
Physiological and Biochemical Responses of Ungrafted and Grafted Bell Pepper Plants (Capsicum annuum L. var. grossum (L.) Sendtn.) Grown under Moderate Salt Stress

 , ,
, , 

 ,
,
Abstract
1. Introduction
2. Results
2.1. Fruit Yield
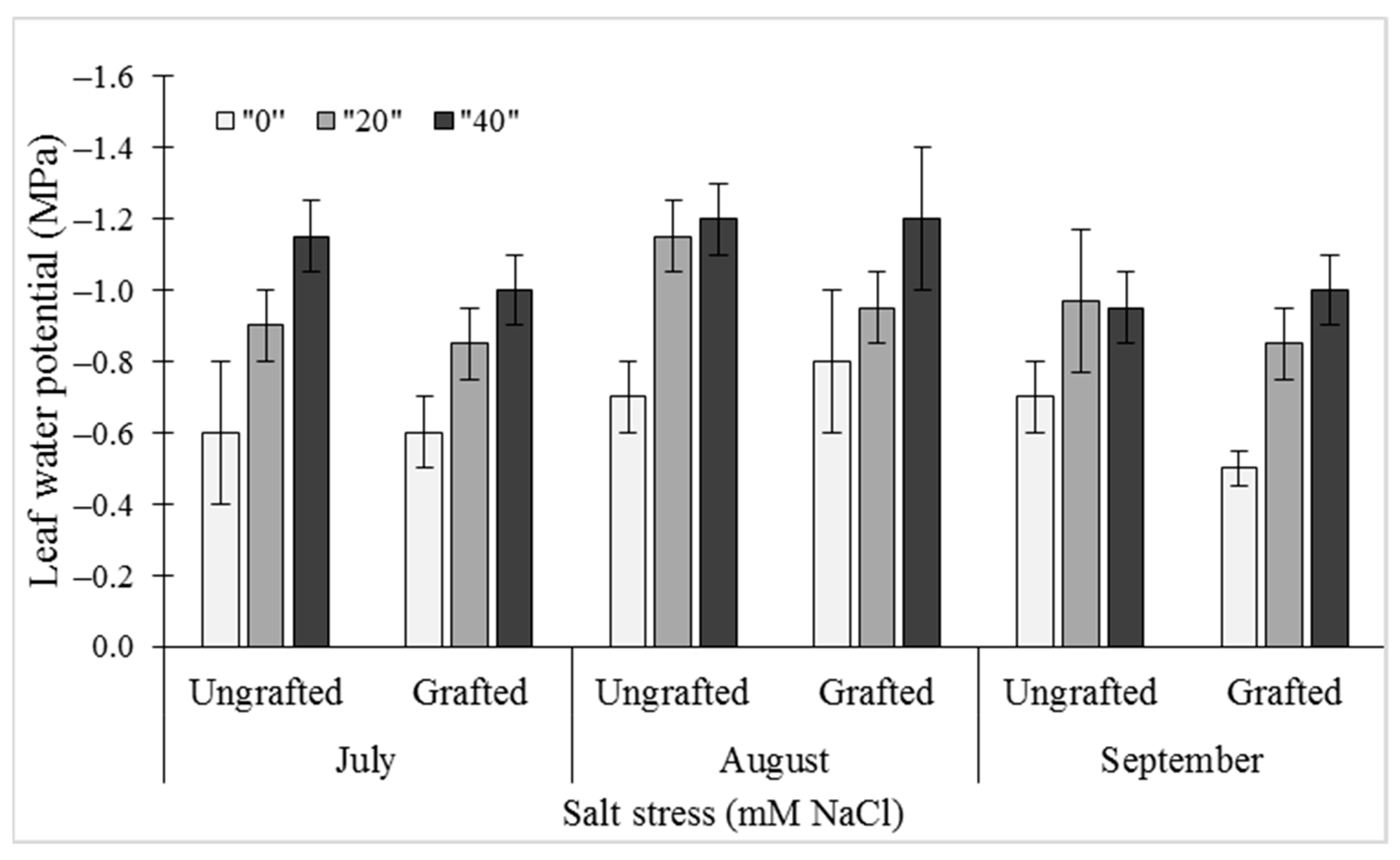
2.2. Water Relations
2.3. Photosynthetic Traits
2.4. Photosynthetic Pigment Characteristics and Xanthophyll Cycle Components
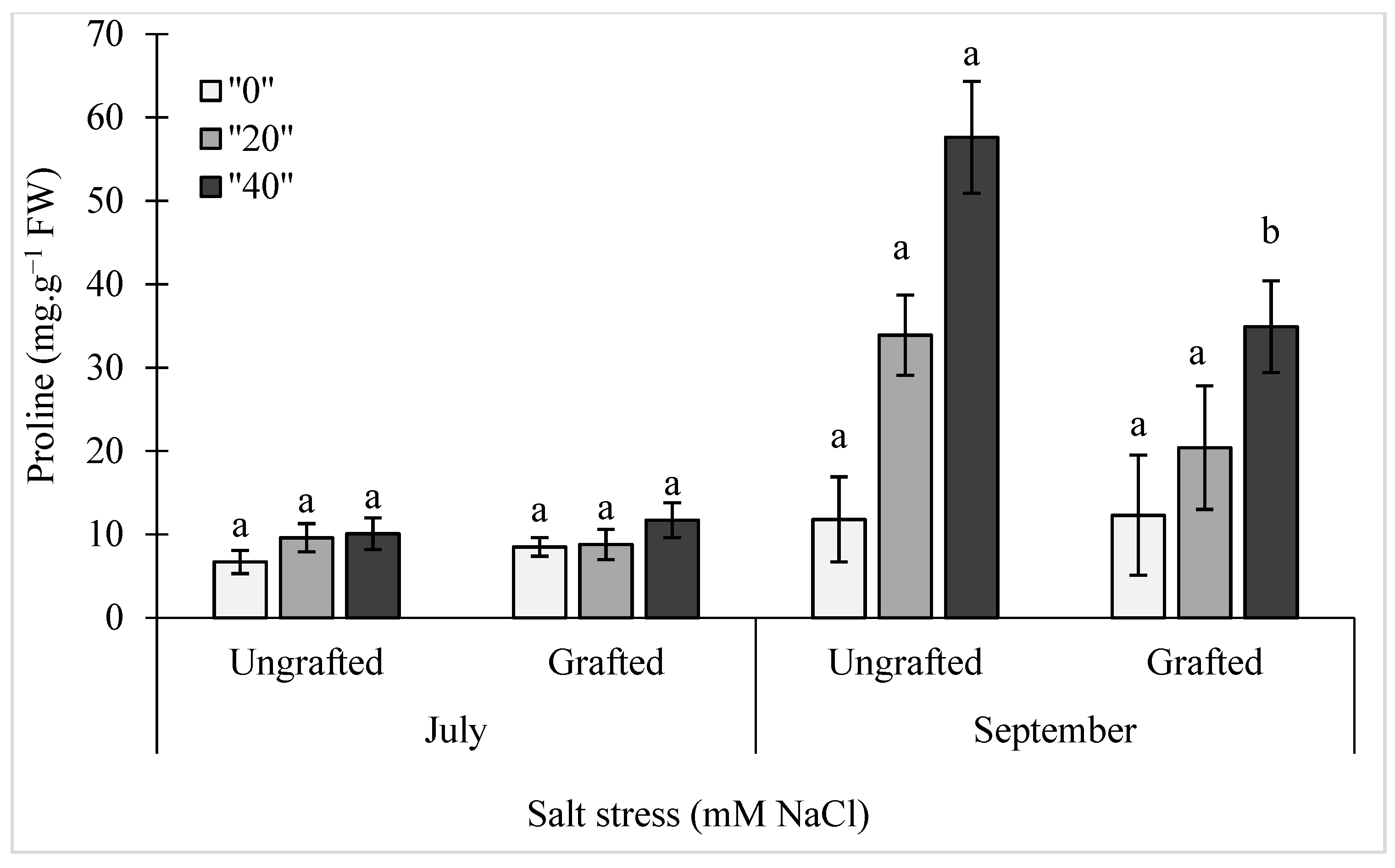
2.5. Proline Levels
2.6. Mineral Contents
2.7. Biochemical Parameters of the Pepper Fruit
3. Discussion
3.1. Physiological and Biochemical Parameters of Pepper Plants
3.2. Fruit Quality
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Leaf and Fruit Sampling
4.3. Proline Determination
4.4. Photosynthetic Measurements
4.5. Analysis of Photosynthetic Pigments
4.6. Analysis of Chloride Content
4.7. Analysis of Mineral Content
4.8. Analysis of Sugars and Organic Acids
4.9. Analysis of Ascorbic Acid
4.10. Analysis and HPLC-MSn Identification of Phenolic Compounds
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chartzoulakis, K.; Klapaki, G. Response of two greenhouse pepper hybrids to NaCl salinity during different growth stages. Sci. Hortic. 2000, 86, 247–260. [Google Scholar] [CrossRef]
- Navarro, J.; Garrido, C.; Carvajal, M.; Martinez, V. Yield and fruit quality of pepper plants under sulphate and chloride salinity. J. Hortic. Sci. Biotech. 2002, 77, 52–57. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M.; Al-Qurainy, F.; Harris, P.J. Salt tolerance in selected vegetable crops. Crit. Rev. Plant. Sci. 2012, 31, 303–320. [Google Scholar] [CrossRef]
- Martınez-Ballesta, M.; Martınez, V.; Carvajal, M. Osmotic adjustment, water relations and gas exchange in pepper plants grown under NaCl or KCl. Environ. Exp. Bot. 2004, 52, 161–174. [Google Scholar] [CrossRef]
- Aktas, H.; Abak, K.; Cakmak, I. Genotypic variation in the response of pepper to salinity. Sci. Hortic. 2006, 110, 260–266. [Google Scholar] [CrossRef]
- Bojórquez-Quintal, E.; Velarde-Buendía, A.; Ku-González, Á.; Carillo-Pech, M.; Ortega-Camacho, D.; Echevarría-Machado, I.; Pottosin, I.; Martínez-Estévez, M. Mechanisms of salt tolerance in habanero pepper plants (Capsicum chinense Jacq.): Proline accumulation, ions dynamics and sodium root-shoot partition and compartmentation. Front. Plant. Sci. 2014, 5, 605. [Google Scholar]
