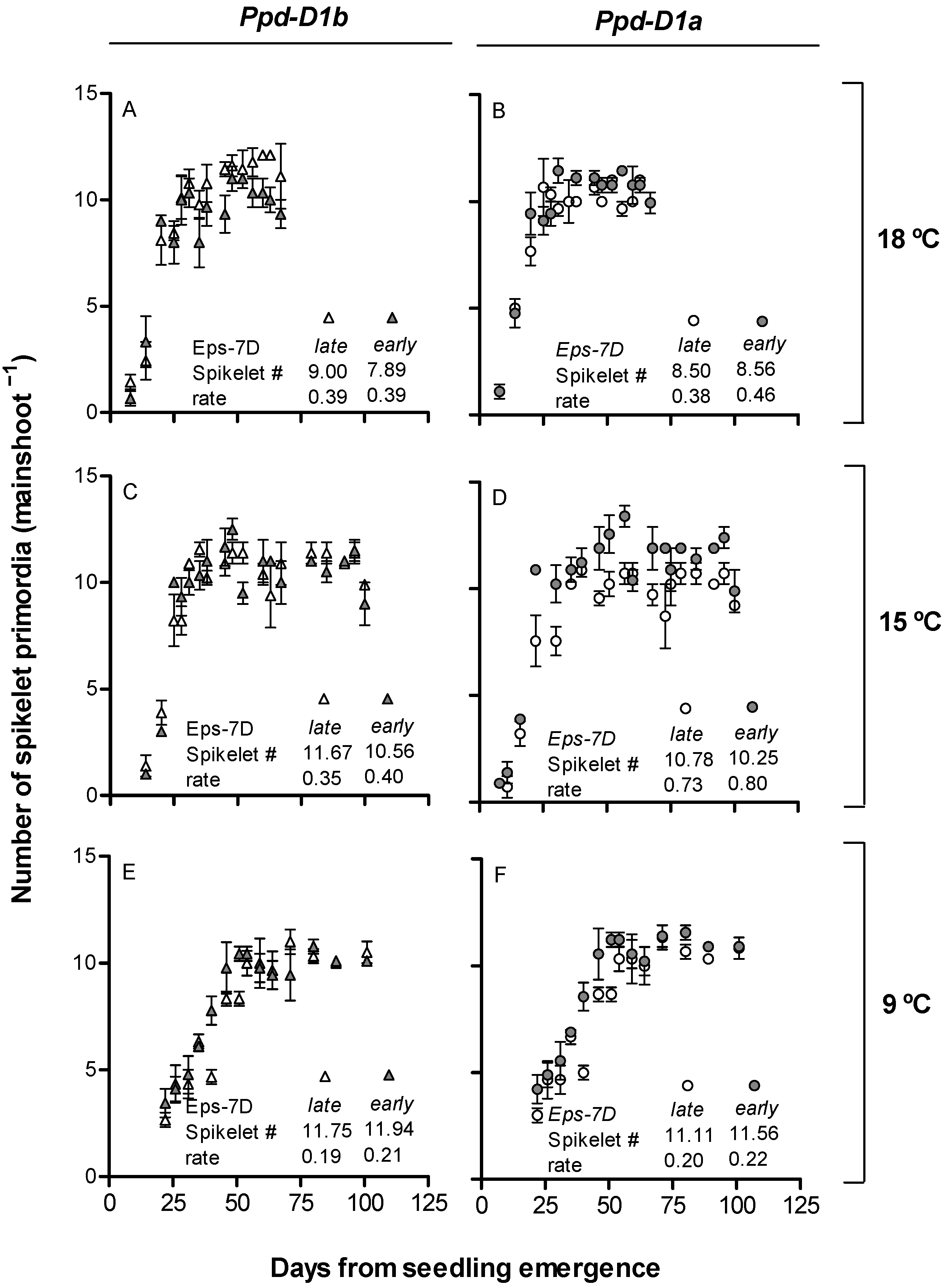
2.2. Dynamics of Leaf Appearance and Spikelet Primordia Development
2.3. Spike Fertility and Dynamics of Floret Development
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Basavaraddi, P.A.; Savin, R.; Bencivenga, S.; Griffiths, S.; Slafer, G.A. Wheat Developmental Traits as Affected by the Interaction between Eps-7D and Temperature under Contrasting Photoperiods with Insensitive Ppd-D1 Background. In press.
