
Accumulation of Silicon and Changes in Water Balance under Drought Stress in Brassica napus var. napus L.




Abstract
1. Introduction
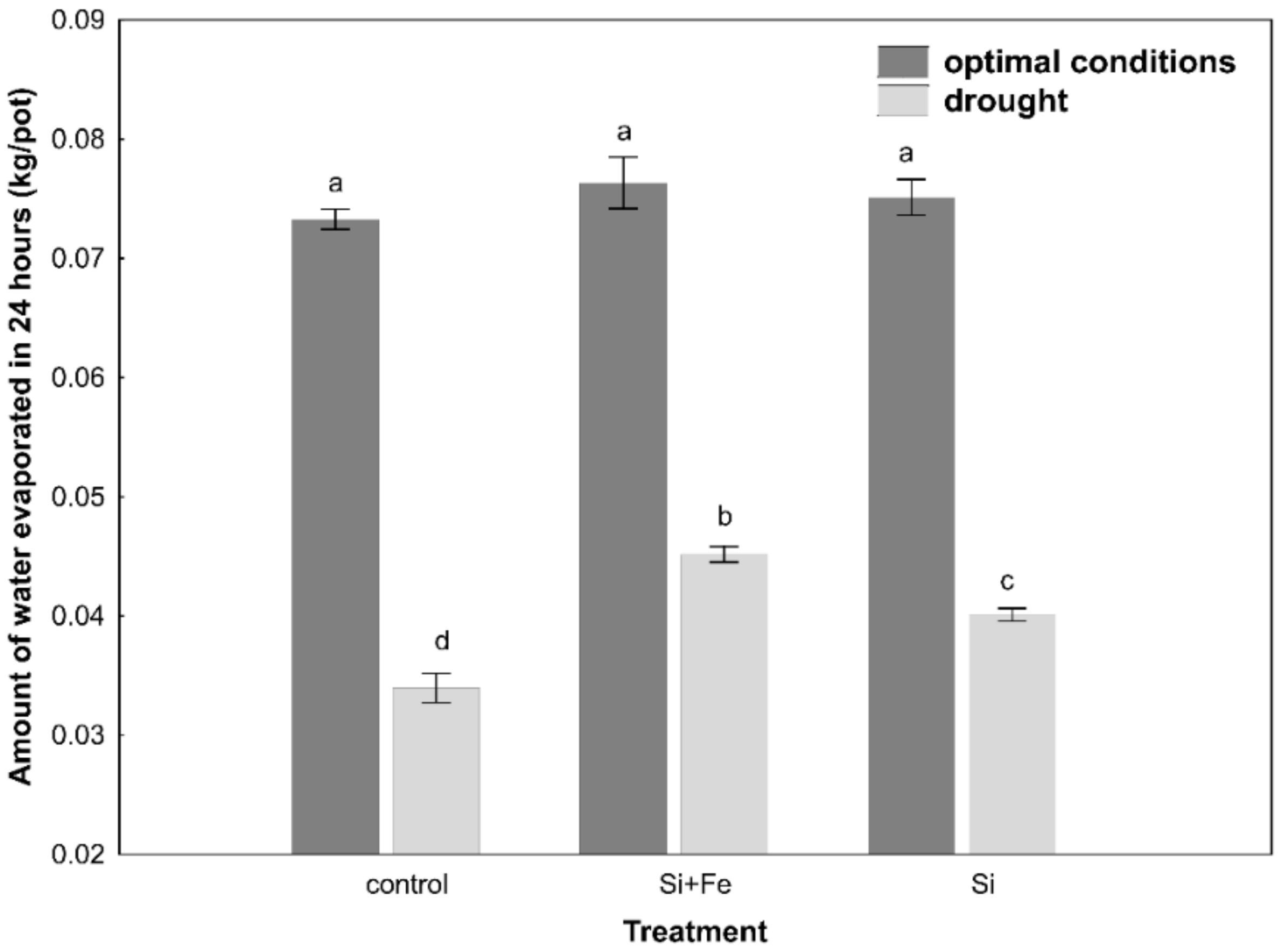
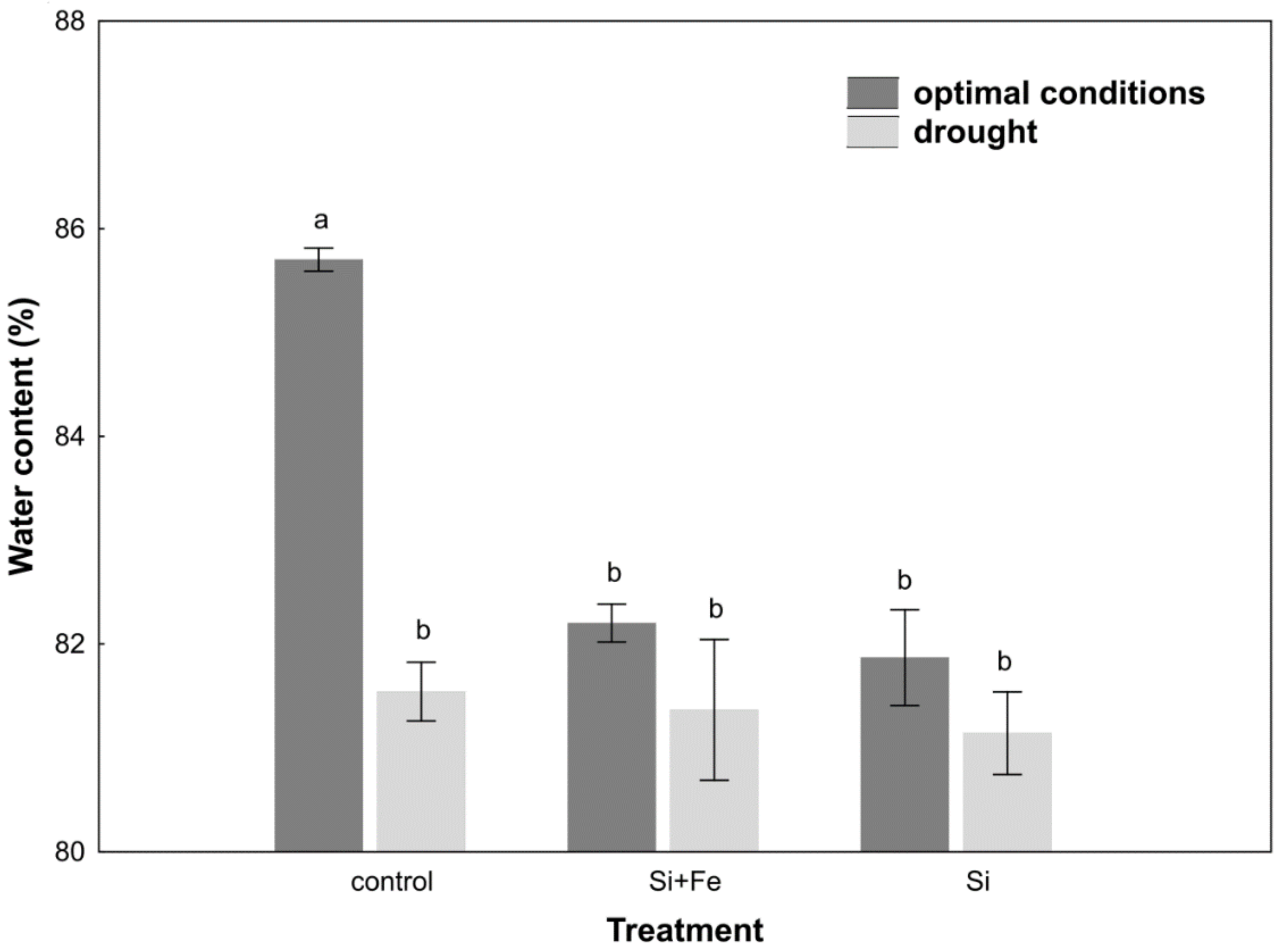
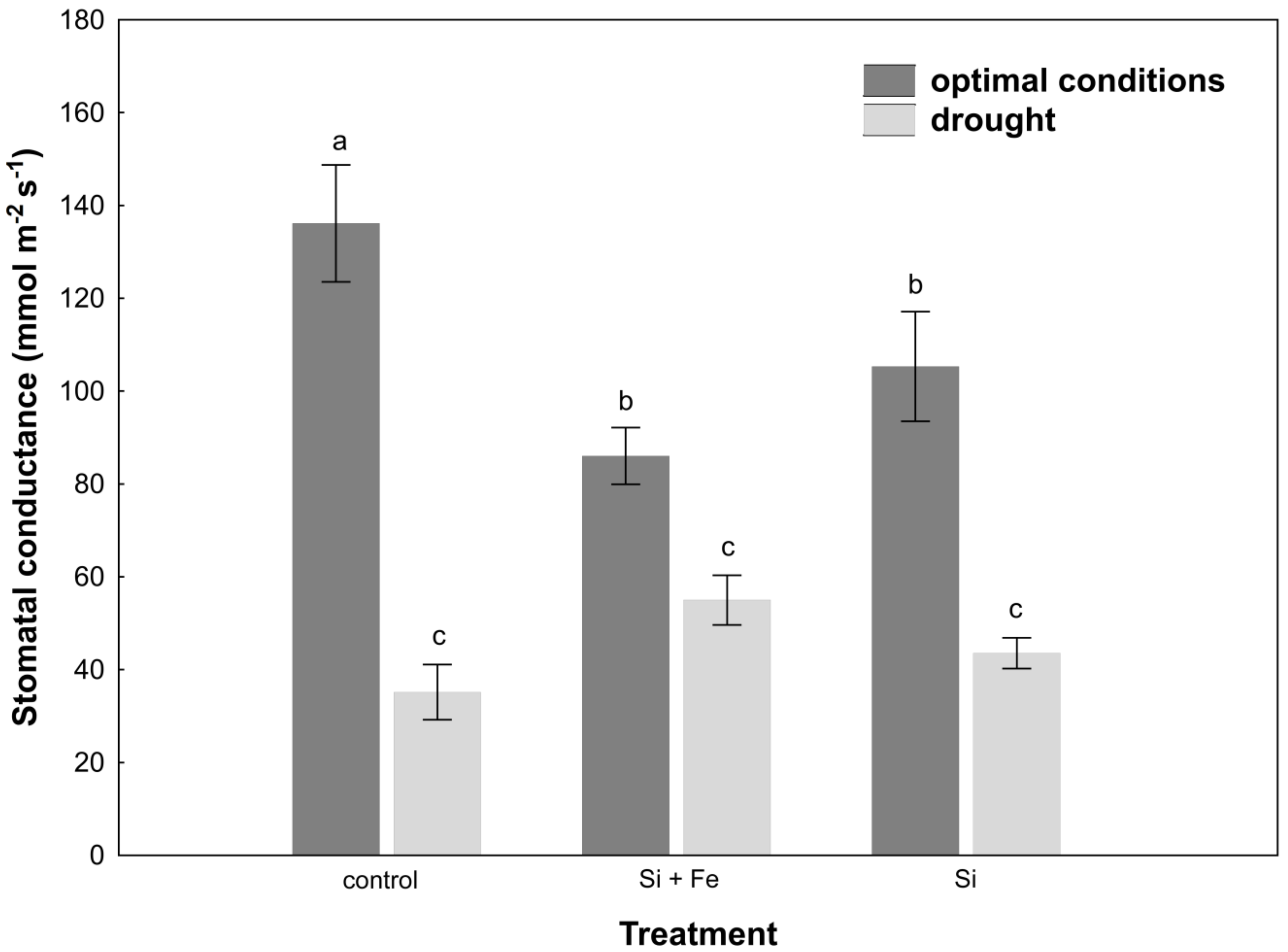
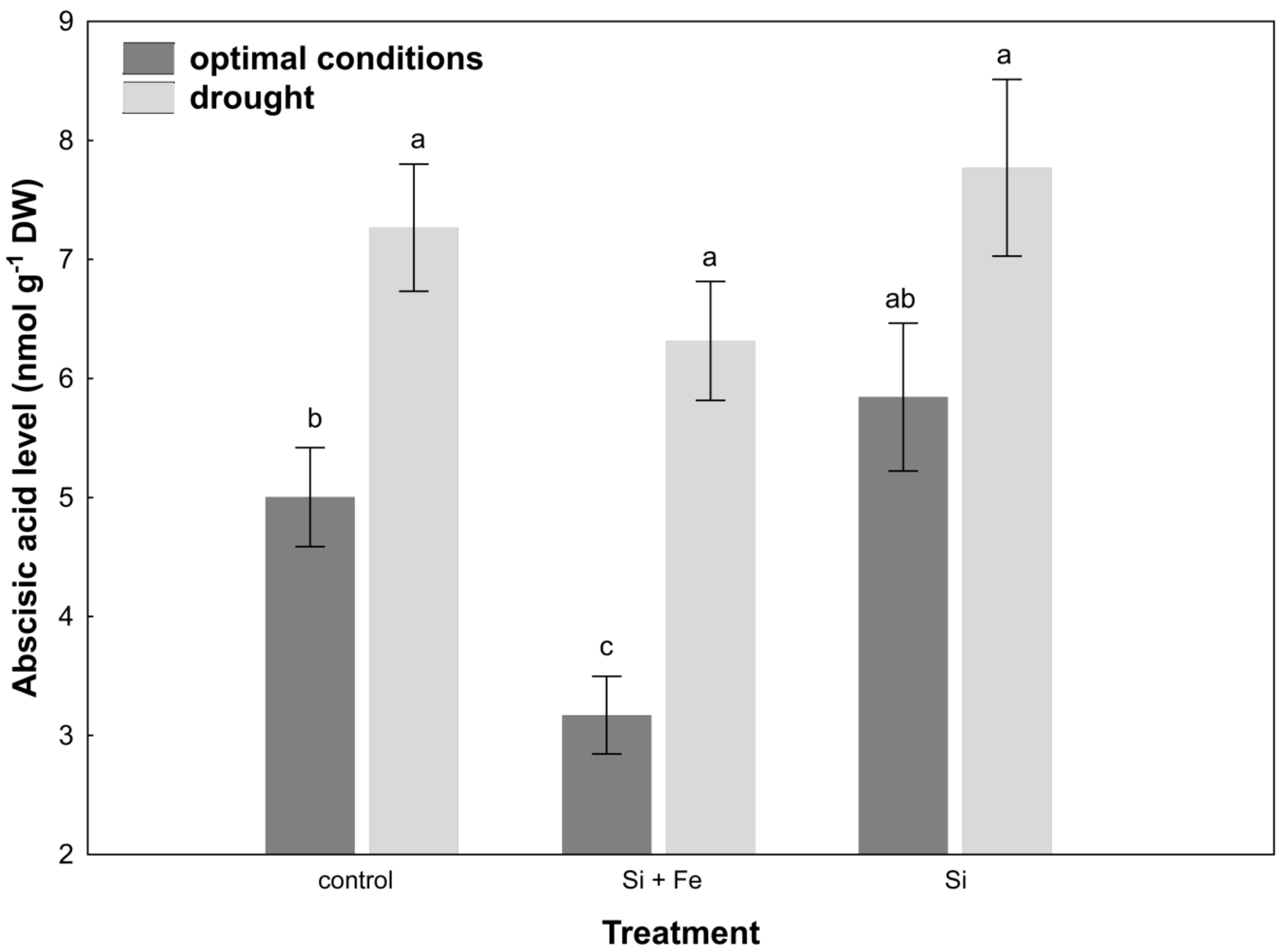
2. Results
3. Discussion
4. Materials and Methods
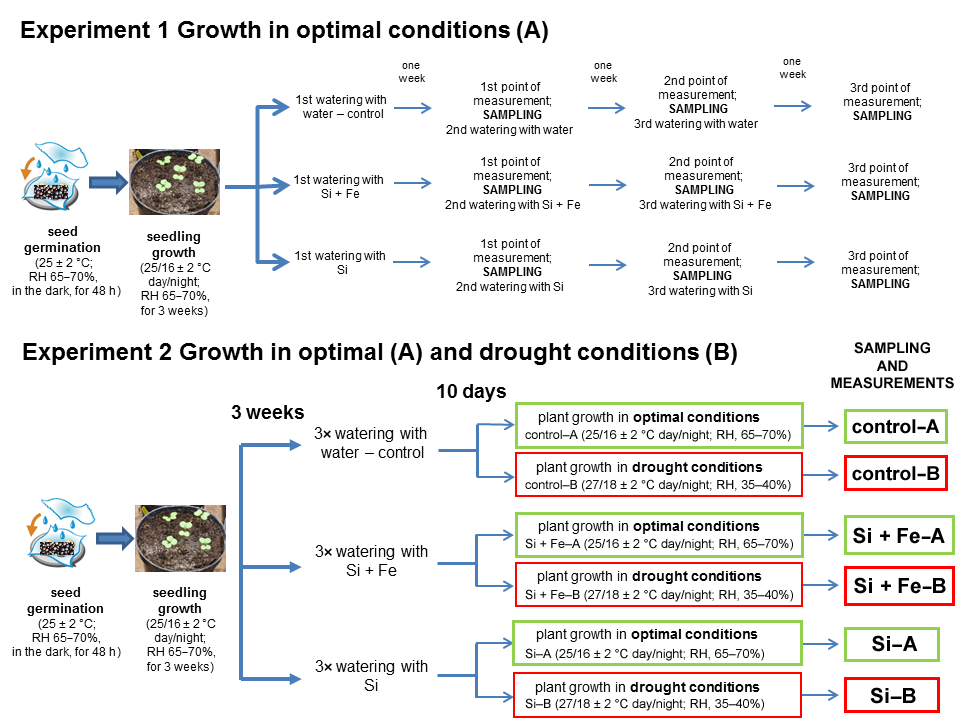
4.1. Plant Materials and Growth under Control Conditions