- Silva, C.; Martínez, V.; Carvajal, M. Osmotic versus toxic effects of NaCl on pepper plants. Biol. Plant. 2008, 52, 72–79. [Google Scholar] [CrossRef]
- Debez, A.; Koyro, H.W.; Grignon, C.; Abdelly, C.; Huchzermeyer, B. Relationship between the photosynthetic activity and the performance of Cakile maritima after long-term salt treatment. Physiol. Plant. 2008, 133, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yan, C.Q.; Cao, B.H.; Xu, H.X.; Chen, J.P.; Jiang, D.A. Some photosynthetic responses to salinity resistance are transferred into the somatic hybrid descendants from the wild soybean Glycine cyrtoloba ACC547. Physiol. Plantarum 2007, 129, 658–669. [Google Scholar] [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef]
- Khatri, K.; Rathore, M. Photosystem photochemistry, prompt and delayed fluorescence, photosynthetic responses and electron flow in tobacco under drought and salt stress. Photosynthetica 2019, 57, 61–74. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, V.; Prasad, S. Proline and salinity tolerance in plants. Biochem. Pharmacol. 2014, 3, e170. [Google Scholar] [CrossRef]
- Kotula, L.; Garcia Caparros, P.; Zörb, C.; Colmer, T.D.; Flowers, T.J. Improving crop salt tolerance using transgenic approaches: An update and physiological analysis. Plant Cell Environ. 2020, 43, 2932–2956. [Google Scholar] [CrossRef]
- Rouphael, Y.; Schwarz, D.; Krumbein, A.; Colla, G. Impact of grafting on product quality of fruit vegetables. Sci. Hortic. 2010, 127, 172–179. [Google Scholar] [CrossRef]
- Penella, C.; Landi, M.; Guidi, L.; Nebauer, S.G.; Pellegrini, E.; San Bautista, A.; Remorini, D.; Nali, C.; López-Galarza, S.; Calatayud, A. Salt-tolerant rootstock increases yield of pepper under salinity through maintenance of photosynthetic performance and sinks strength. J. Plant Physiol. 2016, 193, 1–11. [Google Scholar] [CrossRef]
- Penella, C.; Nebauer, S.G.; López-Galarza, S.; Quiñones, A.; San Bautista, A.; Calatayud, Á. Grafting pepper onto tolerant rootstocks: An environmental-friendly technique overcome water and salt stress. Sci. Hortic. 2017, 226, 33–41. [Google Scholar] [CrossRef]
- Singh, H.; Sethi, S.; Kaushik, P.; Fulford, A. Grafting vegetables for mitigating environmental stresses under climate change: A review. J. Water. Clim. Chang. 2020, 11, 1784–1797. [Google Scholar] [CrossRef]
