Abstract
Mexico is floristically the fourth most species-rich country in the world, and Asteraceae is the most diverse vascular plant family in this country. The species exhibits a wide range of growth forms, but the tree-like habit, appropriately named daisy trees, is heavily underestimated, even though slightly different tree definitions are handled. Very little is known about their precise species number or conservation status in Mexico, so we update here the list of known Mexican daisy tree species, summarize their very diverse uses, present a general panorama of their present and future distribution, and discuss their conservation status. A bibliographic review and herbarium study were carried out, carefully curated taxonomical ocurrence maps were prepared for each species, and a climatic suitability modelling approach was used to characterise the spatial patterns of Mexican Asteraceae trees. With 149 daisy tree species, the country ranks second at a global level; within the country, their greatest diversity is found in central and western Mexico. A decrease in diversity is estimated in areas that currently host the highest species richness, whereas the hotspot regions are estimated to show an increase in species diversity, so climate change is not a threat to all Mexican daisy tree species.
1. Introduction
With more than 23,000 vascular plant species, Mexico is floristically the fourth most species-rich country in the world, after Brazil, China, and Colombia [1,2]; 11,600 of the Mexican plant species are endemic [1]. Asteraceae is the most diverse family of vascular plants in Mexico, with 417 genera and 3050 native species, of which 1988 are endemic, representing about 65% of the family in Mexico [3]. In the flora of North America, 418 genera and 2413 species of this family are registered [4]; in Brazil, 310 genera (64 endemic, 17 exotic) and 2113 species (42 introduced) are registered [5]; in the flora of China, 248 genera (18 endemic, 49 introduced) and 2336 species (1145 endemic, 109 introduced) are registered [6]; in Colombia, 258 genera and 1302 species are registered [7]; while in Ecuador, 217 genera and 918 species (360 endemic) are registered [8]. Its representatives are found practically everywhere on the planet, except in Antarctica and polar regions with permanent ice [9,10]. In Mexico, they are distributed from sea level in coastal dunes to the alpine grasslands of mountainous regions at more than 4000 m elevation [3].
Asteraceae is characterized by its inflorescences called head or capitulum, that simulate a flower that contains numerous florets with unilocular, bicarpellate, inferior ovary, and syngeneic stamens. Their diversity and distribution are due, amongst others, to effective dispersal mechanisms of its fruits by the pappus, the modified calyx (in some cases, the apex of the cypsela lengthens, forming a hook-like structure, as, e.g., in dandelion, Taraxacum officinale, and several Mutisieae, which functions as an aerodynamic structure similar to a propeller to disperse the fruits with the help of the wind), and short life cycles in most of its members. The latter characteristic allows them to colonize disturbed environments or sites where the original vegetation has been removed, thus being essential elements of secondary vegetation, ruderal and weeds in various crops [10,11].
Members of the Asteraceae exhibit a wide range of growth forms, including short-lived annual or perennial herbs, subshrubs, shrubs, trees, and even climbing, epiphytic and (sub)aquatic plants [10]. In the particular case of trees, there are some studies dealing with Mexican species. Standley [12] was one of the first to document the diversity of woody species in Mexico; in the case of Asteraceae, he recorded mainly shrubs and only 14 tree or tree-like species. Other studies where tree species have been included correspond to the taxonomic reviews of some tribes [13,14], genera [15,16,17,18,19,20,21], or sections of these [22,23]. The most recent publications that include arborescent Asteraceae [24,25,26] consider only 36, 62, and 41 species, respectively. Even though at least some of these discrepancies might be due to different tree definitions applied, we consider that a considerable cause is what we might call “Asteraceae tree blindness”; most people, including botanists, picture representatives of this family as annual herbs or short-lived perennials, contrasting with the surprisingly high number of woody species it contains. These trees with their beautiful and striking inflorescences are appropriately called daisy trees (Figure 1).

Figure 1.
Selection of Mexican arborescent species of Asteraceae. (A) Montanoa hexagona (Heliantheae), (B) Pittocaulon praecox (Senecioneae), (C). Telanthophora grandiflora (Senecioneae), (D) Nahuatlea smithii (Gochnatieae), (E) Critoniopsis uniflora (Vernonieae), (F) Sinclairia glabra (Liabeae). Photo credits: (A–D) Rosario Redonda-Martínez; (E–F) Fernando Araujo-Mondragón.
Very little is known about the conservation status of Mexican Asteraceae species in general, and of tree species of this family in particular. Therefore, within the framework of the Global Tree Assessment, in cooperation with the IUCN/SSC Global Tree Specialist Group and Botanic Gardens Conservation International, all arborescent Asteraceae species that are endemic or near-endemic to Mexico (i.e., those shared with the south of the United States of America north of Mexico, and those shared with Central America south of the country) are being assessed for the IUCN Red List. Therefore, we use here the tree definition agreed on by the IUCN/SSC Global Tree Specialist Group, which has also been applied by [25]: a woody plant, usually with a single stem growing to a height of at least 2 m, or if multi-stemmed, then at least one vertical stem 5 cm in diameter at breast height.
A recent exploratory study, including species distribution and spatial analyses of a comprehensive list of native Mexican trees, carried out by [26], included 41 arborescent Asteraceae species. However, based on our knowledge of Asteraceae on the one hand, and our ongoing red listing work on the other hand, we realized that (1) this number is heavily underestimated, even though we handle a slightly different tree definition, and (2) the data analyzed were obtained from the National Biodiversity Information System database of Mexico [27] which, although it compiles and georeferences information, has insufficient taxonomic curation. Moreover, during the preparation of our Red List assessments, we noticed that information on arborescent Asteraceae, as is also the case for tree species in general, is very scattered and knowledge quite limited. As a consequence, a first step in the conservation of these species is the compilation of relevant information in order to obtain a general overview of their distribution, threats, and conservation status.
Based on our ongoing Red List assessments of endemic and near-endemic Mexican Asteraceae and a meta-analysis of carefully curated distribution data, the objectives of this study are the following: (1) to document the precise number of Asteraceae trees that are distributed in Mexico and update the list of Mexican arborescent Asteraceae; (2) to summarize their very diverse uses; (3) to present a general panorama of their present and future distribution, including characterization of climatic suitability; and (4) to discuss the impact on their conservation in protected areas and biogeographical provinces.
2. Results
2.1. Species List
The list, generated from the bibliographic review and study of herbarium specimens, includes 149 tree species of Asteraceae, distributed in three subfamilies and 12 tribes (Appendix A), with Asteroideae having 129 species, being the most diverse. The latter subfamily consists of the tribes Heliantheae (54 species), Eupatorieae (42), and Senecioneae (20), which contain the highest number of species, whereas the remaining six tribes are each represented by only one to four species. Following in order of importance, there is subfamily Vernonioideae, in which the tribe Vernonieae groups 16 taxa and Liabeae only one. Finally, the subfamily Gochnatioideae represented by the Gochnatieae tribe includes only three species (Appendix A, Table 1). At the tribe level, Eupatorieae, Heliantheae, Senecioneae, and Vernonieae represent about 89%, while the other eight are equivalent to the remaining 11% of the total number of daisy trees.

Table 1.
Number of Mexican Asteraceae tree species grouped by subfamilies and tribes based on the most recent classification by Susanna et al. [28]. The percentage represents the number of species of each tribe with respect to the 149 that represent the family in Mexico.
Some species reported as arborescent both in the literature and on the labels of herbarium specimens were excluded from the list because they have been synonimized, e.g., Roldana cordovensis.
2.2. Uses of Daisy Trees
Of the 149 daisy tree species, just under 50% have a registered use. Of the 65 potentially used species, 37 have medicinal purposes, the leaves or young branches being the most used parts. Regarding the diseases they cure or the healing properties attributed to them, 12 species stand out as anti-inflammatory, 11 are used to treat stomach diseases, and 10 for skin conditions, followed by five used as antiseptics and five as febrifugals; they are also used to treat oral, heart, kidney, rheumatism, and vertigo conditions. Moreover, eight species with various medicinal uses were recorded.
Their usefulness as nectariferous species also stands out, distinguishing two main groups of insects and a group of birds, for which they serve as food for honeybees (Apis mellifera), butterflies, and hummingbirds, with 17, 3, and 1 species, respectively.
Six species are applied as forage, and of these, four are used only for that purpose, mainly when they are found in arid or semi-arid zones. Other documented uses for Asteraceae trees are as a living fence, cut flower, artisanal, ceremonial, fuel, construction, insecticide, ritual, and shade for coffee [29] (Table 2).
Table 2.
Use of Mexican arborescent Asteraceae species.
2.3. Diversity per Vegetation Type
The Mexican daisy trees occur in practically all vegetation types and most grow in several vegetation types, although the majority show an affinity for temperate and humid environments. Hence, the highest number of species are recorded in pine forests (107 spp.), followed by oak forests (104 spp.) and cloud forests (90 spp.). However, dry areas also host an important diversity, as 85 species are found in low deciduous forests and 47 in crassicaule shrubland. It should be noted that the genus Nahuatlea is exclusively distributed in arid and semi-arid areas of Mexico, being an important part of the vegetation structure in the crassicaule and thorny shrublands of the south and central part of the country. Finally, disturbed sites also host a significant number of arborescent Asteraceae, with 86 species, thus demonstrating the importance of this family as dominant elements of secondary vegetation [29].
2.4. Distribution in Mexico
Asteraceae trees are found in almost the entire territory; however, the highest number of species is found in the center and south of the country, mainly in the Trans-Mexican Volcanic Belt (Hidalgo, Jalisco, Mexico City, Michoacán, Morelos, State of Mexico, Puebla, Veracruz), the Sierra Madre del Sur (Chiapas, Guerrero, Oaxaca) and the southern portion of the Sierra Madre Oriental (Puebla, Querétaro, Veracruz). (Figure 2).
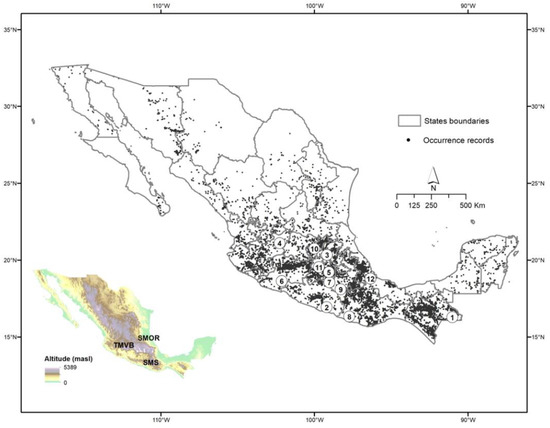
Figure 2.
Occurrence records of Asteraceae tree species in Mexico. The states with the highest number of species are the following: 1 Chiapas, 2. Guerrero, 3. Hidalgo, 4. Jalisco, 5. Mexico City, 6. Michoacán, 7. Morelos, 8. Oaxaca, 9. Puebla, 10. Querétaro, 11. State of Mexico, 12. Veracruz. The map at the left shows the principal mountain regions mentioned in the text: Trans-Mexican Volcanic Belt (TMVB), Sierra Madre del Sur (SMS) and Sierra Madre Oriental (SMOr).
2.5. Climatic Suitability Patterns
The climatic suitability patterns of Asteraceae tree species in Mexico were characterized using models of 86 species, 17 of which show an expansion of over 10% of their current range, whereas 33 species exhibit a contraction of over 50% of their current range; both cases occurs under future scenario (Appendix B). Figure 3 depicts the current and future climatic suitability patterns in Mexico and the difference between scenarios. In the current scenario (1970 to 2000), it is clearly observed that the greatest diversity is found in the west, center, and south of the country, with the states of Jalisco, Michoacán, Mexico, Guerrero, Oaxaca, and Chiapas being those that host the greatest diversity of daisy tree species. The future model (2080 to 2100) estimates a drastic decrease in the number of species in the aforementioned states, although it is more noticeable in Oaxaca. As can be seen on the map that summarizes current and future differences, this state, together with Guerrero, Chiapas, and Jalisco, are those that are estimated to lose the greatest diversity. However, the results also show that the mountain regions of Guerrero and Oaxaca belonging to the Sierra Madre del Sur (SMS) and the Sierra Norte de Oaxaca (SNO), the Tacaná Volcano (TV) in Chiapas on the border with Guatemala, the south of the State of Mexico, northern Michoacán, and the western portion of Jalisco, corresponding to the Trans-Mexican Volcanic Belt (TMVB), will maintain a considerable diversity, indicating that these areas could function as Anthropocene refugia for daisy trees.
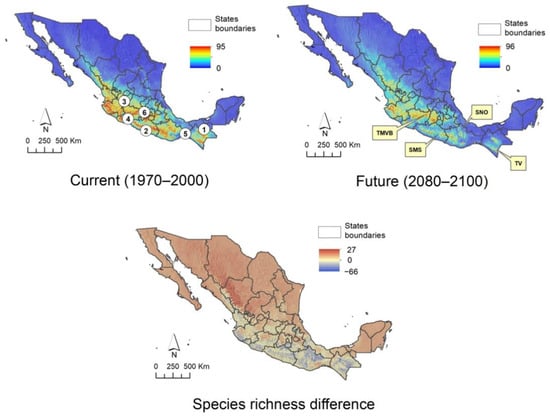
Figure 3.