- Snape, J.W.; Butterworth, K.; Whitechurch, E.; Worland, A.J. Waiting for Fine Times: Genetics of Flowering Time in Wheat. Euphytica 2001, 119, 185–190. [Google Scholar] [CrossRef]
- Hyles, J.; Bloomfield, M.T.; Hunt, J.R.; Trethowan, R.M.; Trevaskis, B. Phenology and Related Traits for Wheat Adaptation. Heredity 2020, 125, 417–430. [Google Scholar] [CrossRef]
- Griffiths, S.; Simmonds, J.; Leverington, M.; Wang, Y.; Fish, L.; Sayers, L.; Alibert, L.; Orford, S.; Wingen, L.; Herry, L.; et al. Meta-QTL Analysis of the Genetic Control of Ear Emergence in Elite European Winter Wheat Germplasm. Theor. Appl. Genet. 2009, 119, 383–395. [Google Scholar] [CrossRef]
- Börner, A.; Worland, A.J.; Plaschke, J.; Schumann, E.; Law, C.N. Pleiotropic Effects of Genes for Reduced Height (Rht) and Day-Length Insensitivity (Ppd) on Yield and Its Components for Wheat Grown in Middle Europe. Plant Breed. 1993, 111, 204–216. [Google Scholar] [CrossRef]
- Foulkes, M.J.; Sylvester-Bradley, R.; Worland, A.J.; Snape, J.W. Effects of a Photoperiod-Response Gene Ppd-D1 on Yield Potential and Drought Resistance in UK Winter Wheat. Euphytica 2004, 135, 63–73. [Google Scholar] [CrossRef]
- Appendino, M.L.; Slafer, G.A. Earliness per Se and Its Dependence upon Temperature in Diploid Wheat Lines Differing in the Major Gene Eps-Am1 Alleles. J. Agric. Sci. 2003, 141, 149–154. [Google Scholar] [CrossRef]
- Prieto, P.; Ochagavía, H.; Savin, R.; Griffiths, S.; Slafer, G.A. Physiological Determinants of Fertile Floret Survival in Wheat as Affected by Earliness per Se Genes under Field Conditions. Eur. J. Agron. 2018, 99, 206–213. [Google Scholar] [CrossRef]
- Lewis, S.; Faricelli, M.E.; Appendino, M.L.; Valárik, M.; Dubcovsky, J. The Chromosome Region Including the Earliness per Se Locus Eps-A M1 Affects the Duration of Early Developmental Phases and Spikelet Number in Diploid Wheat. J. Exp. Bot. 2008, 59, 3595–3607. [Google Scholar] [CrossRef]
- Pérez-Gianmarco, T.I.; Slafer, G.A.; González, F.G. Wheat Pre-Anthesis Development as Affected by Photoperiod Sensitivity Genes (Ppd-1) under Contrasting Photoperiods. Funct. Plant Biol. 2018, 45, 645–657. [Google Scholar] [CrossRef]
- Gawroński, P.; Ariyadasa, R.; Himmelbach, A.; Poursarebani, N.; Kilian, B.; Stein, N.; Steuernagel, B.; Hensel, G.; Kumlehn, J.; Sehgal, S.K.; et al. A Distorted Circadian Clock Causes Early Flowering and Temperature-Dependent Variation in Spike Development in the Eps-3Am Mutant of Einkorn Wheat. Genetics 2014, 196, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Slafer, G.A.; Rawson, H.M. Responses to Photoperiod Change with Phenophase and Temperature during Wheat Development. Field Crops Res. 1996, 46, 1–13. [Google Scholar] [CrossRef]
- Alvarez, M.A.; Tranquilli, G.; Lewis, S.; Kippes, N.; Dubcovsky, J. Genetic and Physical Mapping of the Earliness per Se Locus Eps-A m 1 in Triticum Monococcum Identifies EARLY FLOWERING 3 (ELF3) as a Candidate Gene. Funct. Integr. Genom. 2016, 16, 365–382. [Google Scholar] [CrossRef]
- Ochagavía, H.; Prieto, P.; Zikhali, M.; Griffiths, S.; Slafer, G.A. Earliness Per Se by Temperature Interaction on Wheat Development. Sci. Rep. 2019, 9, 2584. [Google Scholar] [CrossRef] [PubMed]