4.2. Silicon Treatment
4.3. Growth under Drought Conditions
4.4. Total Silicon Content in Plants
4.5. Leaf Water Content (LWC)
4.6. Osmotic and Water Potential
4.7. Estimation of Evapotranspiration
4.8. Leaf Stomatal Conductance Measurement
4.9. ABA Level Measurement
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Afzal, Z.; Howton, T.C.; Sun, Y.L.; Mukhtar, M.S. The roles of aquaporins in plant stress responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef]
- Wang, Z.L.; Wang, J.; Wang, J.S. Risk assessment of agricultural drought disaster in Southern China. Discret. Dyn. Nat. Soc. 2015, 2015, 172919. [Google Scholar] [CrossRef]
- Tubiello, F.N.; Fischer, G.I. Reducing climate change impacts on agriculture: Global and regional effects of mitigation, 2000–2080. Technol. Forecast. Soc. 2007, 74, 1030–1056. [Google Scholar] [CrossRef]
- Engelbrecht, B.M.J.; Comita, L.S.; Condit, R.; Kursar, T.A.; Tyree, M.T.; Turner, B.L.; Hubbell, S.P. Drought sensitivity shapes species distribution patterns in tropical forests. Nature 2007, 447, U80–U82. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Biswas, D.K.; Li, W.D.; Chen, S.B.; Zhang, L.; Jiang, G.M.; Li, Y.G. Photosynthesis and yield responses of ozone-polluted winter wheat to drought. Photosynthetica 2007, 45, 582–588. [Google Scholar] [CrossRef]
- Ashraf, M.; Mehmood, S. Response of four Brassica species to drought stress. Environ. Exp. Bot. 1990, 30, 93–100. [Google Scholar] [CrossRef]
- Plonowanie Odmian Rzepaku Jarego w 2019 Roku. Available online: https://www.agropolska.pl/uprawa/rosliny-oleiste/plonowanie-odmian-rzepaku-jarego-w-2019-roku,205.html (accessed on 10 January 2021).
- Światowe Zbiory Rzepaku w Sezonie 2019/20. Available online: https://www.agrofakt.pl/swiatowe-zbiory-rzepaku-w-sezonie-2019-20/ (accessed on 10 January 2021).