Effect of climate change on Mexican Asteraceae tree species in two different scenarios: current (1970 to 2000) and future (2080 to 2100) and the difference between both. States with the highest species richness: 1. Chiapas, 2. Guerrero, 3. Jalisco. 4. Michoacán, 5. Oaxaca, 6. State of Mexico. Regions identified as Anthropocene refugia: Trans-Mexican Volcanic Belt (TMVB), Sierra Madre del Sur (SMS), Sierra Norte de Oaxaca (SNO), and Tacaná Volcano (TV).
2.6. Protected Area Network and Biogeographic Provinces
The protected area network of Mexico (Figure 4) and biogeographic provinces (Figure 5) show an uneven distribution of climatic suitability. Observing spatial changes demonstrates that protected areas and provinces have a decrease in low suitability zones and an increase in high suitability zones in the future scenario, respectively. In the case of protected natural areas, the possible decrease that will occur in the future in protected areas such as the Sierra Gorda s.l. (Sierra Gorda and Sierra Gorda de Guanajuato), Los Tuxtlas, and Tehuacán-Cuicatlán Valley is notable. Some exceptions to this trend are the Flora and Fauna Protection Area Cuenca Alimentadora del Distrito Nacional de Riego 043, Estado de Nayarit, as well as the biosphere reserves of the Sierra de Manantlán (2), Monarch Butterfly (4) and El Triunfo (7). The models estimate in these areas that diversity could be maintained or increased in the long term, although this is uncertain. When modeling the climate change scenario on the map of biogeographic provinces, the results are similar. The current scenario shows that the greatest diversity is found along the Pacific coast, and in the Balsas Depression, Sierra Madre Occidental, Sierra Madre del Sur, Oaxaca, Altos de Chiapas, Soconusco, and Trans-Mexican Volcanic Belt. It is estimated that in the future, there will be a decrease in the Sierra Madre Occidental, Sierra Madre Oriental, and Altos de Chiapas. On the other hand, the difference between the two models indicates that there will be a small increase in the Trans-Mexican Volcanic Belt, Northern Altiplano (Chihuahuan desert) and Cape provinces, thereby maintaining the trend observed in the other models: a decrease in the sites that currently host the highest species richness, as well as an increase or no change in areas were the actual daisy tree diversity is considerable, such as the Trans-Mexican Volcanic Belt, Balsas Depression, Sierra Madre del Sur, Pacific coast, and Soconusco.
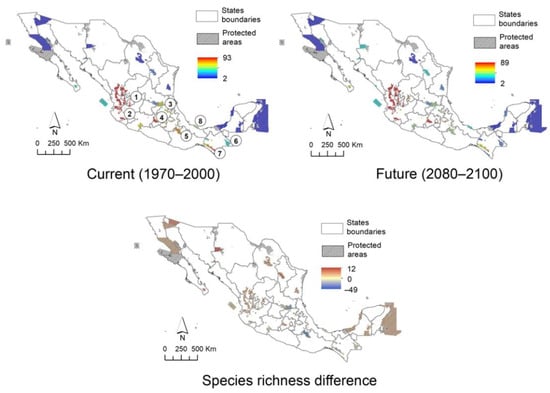
Figure 4.
Actual and future scenarios for Mexican Asteraceae tree richness in the system of Natural Protected Areas at federal level. 1. Flora and Fauna Protection Area “Cuenca Alimentadora del Distrito Nacional de Riego 043, Estado de Nayarit”, 2–8. Biosphere Reserves. 2 Sierra de Manantlán, 3. Sierra Gorda, 4. Monarch Butterfly, 5. Tehuacán-Cuicatlán Valley, 6. Montes Azules, 7. El Triunfo, 8. Los Tuxtlas.
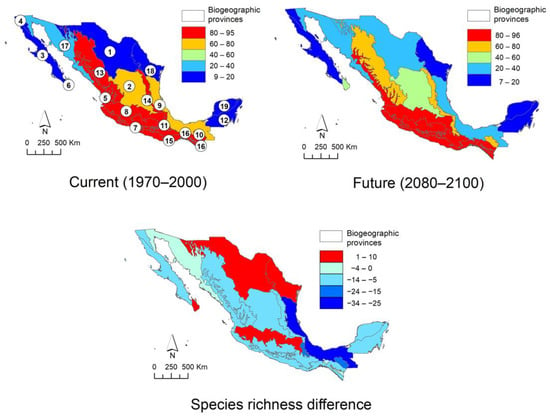
Figure 5.
Estimation of the changes in the distribution of Mexican Asteraceae trees in the different biogeographical provinces of Mexico. 1. Northern Altiplano (Chihuahuense), 2. Southern Altiplano (Zacatecano–Potosina), 3. Baja California, 4. California, 5. Pacific Coast, 6. Cape, 7. Balsas Depression, 8. Trans-Mexican Volcanic Belt, 9. Gulf of Mexico, 10. Altos de Chiapas, 11. Oaxaca, 12. Petén, 13. Sierra Madre Occidental, 14. Sierra Madre Oriental, 15. Sierra Madre del Sur, 16. Soconusco, 17. Sonorense, 18. Tamaulipeca, 19. Yucatán.
2.7. Mexican Daisy Tree Conservation
In the recently updated version of the Mexican decree of endangered species NOM-059-SEMARNAT-2010 [30], only 11 species of Asteraceae are included, of which none correspond to trees, despite the fact that some of them are only known from a few collections, or from the type collection only, and are distributed in areas with strong anthropogenic pressures derived from the change in land use, such as in the Uxpanapa-Chimalapas area in the states of Veracruz and Oaxaca. No Mexican species of Asteraceae are included in the updated appendices of the Convention of International Trade in Endangered Species of Wild Fauna and Flora [31], where only one species of Asteraceae is found—Aucklandia costus Falc. (cited as Saussurea costus (Falc.) Lipsch.)—due to its use in traditional Chinese medicine [32].
With respect to our ongoing assessments of the conservation status of the Mexican Asteraceae trees for the IUCN Red List, less than 10 will be categorized as critically endangered (CR), an estimated 15 to 20 as endangered (EN), about 20 to 25 as vulnerable, and the remainder as species of least concern (LC). The assessments with final conservation statuses will be published on the IUCN Red List later this year.
3. Discussion
3.1. Daisy Tree Diversity in Mexico and Comparison with Other Diverse Areas
The study by Beech et al. [25] reported 3364 tree species for Mexico, positioning this country in the top 10 of the most tree species-rich countries, and due to our efforts in listing additional tree species since then, this has been increased to 3522 species [33]. Similarly, for native Mexican Asteraceae trees, in contrast to the previous studies and reports, we report a much higher number of arborescent Asteraceae taxa. Asteraceae is a main component of vegetation and bioregions along the Americas, with Mexico standing out as the most species-rich country for this family at a global level [1,2].
With respect to the tree species richness of Asteraceae in megadiverse countries and areas, Mexico and Central America rank second in the number of genera and species with 45 and 149, respectively. The first corresponds to Colombia as it is home to 169 species [7], followed by Brazil with 38 genera [34], and Ecuador, with 14 genera and 55 species, all of them in some risk category according to the IUCN Red List criteria [8]. Considering regional scales, the Amazonian (an area that includes the territory of nine South American countries: Bolivia, Brazil, Colombia, Ecuador, French Guyana, Guyana, Peru, Suriname, and Venezuela) is an area of high diversity with 37 genera and 107 species [35]. With respect to the number of Asteraceae trees registered in the flora of North America [4], eight genera and 10 species were found, of which five are mainly shared with the northern part of Mexico; hence, the high diversity of daisy trees in the country stands out. A similar situation occurs with the flora of China [6]; in this case, the numbers are even more contrasting, since in that area only five genera and nine species of native trees are registered. In addition, in China there are also two genera and three cultivated species that are native to Mexico or Central America and that have become naturalized in Chinese territory. These are the only daisy tree species shared between Mexico, Central America, and that region in Asia.
Tree-like Asteraceae are generally not prominently visible in the forests where they occur, as they grow in the understory or in open places, whereas they can reach up to 20 m in cloud forest, but they do not form a prominent part of the forest structure. In contrast, in scrubland vegetation and semi-evergreen low forest, they may be dominant, and an important part of the structure of the forest. It has been documented that Mexican coniferous forests show a relationship between forest structure and tree diversity [36]. In the particular case of Asteraceae, the highest number of arborescent species is found in pine forests (71.8%), while Abies forests concentrate just over 11% of the 149 species present in the country. Even considering that one and the same species can be found in various vegetation types, the percentages for pine and Abies forests are considerable.
3.2. Uses of Mexican Daisy Trees
A considerable amount of ruderal or malezoid Asteraceae are nectariferous and therefore are particularly important for honey-producing bees [37,38,39,40], or other pollinators, which are also attracted in addition to nectar, by the yellow colour of the flowers of many species [41]; hence, they do not depend on a single vector that carries out cross-pollination. The nectar produced by the Asteraceae is rich in glucose, fructose and sucrose [42], which encourages various groups of insects, including Hymenoptera, Diptera, Lepidoptera, and Coleoptera, to obtain food and assist in the pollination of these species [42,43,44]. Even several groups within Mutisieae are hummingbird-pollinated [45,46]. The family is of high economic importance as a honey supplier in several regions in Mexico. The worldwide known migratory phenomenon of the monarch butterfly occurs every year during the fall when millions of butterflies travel from the south of Canada and the north of the United States of America to Mexico to spend the winter season in the Monarch Butterfly Biosphere Reserve, located on the limits of Michoacán and the State of Mexico [47]. The presence of 103 Asteraceae species has been documented in its core zone [48]. From the illustrated flora of the Reserve [49], it can be observed that the butterflies feed on practically all Asteraceae that grow in the area.
Asteraceae are also an important source of food for honey-producing bees in Mexico, both European and native [37,38,40]. Among the species most used by these insects, there is a significant percentage of those that have been classified as “weeds” [40]. Indirectly, these plants are a source of income for beekeepers around the world. Mexico is among the top 10 honey producers worldwide, ranking fourth in exports of this product. In 2019 alone, 61.9 million tons of honey were produced, thus achieving an increase of just over six percent compared to the previous five-year period [50]. Eight states account for 70% of the national production, with Yucatán, Campeche, Jalisco, and Chiapas as the main producers [51].
Asteraceae are used in traditional medicine to treat various conditions such as the treatment of stomach and respiratory diseases, since around 6000 species contain sesquiterpene lactones, chemical compounds with antimicrobial, antiprotozoal, anti-inflammatory and cytotoxic properties [44,52,53]. Others are used as food, whether they are cultivated to obtain leaves or meristems, roots, tubers, heads, seeds to produce oil, natural dyes, bio-insecticides, or as ornamental or florist plants [52]. Although only eight Mexican daisy tree species are used for ornamental purposes, it is important to highlight the potential that other species of the family could have, since, considering their importance as species that produce nectar and pollen, they would be helpful in reducing the loss of bees and other pollinator groups.
The dahlia deserves a special mention as it has been the national flower of Mexico since 1963 [54], since the country has the largest number of wild and endemic species, with 38 and 35, respectively. These are a source of germplasm for the more than 50,000 varieties grown around the world [55]. In addition to the dahlia, there are other wild species with ornamental potential due to their visible inflorescences, among which the following stand out: Montanoa bipinnatifida and Bartlettina sordida. The first is highly appreciated in Mexico [56,57], Spain [58], Australia [59], and New Zealand [60], and the second in Spain [61]. Although there are no published data, some shrub or tree species of Asteraceae are used as living fences, in seasonal crops such as corn or beans, or in vegetables or gardens, among which the following stand out: Barkleyanthus salicifolius, Baccharis heterophylla, Baccharis salicifolia, Montanoa tomentosa, M. leucantha, and M. grandiflora. Asteraceae species associated with corn (milpa) are mainly Tithonia tubiformis, Cosmos bipinnatus, C. sulphureus, Bidens odorata, B. pilosa, Melampodium perfoliatum, Simsia amplexicaulis, and Viguiera dentata, weedy plants that farmers allow to grow alongside the milpa to serve as crop protection, as the loss of the harvest is lessened in the case of a grasshopper or locust plague [29] (pers. obs.).
3.3. Distribution, Including Characterization of Climatic Suitability
The largest quantity of daisy trees is found in the west, center, and south of the country, particularly where the important mountain ranges of the Sierra Madre Occidental, Sierra Madre del Sur, and Trans-Mexican Volcanic Belt converge. Based on the results obtained from the climate suitability model in the future (2080 to 2100), populations will tend to decrease in the sites that are currently particularly rich in tree-like Asteraceae species, e.g., Chiapas, Sierra Norte de Oaxaca, as well as the northern portion of the Sierra Madre Occidental, in the territory occupied by the states of Durango and Sinaloa. There will also be a considerable decrease in the number of species in the Sierra Madre del Sur, particularly in Guerrero, a state that currently ranks fourth in species richness at a national level [3]. If these predictions materialize, several populations of species that are currently found in sites considered Pleistocene refugia [62,63,64] would be lost. This may be due to the fact that their ecosystems would not withstand a scenario of abrupt climate change, such as the one that is estimated to occur in the next 80 years [65]. In this way, the states with the greatest diversity of Asteraceae—Oaxaca, Jalisco, Durango, Guerrero, and Michoacán [3]—would lose a significant number of species and endemism (Figure 2, Figure 3 and Figure 4).