- Albacete, A.; Martínez-Andújar, C.; Ghanem, M.E.; Acosta, M.; Sánchez-Bravo, J.; Asins, M.J.; Cuartero, J.; Lutts, S.; Dodd, I.C.; Pérez-Alfocea, F. Rootstock-mediated changes in xylem ionic and hormonal status are correlated with delayed leaf senescence, and increased leaf area and crop productivity in salinized tomato. Plant Cell Environ. 2009, 32, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Giuffrida, F.; Cassaniti, C.; Leonardi, C. The influence of rootstock on growth and ion concentrations in pepper (Capsicum annuum L.) under saline conditions. Hortic. Sci. Biotech. 2013, 88, 110–116. [Google Scholar] [CrossRef]
- Serrano, L.L.; Penella, C.; San Bautista, A.; Galarza, S.L.; Chover, A.C. Physiological changes of pepper accessions in response to salinity and water stress. Spanish. J. Agric. Res. 2017, 15, 15. [Google Scholar]
- Lee, J.-M.; Kubota, C.; Tsao, S.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable grafting: The implications of a growing agronomic imperative for vegetable fruit quality and nutritive value. Front. Plant. Sci. 2017, 8, 741. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Temperini, O.; Rea, E.; Salerno, A.; Pierandrei, F. Influence of grafting on yield and fruit quality of pepper (Capsicum annuum L.) grown under greenhouse conditions. In Proceedings of the IV International Symposium on Seed, Transplant and Stand Establishment of Horticultural Crops, San Antonio, TX, USA, 3 December 2006; Translating Seed and Seedling 782; International Society for Horticultural Science: Leuven, Belgium, 2006; pp. 359–364. [Google Scholar]
- Chávez-Mendoza, C.; Sánchez, E.; Carvajal-Millán, E.; Muñoz-Márquez, E.; Guevara-Aguilar, A. Characterization of the nutraceutical quality and antioxidant activity in bell pepper in response to grafting. Molecules 2013, 18, 15689–15703. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Leonardi, C.; Bie, Z. Role of grafting in vegetable crops grown under saline conditions. Sci. Hortic. 2010, 127, 147–155. [Google Scholar] [CrossRef]
- Leal-Fernández, C.; Godoy-Hernández, H.; Núñez-Colín, C.; Anaya-López, J.; Villalobos-Reyes, S.; Castellanos, J. Morphological response and fruit yield of sweet pepper (Capsicum annuum L.) grafted onto different commercial rootstocks. Biol. Agric. Hortic. 2013, 29, 1–11. [Google Scholar] [CrossRef]
- Ropokis, A.; Ntatsi, G.; Kittas, C.; Katsoulas, N.; Savvas, D. Effects of temperature and grafting on yield, nutrient uptake, and water use efficiency of a hydroponic sweet pepper crop. Agron 2019, 9, 110. [Google Scholar] [CrossRef]