- Prieto, P.; Ochagavía, H.; Griffiths, S.; Slafer, G.A. Earliness per Se×temperature Interaction: Consequences on Leaf, Spikelet, and Floret Development in Wheat. J. Exp. Bot. 2020, 71, 1956–1968. [Google Scholar] [CrossRef]
- Basavaraddi, P.A.; Savin, R.; Wingen, L.U.; Bencivenga, S.; Przewieslik-Allen, A.M.; Griffiths, S.; Slafer, G.A. Interactions between Two QTLs for Time to Anthesis on Spike Development and Fertility in Wheat. Sci. Rep. 2021, 11, 2451. [Google Scholar] [CrossRef]
- Kamran, A.; Iqbal, M.; Navabi, A.; Randhawa, H.; Pozniak, C.; Spaner, D. Earliness per Se QTLs and Their Interaction with the Photoperiod Insensitive Allele Ppd-D1a in the Cutler × AC Barrie Spring Wheat Population. Theor. Appl. Genet. 2013, 126, 1965–1976. [Google Scholar] [CrossRef] [PubMed]
- Bentley, A.R.; Horsnell, R.; Werner, C.P.; Turner, A.S.; Rose, G.A.; Bedard, C.; Howell, P.; Wilhelm, E.P.; Mackay, I.J.; Howells, R.M.; et al. Short, Natural, and Extended Photoperiod Response in BC2F 4 Lines of Bread Wheat with Different Photoperiod-1 (Ppd-1) Alleles. J. Exp. Bot. 2013, 64, 1783–1793. [Google Scholar] [CrossRef] [PubMed]
- Kiss, T.; Balla, K.; Veisz, O.; Láng, L.; Bedő, Z.; Griffiths, S.; Isaac, P.; Karsai, I. Allele Frequencies in the VRN-A1, VRN-B1 and VRN-D1 Vernalization Response and PPD-B1 and PPD-D1 Photoperiod Sensitivity Genes, and Their Effects on Heading in a Diverse Set of Wheat Cultivars (Triticum aestivum L.). Mol. Breed. 2014, 34, 297–310. [Google Scholar] [CrossRef]
- Jones, H.E.; Lukac, M.; Brak, B.; Martinez-Eixarch, M.; Alhomedhi, A.; Gooding, M.J.; Wingen, L.U.; Griffiths, S. Photoperiod Sensitivity Affects Flowering Duration in Wheat. J. Agric. Sci. 2017, 155, 32–43. [Google Scholar] [CrossRef]
- Ochagavía, H.; Prieto, P.; Savin, R.; Griffiths, S.; Slafer, G.A. Duration of Developmental Phases, and Dynamics of Leaf Appearance and Tillering, as Affected by Source and Doses of Photoperiod Insensitivity Alleles in Wheat under Field Conditions. Field Crops Res. 2017, 214, 45–55. [Google Scholar] [CrossRef]
- González, F.G.; Slafer, G.A.; Miralles, D.J. Pre-Anthesis Development and Number of Fertile Florets in Wheat as Affected by Photoperiod Sensitivity Genes Ppd-D1 and Ppd-B1. Euphytica 2005, 146, 253–269. [Google Scholar] [CrossRef]
- Parent, B.; Tardieu, F. Temperature Responses of Developmental Processes Have Not Been Affected by Breeding in Different Ecological Areas for 17 Crop Species. New Phytol. 2012, 194, 760–774. [Google Scholar] [CrossRef] [PubMed]
- Ochagavía, H.; Prieto, P.; Savin, R.; Griffiths, S.; Slafer, G.A. Earliness per Se Effects on Developmental Traits in Hexaploid Wheat Grown under Field Conditions. Eur. J. Agron. 2018, 99, 214–223. [Google Scholar] [CrossRef]
- Hay, R.K.M.; Kirby, E.J.M. Convergence and Synchrony-a Review of the Coordination of Development in Wheat. Aust. J. Agric. Res. 1991, 42, 661–700. [Google Scholar] [CrossRef]
- Slafer, G.A.; Savin, R. Developmental Base Temperature in Different Phenological Phases of Wheat (Triticum aestivum). J. Exp. Bot. 1991, 42, 1077–1082. [Google Scholar] [CrossRef]