3.4. Conservation
The occurrence points of the known records, as well as the potential distribution models of Asteraceae trees compared with the areas occupied by the main natural protected areas present in the country, show a tendency to reduce their presence in some areas that are currently particularly rich in species; however, in other zones they will remain or increase; some of these correspond to protected natural areas.
Mexico has 182 protected natural areas distributed in maritime and continental territory. Of these, 67 correspond to National Parks, 44 are Biosphere Reserves, 40 Flora and Fauna Protection Areas, 18 Sanctuaries, eight Natural Resources Protection Areas, and five Natural Monuments [66]. Those that are located in continental territory are equivalent to 10.88% of the country’s land surface [66]. One of the terrestrial protected areas with the largest territorial extension is the Flora and Fauna Protection Area Cuenca Alimentadora del Distrito Nacional de Riego 043, Estado de Nayarit, located in the west of the country, comprising part of the territory of the states of Aguascalientes, Jalisco, Durango, Nayarit, and Zacatecas [66]. It is home to around 11 types of vegetation, more than 2000 species of vascular plants and at least two endemic daisy tree species [67,68]. The territorial extension and biological diversity of this protected area is considerable. In the particular case of Asteraceae trees, the climatic suitability models estimate that the diversity of species will remain in the western part of the country, a situation similar to what could occur in the Sierra de Manantlán, one of the most important biosphere reserves in the western region with an area of 139,577.12 ha [66]. The latter was recognized as a biosphere reserve for the biological diversity that it houses in its territory, which includes vegetation of dry, temperate, and humid environments, in addition to being the main water source for more than 430,000 inhabitants of southern Jalisco and northern Colima [69].
The biosphere reserves are distributed throughout the country; 70% have territorial extensions of more than 100 ha or a high species diversity [66]. In the center of the country, the Sierra Gorda stands out with an extension of 383,567.45 ha, located on the limits of Querétaro, Guanajuato, San Luis Potosí and Hidalgo, and Sierra Gorda de Guanajuato (236,882.76 ha) [66], which together occupy the seventh place in size of all the protected natural areas of Mexico [70]. In its territory, there are dry shrublands and temperate forests, which host a great diversity of plants, many of them endemic to the Sierra Madre Oriental [70]. This region has three hydrological sub-basins and a dam declared a Ramsar site, as it is a wetland of global importance [70]. The climate suitability models estimate a slight decrease in the number of tree species of Asteraceae (Figure 4). Among those that would be affected are Baccharis heterophylla, Barkleyanthus salicifolius, Critonia morifolia, Koanophyllon albicaulis, and Nahuatlea hypoleuca, species that fortunately are not restricted to Mexico, since the first four also occur in Central America and the fifth in the south of the United States of America.
One of the most important protected areas in central Mexico is the Monarch Butterfly Biosphere Reserve with an area of 56,259.05 ha [66]. In its territory, there are pine forests, Abies forests, and oak forests that contribute in an important way to the carbon capture from industrial areas and favour the recharge of aquifers that provide water to the metropolitan area of Mexico City, as well as various areas of the states of Michoacán and Mexico [47]. Our models estimate that the daisy tree diversity in this region will be maintained in the next century, if this also happens with the ecosystems where they are found, as well as with the environmental services that they provide to this area of the country.
The Tehuacán-Cuicatlán Valley Biosphere Reserve, with a surface area of 490,186.87 ha, is the largest of all those found in arid and semi-arid zones [66]. In this case, the climatic suitability models estimate a possible decline in species (Figure 3, Figure 4 and Figure 5). Although the number of trees that are currently in its territory is minimal, several species dominate and give structure to the vegetation of some areas, among them: Baccharis heterophylla, Montanoa leucantha, M. tomentosa, Nahuatlea hypoleuca, N. smithii, Parthenium tomentosum, Pittocaulon praecox, P. velatum, Roldana eriophylla, and R. oaxacana. Of these, Nahuatlea smithii and Roldana eriophylla are practically endemic to this region and could be at risk if the scenario predicted by the models takes place.
In the case of tropical ecosystems and particularly rainforests, the main biosphere reserves are: Montes Azules (331,200 ha) and El Triunfo (119,177.29 ha) in Chiapas, and Los Tuxtlas (155,122.47 ha), in Veracruz [66]. Interestingly, the three are in territory belonging to Pleistocene refugia; Montes Azules corresponds to the refugium called Lacandonia and El Triunfo to Soconusco [62]. Both are considered primary Pleistocene refugia; i.e., these zones maintained constant temperature and precipitation conditions during the dry and cold periods that occurred during this period, which allowed them to safeguard common species in humid tropical forests [62]. The climate suitability models estimate different situations in the event of abrupt climatic events, since a probable reduction would occur in Montes Azules, while in El Triunfo, the Asteraceae tree populations would remain. This may be due to the climatic conditions that currently prevail in each of these regions, since in Montes Azules the current oscillation of temperature and precipitation is lower (average annual temperature between 22 and 24 °C; average annual precipitation 2000 to 3000 mm [71]), compared to the values recorded in El Triunfo (mean annual temperature of 18 to 22 °C, with a mean annual rainfall of 1000 to 3000 mm [72]). However, both sites have invaluable biological potential and value. Montes Azules protects 20% of the plant species present in Mexico; recent calculations indicate that one hectare of this reserve protects about 160 tree species and about 700 vascular plants [73]. In El Triunfo, there are important extensions of mountain cloud forest, considered one of the ecosystems that hosts the greatest diversity of trees in North and Central America [74]. The Los Tuxtlas reserve is located in an area recognized as a secondary Pleistocene refugium, because it corresponds to an area that only managed to preserve itself from the drastic drop in temperature or precipitation, during the alternation of cold and dry periods of the Pleistocene [62].
Los Tuxtlas homes coniferous forest, oak forest, mountain cloud forest, high evergreen forest, and mangroves, where around 3000 plant species have been documented, and it is also one of the five areas with the highest amount of tree endemism in Mexico [75]. For this site, the climatic suitability models estimate a decrease in the number of daisy trees. Considering the importance of Los Tuxtlas as an area of endemism for tree species, it would be interesting to explore what happens with other angiosperm groups. In this area, the average annual temperature ranges between 22 and 26 °C and rainfall varies from 1500 to 4500 mm [76]. Although the temperature is relatively constant, the precipitation has a considerable range of variation, which would support the proposal that it was a secondary Pleistocene refugium [62].
Apparently, the aforementioned biosphere reserves will allow that, given a scenario such as that estimated by our models, the probable reduction of the populations does not become catastrophic as in other areas of the country, which are particularly rich in diversity and Asteraceae endemisms, as occurs with the Tehuacán-Cuicatlán Valley.
Anthropocene refugia correspond to territories that meet the following qualities: being ecologically suitable areas to house the diversity units analyzed and having relatively low levels of observed and predicted anthropogenic pressure to allow their long-term persistence in this area, i.e., through several generations [77]. The main difference between Pleistocene and Anthropocene refugia is that the former are sites where organisms resisted and responded to glacial and interglacial oscillations of the late Quaternary, having the possibility of expanding their distribution once environmental stress conditions decreased [78]. In contrast, in order to characterize probable refugia from the Anthropocene, climate change derived from anthropogenic pressures is taken into account [77]. Therefore, the identification of Anthropocene refugia is useful to categorize, plan, and decide where to establish conservation areas for the group of interest [77].
In the case of daisy trees, the models allow the identification of some areas that meet sufficient characteristics to be considered Anthropocene refugia and, therefore, to be maintained or proposed as conservation areas, although some of them are already cataloged like this. This is the case of the western region of Jalisco where the Flora and Fauna Protection Zone Cuenca Alimentadora del Distrito Nacional de Riego 043, Estado de Nayarit and the Biosphere Reserve Sierra de Manantlán are located. Moreover, the Biosphere Reserves of the Monarch Butterfly, on the limits of Michoacán and the State of Mexico, and of the Tacaná Volcano and El Triunfo in Chiapas are also already existing conservation areas; the latter has previously been proposed as a Pleistocene refugium [62]. Based on the results obtained, other areas that could function as Anthropocene refugia correspond to the northern portion of Michoacán and southern part of the State of Mexico, which together with western Jalisco form part of the Trans-Mexican Volcanic Belt, an area that currently contains a high richness of Asteraceae trees, which, according to our models, is estimated to remain or increase (Figure 3, Figure 4 and Figure 5). The mountainous regions of Guerrero and Oaxaca, corresponding to the Sierra Madre del Sur (Figure 3), also seem to meet the characteristics of Anthropocene refugia. Although there are currently no natural protected areas decreed in these areas, despite being sites with high biological diversity and a large number of endemisms, it is a fact that their geographical location and the difficulty of accessing them has kept them safe from human damage. Special mention should be made of the Sierra Norte de Oaxaca, an area particularly rich in diversity and endemism of Asteraceae [79], in which the models indicate that there are also adequate conditions to serve as an Anthropocene refugium, which is confirmed by the fact that it has also been considered as a secondary Pleistocene refugium [62]. However, unlike the mountainous region of central Oaxaca, the Sierra Norte has sufficient infrastructure to access its territory and forest management of the coniferous forests, although the other ecosystems remain almost intact. This shows that, despite the fact that this area is not recognized as a protected area at the federal level, the community forest management that the inhabitants of the region have carried out has been adequate and successful, since in addition to generating jobs and resources for the inhabitants of the region, the forest area has increased in the last four decades [80,81,82,83].
Based on the aforementioned, the sites identified as priority areas to conserve the diversity and endemism of daisy trees are the following: Trans-Mexican Volcanic Belt (including Protected Areas of western of Jalisco), Sierra Madre del Sur, Sierra Norte de Oaxaca, El Triunfo and Tacaná Volcano. All these regions have previously been identified as diversity hotspots of other groups of plants [84,85] and animals [86].
As a consequence, whether the efforts and proposals to conserve nature are federal or local, everything seems to indicate that the establishment, maintenance and conservation of protected natural areas that currently exist in Mexico have been adequate. However, the ideal would be to keep them intact in the long term or, as far as possible, to extend their territory in order to safeguard a greater number of species, both Asteraceae and other families of angiosperms.
In conclusion, relatively few Mexican daisy tree species are currently seriously threatened by climate change or other factors, as most species are widely distributed. Direct exploitation for human use is also not generally a threatening factor. Mexico ranks first at the global level with respect to daisy diversity and second with respect to daisy tree diversity. As mentioned above, Asteraceae are ecologically successful, and the same goes for tree-like representatives of this family. However, it will be important to include those endemic species whose IUCN Red List assessment indicates that they are endangered or critically endangered in the NOM-059-SEMARNAT-2010, as several of these, e.g., Ageratina chimalapana, Lepidonia wendtiana, Mixtecalia teitaensis, Montanoa revealii, and Verbesina sousae, occur in areas subject to anthropogenic pressures that puts their survival at risk, either due to changes in land use, excessive or unplanned tourism, and the extraction of stone material.
4. Materials and Methods
4.1. Study Area
Mexico is located in North America, between the extreme coordinates 32°43′06′′ and 19°32′25′′ latitude N and 114°43′22′′and 84°38′30′′ longitude W. It limits to the north with the United States of America and to the south with Belize and Guatemala. It has a territorial extension of 1,960,189 km2; which positions it in sixth place among the American countries and as 14th in the world. Nearly half of the country is located below the Tropic of Cancer, which favours the presence of temperate and cold climates in the north, as well as temperate and warm climates in the south. In addition, the geographical position of the country, the presence of large mountain ranges such as the Sierra Madre Oriental to the east, the Sierra Madre Occidental to the west and the Trans-Mexican Volcanic Belt that crosses the territory from east to west in the central-southern area of the country, together with geological, edaphic, and microclimatic variations, favour the existence of a great diversity of vegetation types and therefore a high biodiversity [87]. Mexico shares floristic diversity with neighbouring countries; in the northern part it has affinities with some regions of the United States of America, in the southern part with Central America, and to a lesser extent with South America. The species analyzed in this work are those that are endemic or near-endemic to Mexico, i.e., those shared with the south of the United States of America north of Mexico, and those shared with Central America south of the country.
For the purpose of this study, we only focus on the occurrence, distribution pattern uses, and conservation of the daisy tree species within the Mexico territory.
4.2. Compilation of Taxonomic List and Species Information (Use and Habitat)
From a bibliographic review and the consultation of herbarium specimens available online, a list of tree species of Asteraceae was made (Appendix A), considering in this category those that have been described as trees, arborescent or small trees and even some that have been registered as shrubs, but that sometimes also develop a tree habit according to the definition we used. This list was generated based on the revision of regional floras, such as: Flora Novogaliciana [56], Flora of Chiapas [14], Flora Mesoamericana [88], treatments of the Flora del Valle de Tehuacán-Cuicatlán [89,90,91], Flora del Bajío y de Regiones Adyacentes [57]; taxonomic reviews at tribe level (Eupatorieae [13]), genus level [15,16,17,18,19,20,21,92,93], and section level [22,23]. Moreover, online available collections were also consulted: the National Herbarium of Mexico (MEXU) [29], United States National Herbarium (US) [94], herbarium of the Missouri Botanical Garden (MO) [95], and several herbaria of northern Mexico and Arizona whose collections are available via the portal SEinet Arizona-New Mexico Chapter [96].
In addition to the compilation of Mexican species, we looked up the number of daisy tree species in at least five other megadiverse countries: Brazil [5], China [6], Colombia [7], Ecuador [8], and the United States of America [4].
