
Evaluation of Cadmium Bioaccumulation-Related Physiological Effects in Salvinia biloba: An Insight towards Its Use as Pollutant Bioindicator in Water Reservoirs

 ,
,  and
and 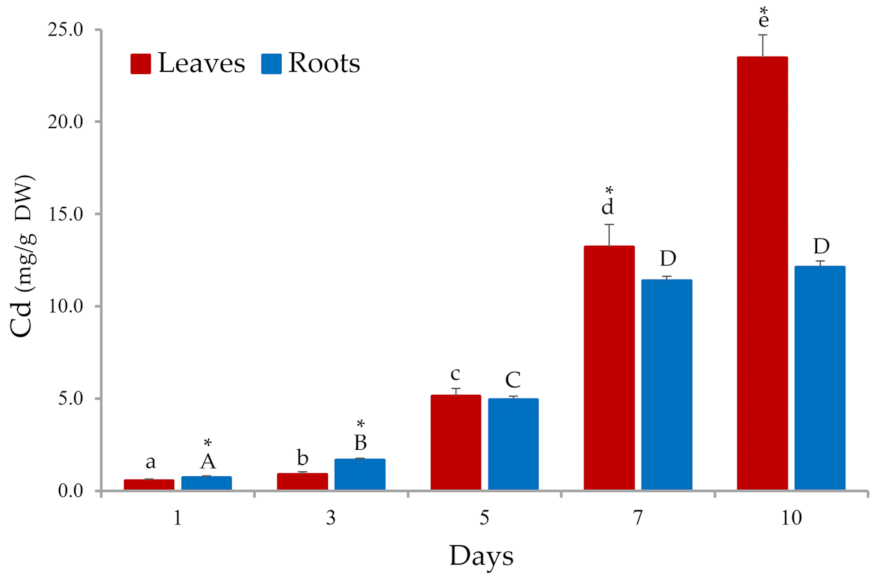{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Collection and Characterization
2.2. Cd Bioaccumulation Studies
2.3. Physiological Parameters
2.3.1. Quantification of Photosynthetic Pigments
2.3.2. Flavonoid Determination
2.3.3. Soluble Carbohydrates
2.4. Statistical Analysis
3. Results and Discussion
3.1. Cd Bioaccumulation
3.2. Phenotypic Evaluation of Cd Phytotoxicity in S. biloba
3.3. Cd Phytotoxicity in S. biloba
3.3.1. Photosynthetic Pigments
3.3.2. Flavonoid Content
3.3.3. Soluble Carbohydrates
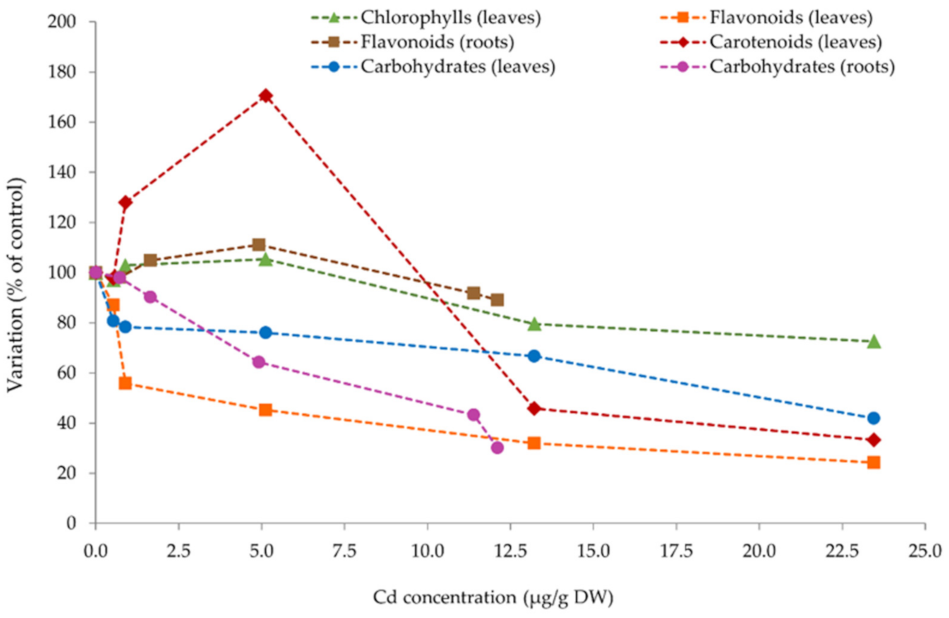
3.4. Further Analysis—Cd Bioaccumulation vs. Physiological Parameters
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lesiv, M.S.; Polishchuk, A.I.; Antonyak, H.L. Aquatic macrophytes: Ecological features and functions. Biol. Stud. 2020, 14, 79–94. [Google Scholar] [CrossRef]
- Rejmánková, E. The Role of Macrophytes in Wetland Ecosystems. J. Ecol. Field Biol. 2011, 34, 333–345. [Google Scholar] [CrossRef]
- Bonanno, G.; Borg, J.A.; Di Martino, V. Levels of Heavy Metals in Wetland and Marine Vascular Plants and Their Biomonitoring Potential: A Comparative Assessment. Sci. Total Environ. 2017, 576, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Alonso, X.; Hadad, H.R.; Córdoba, C.; Polla, W.; Reyes, M.S.; Fernández, V.; Granados, I.; Marino, L.; Villalba, A. Macrophytes as Potential Biomonitors in Peri-Urban Wetlands of the Middle Parana River (Argentina). Environ. Sci. Pollut. Res. 2018, 25, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Fawzy, M.A.; Badr, N.; El-Khatib, A.; Abo-El-Kassem, A. Heavy Metal Biomonitoring and Phytoremediation Potentialities of Aquatic Macrophytes in River Nile. Environ. Monit. Assess. 2012, 184, 1753–1771. [Google Scholar] [CrossRef] [PubMed]
- Freitas, F.; Battirola, L.D.; de Andrade, R.L.T. Adsorption of Cu2+ and Pb2+ Ions by Pontederia rotundifolia (L.f.) (Pontederiaceae) and Salvinia biloba Raddi (Salviniaceae) biomass. Water Air Soil Pollut. 2018, 229, 349. [Google Scholar] [CrossRef]
- Dixit, R.; Wasiullah, E.; Malaviya, D.; Pandiyan, K.; Singh, U.B.; Sahu, A.; Shukla, R.; Singh, B.P.; Rai, J.P.; Sharma, P.K.; et al. Bioremediation of Heavy Metals from Soil and Aquatic Environment: An Overview of Principles and Criteria of Fundamental Processes. Sustainability 2015, 7, 2189–2212. [Google Scholar] [CrossRef]
- Sharma, P.; Pandey, S. Status of Phytoremediation in World Scenario. Int. J. Environ. Bioremediat. Biodegrad. 2014, 2, 178–191. [Google Scholar] [CrossRef]