- Gisbert Domenech, M.C.; Sánchez-Torres, P.; Raigón Jiménez, M.; Nuez Viñals, F. Phytophthora capsici resistance evaluation in pepper hybrids. Agronomic performance and fruit quality of pepper grafted plants. J. Food Agric. Environ. 2010, 8, 116–121. [Google Scholar]
- López-Marín, J.; González, A.; Pérez-Alfocea, F.; Egea-Gilabert, C.; Fernández, J.A. Grafting is an efficient alternative to shading screens to alleviate thermal stress in greenhouse-grown sweet pepper. Sci. Hortic. 2013, 149, 39–46. [Google Scholar] [CrossRef]
- López-Marín, J.; Gálvez, A.; del Amor, F.M.; Albacete, A.; Fernández, J.A.; Egea-Gilabert, C.; Pérez-Alfocea, F. Selecting vegetative/generative/dwarfing rootstocks for improving fruit yield and quality in water stressed sweet peppers. Sci. Hortic. 2017, 214, 9–17. [Google Scholar] [CrossRef]
- Sánchez-Torres, P.; Raigón, M.D.; Gammoudi, N.; Gisbert, C. Effects of grafting combinations on the nutritional composition of pepper fruit. Fruits 2016, 71, 249–256. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable grafting: A toolbox for securing yield stability under multiple stress conditions. Front. Plant. Sci. 2018, 8, 2255. [Google Scholar] [CrossRef]
- López-Serrano, L.; Canet-Sanchis, G.; Selak, G.V.; Penella, C.; San Bautista, A.; López-Galarza, S.; Calatayud, Á. Physiological characterization of a pepper hybrid rootstock designed to cope with salinity stress. Plant Physiol. Biochem. 2020, 148, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Wen, D.; VandenLangenberg, K.; Wei, M.; Yang, F.; Shi, Q.; Wang, X. Comparative effects of NaCl and NaHCO3 stress on photosynthetic parameters, nutrient metabolism, and the antioxidant system in tomato leaves. Sci. Hortic. 2013, 157, 1–12. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Phys. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhu, Z.; Yang, J.; Ni, X.; Zhu, B. Grafting increases the salt tolerance of tomato by improvement of photosynthesis and enhancement of antioxidant enzymes activity. Environ. Exp. Bot. 2009, 66, 270–278. [Google Scholar] [CrossRef]
- Penella, C.; Nebauer, S.G.; Quinones, A.; San Bautista, A.; Lopez-Galarza, S.; Calatayud, A. Some rootstocks improve pepper tolerance to mild salinity through ionic regulation. Plant. Sci. 2015, 230, 12–22. [Google Scholar] [CrossRef]
- Esasem. 2018. Available online: https://www.esasem.com/en/sale-of-vegetable-seeds/rootstock/ (accessed on 10 November 2019).