The most recent classification of Asteraceae for subfamilies and tribes is used in Appendix A [28]. The names of some genera and species are based on specialist reviews and criteria, for example, Ageratina [13,97], Critoniopsis [98], and Pachythamnus [99].
4.3. Compilation of Species Ocurrence Geographical Data
The geographical coordinates were obtained from GBIF [100] via GeoCAT [101], carrying out an exhaustive curation of the information, which consisted mainly of eliminating records of human observations without support by vouchers or photos, as well as those that lacked information regarding the collector or herbarium where the voucher is located, or that only presented decimal coordinates. Moreover, duplicate records in the same locality were also deleted. Records that did not correspond to the known distribution of the species were also eliminated, based on expert knowledge, either due to possible misidentifications or because they correspond to specimens grown outside the natural distribution area of a certain species, or invasive groups in areas other than their natural distribution.
When the records were minimal or there was no information available in GBIF [100], this was complemented with data obtained from the labels of herbarium specimens available online through the digital platforms of the National Herbarium of Mexico (MEXU) [29], the National Herbarium of the United States of America (US) [94] and several herbaria of Northern Mexico and Arizona, USA [96]. The localities that lacked geographical coordinates were georeferenced using Google Earth Pro [102]. To locate some little-known localities, the Historical Archive of geostatistical localities [103], Mapcarta [104], and Pueblos de México [105] were used. The data for each species were generated in Excel tables in comma delimited text format (.csv).
4.4. Spatial Analyses
A species distribution modelling (SDM) approach was used to characterise the spatial patterns of tree Asteraceae species in Mexico. SDM allows identifying the geographic areas with the highest climatic suitability in the current period and projects this suitability in future scenarios. The modelling approach was maximum entropy using Maxent software [106]. To model climate adequacy, the compiled occurrence database is used together with a set of explanatory climate variables. Only species with more than 50 unique occurrences were selected for modelling, to ensure good performance [107]. In this study, the climate variables were obtained from the Worldclim 2 database [108] for the current period (1970 to 2000). The variables were selected from the set of 19 bioclimatic variables available in Worldclim at a spatial resolution of 1 km, which were analysed for their degree of correlation in the Americas’ total extent of occurrence. Correlation values between variables higher than 0.7 were excluded, obtaining a set of six variables with low correlation. The variables were temperature seasonality (BIO4), minimum temperature of coldest month (BIO6), temperature annual range (BIO7), annual precipitation (BIO12), precipitation seasonality (BIO15), and precipitation of coldest quarter (BIO19). Each species’ projection was carried out by maintaining the default Maxent regularisation parameters (auto features) and avoiding extrapolation and clamping options. Records less than 1 km apart per species were excluded to avoid spatial autocorrelation. Occurrences were divided into a training set (70% of the total) and a test set (30%). Maxent probability models were projected over the entire distribution of the species occurrences and then were restricted to the continental area of Mexico by cropping the total raster extent area. This step was done to avoid the loss of potential climatic space combinations where species are present outside Mexico and improving final model accuracy [109]. Models were transformed into a binary format, using a threshold of maximum training sensitivity plus specificity [110]. The binary models per species were summed to obtain the current climate suitability pattern. Future projections were obtained using the global circulation model MIROC6 [111], which has been assessed to represent the average conditions of different climatic factors on a global scale [112]. From this global circulation model, the most extreme scenario SSP585 was selected for the future period 2081 to 2100. The same transformation procedure to binary and summation by species was repeated to obtain the future pattern of climate suitability. Three spatial analyses were performed with the current and future climate suitability models: the calculation of the difference between the future and current patterns, the extraction of the climate suitability in Mexico’s protected areas, and finally the extraction for the biogeographic provinces of Mexico [113]. All spatial analyses were performed with ESRI Arcgis software (version 10.8).
4.5. IUCN Red List Assessments
Red List categories were applied according to the IUCN red list criteria [114] and all relevant information was completed in the IUCN SIS database for pending publication on the publicly available IUCN Red List. Data on species occurrence, uses and habitat are those that were obtained for the abovementioned analyses.
Author Contributions
Conceptualization, R.R.-M., E.M.M.S., M.-S.S., data generation and analyses, R.R.-M., P.P., M.-S.S.; writing—original draft preparation, R.R.-M., M.-S.S.; writing—review and editing, all authors; visualization, R.R.-M., P.P.; funding acquisition, R.R.-M., P.P., M.-S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fondation Franklinia, project number 2019-04, the Instituto de Ecología, A.C., project numbers 20006-11337 and 20006-11617, and FONDECYT grant N°1181677.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used in this study were obtained from databases and publicly available literature sources, all of them were cited in the references.
Acknowledgments
R.R.-M. thanks Fernando Araujo Mondragón for his help in capturing information and georeferencing localities in Vernonieae, as well as for providing photographs of some tree species; Karina Machuca Machuca and Aarón Hernández Miranda (Critoniinae) for searching and compiling the information required to carry out the IUCN Red List evaluations; Francisco Galán and Adrián García for the support provided to capture specimen information from online collections. We are very grateful to Sara Oldfield for the critical revision of the manuscript, as well as to the three anonymous reviewers, whose comments have greatly improved our work.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A
Table A1.
List of Mexican endemic and near-endemic arborescent Asteraceae species.
Table A1.
List of Mexican endemic and near-endemic arborescent Asteraceae species.
| Subfamily | Tribe | Species |
|---|---|---|
| Gochnatioideae | Gochnatieae | Nahuatlea arborescens (Brandegee) V.A. Funk |
| Gochnatieae | Nahuatlea hypoleuca (DC.) V.A. Funk | |
| Gochnatieae | Nahuatlea smithii (B.L. Rob. & Greenm.) V.A. Funk | |
| Vernonioideae | Liabeae | Sinclairia glabra (Hemsl.) Rydb. |
| Vernonieae | Critoniopsis baadii (McVaugh) H. Rob. | |
| Vernonieae | Critoniopsis heydeana (J.M. Coult.) H. Rob. | |
| Vernonieae | Critoniopsis leiocarpa (DC.) H. Rob. | |
| Vernonieae | Critoniopsis macvaughii (S.B. Jones) H. Rob. | |
| Vernonieae | Critoniopsis obtusa (Gleason) H. Rob. | |
| Vernonieae | Critoniopsis salicifolia (DC.) H. Rob. | |
| Vernonieae | Critoniopsis shannonii (J.M. Coult.) H. Rob. | |
| Vernonieae | Critoniopsis tomentosa (Lex.) H. Rob. | |
| Vernonieae | Critoniopsis triflosculosa (Kunth) H. Rob. | |
| Vernonieae | Critoniopsis uniflora (Sch. Bip.) H. Rob. | |
| Vernonieae | Critoniopsis villaregalis (Carvajal) H. Rob. | |
| Vernonieae | Lepidaploa polypleura (S.F. Blake) H. Rob. | |
| Vernonieae | Lepidonia salvinae (Hemsl.) H. Rob. & V.A. Funk | |
| Vernonieae | Lepidonia wendtiana (B.L. Turner) Redonda-Mart. & Villaseñor | |
| Vernonieae | Vernonanthura cordata (Kunth) H. Rob. | |
| Vernonieae | Vernonanthura patens (Kunth) H. Rob. | |
| Asteroideae | Astereae | Baccharis glandulifera G.L. Nesom |
| Astereae | Baccharis heterophylla Kunth | |
| Astereae | Baccharis lancifolia Less. | |
| Astereae | Baccharis salicifolia (Ruiz & Pav.) Pers. subsp. monoica (G.L. Nesom) Joch. Müll. | |
| Bahieae | Peucephyllum schottii A.Gray | |
| Coreopsideae | Dahlia imperialis Roezl ex Ortgies | |
| Coreopsideae | Electranthera mutica (DC.) Mesfin, D.J.Crawford & Pruski | |
| Eupatorieae | Ageratina areolaris (DC.) Gage ex B.L.Turner | |
| Eupatorieae | Ageratina cerifera (McVaugh) R.M. King & H. Rob. | |
| Eupatorieae | Ageratina chiapensis (B.L. Rob.) R.M. King & H. Rob. | |
| Eupatorieae | Ageratina chimalapana B.L. Turner | |
| Eupatorieae | Ageratina cylindrica (McVaugh) R.M. King & H. Rob. | |
| Eupatorieae | Ageratina glabrata (Kunth) R.M. King & H. Rob. | |
| Eupatorieae | Ageratina grandifolia (Regel) R.M. King & H. Rob. | |
| Eupatorieae | Ageratina havanensis (Kunth) R.M. King & H. Rob. | |
| Eupatorieae | Ageratina ligustrina (DC.) R.M. King & H. Rob. | |
| Eupatorieae | Ageratina mairetiana (DC.) R.M.King & H.Rob. | |
| Eupatorieae | Ageratina vernalis (Vatke & Kurtz) R.M. King & H. Rob. | |
| Asteroideae | Eupatorieae | Amolinia heydeana (B.L.Rob.) R.M.King & H.Rob. |
| Eupatorieae | Bartlettina luxii (B.L. Rob.) R.M. King & H. Rob. | |
| Eupatorieae | Bartlettina pansamalensis (B.L. Rob.) R.M. King & H. Rob. | |
| Eupatorieae | Bartlettina pinabetensis (B.L. Rob.) R.M. King & H. Rob. | |
| Eupatorieae | Bartlettina platyphylla (B.L. Rob.) R.M. King & H. Rob. | |
| Eupatorieae | Bartlettina prionophylla (B.L. Rob.) R.M. King & H. Rob. | |
| Eupatorieae | Bartlettina sordida (Less.) R.M. King & H. Rob. | |
| Eupatorieae | Bartlettina tuerckheimii (Klatt) R.M. King & H. Rob. | |
| Eupatorieae | Bartlettina williamsii R.M. King & H. Rob. | |
| Eupatorieae | Chromolaena collina (DC.) R.M. King & H. Rob. | |
| Eupatorieae | Chromolaena glaberrima (DC.) R.M.King & H.Rob. | |