- Dhir, B. Potential of Biological Materials for Removing Heavy Metals from Wastewater. Environ. Sci. Pollut. Res. Int. 2014, 21, 1614–1627. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of Heavy Metals—Concepts and Applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Rai, P.K. Heavy Metal Phytoremediation from Aquatic Ecosystems with Special Reference to Macrophytes. Crit. Rev. Environ. Sci. Technol. 2009, 39, 697–753. [Google Scholar] [CrossRef]
- Emiliani, J.; Llatance Oyarce, W.G.; Bergara, C.D.; Salvatierra, L.M.; Novo, L.A.B.; Pérez, L.M. Variations in the Phytoremediation Efficiency of Metal-Polluted Water with Salvinia biloba: Prospects and Toxicological Impacts. Water 2020, 12, 1737. [Google Scholar] [CrossRef]
- Castillo Loría, K.; Emiliani, J.; Herrero, M.S.; Bergara, C.D.; Salvatierra, L.M.; Pérez, L.M. Effect of Daily Exposure to Pb-Contaminated Water into Salvinia biloba Physiology and Phytoremediation Performance. Aquat. Toxicol. 2019, 210, 158–166. [Google Scholar] [CrossRef]
- Prado, C.; Ponce, S.C.; Pagano, E.; Prado, F.E.; Rosa, M. Differential Physiological Responses of Two Salvinia Species to Hexavalent Chromium at a Glance. Aquat. Toxicol. 2016, 175, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Bizzo, A.L.; Intorne, A.C.; Gomes, P.H.; Susuki, M.S.; Esteves, B.D. Short-Term Physiological Responses to Copper Stress in Salvinia auriculata Aubl. Acta Limnol. Bras. 2014, 26, 268–277. [Google Scholar] [CrossRef][Green Version]
- Dhir, B.; Sharmila, P.; Pardha-Saradhi, P.; Sharma, P.; Kumar, R.; Mehta, D. Heavy Metal Induced Physiological Alterations in Salvinia natans. Ecotoxicol. Environ. Saf. 2011, 74, 1678–1684. [Google Scholar] [CrossRef]
- Wolff, G.; Pereira, G.C.; Castro, E.M.; Louzada, J.; Coelho, F.F. The Use of Salvinia auriculata as a Bioindicator in Aquatic Ecosystems: Biomass and Structure Dependent on the Cadmium Concentration. Braz. J. Biol. 2012, 72, 71–77. [Google Scholar] [CrossRef]
- Freitas, F.; Lunardi, S.; Souza, L.B.; von der Osten, J.S.C.; Arruda, R.; Andrade, R.L.T.; Battirola, L.D. Accumulation of Copper by the Aquatic Macrophyte Salvinia biloba Raddi (Salviniaceae). Braz. J. Biol. 2018, 78, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Tello Zevallos, W.; Salvatierra, L.M.; Loureiro, D.B.; Morató, J.; Pérez, L.M. Evaluation of the Autochthonous Free-Floating Macrophyte Salvinia biloba Raddi for Use in the Phytoremediation of Water Contaminated. Desalin. Water Treat. 2018, 103, 282–289. [Google Scholar] [CrossRef]
- Ferreira, F.A.; Mormul, R.P.; Thomaz, S.M.; Pott, A.; Pott, V.J. Macrophytes in the Upper Paraná River floodplain: Checklist and Comparison with Other Large South American Wetlands. Rev. Biol. Trop. 2011, 59, 541–556. [Google Scholar] [PubMed]