- Calatayud, A.; Barreno, E. Response to ozone in two lettuce varieties on chlorophyll a fluorescence, photosynthetic pigments and lipid peroxidation. Plant. Physiol. Bioch. 2004, 42, 549–555. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Bioch. Biophys. Acta (BBA)-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Estan, M.T.; Martinez-Rodriguez, M.M.; Perez-Alfocea, F.; Flowers, T.J.; Bolarin, M.C. Grafting raises the salt tolerance of tomato through limiting the transport of sodium and chloride to the shoot. J. Exp. Bot. 2005, 56, 703–712. [Google Scholar] [CrossRef]
- Zhu, J.; Bie, Z.; Huang, Y.; Han, X. Effect of grafting on the growth and ion concentrations of cucumber seedlings under NaCl stress. Soil. Sci. Plant. Nutr. 2008, 54, 895–902. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef] [PubMed]
- Rengel, Z. The role of calcium in salt toxicity. Plant Cell Environ. 1992, 15, 625–632. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier Science Publishing Co Inc., Academic Press Inc.: San Diego, CA, USA, 2011; 672p. [Google Scholar]
- Geilfus, C.-M. Chloride: From nutrient to toxicant. Plant Cell Physiol. 2018, 59, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends. Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [PubMed]
- Louws, F.J.; Rivard, C.L.; Kubota, C. Grafting fruiting vegetables to manage soilborne pathogens, foliar pathogens, arthropods and weeds. Sci. Hortic. 2010, 127, 127–146. [Google Scholar] [CrossRef]
- Marin, A.; Rubio, J.S.; Martínez, V.; Gil, M.I. Antioxidant compounds in green and red peppers as affected by irrigation frequency, salinity and nutrient solution composition. J. Sci. Food. Agric. 2009, 89, 1352–1359. [Google Scholar] [CrossRef]
- Serio, F.; Gara, L.D.; Caretto, S.; Leo, L.; Santamaria, P. Influence of an increased NaCl concentration on yield and quality of cherry tomato grown in posidonia (Posidonia oceanica (L) Delile). J. Sci. Food Agric. 2004, 84, 1885–1890. [Google Scholar] [CrossRef]
- Navarro, J.M.; Flores, P.; Garrido, C.; Martinez, V. Changes in the contents of antioxidant compounds in pepper fruits at different ripening stages, as affected by salinity. Food Chem. 2006, 96, 66–73. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef]
- Kacjan Marsic, N.; Vodnik, D.; Mikulic-Petkovsek, M.; Veberic, R.; Sircelj, H. Photosynthetic traits of plants and the biochemical profile of tomato fruits are influenced by grafting, salinity stress, and growing season. J. Agric. Food. Chem. 2018, 66, 5439–5450. [Google Scholar] [CrossRef] [PubMed]
- Koleška, I.; Hasanagić, D.; Oljača, R.; Murtić, S.; Bosančić, B.; Todorović, V. Influence of grafting on the copper concentration in tomato fruits under elevated soil salinity. АГРОЗНАЊЕ 2019, 20, 37–44. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Borošić, J.; Benko, B.; Fabek, S.; Novak, B.; Dobričević, N.; Bućan, L. Agronomic traits of soilless grown bell pepper. In Proceedings of the28th International Horticultural Congress on Science and Horticulture for People (IHC2010), Lisbon, Portugal, 22–27 August 2010; pp. 421–428. [Google Scholar]
- Drăgici, E.M.; Brezeanu, P.M.; Muñoz, P.; Brezeanu, C. Pepper and eggplant. In Good Agricultural Practices for Greenhouse Vegetable Production in the South East European Countries-Principles for Sustainable Intensification of Smallholder Farms, 1st ed.; Baudoin, W., Nersisyan, A., Shamilov, A., Hodder, A., Gutierrez, D., Eds.; FAO: Rome, Italy, 2017; Volume 230, pp. 301–307. [Google Scholar]
- Nagaz, K.; Masmoudi, M.M.; Mechlia, N.B. Effects of deficit drip-irrigation scheduling regimes with saline water on pepper yield, water productivity and soil salinity under arid conditions of Tunisia. J. Agric. Environ. Inter. Develop. (JAEID) 2012, 106, 85–103. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant. Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Guidi, L.; Degl’Innocenti, E.; Remorini, D.; Massai, R.; Tattini, M. Interactions of water stress and solar irradiance on the physiology and biochemistry of Ligustrum vulgare. Tree Physiol. 2008, 28, 873–883. [Google Scholar] [CrossRef]
- Sircelj, H.; Batic, F. Evaluation of selected nutritional factors in Aposeris foetida (L.) Less. during the harvesting period. J. Appl. Bot. Food. Qual.-Angew. Bot. 2007, 81, 121–125. [Google Scholar]
- Nielsen, S.S. Sodium determination using ion-selective electrodes, Mohr titration, and test strips. In Food Analysis Laboratory Manual, Springer: Berlin, Germany, 2017; pp. 161–170.