| Eupatorieae | Critonia breedlovei R.M. King & H. Rob. | |
| Eupatorieae | Critonia conzatti (Greenm.) R.M. King & H. Rob. | |
| Eupatorieae | Critonia daleoides DC. | |
| Eupatorieae | Critonia hebebotrya DC. | |
| Eupatorieae | Critonia hospitalis (B.L. Rob.) R.M. King & H. Rob. | |
| Eupatorieae | Critonia iltisii R.M. King & H. Rob. | |
| Eupatorieae | Critonia morifolia (Mill.) R.M. King & H. Rob. | |
| Eupatorieae | Critonia paneroi B.L.Turner | |
| Eupatorieae | Critonia quadrangularis (DC.) R.M. King & H. Rob. | |
| Eupatorieae | Critonia sexangularis (Klatt) R.M. King & H. Rob. | |
| Eupatorieae | Critonia tuxtlae R.M. King & H. Rob. | |
| Eupatorieae | Critoniadelphus microdon (B.L. Rob.) R.M. King & H. Rob. | |
| Eupatorieae | Critoniadelphus nubigenus (Benth.) R.M. King & H. Rob. | |
| Eupatorieae | Koanophyllon albicaule (Sch. Bip. ex Klatt) R.M. King & H. Rob. | |
| Eupatorieae | Koanophyllon galeottii (B.L. Rob.) R.M. King & H. Rob. | |
| Eupatorieae | Koanophyllon palmeri (A. Gray) R.M. King & H. Rob. | |
| Eupatorieae | Koanophyllon pittieri (Klatt) R.M. King & H. Rob. | |
| Eupatorieae | Koanophyllon revealii B.L. Turner | |
| Eupatorieae | Kyrsteniopsis nelsonii (B.L. Rob.) R.M. King & H. Rob. | |
| Eupatorieae | Pachythamnus crassirameus (B.L. Rob.) R.M. King & H.Rob. | |
| Heliantheae | Clibadium arboreum Donn. Sm. | |
| Heliantheae | Clibadium leiocarpum Steetz in Seemann | |
| Heliantheae | Clibadium surinamense L. | |
| Heliantheae | Dendroviguiera puruana (Paray) E.E. Schill. & Panero | |
| Heliantheae | Dendroviguiera quinqueradiata (Cav.) E.E. Schill. & Panero | |
| Heliantheae | Dendroviguiera sphaerocephala (DC.) E.E. Schill. & Panero | |
| Heliantheae | Dendroviguiera splendens (Panero & E.E. Schill.) E.E. Schill. & Panero | |
| Heliantheae | Lagascea palmeri (B.L. Rob.) B.L. Rob. | |
| Heliantheae | Lasianthaea fruticosa (L.) K.M. Becker | |
| Heliantheae | Montanoa andersonii McVaugh | |
| Asteroideae | Heliantheae | Montanoa bipinnatifida (Kunth) K. Koch |
| Heliantheae | Montanoa frutescens (Mairet ex DC.) Hemsl. | |
| Heliantheae | Montanoa grandiflora Alamán ex DC. | |
| Heliantheae | Montanoa hexagona B.L. Rob. & Greenm. | |
| Heliantheae | Montanoa imbricata V.A. Funk | |
| Heliantheae | Montanoa karwinskii DC. | |
| Heliantheae | Montanoa leucantha (Lag.) S.F. Blake | |
| Heliantheae | Montanoa revealii H. Rob. | |
| Heliantheae | Montanoa speciosa DC. | |
| Heliantheae | Montanoa tomentosa Cerv. | |
| Heliantheae | Parthenium fruticosum Less. ex Schltdl. & Cham. | |
| Heliantheae | Parthenium schottii Greenm. ex Millsp. & Chase | |
| Heliantheae | Parthenium tomentosum DC. | |
| Heliantheae | Perymenium grande Hemsl. | |
| Heliantheae | Perymenium hintonii McVaugh | |
| Heliantheae | Podachaenium chiapanum B.L. Turner & Panero | |
| Heliantheae | Podachaenium eminens (Lag.) Sch. Bip. ex Sch. Bip. | |
| Heliantheae | Podachaenium standleyi (Steyerm.) B.L.Turner & Panero | |
| Heliantheae | Rensonia salvadorica S.F. Blake | |
| Heliantheae | Rojasianthe superba Standl. & Steyerm. | |
| Heliantheae | Squamopappus skutchii (S.F. Blake) R.K. Jansen, N.A. Harriman & Urbatsch | |
| Heliantheae | Tetrachyron orizabaensis (Klatt) Wussow & Urbatsch | |
| Heliantheae | Tithonia koelzii McVaugh | |
| Heliantheae | Tithonia longiradiata (Bertol.) S.F. Blake | |
| Heliantheae | Verbesina apleura S.F. Blake | |
| Heliantheae | Verbesina breedlovei B.L. Turner | |
| Heliantheae | Verbesina culminicola McVaugh | |
| Heliantheae | Verbesina fastigiata B.L. Rob. & Greenm. | |
| Heliantheae | Verbesina furfuracea McVaugh | |
| Heliantheae | Verbesina guatemalensis B.L. Rob. & Greenm. | |
| Heliantheae | Verbesina hypargyrea B.L. Rob. & Greenm. | |
| Heliantheae | Verbesina hypoglauca Sch. Bip. ex Klatt | |
| Heliantheae | Verbesina klattii B.L. Rob. & Greenm. | |
| Heliantheae | Verbesina lanata B.L. Rob. & Greenm. | |
| Heliantheae | Verbesina montanoifolia B.L. Rob. & Greenm. | |
| Heliantheae | Verbesina oncophora B.L. Rob. & Seaton | |
| Heliantheae | Verbesina oligantha B.L. Rob. | |
| Heliantheae | Verbesina ovatifolia A. Gray | |
| Heliantheae | Verbesina perymenioides Sch. Bip. ex Klatt | |
| Heliantheae | Verbesina platyptera Sch. Bip. ex Klatt | |
| Heliantheae | Verbesina sousae J.J. Fay | |
| Asteroideae | Heliantheae | Verbesina sphaerocephala A. Gray |
| Heliantheae | Verbesina turbacensis Kunth | |
| Heliantheae | Verbesina villaregalis McVuagh | |
| Heliantheae | Wamalchitamia aurantiaca (Klatt) Strother | |
| Inuleae | Pluchea sericea (Nutt.) Coville | |
| Millerieae | Desmanthodium perfoliatum Benth. | |
| Millerieae | Rumfordia floribunda DC. | |
| Millerieae | Schistocarpha longiligula Rydb. | |
| Neurolaeneae | Neurolaena macrophylla Greenm. | |
| Senecioneae | Barkleyanthus salicifolius (Kunth) H. Rob. & Brettell | |
| Senecioneae | Lepidospartum squamatum (A. Gray) A. Gray | |
| Senecioneae | Mixtecalia teitaensis Redonda-Mart., García-Mend., & D. Sandoval | |
| Senecioneae | Pittocaulon filare (McVaugh) H. Rob. & Brettell | |
| Senecioneae | Pittocaulon praecox (Cav.) H. Rob. & Brettell | |
| Senecioneae | Pittocaulon velatum (Greenm.) H. Rob. & Brettell | |
| Senecioneae | Roldana albonervia (Greenm.) H. Rob. & Brettell | |
| Senecioneae | Roldana angulifolia (DC.) H. Rob. & Brettell | |
| Senecioneae | Roldana barba-johannis (DC.) H. Rob. & Brettell | |
| Senecioneae | Roldana eriophylla (Greenm.) H. Rob. & Brettell | |
| Senecioneae | Roldana gentryi H. Rob. & Brettell | |
| Senecioneae | Roldana greenmanii H. Rob. & Brettell | |
| Senecioneae | Roldana neogibsonii (B.L. Turner) B.L. Turner | |
| Senecioneae | Roldana oaxacana (Hemsl.) H.Rob. & Brettell | |
| Senecioneae | Roldana schaffneri (Sch. Bip. ex Klatt) H. Rob. & Brettell | |
| Senecioneae | Telanthophora cobanensis (J.M. Coult.) H. Rob. & Brettell | |
| Senecioneae | Telanthophora grandifolia (Less.) H.Rob. & Brettell | |
| Senecioneae | Telanthophora jaliscana H. Rob. & Brettell | |
| Senecioneae | Telanthophora standleyi (Greenm.) H. Rob. & Brettell | |
| Senecioneae | Telanthophora uspantanensis (J.M. Coult.) H. Rob. & Brettell |
Appendix B
Table A2.
Species list and number of records used for the climatic suitability models. I. Records in whole distribution area, II. Records in Mexico, III. Current suitable area (km2), IV. Future suitable area (km2), V. Future range expansion in Mexico (km2), VI. Future range stability in Mexico (km2), VII. Future range contraction in Mexico (km2), VIII. Future range expansion in Mexico (%), IX. Future range stability in Mexico (%), X. Future range contraction in Mexico (%).
Table A2.
Species list and number of records used for the climatic suitability models. I. Records in whole distribution area, II. Records in Mexico, III. Current suitable area (km2), IV. Future suitable area (km2), V. Future range expansion in Mexico (km2), VI. Future range stability in Mexico (km2), VII. Future range contraction in Mexico (km2), VIII. Future range expansion in Mexico (%), IX. Future range stability in Mexico (%), X. Future range contraction in Mexico (%).
| ID | Species | I | II | III | IV | V | VI | VII | VIII | IX | X |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Ageratina areolaris (DC.) Gage ex B.L.Turner | 552 | 538 | 439,361 | 258,128 | 37,770 | 167,455 | 184,977 | 9.0 | 39.8 | 43.9 |
| 2 | Ageratina chiapensis (B.L. Rob.) R.M. King & H. Rob. | 97 | 94 | 364,784 | 246,875 | 3687 | 10,875 | 237,846 | 1.0 | 3.0 | 65.2 |
| 3 | Ageratina grandifolia (Regel) R.M. King & H. Rob. | 68 | 68 | 412,400 | 139,148 | 1401 | 110,475 | 221,618 | 0.3 | 26.8 | 53.7 |
| 4 | Ageratina ligustrina (DC.) R.M. King & H. Rob. | 824 | 716 | 625,473 | 241,772 | 2746 | 191,062 | 309,808 | 0.4 | 30.5 | 49.5 |
| 5 | Ageratina mairetiana (DC.) R.M.King & H.Rob. | 577 | 567 | 387,878 | 198,528 | 12,833 | 146,160 | 165,799 | 3.3 | 37.7 | 42.7 |
| 6 | Ageratina vernalis (Vatke & Kurtz) R.M. King & H. Rob. | 106 | 102 | 557,923 | 183,660 | 3409 | 142,534 | 301,187 | 0.6 | 25.5 | 54.0 |
| 7 | Baccharis heterophylla Kunth | 502 | 499 | 650,100 | 444,872 | 45,738 | 307,814 | 211,270 | 7.0 | 47.3 | 32.5 |
| 8 | Baccharis lancifolia Less. | 71 | 58 | 372,699 | 119,928 | 4076 | 92,403 | 206,874 | 1.1 | 24.8 | 55.5 |
| 9 | Baccharis monoica G.L. Nesom | 146 | 55 | 336,151 | 128,384 | 3697 | 101,109 | 172,325 | 1.1 | 30.1 | 51.3 |
| 10 | Barkleyanthus salicifolius (Kunth) H. Rob. & Brettell | 416 | 411 | 871,464 | 563,600 | 41,211 | 402,173 | 283,498 | 4.7 | 46.1 | 32.5 |
| 11 | Bartlettina platyphylla (B.L. Rob.) R.M. King & H. Rob. | 77 | 37 | 279,330 | 118,849 | 3467 | 92,476 | 132,519 | 1.2 | 33.1 | 47.4 |
| 12 | Bartlettina sordida (Less.) R.M. King & H. Rob. | 179 | 176 | 311,664 | 64,849 | 258 | 51,960 | 199,004 | 0.1 | 16.7 | 63.9 |
| 13 | Bartlettina tuerckheimii (Klatt) R.M. King & H. Rob. | 146 | 132 | 420,782 | 100,855 | 95 | 81,591 | 258,139 | 0.0 | 19.4 | 61.3 |
| 14 | Chromolaena collina (DC.) R.M. King & H. Rob. | 742 | 608 | 862,983 | 918,322 | 189,840 | 531,576 | 156,402 | 22.0 | 61.6 | 18.1 |
| 15 | Chromolaena glaberrima (DC.) R.M.King & H.Rob. | 227 | 97 | 386,145 | 174,246 | 5085 | 136,090 | 176,815 | 1.3 | 35.2 | 45.8 |
| 16 | Clibadium arboreum Donn. Sm. | 509 | 443 | 312,076 | 118,778 | 9241 | 87,285 | 164,964 | 3.0 | 28.0 | 52.9 |
| 17 | Clibadium surinamense L. | 359 | 9 | 17,074 | 2561 | 1116 | 978 | 12,947 | 6.5 | 5.7 | 75.8 |
| 18 | Critonia daleoides DC. | 417 | 228 | 534,699 | 215,084 | 11,819 | 160,473 | 270,312 | 2.2 | 30.0 | 50.6 |
| 19 | Critonia hebebotrya DC. | 121 | 95 | 725,959 | 587,158 | 37,854 | 430,086 | 153,176 | 5.2 | 59.2 | 21.1 |
| 20 | Critonia hospitalis (B.L. Rob.) R.M. King & H. Rob. | 78 | 75 | 300,071 | 64,605 | 241 | 52,128 | 190,791 | 0.1 | 17.4 | 63.6 |
| 21 | Critonia morifolia (Mill.) R.M. King & H. Rob. | 802 | 301 | 532,910 | 281,050 | 36,494 | 188,731 | 241,060 | 6.8 | 35.4 | 45.2 |
| 22 | Critonia quadrangularis (DC.) R.M. King & H. Rob. | 147 | 112 | 823,247 | 1,190,328 | 288,440 | 648,289 | 5097 | 35.0 | 78.7 | 0.6 |