- Guerrero, E.L.; Agnolin, F.L.; Benedictto, M.; Gambeta, D.; Suazo Lara, F.A.; Derguy, M.R.; Apodaca, M.J. Vascular Plant Species of the Floating Vegetation Rafts from the Río de la Plata (Argentina). Rodriguésia 2018, 69, 1965–1972. [Google Scholar] [CrossRef]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef] [PubMed]
- Rahimzadeh, M.R.; Kazemi, S.; Moghadamnia, A.A. Cadmium Toxicity and Treatment: An Update. Casp. J. Intern. Med. 2017, 8, 135–145. [Google Scholar] [CrossRef]
- Qu, R.J.; Wang, X.H.; Feng, M.B.; Li, Y.; Liu, H.X.; Wang, L.S.; Wang, Z.Y. The Toxicity of Cadmium to Three Aquatic Organisms (Photobacterium phosphoreum, Daphnia magna and Carassius auratus) under Different pH Levels. Ecotoxicol. Environ. Saf. 2013, 95, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Zaki, M.S.; Zakaria, A.; Eissa, I.A.; Eldeen, A.I. Effect of Cadmium Toxicity on Vertebrates. Electron. Physician 2016, 8, 1964–1965. [Google Scholar] [CrossRef]
- Hayat, M.T.; Nauman, M.; Nazir, N.; Ali, S.; Bangash, N. Environmental Hazards of Cadmium: Past, Present, and future. In Cadmium Toxicity and Tolerance in Plants: Agronomic, Genetic, Molecular and Omic Approaches, 1st ed.; Hasanuzzaman, M., Prasad, M.N.V., Fujita, M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 163–183. [Google Scholar] [CrossRef]
- WHO. Chemical Safety—Activity Report; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of Total Carotenoids and Chlorophylls A and B of Leaf Extracts in Different Solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Robyt, I.F.; White, J.B. Biochemical Techniques: Theory and Practice, 1st ed.; Brooks and Cole Publishing: Monterey, CA, USA, 1987; 407p. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Sánchez-Galván, G.; Monroy, O.; Gómez, J.; Olguín, E.J. Assessment of the Hyperaccumulating Lead Capacity of Salvinia minima Using Bioadsorption and Intracellular Accumulation Factors. Water Air Soil Pollut. 2008, 194, 77–90. [Google Scholar] [CrossRef]
- Olguín, E.J.; Sánchez-Galván, G.; Pérez-Pérez, T.; Pérez-Orozco, A. Surface Adsorption, Intracellular Accumulation and Compartmentalization of Lead in Batch-Operated Lagoons with Salvinia minima as Affected by Environmental Conditions, EDTA and Nutrients. J. Ind. Microbiol. Biotechnol. 2005, 32, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Estrella-Gómez, N.; Mendoza-Cózatl, D.; Romero-Sánchez, R.; Gonzáles-Mendoza, D.; Zapata-Pérez, O.; Martínez-Hernández, A.; Santamaría, J.M. The Pb-Hyperaccumulator Aquatic Fern Salvinia minima Baker, Responds to Pb2+ by Increasing Phytochelatins via Changes in SmPCS Expression and in Phytochelatin Synthase Activity. Aquat. Toxicol. 