- Kowalska, J.; Tyburski, J.; Bocianowski, J.; Krzymińska, J.; Matysiak, K. Methods of Silicon Application on Organic Spring Wheat (Triticum aestivum L. spp. vulgare) Cultivars Grown across Two Contrasting Precipitation Years. Agronomy 2020, 10, 1655. [Google Scholar] [CrossRef]
- Liu, H.X.; Guo, Z.G. Forage yield and water use efficiency of alfalfa applied with silicon under water deficit conditions. Philipp. Agric. Sci. 2013, 96, 370–376. [Google Scholar]
- Shi, Y.; Zhang, Y.; Han, W.H.; Feng, R.; Hu, Y.H.; Guo, J.; Gong, H.J. Silicon enhances water stress tolerance by improving root hydraulic conductance in Solanum lycopersicum L. Front. Plant Sci. 2016, 7, 196. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, S.; Chauhan, D.K.; Dubey, N.K.; Prasad, R. Silicon as A Beneficial Element to Combat the Adverse Effect of Drought in Agricultural Crops: Capabilities and Future Possibilities. In Water Stress and Crop Plants: A Sustainable Approach; Parvaiz, A., Ed.; John Wiley & Sons, Ltd.: Pondicherry, India, 2016; Volume 2, pp. 682–694. [Google Scholar]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon nanoparticles more efficiently alleviate arsenate toxicity than silicon in maize cultiver and hybrid differing in arsenate tolerance. Front. Environ. Sci. 2016, 4, 46. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K.; Rai, A.K. LIB spectroscopic and biochemical analysis to characterize lead toxicity alleviative nature of silicon in wheat (Triticum aestivum L.) seedlings. J. Photoch. Photobio. B 2016, 154, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.X.; Gong, H.J. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.-S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.M.; Ramezani, A.; Far, S.M.; Sadat, K.A.; Moradi-Ghahderijani, M.; Jamian, S.S. Application of silicon ameliorates salinity stress in sunflower (Helianthus annuus L.) plants. Int. J. Agric. Crop Sci. 2013, 6, 1367–1372. [Google Scholar]
- Noman, A.; Ali, S.; Naheed, F.; Ali, Q.; Farid, M.; Rizwan, M.; Irshad, M.K. Foliar application of ascorbate enhances the physiological and biochemical attributes of maize (Zea mays L.) cultivars under drought stress. Arch. Agron. Soil Sci. 2015, 61, 1659–1672. [Google Scholar] [CrossRef]
- Kurdali, F.; Al-Chammaa, M. Growth and nitrogen fixation in silicon and/or potassium fed chickpeas grown under drought and well watered conditions. J. Stress Physiol. Biochem. 2013, 9, 385–406. [Google Scholar]
- Ming, D.F.; Pei, Z.F.; Naeem, M.S.; Gong, H.J.; Zhou, W.J. Silicon alleviates PEG-induced water-deficit stress in upland rice seedlings by enhancing osmotic adjustment. J. Agron. Crop. Sci. 2012, 198, 14–26. [Google Scholar] [CrossRef]
- Saud, S.; Li, X.; Chen, Y.; Zhang, L.; Fahad, S.; Hussain, S.; Sadiq, A.; Chen, Y. Silicon application increases drought tolerance of Kentucky Bluegrass by improving plant water relations and morphophysiological functions. Sci. World J. 2014. [Google Scholar] [CrossRef]
- Amin, M.; Ahmad, R.; Basra, S.M.A.; Murtaza, G. Silicon induced improvement in morpho-physiological traits of maize (Zea mays L.) under water deficit. Pak. J. Agric. Sci. 2014, 51, 187–196. [Google Scholar]
- Sonobe, K.; Hattori, T.; An, P.; Tsuji, W.; Eneji, A.E.; Kobayashi, S.; Kawamura, Y.; Tanaka, K.; Inanaga, S. Effect of silicon application on sorghum root responses to water stress. J. Plant Nutr. 2010, 34, 71–82. [Google Scholar] [CrossRef]
- Ahmed, M.; Fayyaz Ul, H.; Asif, M. Amelioration of Drought in Sorghum (Sorghum bicolor L.) by Silicon. Commun. Soil Sci. Plan. 2014, 45, 470–486. [Google Scholar] [CrossRef]