| 23 | Critonia sexangularis (Klatt) R.M. King & H. Rob. | 178 | 31 | 77,991 | 11,474 | 4005 | 5362 | 58,177 | 5.1 | 6.9 | 74.6 |
| 24 | Critoniadelphus nubigenus (Benth.) R.M. King & H. Rob. | 57 | 37 | 313,638 | 89,891 | 0 | 73,133 | 182,011 | 0.0 | 23.3 | 58.0 |
| 25 | Critoniopsis leiocarpa (DC.) H. Rob. | 315 | 221 | 311,807 | 103,670 | 1117 | 82,904 | 170,099 | 0.4 | 26.6 | 54.6 |
| 26 | Critoniopsis obtusa (Gleason) H. Rob. | 118 | 118 | 461,220 | 360,251 | 54,851 | 230,966 | 133,515 | 11.9 | 50.1 | 28.9 |
| 27 | Critoniopsis salicifolia (DC.) H. Rob. | 115 | 115 | 587,539 | 563,699 | 46,612 | 404,981 | 68,241 | 7.9 | 68.9 | 11.6 |
| 28 | Critoniopsis tomentosa (Lex.) H. Rob. | 225 | 225 | 426,071 | 293,206 | 27,519 | 206,722 | 136,001 | 6.5 | 48.5 | 31.9 |
| 29 | Critoniopsis uniflora (Sch. Bip.) H. Rob. | 170 | 267 | 686,446 | 717,520 | 121,094 | 443,992 | 103,775 | 17.6 | 64.7 | 15.1 |
| 30 | Dahlia imperialis Roezl ex Ortgies | 143 | 197 | 536,126 | 473,635 | 33,319 | 345,751 | 84,478 | 6.2 | 64.5 | 15.8 |
| 31 | Dendroviguiera quinqueradiata (Cav.) E.E. Schill. & Panero | 114 | 42 | 218,168 | 33,681 | 0 | 27,472 | 150,172 | 0.0 | 12.6 | 68.8 |
| 32 | Dendroviguiera sphaerocephala (DC.) E.E. Schill. & Panero | 110 | 114 | 182,080 | 173,000 | 36,608 | 101,219 | 44,420 | 20.1 | 55.6 | 24.4 |
| 33 | Desmanthodium perfoliatum Benth. | 94 | 110 | 328,888 | 198,100 | 5352 | 154,402 | 111,546 | 1.6 | 46.9 | 33.9 |
| 34 | Electranthera mutica (DC.) Mesfin, D.J.Crawford & Pruski | 730 | 94 | 264,289 | 105,968 | 4255 | 81,480 | 132,739 | 1.6 | 30.8 | 50.2 |
| 35 | Critoniopsis triflosculosa (Kunth) H. Rob. | 364 | 679 | 540,285 | 336,732 | 18,823 | 252,324 | 183,402 | 3.5 | 46.7 | 33.9 |
| 36 | Koanophyllon albicaule (Sch. Bip. ex Klatt) R.M. King & H. Rob. | 501 | 407 | 694,526 | 919,970 | 189,819 | 548,328 | 9384 | 27.3 | 78.9 | 1.4 |
| 37 | Koanophyllon galeottii (B.L. Rob.) R.M. King & H. Rob. | 78 | 67 | 481,665 | 331,545 | 7205 | 262,540 | 128,231 | 1.5 | 54.5 | 26.6 |
| 38 | Koanophyllon palmeri (A. Gray) R.M. King & H. Rob. | 93 | 93 | 1,131,471 | 1,447,606 | 263,411 | 867,251 | 17,763 | 23.3 | 76.6 | 1.6 |
| 39 | Koanophyllon pittieri (Klatt) R.M. King & H. Rob. | 406 | 127 | 333,021 | 152,754 | 10,319 | 113,018 | 157,329 | 3.1 | 33.9 | 47.2 |
| 40 | Lasianthaea fruticosa (L.) K.M. Becker | 878 | 467 | 972,478 | 984,137 | 227,609 | 550,550 | 228,008 | 23.4 | 56.6 | 23.4 |
| 41 | Lepidaploa polypleura (S.F. Blake) H. Rob. | 122 | 121 | 247,682 | 36,979 | 0 | 30,114 | 171,540 | 0.0 | 12.2 | 69.3 |
| 42 | Lepidospartum squamatum (A. Gray) A. Gray | 334 | 27 | 132,657 | 74,277 | 0 | 54,872 | 43,738 | 0.0 | 41.4 | 33.0 |
| 43 | Montanoa frutescens (Mairet ex DC.) Hemsl. | 254 | 254 | 565,398 | 375,408 | 39,691 | 259,895 | 195,086 | 7.0 | 46.0 | 34.5 |
| 44 | Montanoa hexagona B.L. Rob. & Greenm. | 50 | 45 | 294,889 | 68,068 | 0 | 55,131 | 184,037 | 0.0 | 18.7 | 62.4 |
| 45 | Montanoa karwinskii DC. | 67 | 67 | 579,350 | 565,442 | 99,152 | 349,899 | 115,859 | 17.1 | 60.4 | 20.0 |
| 46 | Montanoa leucantha (Lag.) S.F. Blake | 826 | 826 | 1,004,151 | 833,182 | 63,699 | 588,544 | 202,978 | 6.3 | 58.6 | 20.2 |
| 47 | Montanoa revealii H. Rob. | 51 | 51 | 90,690 | 55,06 | 0 | 4477 | 69,263 | 0.0 | 4.9 | 76.4 |
| 48 | Montanoa speciosa DC. | 93 | 93 | 1,142,963 | 1,139,462 | 70,460 | 839,795 | 74,223 | 6.2 | 73.5 | 6.5 |
| 49 | Montanoa tomentosa Cerv. | 714 | 611 | 741,882 | 692,138 | 59,744 | 493,406 | 102,033 | 8.1 | 66.5 | 13.8 |
| 50 | Nahuatlea arborescens (Brandegee) V.A. Funk | 64 | 59 | 14,120 | 36,404 | 17,276 | 11,072 | 0 | 122.4 | 78.4 | 0.0 |
| 51 | Nahuatlea hypoleuca (DC.) V.A. Funk | 185 | 179 | 695,788 | 568,631 | 100,275 | 341,675 | 196,511 | 14.4 | 49.1 | 28.2 |
| 52 | Parthenium fruticosum Less. ex Schltdl. & Cham. | 58 | 58 | 268,619 | 625,789 | 309,243 | 174,803 | 36,718 | 115.1 | 65.1 | 13.7 |
| 53 | Parthenium tomentosum DC. | 302 | 302 | 619,554 | 1,277,564 | 529,608 | 468,614 | 19,320 | 85.5 | 75.6 | 3.1 |
| 54 | Perymenium grande Hemsl. | 329 | 176 | 454,542 | 224,368 | 743 | 181,786 | 187,402 | 0.2 | 40.0 | 41.2 |
| 55 | Peucephyllum schottii A. Gray | 237 | 15 | 164,105 | 182,993 | 17,727 | 118,339 | 3981 | 10.8 | 72.1 | 2.4 |
| 56 | Pittocaulon praecox (Cav.) H. Rob. & Brettell | 496 | 496 | 592,825 | 470,700 | 22,228 | 353,993 | 120,575 | 3.7 | 59.7 | 20.3 |
| 57 | Pittocaulon velatum (Greenm.) H. Rob. & Brettell | 163 | 162 | 509,460 | 637,622 | 138,987 | 369,228 | 40,493 | 27.3 | 72.5 | 7.9 |
| 58 | Pluchea sericea (Nutt.) Coville | 138 | 46 | 231,437 | 225,132 | 9077 | 158,609 | 14,129 | 3.9 | 68.5 | 6.1 |
| 59 | Podachaenium eminens (Lag.) Sch. Bip. ex Sch. Bip. | 683 | 564 | 619,757 | 294,788 | 7450 | 229,798 | 270,875 | 1.2 | 37.1 | 43.7 |
| 60 | Rensonia salvadorica S.F. Blake | 96 | 52 | 126,496 | 22,543 | 0 | 18,456 | 84,920 | 0.0 | 14.6 | 67.1 |
| 61 | Roldana albonervia (Greenm.) H. Rob. & Brettell | 441 | 441 | 448,148 | 251,806 | 10,237 | 191,290 | 168,160 | 2.3 | 42.7 | 37.5 |
| 62 | Roldana angulifolia (DC.) H. Rob. & Brettell | 767 | 767 | 587,170 | 258,071 | 1712 | 204,876 | 264,237 | 0.3 | 34.9 | 45.0 |
| 63 | Roldana barba-johannis (DC.) H. Rob. & Brettell | 821 | 817 | 550,253 | 228,428 | 1797 | 181,755 | 260,052 | 0.3 | 33.0 | 47.3 |
| 64 | Roldana eriophylla (Greenm.) H. Rob. & Brettell | 138 | 138 | 500,630 | 608,966 | 94,727 | 396,654 | 7695 | 18.9 | 79.2 | 1.5 |
| 65 | Roldana gentryi H. Rob. & Brettell | 56 | 56 | 690,226 | 475,248 | 27,835 | 344,349 | 200,325 | 4.0 | 49.9 | 29.0 |
| 66 | Roldana schaffneri (Sch. Bip. ex Klatt) H. Rob. & Brettell | 264 | 209 | 436,186 | 118,254 | 1102 | 94,159 | 257,213 | 0.3 | 21.6 | 59.0 |
| 67 | Rumfordia floribunda DC. | 434 | 434 | 376,503 | 147,338 | 5695 | 112,434 | 190,713 | 1.5 | 29.9 | 50.7 |
| 68 | Schistocarpha longiligula Rydb. | 101 | 74 | 81,642 | 7568 | 494 | 5677 | 60,918 | 0.6 | 7.0 | 74.6 |
| 69 | Sinclairia glabra (Hemsl.) Rydb. | 495 | 390 | 522,077 | 402,878 | 53,658 | 267,867 | 153,473 | 10.3 | 51.3 | 29.4 |
| 70 | Telanthophora cobanensis (J.M. Coult.) H. Rob. & Brettell | 179 | 156 | 224,845 | 41,074 | 0 | 33,393 | 149,708 | 0.0 | 14.9 | 66.6 |
| 71 | Telanthophora grandifolia (Less.) H.Rob. & Brettell | 801 | 602 | 521,994 | 217,796 | 4899 | 169,326 | 249,901 | 0.9 | 32.4 | 47.9 |
| 72 | Telanthophora uspantanensis (J.M. Coult.) H. Rob. & Brettell | 160 | 157 | 375,618 | 68,252 | 0 | 55,137 | 248,274 | 0.0 | 14.7 | 66.1 |
| 73 | Tithonia longiradiata (Bertol.) S.F. Blake | 341 | 256 | 187,591 | 35,500 | 78 | 28,755 | 123,464 | 0.0 | 15.3 | 65.8 |
| 74 | Verbesina fastigiata B.L. Rob. & Greenm. | 343 | 343 | 526,821 | 363,759 | 47,946 | 242,226 | 182,200 | 9.1 | 46.0 | 34.6 |
| 75 | Verbesina guatemalensis B.L. Rob. & Greenm. | 88 | 9 | 144,376 | 43,261 | 0 | 35,374 | 82,631 | 0.0 | 24.5 | 57.2 |
| 76 | Verbesina hypargyrea B.L. Rob. & Greenm. | 58 | 50 | 298,605 | 131,918 | 22,497 | 84,905 | 157,747 | 7.5 | 28.4 | 52.8 |
| 77 | Verbesina hypoglauca Sch. Bip. ex Klatt | 149 | 141 | 488,252 | 171,872 | 12,271 | 125,163 | 265,497 | 2.5 | 25.6 | 54.4 |
| 78 | Verbesina klattii B.L. Rob. & Greenm. | 155 | 155 | 266,302 | 108,800 | 9019 | 77,940 | 136,101 | 3.4 | 29.3 | 51.1 |
| 79 | Verbesina lanata B.L. Rob. & Greenm. | 63 | 32 | 155,973 | 24,034 | 4323 | 15,346 | 111,813 | 2.8 | 9.8 | 71.7 |
| 80 | Verbesina montanoifolia B.L. Rob. & Greenm. | 80 | 80 | 504,621 | 339,200 | 24,377 | 244,898 | 157,276 | 4.8 | 48.5 | 31.2 |
| 81 | Verbesina oligantha B.L. Rob. | 91 | 90 | 437,165 | 277,890 | 5494 | 218,217 | 135,095 | 1.3 | 49.9 | 30.9 |
| 82 | Verbesina oncophora B.L. Rob. & Seaton | 292 | 292 | 431,477 | 196,816 | 6150 | 151,754 | 195,615 | 1.4 | 35.2 | 45.3 |
| 83 | Verbesina perymenioides Sch. Bip. ex Klatt | 215 | 203 | 463,276 | 279,876 | 2385 | 224,930 | 151,418 | 0.5 | 48.6 | 32.7 |
| 84 | Verbesina turbacensis Kunth | 658 | 396 | 482,089 | 157,618 | 6000 | 121,700 | 269,119 | 1.2 | 25.2 | 55.8 |
| 85 | Vernonanthura cordata (Kunth) H. Rob. | 221 | 220 | 472,583 | 382,946 | 25,378 | 281,877 | 99,033 | 5.4 | 59.6 | 21.0 |
| 86 | Vernonanthura patens (Kunth) H. Rob. | 748 | 206 | 360,749 | 56,005 | 786 | 44,745 | 248,219 | 0.2 | 12.4 | 68.8 |
References
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex. Biodivers. 2016, 87, 559–902. [Google Scholar] [CrossRef]
- Ulloa Ulloa, C.; Acevedo-Rodríguez, P.; Beck, S.; Belgrano, M.J.; Bernal, R.; Berry, P.E.; Brako, L.; Celis, M.; Davidse, G.; Forzza, R.C.; et al. An integrated assessment of the vascular plant species of the Americas. Science 2017, 358, 1614–1617. [Google Scholar] [CrossRef] [PubMed]
- Villaseñor, J.L. Diversidad y distribución de la familia Asteraceae en México. Bot. Sci. 2018, 96, 332–358. [Google Scholar] [CrossRef]
- FNA, Flora of North America. Asteraceae. Available online: http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=10074#:~:text=With%20418%20genera%20and%202413,alpine%20habitats%20to%20salt%20marshes (accessed on 1 February 2021).
- Roque, N.; Magalhães Teles, A.; Naoki Nakajima, J. (Eds.) A Família Asteraceae no Brasil Clasifição e Diversidade; Editora da Universidade Federal da Bahia: Salvador, Brazil, 2017; p. 260. [Google Scholar] [CrossRef]
- Flora of China. Asteraceae. Available online: http://www.efloras.org/florataxon.aspx?flora_id=2&taxon_id=10074 (accessed on 28 January 2021).
- Avila, F.; Funk, V.A.; Diazgranados, M.; Díaz-Piedrahíta, S.; Vargas, O. Asteraceae. In Catálogo de Plantas y Líquenes de Colombia; Bernal, R., Gradstein, S.R., Celis, M., Eds.; Instituto de Ciencias Naturales, Universidad Nacional de Colombia: Bogotá, Colombia; Available online: http://catalogoplantasdecolombia.unal.edu.co/es/resultados/familia/Asteraceae/ (accessed on 2 February 2021).
- Barriga, P.; Toasa, G.; Montúfar, R.; Tye, A. Asteraceae. In Libro Rojo de Plantas Endémicas del Ecuador; León-Yáñez, S., Valencia, R., Pitmam, N., Endara, L., Ulloa Ulloa, C., Navarrete, H., Eds.; Publicaciones del Herbario QCA, Pontificia Universidad Católica del Ecuador: Quito, Ecuador; Available online: https://bioweb.bio/floraweb/librorojo/ListaEspeciesPorFamilia/500047 (accessed on 29 January 2021).