- Slafer, G.A.; Rawson, H.M. Sensitivity of Wheat Phasic Development to Major Environmental Factors: A Re-Examination of Some Assumptions Made by Physiologists and Modellers. Aust. J. Plant Physiol. 1994, 21, 393–426. [Google Scholar] [CrossRef]
- Mosaad, M.G.; Ortiz-Ferrara, G.; Mahalakshmi, V.; Fischer, R.A. Phyllochron Response to Vernalization and Photoperiod in Spring Wheat. Crop Sci. 1995, 35, 168–171. [Google Scholar] [CrossRef]
- Scarth, R.; Kirby, E.J.M.; Law, C.N. Effects of the Photoperiod Genes Ppd1 and Ppd2 on Growth and Development of the Shoot Apex in Wheat. Ann. Bot. 1985, 55, 351–359. [Google Scholar] [CrossRef]
- John, R.P.; Megan, G. Temperatures and the Growth and Development of Wheat: A Review. Eur. J. Agron. 1999, 10, 23–36. [Google Scholar]
- Matsuyama, H.; Fujita, M.; Masako, S.; Hisayo, K.; Shimazaki, Y.; Matsunaka, H.; Chono, M.; Hatta, K.; Kubo, K.; Takayama, T.; et al. Growth and Yield Properties of Near-Isogenic Wheat Lines Carrying Different Photoperiodic Response Genes. Plant Prod. Sci. 2015, 18, 57–68. [Google Scholar] [CrossRef]
- Langer, S.M.; Longin, C.F.H.; Würschum, T. Flowering Time Control in European Winter Wheat. Front. Plant Sci. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Farré, A.; Sayers, L.; Leverington-Waite, M.; Goram, R.; Orford, S.; Wingen, L.; Mumford, C.; Griffiths, S. Application of a Library of near Isogenic Lines to Understand Context Dependent Expression of QTL for Grain Yield and Adaptive Traits in Bread Wheat. BMC Plant Biol. 2016, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Slafer, G.A.; Rawson, H.M. Intrinsic Earliness and Basic Development Rate Assessed for Their Response to Temperature in Wheat. Euphytica 1995, 83, 175–183. [Google Scholar] [CrossRef]
- Bullrich, L.; Appendino, M.L.; Tranquilli, G.; Lewis, S.; Dubcovsky, J. Mapping of a Thermo-Sensitive Earliness per Se Gene on Triticum Monococcum Chromosome 1Am. Theor. Appl. Genet. 2002, 105, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Slafer, G.A.; Rawson, H.M. Photoperiod × Temperature Interactions in Contrasting Wheat Genotypes: Time to Heading and Final Leaf Number. Field Crops Res. 1995, 44, 73–83. [Google Scholar] [CrossRef]
- Kiss, T.; Dixon, L.E.; Soltész, A.; Bányai, J.; Mayer, M.; Balla, K.; Allard, V.; Galiba, G.; Slafer, G.A.; Griffiths, S.; et al. Effects of Ambient Temperature in Association with Photoperiod on Phenology and on the Expressions of Major Plant Developmental Genes in Wheat (Triticum aestivum L.). Plant Cell Environ. 2017, 40, 1629–1642. [Google Scholar] [CrossRef]
- Abbate, P.E.; Andrade, F.H.; Culot, J.P.; Bindraban, P.S. Grain Yield in Wheat: Effects of Radiation during Spike Growth Period. Field Crops Res. 1997, 54, 245–257. [Google Scholar] [CrossRef]
- Fischer, R.A. Understanding the Physiological Basis of Yield Potential in Wheat. J. Agric. Sci. 2007, 145, 99–113. [Google Scholar] [CrossRef]
- Serrago, R.A.; Miralles, D.J.; Slafer, G.A. Floret Fertility in Wheat as Affected by Photoperiod during Stem Elongation and Removal of Spikelets at Booting. Eur. J. Agron. 2008, 28, 301–308. [Google Scholar] [CrossRef]