- Ahmed, M.; Asif, M.; Hassan, F.-U. Augmenting drought tolerance in sorghum by silicon nutrition. Acta. Physiol. Plant. 2014, 36, 473–483. [Google Scholar] [CrossRef]
- Keller, C.; Rizwan, M.; Davidian, J.C.; Pokrovsky, O.S.; Bovet, N.; Chaurand, P.; Meunier, J.D. Effect of silicon on wheat seedlings (Triticum turgidum L.) grown in hydroponics and exposed to 0 to 30 μMCu. Planta 2015, 241, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Haynes, R.J. A contemporary overview of silicon availability in agricultural soils. J. Plant Nutr. Soil Soc. 2014, 177, 831–844. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Huynh, W.Q.; Kronzucker, H.J. The role of silicon in higher plants under salinity and drought stress. Front. Plant Sci. 2016, 7, 1072. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Belanger, R.; Gong, H.; Song, A. Silicon in Agriculture: From Theory to Practice; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2015; pp. 978–994. [Google Scholar]
- Epstein, E.; Bloom, A.J. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates, Inc.: Sunderland, UK, 2005. [Google Scholar]
- Artyszak, A. Effect of Silicon Fertilization on Crop Yield Quantity and Quality—A Literature Review in Europe. Plants 2018, 7, 54. [Google Scholar] [CrossRef]
- Kowalska, J.; Tyburski, J.; Jakubowska, M.; Krzymińska, J. Effect of Different Forms of Silicon on Growth of Spring Wheat Cultivated in Organic Farming System. Silicon 2021, 13, 211–217. [Google Scholar] [CrossRef]
- Guntzer, F.; Keller, C.; Meunier, J.D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Khan, M.I.R.; Fujita, M. Silicon-mediated regulation of antioxidant defense and glyoxalase systems confers drought stress tolerance in Brassica napus L. S. Afr. J. Bot. 2018, 115, 50–57. [Google Scholar] [CrossRef]
- Becker, M.; Ngo, N.S.; Schenk, M.K.A. Silicon reduces the iron uptake in rice and induces iron homeostasis related genes. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Gonzalo, M.J.; Lucena, J.J.; Hernandez-Apaolaza, L. Effect of silicon addition on soybean (Glycine max) and cucumber (Cucumis sativus) plants grown under iron deficiency. Plant Physiol. Bioch. 2013, 70, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Functions of Mineral Nutrients: Micronutrients, Iron. In Mineral Nutrition of Higher Plants; Academic Press Limited: London, UK; San Diego, CA, USA; New York, NY, USA; Boston, MA, USA; Sydney, Australia; Tokyo, Japan; Toronto, ON, Canada, 1995; pp. 313–324. [Google Scholar]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. R. 2015, 22, 15416–15431. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Q.; Wang, S.W.; Yin, L.; Deng, X.P. How does silicon mediate plant water uptake and loss under water deficiency? Front. Plant Sci. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Sonah, H.; Deshmukh, R.K.; Labbé, C.; Bélanger, R.R. Analysis of aquaporins in Brassicaceae species reveals high-level of conservation and dynamic role against biotic and abiotic stress in canola. Sci. Rep. 2017, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.M. Osmoregulation and water stress in higher plants. Annu. Rev. Plant Phys. 1984, 35, 299–319. [Google Scholar] [CrossRef]
- Agarie, S.; Uchida, H.; Agata, W.; Kubota, F.; Kaufman, P.B. Effects of silicon on transpiration and leaf conductance in rice plants (Oryza sativa L.). Plant. Prod. Sci. 1998, 1, 89–95. [Google Scholar] [CrossRef]
- Gao, D.; Cai, K.Z.; Chen, J.N.; Luo, S.M.; Zeng, R.S.; Yang, J.Y.; Zhu, X.Y. Silicon enhances photochemical efficiency and adjusts mineral nutrient absorption in Magnaporthe oryzae infected rice plants. Acta. Physiol. Plant. 2011, 33, 675–682. [Google Scholar] [CrossRef]