- Mikulic-Petkovsek, M.; Ivancic, A.; Schmitzer, V.; Veberic, R.; Stampar, F. Comparison of major taste compounds and antioxidative properties of fruits and flowers of different Sambucus species and interspecific hybrids. Food Chem. 2016, 200, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Mikulic-Petkovsek, M.; Schmitzer, V.; Jakopic, J.; Cunja, V.; Veberic, R.; Munda, A.; Stampar, F. Phenolic compounds as defence response of pepper fruits to Colletotrichum coccodes. Physiol. Mol. Plant P 2013, 84, 138–145. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Slatnar, A.; Stampar, F.; Veberic, R. HPLC–MSn identification and quantification of flavonol glycosides in 28 wild and cultivated berry species. Food Chem. 2012, 135, 2138–2146. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Zheng, W.; Galletta, G.J. Cultural system affects fruit quality and antioxidant capacity in strawberries. J. Agric. Food. Chem. 2002, 50, 6534–6542. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing (Version 3.1. 2); R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]



{kind=link}
{kind=link}
| Salinity (mM NaCl) | Marketable Yield (kg Plant–1) | Blossom End Rot Infection (%) | Plant Biomass (kg) | |||
|---|---|---|---|---|---|---|
| Ungrafted | Grafted | Ungrafted | Grafted | Ungrafted | Grafted | |
| 0 | 1.34 ± 0.2 | 1.12 ± 0.1 | 25.5 ± 5 *** | 42.2 ± 5 | 0.970 ± 0.20 | 0.78 ± 0.06 |
| 20 | 0.96 ± 0.3 | 0.84 ± 0.1 | 65.9 ± 11 | 65.6 ± 11 | 0.777 ± 0.03 | 0.63 ± 0.02 |
| 40 | 0.64 ± 0.3 | 0.51 ± 0.1 | 72.5 ± 8 *** | 52.0 ± 8 | 0.577 ± 0.02 | 0.44 ± 0.07 |
| Significance | ||||||
| Salinity (S) | * | * | * | |||
| Grafting (G) | ns | ns | ns | |||
| S × G | ns | ** | ns | |||
| Salinity | Grafting | Net CO2 Assimilation Rate (Pn) | Stomatal Conductance (gs) | Transpiration Rate (E) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (mM NaCl) | July | August | September | July | August | September | July | August | September | |
| 0 | Ungrafted | 20.3 ± 1.1 | 14.4 ± 1.9 | 17.7 ± 0.6 | 0.69 ± 0.12 | 0.25 ± 0.04 | 0.50 ± 0.02 | 5.83 ± 0.55 | 3.91 ± 0.74 | 3.28 ± 0.43 |
| Grafted | 17.4 ± 2.2 | 12.0 ± 0.6 | 18.0 ± 0.6 | 0.53 ± 0.2 | 0.22 ± 0.04 | 0.61 ± 0.03 | 5.05 ± 1.42 | 3.80 ± 0.57 | 3.50 ± 0.40 | |
| 20 | Ungrafted | 18.0 ± 0.8 | 11.3 ± 0.8 | 15.2 ± 2.3 | 0.51 ± 0.12 | 0.15 ± 0.02 | 0.42 ± 0.09 | 5.45 ± 0.70 | 2.82 ± 0.33 | 2.72 ± 0.45 |
| Grafted | 20.5 ± 1.6 | 15.6 ± 0.6 | 18.1 ± 0.9 | 0.76 ± 0.09 | 0.27 ± 0.04 | 0.66 ± 0.08 | 6.92 ± 0.50 | 4.70 ± 0.64 | 3.87 ± 0.41 | |
| 40 | Ungrafted | 18.2 ± 1.7 | 8.6 ± 0.6 | 9.0 ± 2.4 | 0.54 ± 0.14 | 0.12 ± 0.01 | 0.21 ± 0.05 | 5.19 ± 0.97 | 2.10 ± 0.31 | 1.76 ± 0.14 |
| Grafted | 17.3 ± 3.5 | 12.1 ± 2.6 | 12.2 ± 2.3 | 0.45 ± 0.13 | 0.17 ± 0.05 | 0.27 ± 0.05 | 4.87 ± 1.12 | 3.36 ± 0.97 | 2.10 ± 0.47 | |
| Significance | ||||||||||
| Salinity (S) | * | * | ns | |||||||
| Grafting (G) | ns | ns | ns | |||||||
| Date (D) | *** | *** | *** | |||||||
| S × G | * | ns | ns | |||||||
| S × D | ns | ns | ns | |||||||
| G × D | ns | ns | ns | |||||||
| S × G × D | ns | ns | ns | |||||||