- Funk, V.A.; Randall, J.B.; Keeley, S.C.; Chan, R.; Watson, L.; Gemeinholzer, B.; Schilling, E.; Panero, J.L.; Baldwin, B.G.; García-Jacas, N.; et al. Everywhere but Antarctica: Using a supertree to understand the diversity and distribution of the Compositae. Biol. Skrif. 2005, 55, 343–373. [Google Scholar]
- Funk, V.A.; Susanna, A.; Stuessy, T.F.; Robinson, H. Classification of Compositae. In Systematics, Evolution and Biogeography of the Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; IAPT: Vienna, Austria, 2009; pp. 171–189. [Google Scholar]
- Crawford, D.J.; Lowrey, T.K.; Anderson, G.J.; Bernardello, G.; Santos-Guerra, A.; Stuessy, T.F. Genetic diversity in Asteraceae endemic to oceanic islands: Baker’s Law and polyploidy. In Systematics, Evolution and Biogeography of the Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; IAPT: Vienna, Austria, 2009; pp. 139–151. [Google Scholar]
- Standley, P.C. Trees and shrubs of Mexico (Bignoniaceae-Asteraceae). Constr. U. S. Natl. Herb. 1926, 23, 1401–1641. [Google Scholar]
- Turner, B.L. The comps of Mexico. A systematic account of the Family Asteraceae, Vol. 1 Eupatorieae. Phytol. Mem. 1997, 11, 1–272. [Google Scholar]
- Strother, J.L. Compositae-Heliantheae s.l. In Flora of Chiapas; Daniel, T.F., Ed.; California Academy of Sciences: San Francisco, CA, USA, 1999; Volume 5, pp. 1–232. [Google Scholar]
- Cabrera, A.L. Revisión del género Gochnatia (Compositae). Rev. Mus. La Plata Nueva Ser. 1971, 12, 1–160. [Google Scholar]
- Robinson, H.; Brettell, R.D. Studies in the Senecioneae (Asteraceae). I. A new genus Pittocaulon. Phytologia 1973, 26, 451–453. [Google Scholar]
- Robinson, H.; Brettell, R.D. Studies in the Senecioneae (Asteraceae). V. The genera Psacaliopsis, Barkleyanthus, Telantophora and Roldana. Phytologia 1974, 27, 402–439. [Google Scholar]
- Funk, V.A. The Systematics of Montanoa (Asteraceae, Heliantheae). Mem. N. Y. Bot. Gard. 1982, 36, 1–133. [Google Scholar]
- Turner, B.L. A recension of Mexican species of Roldana (Asteraceae: Senecioneae). Phytologia 2005, 87, 204–249. [Google Scholar]
- Funston, A.M. Taxonomic revision of Roldana (Asteraceae: Senecioneae), a genus of Southwestern U.S.A., Mexico, and Central America. Ann. Missouri Bot. Gard. 2008, 95, 282–337. [Google Scholar] [CrossRef]
- Funk, V.A.; Sancho, G.; Roque, N. Nahuatlea: A new genus of Compositae (Gochnatieae) from North America. Phytokeys 2017, 91, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.B. Revision of Vernonia section Eremosis (Compositae) in North America. Brittonia 1973, 25, 86–115. [Google Scholar] [CrossRef]
- Turner, B.L. Revision of Verbesina sect. Pseudomontanoa (Asteraceae). Plant Syst. Evol. 1985, 150, 237–262. [Google Scholar] [CrossRef]
- Ricker, M.; Hernández, H.M.; Sousa, M.; Ochoterena, H. Tree and tree-like species of Mexico: Asteraceae, Leguminosae, and Rubiaceae. Rev. Mex. Biodivers. 2013, 84, 439–470. [Google Scholar] [CrossRef]
- Beech, E.; Rivers, M.; Oldfield, S.; Smith, P. GlobalTreeSearch: The first complete global database of tree species and country distribution. J. Sustain. For. 2017, 36, 454–489. [Google Scholar] [CrossRef]
- Téllez, O.; Mattana, E.; Diazgranados, M.; Kühn, N.; Castillo-Lorenzo, E.; Lira, R.; Montes-Leyva, L.; Flores Ortiz, C.M.; Way, M.; Dávila, P.; et al. Native trees of Mexico: Diversity, distribution, uses and conservation. PeerJ 2020, 8, e9898. [Google Scholar] [CrossRef]
- Sistema Nacional de Información Sobre la Biodiversidad de México (SNIB), Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad (CONABIO). Available online: https://www.snib.mx/ (accessed on 19 January 2021).
- Susanna, A.; Baldwin, B.G.; Bayer, R.J.; Bonifacio, J.M.; García-Jacas, N.; Keeley, S.C.; Mandel, J.R.; Ortiz, S.; Robinson, H.; Stuessy, T.F. The classification of the Compositae: A tribute to Vicki Ann Funk (1947–2019). Taxon 2020, 69, 807–814. [Google Scholar] [CrossRef]
- DGRU, Dirección General de Repositorios Universitartios, Universidad Nacional Autónoma de México. Portal de Datos Abiertos UNAM, Colecciones Universitarias, Herbario Nacional de México (MEXU). Available online: https://datosabiertos.unam.mx/biodiversidad/ (accessed on 27 November 2020).
- Diario Oficial de la Federación, Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección Ambiental-Especies Nativas de México de Flora y Faina Silvestres-Categorías de Riesgo y Especificaciones para su Inclusion, Exclusion o Cambio-Lista de Especies en Riesgo. Available online: https://dof.gob.mx/nota_detalle_popup.php?codigo=5173091 (accessed on 26 January 2021).
- CITES, Convención Sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres. Apéndices I, II y III en vigor a Partir del 26 de Noviembre de 2019. Available online: https://cites.org/sites/default/files/esp/app/2019/S-Appendices-2019-11-26.pdf (accessed on 26 January 2021).
- Hempen, C.H.; Fischer, T. A Materia Medica for Chinese Medicine; Churchill Livingstone: London, UK, 2009; pp. 466–513. [Google Scholar] [CrossRef]
- BGCI. GlobalTreeSearch Online Database; Botanic Gardens Conservation International: Richmond, UK, 2017. [Google Scholar]
- CASTUERA-OLIVEIRA, L.; OLIVEIRA-FILHO, A.T.; EISENLOHR, P.V. Emerging hotspots of tree in Brazil. Acta Bot. Bras. 2020, 34, 117–134. [Google Scholar] [CrossRef]
- Ter Steege, H.; Vaessen, R.W.; Cárdenas-López, D.; Sabtier, D.; Antonelli, A.; Mota de Oliveira, S.; Pitman, N.C.A.; Møller Jørgensen, P. The discovery of the Amazonian tree flora with an updated checklist of all know tree taxa. Sci. Rep. 2016, 6, 29549. [Google Scholar] [CrossRef]
- Návar, J. Modeling tree diversity, stand structure and productivity of northern temperate coniferous forests of Mexico. PeerJ 2019, 7, e7051. [Google Scholar] [CrossRef] [PubMed]
- Alaniz-Gutiérrez, L.; Ail-Catzim, C.E.; Villanueva-Gutiérrez, R.; Delgadillo-Rodríguez, J.; Ortiz-Acosta, M.E.; García-Moya, E.; Medina-Cervantes, T.S. Caracterización palinológica de mieles del Valle de Mexicali, Baja California, México. Polibotánica 2017, 43, 255–283. [Google Scholar] [CrossRef]
- Araujo-Mondragón, F.; Redonda-Martínez, R. Flora melífera de la región centro-este del municipio de Pátzcuaro, Michoacán, México. Acta Bot. Mex. 2019, 126, e1444. [Google Scholar] [CrossRef]
- Andrada, A.C. Flora utilizada por Apis mellifera L. en el sur del Caldenal (Provincia Fitogeográfica del Espinal), Argentina. Rev. Mus. Argent. Cienci. Nat. 2003, 5, 329–336. [Google Scholar] [CrossRef]
- Santana-Michel, F.J.; Cervantes-Aceves, N.; Jiménez-Reyes, N. Flora melífera del estado de Colima, México. Ibugana 1998, 6, 251–277. [Google Scholar]
- Calabria, L.M.; Emerenciano, V.P.; Scotti, M.T.; Mabry, T.J. Secondary chemistry of Compositae. In Systematics, Evolution and Biogeography of the Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; IAPT: Vienna, Austria, 2009; pp. 73–88. [Google Scholar]
- Torres, C.; Galetto, L. Are nectar sugar composition and corolla tube length related to the diversity of insects than visit Asteraceae flowers? Plant Biol. 2002, 4, 360–366. [Google Scholar] [CrossRef]
- Torres, C.; Galetto, L. Importancia de los polinizadores en la reproducción de Asteraceae de Argentina Central. Acta Bot. Venez. 2008, 31, 473–494. [Google Scholar]
- Camina, J.L.; Tourn, E.; Andrada, A.C.; Pellegrini, C.; Ashworth, L. Spatial and temporal distribution of floral rewards within the capitula: The case of Hyalis argentea (Asteraceae). Bol. Soc. Argent. Bot. 2019, 54, 17–27. [Google Scholar] [CrossRef]
- Arroyo, M.T.; Robles, V.; Tamburrino, I.; Martínez-Harms, J.; Garreaud, R.D.; Jara-Arancio, P.; Pliscoff, P.; Copier, A.; Arenas, J.; Keymer, J.; et al. Extreme Drought Affects Visitation and Seed Set in a Plant Species in the Central Chilean Andes Heavily Dependent on Hummingbird Pollination. Plants 2020, 9, 1553. [Google Scholar] [CrossRef]
- Moreira-Muñoz, A.; Scherson, R.A.; Luebert, F.; Román, M.J.; Monge, M.; Diazgranados, M.; Silva, H. Biogeography, phylogenetic relationships and morphological analyses of the South American genus Mutisia L.f. (Asteraceae) shows early connections of two disjunct biodiversity hotspots. Org. Divers. Evol. 2020, 20, 639–656. [Google Scholar] [CrossRef]
- Secretaría del Medio Ambiente y Recursos Naturales. Reserva de la Biosfera Mariposa Monarca. Available online: https://www.gob.mx/semarnat/articulos/reserva-de-la-biosfera-mariposa-monarca-79228 (accessed on 15 January 2021).
- Cornejo-Tenorio, G.; Casas, A.; Farfán, B.; Villaseñor, J.L.; Ibarra-Manríquez, G. Flora y vegetación de las zonas núcleo de la Reserva de la Biosfera Mariposa Monarca, México. Bol. Soc. Bot. Mex. 2003, 73, 43–62. [Google Scholar] [CrossRef][Green Version]
- Cornejo-Tenorio, G.; Ibarra-Manríquez, G. Flora Ilustrada de la Reserva de la Biosfera Mariposa Monarca; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), Universidad Nacional Autónoma de México(UNAM): Mexico City, Mexico, 2008; p. 441. [Google Scholar]
- Secretaría de Agricultura y Desarrollo Rural. La miel Mexicana va Endulzando al Mundo. Available online: https://www.gob.mx/agricultura/articulos/la-miel-mexicana-va-endulzando-el-mundo?idiom=es (accessed on 15 January 2021).
- Secretaría de Agricultura y Desarrollo Rural. Yucatán se Encuentra entre los Principales Productores de miel del país. Available online: https://www.gob.mx/agricultura/yucatan/articulos/yucatan-se-encuentra-entre-los-principales-productores-de-miel-del-pais?idiom=es (accessed on 15 January 2021).
- Simpson, B.B. Economic importance of Compositae. In Systematics, Evolution and Biogeography of the Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; IAPT: Vienna, Austria, 2009; pp. 45–58. [Google Scholar]
- Ruiz-Reyes, E.; Suárez, M. Lactonas sesquiterpénicas. Diversidad estructural y sus actividades biológicas. Rev. CENIC Cienc. Biol. 2015, 46, 9–24. [Google Scholar]
- Diario Oficial de la Federación. Secretaría de Agricultura y Ganadería. Decreto por el que se Declara Símbolo de la Floricultura Nacional la Flor de Dalia en todas sus Especies y Variedades. Available online: http://www.dof.gob.mx/nota_to_imagen_fs.php?codnota=4720563&fecha=13/05/1963&cod_diario=203468 (accessed on 16 January 2021).
- Secretaría de Medio Ambiente y Recursos Naturales. Dalia, flor Representativa de México. Available online: https://www.gob.mx/semarnat/articulos/dalia-flor-representativa-de-mexico?idiom=es (accessed on 16 January 2021).
- McVaugh, R. Compositae. In Flora Novo-Galiciana; Anderson, W.R., Ed.; The University of Muchigan Press: Ann Arbor, MI, USA, 1984; Volume 12, pp. 1–1157. [Google Scholar]
- Rzedowski, J.; Calderón de Rzedowski, G.; Carrillo-Reyes, P. Compositae Tribu Helientheae II (géneros Lagascea-Zinnia). In Flora del Bajío y de Regiones Adyacentes; Rzedowski, J., Calderón de Rzedowski, G., Eds.; Instituto de Ecología, A.C., Centro Regional del Bajío: Pátzcuaro, Michoacán, Mexico, 2011; Volume 172, pp. 1–100. [Google Scholar]
- Flora Ornamental de Barcelona. Available online: https://floraornamentaldebarcelona.com/2020/01/18/flora-del-parc-de-la-guineueta-3-arbustos-2-margaritero-montanoa-bipinnatifida/ (accessed on 8 October 2020).
- Plant This. Available online: http://plantthis.co.nz/plant-information.asp?gardener=18906&tabview=features&plantSpot=1 (accessed on 8 October 2020).
- Auckland Museum. Available online: https://www.aucklandmuseum.com/collections-research/collections/record/am_naturalsciences-object-780123 (accessed on 8 October 2020).