- Habibi, G. Silicon supplementation improves drought tolerance in canola plants. Russ. J. Plant Physl. 2014, 61, 784–791. [Google Scholar] [CrossRef]
- Meier, U. Growth stages of mono- and dicotyledonous plants. BBCH Monogr. 2001. [Google Scholar] [CrossRef]
- Masarovic, D.; Slovakova, L.; Bokor, B.; Bujdos, M.; Lux, A. Effect of silicon application on Sorghum bicolor exposed to toxic concentration of zinc. Biologia 2012, 67, 706–712. [Google Scholar] [CrossRef]
- Quarrie, S.A.; Whitford, P.N.; Appleford, N.E.J.; Wang, T.L.; Cook, S.K.; Henson, I.E.; Loveys, B.R. A monoclonal antibody to (S) abscisic acid: Its characterisation and use in a radioimmunoassay for measuring abscisic acid in crude extracts of cereal and lupin leaves. Planta 1988, 173, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Walker-Simmons, M.; Abrams, S. Use of ABA Immunoassays. In Abscisic Acid, Physiology and Biochemistry; Davies, W., Jones, H., Eds.; Bios Scientific Publishers: Oxford, UK, 1991; pp. 53–63. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Conditions | Treatment | Shoot (Leaves and Stem) Si (mg/kg) | Leaf Blade Si (mg/kg) | Leaf Petiole Si (mg/kg) |
|---|---|---|---|---|
| Optimal conditions | control | 729.8 c | 1487.1 b | 1182.4 b |
| Si + Fe | 2159.25 a | 1804.3 a | 2351.4 a | |
| Si | 1663.35 b | 953.1 c | 1258.2 b |
| Points of Measurement | Treatment | Leaf Water Content (%) | Leaf Osmotic Potential Ψo (Mpa) |
|---|---|---|---|
| One week after first watering | control | 90.09 a | −0.74 a |
| Si + Fe | 90.10 a | −0.82 c | |
| Si | 89.33 a,b | −0.77 a,b | |
| One week after second watering | control | 89.47 a | −0.82 c |
| Si + Fe | 87.87 b,c | −0.83 c | |
| Si | 87.94 b,c | −0.80 b,c | |
| One week after third watering | control | 89.12 a,b | −0.75 a |
| Si + Fe | 86.56 c | −0.78 b,c | |
| Si | 87.62 c | −0.75 a |
| Growth Conditions | Treatment | Shoot (Leaves and Stem) Si (mg/kg) | Leaf Blade Si (mg/kg) | Leaf Petiole Si (mg/kg) |
|---|---|---|---|---|
| Drought | control | 889.1 c | 1111.5 b | 1305.5 c |
| Si + Fe | 2115.1 a | 973.8 b | 1731.3 b | |
| Si | 1422.4 b | 1548.7 a | 3241.5 a |
| Growth Conditions | Treatment | Leaf Osmotic Potential Ψo (MPa) | Leaf Water Potential Ψw (MPa) |
|---|---|---|---|
| Optimal conditions | control | −1.13 a,b | −0.86 a |
| Si + Fe | −1.08 a | −0.87 a | |
| Si | −1.20 b | −0.99 a | |
| Drought conditions | control | −1.76 d | −1.35 b |
| Si + Fe | −1.48 c | −1.30 b | |
| Si | −1.68 d | −1.46 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saja-Garbarz, D.; Ostrowska, A.; Kaczanowska, K.; Janowiak, F. Accumulation of Silicon and Changes in Water Balance under Drought Stress in Brassica napus var. napus L. Plants 2021, 10, 280. https://doi.org/10.3390/plants10020280
Saja-Garbarz D, Ostrowska A, Kaczanowska K, Janowiak F. Accumulation of Silicon and Changes in Water Balance under Drought Stress in Brassica napus var. napus L. Plants. 2021; 10(2):280. https://doi.org/10.3390/plants10020280
Chicago/Turabian StyleSaja-Garbarz, Diana, Agnieszka Ostrowska, Katarzyna Kaczanowska, and Franciszek Janowiak. 2021. "Accumulation of Silicon and Changes in Water Balance under Drought Stress in Brassica napus var. napus L." Plants 10, no. 2: 280. https://doi.org/10.3390/plants10020280
APA StyleSaja-Garbarz, D., Ostrowska, A., Kaczanowska, K., & Janowiak, F. (2021). Accumulation of Silicon and Changes in Water Balance under Drought Stress in Brassica napus var. napus L. Plants, 10(2), 280. https://doi.org/10.3390/plants10020280