| Salinity (mM NaCl) | Grafting | Maximum Quantum Use Efficiency of PSII in Dark-Adapted State (Fv/Fm), | Effective Quantum Use Efficiency of PSII in Light-Adapted State (Fv′/Fm′) | Photochemical Quenching (qP) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| July | August | September | July | August | September | July | August | September | ||
| 0 | Grafted | 0.76 ± 0.00 | 0.76 ± 0.00 | 0.70 ± 0.06 | 0.53 ± 0.02 | 0.51 ± 0.02 | 0.57 ± 0.02 | 0.45 ± 0.02 | 0.38 ± 0.02 | 0.41 ± 0.01 |
| Ungrafted | 0.76 ± 0.00 | 0.76 ± 0.00 | 0.76 ± 0.00 | 0.56 ± 0.02 | 0.52 ± 0.03 | 0.55 ± 0.02 | 0.48 ± 0.02 | 0.43 ± 0.04 | 0.41 ± 0.01 | |
| 20 | Grafted | 0.76 ± 0.00 | 0.76 ± 0.00 | 0.76 ± 0.00 | 0.56 ± 0.01 | 0.53 ± 0.01 | 0.57 ± 0.00 | 0.49 ± 0.02 | 0.46 ± 0.01 | 0.37 ± 0.00 |
| Ungrafted | 0.76 ± 0.00 | 0.76 ± 0.00 | 0.72 ± 0.04 | 0.54 ± 0.02 | 0.48 ± 0.01 | 0.54 ± 0.04 | 0.45 ± 00.02 | 0.43 ± 0.03 | 0.39 ± 0.01 | |
| 40 | Grafted | 0.76 ± 0.00 | 0.76 ± 0.00 | 0.71 ± 0.06 | 0.52 ± 0.04 | 0.49 ± 0.02 | 0.49 ± 0.03 | 0.46 ± 0.02 | 0.42 ± 0.00 | 0.40 ± 0.02 |
| Ungrafted | 0.76 ± 0.00 | 0.76 ± 0.00 | 0.71 ± 0.05 | 0.53 ± 0.02 | 0.45 ± 0.02 | 0.46 ± 0.05 | 0.47 ± 0.01 | 0.43 ± 0.00 | 0.41 ± 0.00 | |
| Significance | ||||||||||
| Salinity (S) | ns | * | ns | |||||||
| Grafting (G) | ns | ns | ns | |||||||
| Date (D) | * | *** | *** | |||||||
| S × G | ns | ns | ns | |||||||
| S × D | ns | ns | ns | |||||||
| G × D | ns | ns | ns | |||||||
| S × G × D | ns | ns | ns | |||||||
| Salinity (mM NaCl) | Grafting | Total Chlorophyll Content (Chla+b) | Total Violaxanthin, Antheraxanthin, and Zeaxanthin | De-Epoxidation State (EOS) | |||
|---|---|---|---|---|---|---|---|
| July | September | July | September | July | September | ||
| 0 | Ungrafted | 10.98 ± 0.22 | 10.88 ± 0.10 | 0.52 ± 0.01 | 0.57 ± 0.01 | 0.57 ± 0.01 | 0.18 ± 0.02 |
| Grafted | 11.02 ± 0.21 | 10.77 ± 0.11 | 0.55 ± 0.03 | 0.54 ± 0.03 | 0.54 ± 0.03 | 0.13 ± 0.03 | |
| 20 | Ungrafted | 10.62 ± 0.42 | 10.42 ± 0.13 | 0.50 ± 0.03 | 0.51 ± 0.03 | 0.51 ± 0.03 | 0.14 ± 0.03 |
| Grafted | 10.78 ± 0.02 | 10.74 ± 0.12 | 0.50 ± 0.01 | 0.53 ± 0.02 | 0.53 ± 0.02 | 0.11 ± 0.01 | |
| 40 | Ungrafted | 10.06 ± 0.22 | 9.59 ± 0.39 | 0.42 ± 0.01 | 0.43 ± 0.03 | 0.43 ± 0.03 | 0.11 ± 0.01 |
| Grafted | 10.79 ± 0.51 | 10.24 ± 0.13 | 0.46 ± 0.03 | 0.43 ± 0.02 | 0.43 ± 0.02 | 0.13 ± 0.01 | |
| Significance | |||||||
| Salinity (S) | ns | ** | ns | ||||
| Grafting (G) | * | ns | ns | ||||
| Date (D) | ** | ns | *** | ||||
| S × G | ns | ns | ns | ||||
| S × D | ns | ns | ns | ||||
| G × D | ns | ns | ns | ||||
| S×G×D | ns | ns | ns | ||||
| Salinity | Grafting | Leaf Mineral Concentration (mg·g–1 Fresh Weight) | ||||
|---|---|---|---|---|---|---|
| (mM NaCl) | Na+ | Cl– | K+ | Ca2+ | Mg2+ | |
| Ungrafted | 0.014 ± 0.01 | 0.17 ± 0.052 | 61.5 ± 7.38 | 20.2 ± 4.90 | 10.2 ± 1.11 | |
| 0 | Grafted | 0.016 ± 0.01 | 0.15 ± 0.042 | 51.5 ± 6.01 | 20.6 ± 5.17 | 10.0 ± 1.48 |
| Ungrafted | 0.557 ± 0.35 | 2.84 ± 0.563 | 54.4 ± 5.40 | 21.1 ± 4.58 | 10.4 ± 2.00 | |
| 20 | Grafted | 0.291 ± 0.09 | 2.98 ± 0.558 | 54.8 ± 7.96 | 23.3 ± 8.35 | 11.5 ± 2.87 |
| Ungrafted | 1.190 ± 0.61 | 4.73 ± 0.414 | 56.4 ± 8.26 | 28.9 ± 5.98 | 11.8 ± 1.01 | |
| 40 | Grafted | 0.955 ± 0.35 | 5.61 ± 0.622 *** | 52.8 ± 4.53 | 27.6 ± 6.98 | 12.0 ± 2.20 |