- González, F.G.; Slafer, G.A.; Miralles, D.J. Floret Development and Survival in Wheat Plants Exposed to Contrasting Photoperiod and Radiation Environments during Stem Elongation. Funct. Plant Biol. 2005, 32, 189–197. [Google Scholar] [CrossRef] [PubMed]
- González, F.G.; Slafer, G.A.; Miralles, D.J. Vernalization and Photoperiod Responses in Wheat Pre-Flowering Reproductive Phases. Field Crops Res. 2002, 74, 183–195. [Google Scholar] [CrossRef]
- Miralles, D.J.; Richards, R.A.; Slafer, G.A. Duration of the Stem Elongation Period Influences the Number of Fertile Florets in Wheat and Barley. Funct. Plant Biol. 2000, 27, 931. [Google Scholar] [CrossRef]
- Zikhali, M.; Griffiths, S. The Effect of Earliness per Se (Eps) Genes on Flowering Time in Bread Wheat. In Advances in Wheat Genetics: From Genome to Field; Springer: Tokyo, Japan, 2015; pp. 339–345. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Haun, J.R. Visual Quantification of Wheat Development. Agron. J. 1973, 65, 116–119. [Google Scholar] [CrossRef]
- Kirby, E.J.M.; Appleyard, M. Development and Structure of the Wheat Plant. In Wheat Breeding; Springer: Dordrecht, The Netherlands, 1987; pp. 287–311. [Google Scholar] [CrossRef]
- Waddington, S.R.; Cartwright, P.M.; Wall, P.C. A Quantitative Scale of Spike Initial and Pistil Development in Barley and Wheat. Ann. Bot. 1983, 51, 119–130. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growing Conditions | Allele at Eps-7D | FLN | RLA (Leaves d−1) | r2 | |
|---|---|---|---|---|---|
| Ppd-D1b | 18 °C | late | 7.0 ± 0.0 | 0.114 ± 0.001 | 0.986 *** |
| early | 6.9 ± 0.1 | 0.123 ± 0.001 | 0.983 *** | ||
| 15 °C | late | 7.9 ± 0.1 | 0.076 ± 0.001 | 0.981 *** | |
| early | 7.6 ± 0.1 | 0.084 ± 0.001 | 0.975 *** | ||
| 9 °C | late | 7.0 ± 0.0 | 0.061 ± 0.000 | 0.987 *** | |
| early | 6.9 ± 0.1 | 0.066 ± 0.001 | 0.982 *** | ||
| Ppd-D1a | 18 °C | late | 7.0 ± 0.0 | 0.126 ± 0.001 | 0.983 *** |
| early | 6.6 ± 0.2 | 0.130 ± 0.002 | 0.975 *** | ||
| 15 °C | late | 7.0 ± 0.0 | 0.067 ± 0.001 | 0.980 *** | |
| early | 6.8 ± 0.0 | 0.072 ± 0.001 | 0.973 *** | ||
| 9 °C | late | 6.7 ± 0.2 | 0.067 ± 0.001 | 0.962 *** | |
| early | 6.1 ± 0.1 | 0.071 ± 0.001 | 0.976 *** | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basavaraddi, P.A.; Savin, R.; Bencivenga, S.; Griffiths, S.; Slafer, G.A. Phenology and Floret Development as Affected by the Interaction between Eps-7D and Ppd-D1. Plants 2021, 10, 533. https://doi.org/10.3390/plants10030533
Basavaraddi PA, Savin R, Bencivenga S, Griffiths S, Slafer GA. Phenology and Floret Development as Affected by the Interaction between Eps-7D and Ppd-D1. Plants. 2021; 10(3):533. https://doi.org/10.3390/plants10030533
Chicago/Turabian StyleBasavaraddi, Priyanka A., Roxana Savin, Stefano Bencivenga, Simon Griffiths, and Gustavo A. Slafer. 2021. "Phenology and Floret Development as Affected by the Interaction between Eps-7D and Ppd-D1" Plants 10, no. 3: 533. https://doi.org/10.3390/plants10030533
APA StyleBasavaraddi, P. A., Savin, R., Bencivenga, S., Griffiths, S., & Slafer, G. A. (2021). Phenology and Floret Development as Affected by the Interaction between Eps-7D and Ppd-D1. Plants, 10(3), 533. https://doi.org/10.3390/plants10030533