- VivoPlant. Available online: https://vivoplant.com/comprar-plantas-arbustivas-exterior/69-257-especies-arbustivas-bartlettina-sordida.html (accessed on 16 January 2021).
- Toledo, V.M. Los Cambios Climáticos del Pleistoceno y sus Efectos Sobre la Vegetación Tropical Cálida y Húmeda de México. Master’s Thesis, Facultad de Ciencias, Universidad Nacional Autónoma de México, Mexico City, Mexico, 29 April 1976. [Google Scholar]
- Torrescano-Valle, N.; Islebe, G.A. Holocene paleoecology, climate history and human influence in the southwestern Yucatan Peninsula. Rev. Palaeobot. Palynol. 2015, 217, 1–8. [Google Scholar] [CrossRef]
- Escobar García, P.; Winkler, M.; Flatscher, R.; Sonnleitner, M.; Krejcikova, J.; Suda, J.; Hülber, K.; Schneeweiss, G.; Schönswetter, P. Extensive range persistence in peripheral and interior refugia characterizes Pleistocene range dynamics in a widespread Alpine plant species (Senecio carniolicus, Asteraceae). Mol. Ecol. 2012, 21, 1255–1270. [Google Scholar] [CrossRef] [PubMed]
- Cuervo-Robayo, A.P.; Ureta, C.; Gómez-Albores, M.A.; Meneses-Mosquera, A.K.; Téllez-Valdés, O.; Martínez-Meyer, E. One hundred years of climate change in Mexico. PLoS ONE 2020, 15, e0209808. [Google Scholar] [CrossRef] [PubMed]
- Comisión Nacional de Áreas Naturales Protegidas. Áreas Naturales Protegidas Decretadas. Available online: http://sig.conanp.gob.mx/website/pagsig/datos_anp.htm (accessed on 19 February 2021).
- Secretaría de Medio Ambiente y Desarrollo Territorial de Jalisco. 15 Cuenca Alimentadora del Distrito Nacional de Riego 043, Nayarit. Available online: https://semadet.jalisco.gob.mx/medio-ambiente/biodiversidad/areas-naturales-protegidas/142 (accessed on 19 February 2021).
- Sistema de Información y Monitoreo para la Conservación, Comisión Nacional de Áreas Naturales Protegidas. CADNR043 Estado de Nayarit. Available online: https://simec.conanp.gob.mx/ficha_pdf.php?anp=4®=11 (accessed on 19 February 2021).
- Secretaría de Medio Ambiente y Recursos Naturales. Reserva de la Biosfera Sierra de Manantlán. Available online: https://www.gob.mx/semarnat/articulos/reserva-de-la-biosfera-sierra-de-manantlan (accessed on 19 February 2021).
- Secretaría de Medio Ambiente y Recursos Naturales. Reserva de la Biosfera Sierra Gorda. Available online: https://www.gob.mx/semarnat/articulos/reserva-de-la-biosfera-sierra-gorda-celebra-su-19-aniversario (accessed on 19 February 2021).
- Centro de Investigación en Ciencias de Información Geoespacial. Selva Lacandona, Mosaico del Paisaje, Paisaje Ecológico. Available online: http://mapas.centrogeo.org.mx/ciberatlas/lacandona/mosaico/clima.htm (accessed on 20 February 2021).
- Instituto Nacional de Ecología, Plan de manejo de la Reserva de la Biosfera El Triunfo. Available online: http://www.paot.org.mx/centro/ine-semarnat/anp/AN14.PDF (accessed on 20 February 2021).
- Secretaría de Medio Ambiente y Recursos Naturales. Reserva de la Biosfera Montes Azules. Available online: https://www.gob.mx/semarnat/articulos/reserva-de-la-biosfera-montes-azules-selva-lancandona-chiapas?idiom=es (accessed on 20 February 2021).
- Secretaría de Medio Ambiente y Recursos Naturales. Reserva de la Biosfera El Triunfo. Available online: https://www.gob.mx/semarnat/articulos/reserva-de-la-biosfera-el-triunfo?idiom=es (accessed on 20 February 2021).
- Secretaría de Medio Ambiente y Recursos Naturales. Reserva de la Biosfera Los Tuxtlas. Available online: https://www.gob.mx/semarnat/articulos/reserva-de-la-biosfera-los-tuxtlas?idiom=es (accessed on 21 February 2021).
- Comisión Nacional de Áreas Naturales Protegidas. Programa de Conservación y Manejo Reserva de la Biosfera Los Tuxtlas, México. Available online: https://www.conanp.gob.mx/que_hacemos/pdf/programas_manejo/tuxtla_final.pdf (accessed on 21 February 2021).
- Montsarrat, S.; Jarvie, S.; Svenning, J.C. Anthropocene refugia: Integrating history and predictive modelling to assess the space available for biodiversity in a human-dominated world. Philos. Trans. R. Soc. B 2019, 374, 20190219. [Google Scholar] [CrossRef]
- Bennett, K.D.; Provan, J. What do we mean by ‘refugia’? Quat. Scient. Rev. 2008, 27, 2449–2455. [Google Scholar] [CrossRef]
- Suárez-Mota, E.; Villaseñor, J.L.; Ramírez-Aguirre, M.B. Sitios prioritarios para la conservación de la riqueza y el endemismo de la Sierra Norte de Oaxaca, México. Acta Bot. Mex. 2018, 124, 49–74. [Google Scholar] [CrossRef]
- Aschentrupp Toledo, R. Las Comunidades Indígenas de la Sierra Norte de Oaxaca; Centro de Estudios Sociales y Opinión Pública (CESOP): Mexico City, Mexico, 2015; p. 15. [Google Scholar]
- Ceballos Pérez, S.G. Manejo Forestal Comunitario Sustentable en el Sierra Norte de Oaxaca; Published by the Autor: Mexico City, Mexico, 2017; p. 310. [Google Scholar]
- Sastre Merino, S. Análisis de la gestión forestal comunitaria y sus implicaciones sociales en Ixtlán de Juárez, Oaxaca (México). Bachelor’s Thesis, Universidad Politécnica de Madrid, Madrid, Spain, 2008. [Google Scholar]
- Gasca Zamora, J. Gobernanza y gestión comunitaria de recursos naturales en la Sierra Norte de Oaxaca. Reg. Soc. 2014, 60, 89–120. [Google Scholar] [CrossRef]
- Villaseñor, J.L.; Delgadillo, C.; Ortiz, E. Hostspots from a multigroup perspective: Mosses and Senecios in the Transmexican Volcaniv Belt. Biodivers. Conserv. 2006, 15, 4045–4058. [Google Scholar] [CrossRef]
- Suárez-Mota, M.E.; Téllez-Valdés, O. Red de áreas prioritarias para la conservación de la biodiversidad del Eje Volvánico Transmexicano analizando su riqueza florística y variabilidad climática. Polibotánica 2014, 38, 67–93. [Google Scholar]
- Critical Ecosystem Partnership Fund. Región Norte del Hotspot de Biodiversidad de Mesoamérica, Belice, Guatemala, México. Available online: https://www.cepf.net/sites/default/files/final.spanish.mesoamerica.northernmesoamerica.ep_.pdf (accessed on 23 February 2021).
- Instituto Nacional de Estadística y Geografía, Territorio de México. Available online: http://www.cuentame.inegi.org.mx/territorio/extension/default.aspx?tema=T (accessed on 6 February 2021).
- Pruski, J.F. Asteraceae. In Flora Mesoamericana; Davidse, G., Sousa, S.M., Knapp, S., Chiang, F., Eds.; Misssouri Botanical Garden Press: St. Louis, MO, USA, 2018; Volume 5, Part 2, pp. 1–608. [Google Scholar]
- Redonda-Martínez, R.; Villaseñor, J.L. Asteraceae, Vernonieae. In Flora del Valle de Tehuacán-Cuicatlán; Medina Lemos, R., Sánchez Ken, J.G., García Mendoza, A., Arias Montes, S., Eds.; Instituto de Biología, Universidad Nacional Autónoma de México: Mexico City, Mexico, 2009; Volume 72, pp. 1–23. [Google Scholar]
- Redonda-Martínez, R.; Villaseñor, J.L. Asteraceae, Senecioneae. In Flora del Valle de Tehuacán-Cuicatlán; Medina Lemos, R., Sánchez Ken, J.G., García Mendoza, A., Arias Montes, S., Eds.; Instituto de Biología, Universidad Nacional Autónoma de México: Mexico City, Mexico, 2011; Volume 89, pp. 1–64. [Google Scholar]
- Redonda-Martínez, R. Asteraceae, Gochnatieae. In Flora del Valle de Tehuacán-Cuicatlán; Medina Lemos, R., García Mendoza, A., Arias Montes, S., Grether González, R., Fonseca Juárez, R.M., Eds.; Instituto de Biología, Universidad Nacional Autónoma de México: Mexico City, Mexico, 2019; Volume 155, pp. 1–16. [Google Scholar]
- Turner, B.L. Taxonomy of Neurolaena (Asteraceae-Heliantheae). Plant Syst. Evol. 1982, 140, 119–139. [Google Scholar] [CrossRef]
- Redonda-Martínez, R.; Villaseñor, J.L. El género Lepidaploa (Familia Asteraceae, Tribu Vernonieae) en México. Rev. Mex. Biodivers. 2011, 82, 782–797. [Google Scholar] [CrossRef]
- Botany Collection Search, Smithsonian National Museum of Natural History. Available online: http://collections.nmnh.si.edu/search/botany/?v=s1#new-search (accessed on 24 November 2020).
- Tropicos.org. Missouri Botanical Garden. Available online: http://www.tropicos.org (accessed on 28 November 2020).
- SEinet Arizona-New Mexico Chapter. Available online: https://swbiodiversity.org/seinet/collections/harvestparams.php (accessed on 6 December 2020).
- Hinojosa-Espinosa, O.; Villaseñor, J.L.; Ortiz, E. On the identity of two Mexican species of Ageratina (Eupatorieae, Asteraceae): A. grandifolia and A. rivalis. Bot. Sci. 2019, 97, 250–259. [Google Scholar] [CrossRef]
- Robinson, H. Generic and subtribal clasification of American Vernonieae. Smithsonian Contr. Bot. 1999, 89, 1–116. [Google Scholar] [CrossRef]
- Schilling, E.; Panero, J.L. The Eupatorieae Website. Available online: http://schillinglab.utk.edu/Danielweb/Eup/genusindex.html (accessed on 22 September 2020).
- GBIF, Global Biodiversity Information Facility. Available online: https://www.gbif.org/ (accessed on 21 September 2020).
- Bachmann, S.; Moat, J.; Hill, A.W.; de la Torre, J.; Scott, B. Supporting Red List threat assessments with GeoCAT: Geospatial conservation assessment tool. e-Infrastructures for data publishing in biodiversity science. ZooKeys 2011, 150, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Google Earth Pro. Available online: https://www.google.com/intl/es/earth/download/gep/agree.html (accessed on 4 December 2020).
- Instituto Nacional de Estadística y Geografía, Archivo Histórico de Localidades Geoestadísticas. Available online: https://www.inegi.org.mx/app/geo2/ahl/ (accessed on 2 December 2020).
- Mapcarta, El Mapa Libre. Available online: https://mapcarta.com/es/ (accessed on 2 December 2020).
- PueblosAmerica.com. Pueblos de México. Available online: https://mexico.pueblosamerica.com/ (accessed on 28 November 2020).
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Van Proosdij, A.S.J.; Sosef, M.S.M.; Wieringa, J.J.; Raes, N. Minimum required number of specimen records to develop accurate species distribution models. Ecography 2016, 39, 542–552. [Google Scholar] [CrossRef]
- WorldClim, Maps, Graphs, Tables, and Data of the Global Climate. Available online: https://www.worldclim.org/ (accessed on 5 January 2021).
- Mendes, P.; Elías Velasco, S.J.; Alves de Andrade, A.F.; De Marco Júnior, P. Dealing with overprediction in species distribution models: How adding distance constraints can improve model accuracy. Ecol. Model. 2020, 431, 109180. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Lobo, J.M. Threshold criteria for conversion of probability of species presence to either-or presence-absence. Acta Oecol. 2007, 31, 361–369. [Google Scholar] [CrossRef]
- Tatebe, H.; Ogura, T.; Nitta, T.; Komuro, Y.; Ogochi, K.; Takemura, T.; Sudo, K.; Sekiguchi, M.; Manabu, A.; Saito, F.; et al. Description and basic evaluation of simulated mean state, internal variability, and climate sensitivity in MIROC6. Geosci. Model Dev. 2019, 12, 2727–2765. [Google Scholar] [CrossRef]
- Fasullo, J.T. Evaluating Simulated Climate Patterns from the CMIP Archives Using Satellite and Reanalysis Datasets. Geosci. Model Dev. 2020, 13, 3627–3642. [Google Scholar] [CrossRef]
- Provincias Biogeográficas de México, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). 1997. Available online: http://conabio.gob.mx/informacion/metadata/gis/rbiog4mgw.xml?_xsl=/db/metadata/xsl/fgdc_html.xsl&_indent=no (accessed on 25 January 2021).
- International Union for Conservation of Nature. Red List Categories and Criteria, Version 3.1, 2nd ed.; IUCN: Gland, Switzerland; Cambridge, UK, 2012; p. 32. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).