| Significance | ||||||
| Salinity (S) | *** | *** | ns | * | * | |
| Grafting (G) | ns | ns | ns | ns | ns | |
| S × G | ns | * | ns | ns | ns | |
| Salinity | Grafting | Total Sugars | Total Organic Acids | Ascorbic Acid | Total Phenolics Analysed | ||||
|---|---|---|---|---|---|---|---|---|---|
| (mM NaCl) | (g·kg–1 Fresh Weight) | (g·kg–1 Fresh Weight) | (mg·[100 g]–1 Fresh Weight) | (mg·[100 g]–1 Fresh Weight) | |||||
| July | September | July | September | July | September | July | September | ||
| 0 | Grafted | 2790 ± 438 | 2460 ± 447 | 226 ± 27 | 192 ± 23 | 50.5 ± 28 | 57.4 ± 27 | 5.96 ± 2.6 | 4.47 ± 1.4 |
| Ungrafted | 2710 ± 638 | 2630 ± 370 | 230 ± 41 | 203 ± 42 | 34.8 ± 15 | 57.8 ± 25 | 6.43 ± 2.1 | 3.34 ± 1.7 | |
| 20 | Grafted | 2680 ± 324 | 3530 ± NA | 220 ± 35 | 245 ± NA | 44.9 ± 19 | 82.9 ± NA | 5.24 ± 0.8 | 1.8 ± NA |
| Ungrafted | 2990 ± 280 | 2940 ± 197 | 252 ± 34 | 317 ± 73 | 51.2 ± 24 | 82.9 ± 14 | 5.84 ± 1.3 | 2.87 ± 0.9 | |
| 40 | Grafted | 2760 ± 359 | 2690 ± 1090 | 249 ± 24 | 371 ± 50 | 57.4 ± 19 | 92.6 ± 9 | 6.77 ± 1.2 * | 2.99 ± 0.5 |
| Ungrafted | 2970 ± 559 | 3480 ± 306 | 235 ± 18 | 415 ± 47 | 49.9 ± 16 | 102 ± 13 | 4.85 ± 1.1 | 3.15 ± 0.4 | |
| Significance | |||||||||
| Salinity (S) | Ns | *** | ** | ns | |||||
| Grafting (G) | Ns | * | ns | ns | |||||
| Date (D) | Ns | *** | *** | *** | |||||
| S × G | Ns | * | ns | ns | |||||
| S × D | Ns | *** | ** | ns | |||||
| G × D | Ns | ns | ns | ns | |||||
| S × G × D | Ns | ns | ns | * | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kacjan Maršić, N.; Štolfa, P.; Vodnik, D.; Košmelj, K.; Mikulič-Petkovšek, M.; Kump, B.; Vidrih, R.; Kokalj, D.; Piskernik, S.; Ferjančič, B.; et al. Physiological and Biochemical Responses of Ungrafted and Grafted Bell Pepper Plants (Capsicum annuum L. var. grossum (L.) Sendtn.) Grown under Moderate Salt Stress. Plants 2021, 10, 314. https://doi.org/10.3390/plants10020314
Kacjan Maršić N, Štolfa P, Vodnik D, Košmelj K, Mikulič-Petkovšek M, Kump B, Vidrih R, Kokalj D, Piskernik S, Ferjančič B, et al. Physiological and Biochemical Responses of Ungrafted and Grafted Bell Pepper Plants (Capsicum annuum L. var. grossum (L.) Sendtn.) Grown under Moderate Salt Stress. Plants. 2021; 10(2):314. https://doi.org/10.3390/plants10020314
Chicago/Turabian StyleKacjan Maršić, Nina, Petra Štolfa, Dominik Vodnik, Katarina Košmelj, Maja Mikulič-Petkovšek, Bojka Kump, Rajko Vidrih, Doris Kokalj, Saša Piskernik, Blaz Ferjančič, and et al. 2021. "Physiological and Biochemical Responses of Ungrafted and Grafted Bell Pepper Plants (Capsicum annuum L. var. grossum (L.) Sendtn.) Grown under Moderate Salt Stress" Plants 10, no. 2: 314. https://doi.org/10.3390/plants10020314
APA StyleKacjan Maršić, N., Štolfa, P., Vodnik, D., Košmelj, K., Mikulič-Petkovšek, M., Kump, B., Vidrih, R., Kokalj, D., Piskernik, S., Ferjančič, B., Dragutinović, M., Veberič, R., Hudina, M., & Šircelj, H. (2021). Physiological and Biochemical Responses of Ungrafted and Grafted Bell Pepper Plants (Capsicum annuum L. var. grossum (L.) Sendtn.) Grown under Moderate Salt Stress. Plants, 10(2), 314. https://doi.org/10.3390/plants10020314