2009, 91, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Schor-Fumbarov, T.; Goldsbrough, P.; Adam, Z.; Tel-Or, E. Characterization and Expression of a Metallothionein Gene in the Aquatic Fern Azolla filiculoides Under Heavy Metal Stress. Planta 2005, 223, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Cobbett, C.S. Phytochelatins and Their Roles in Heavy Metal Detoxification. Plant Physiol. 2000, 123, 825–832. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.; Rodríguez-Zavala, J.; Rodríguez-Enríquez, S.; Mendoza-Hernandez, G.; Briones-Gallardo, G.; Moreno-Sánchez, R. Phytochelatin—Cadmium—Sulfide High-Molecular-Mass Complexes of Euglena gracilis. FEBS J. 2006, 273, 5703–5713. [Google Scholar] [CrossRef] [PubMed]
- Phetsombat, S.; Kruatrachue, M.; Pokethitiyook, P.; Upatham, S. Toxicity and Bioaccumulation of Cadmium and Lead in Salvinia cucullata. J. Environ Biol. 2006, 27, 645–652. [Google Scholar]
- Tello Zevallos, W.; Loureiro, D.B.; Reeves, M.C.; Yujnovsky, F.; Salvatierra, L.M.; Pérez, L.M. Biorremediación de Aguas Contaminadas con Metales Pesados Empleando Macrófitas Autóctonas de Flotación Libre. In IV Simposio Argentino de Procesos Biotecnológicos: Libro de Resúmenes, Proceeding of the 4th Simposio Argentino de Procesos Biotecnológicos (SAPROBIO 2016), Buenos Aires, Argentina, 1–2 December 2016; Carbajal, M.L., Rojas, N.L., Grasselli, M., Eds.; (Universidad Nacional de Quilmes), Bernal: Buenos Aires, Argentina, 2017; Abstract Number: S4.22; pp. 242–243. ISBN 978-987-558-428-0. [Google Scholar]
- Williams, L.E.; Pittman, J.K.; Hall, J.L. Emerging Mechanisms for Heavy Metal Transport in Plants. Biochim. Biophys. Acta 2000, 1465, 104–126. [Google Scholar] [CrossRef]
- Gomes, S.M.S.; De Lima, V.L.A.; De Souza, A.P.; Do Nascimento, J.J.V.R.; Do Nascimento, E.S. Chloroplast Pigments as Indicators of Lead Stress. Eng. Agríc. 2014, 34, 877–884. [Google Scholar] [CrossRef]
- Babenko, L.M.; Shcherbatiuk, M.M.; Polishchuk, O.V.; Kosakivska, I.V. Structural-Functional Peculiarities of Water Fern Salvinia natans (L.) All. Biol. Stud. 2016, 10, 149–162. [Google Scholar] [CrossRef]
- Kosakivska, I.V.; Shcherbatiuk, M.M.; Babenko, L.M.; Polishchuk, O.V. Characteristics of Photosynthetic Apparatus of Aquatic Fern Salvinia natans Floating and Submerged Fronds. Adv. Biol. Earth Sci. 2018, 3, 13–26. [Google Scholar]
- Fryzova, R.; Pohanka, M.; Martinkova, P.; Cihlarova, H.; Brtnicky, M.; Hladky, J.; Kynicky, J. Oxidative Stress and Heavy Metals in Plants. Rev. Environ. Contam. Toxicol. 2018, 245, 129–156. [Google Scholar] [CrossRef]
- Carralero Bon, I.; Salvatierra, L.M.; Lario, L.D.; Morató, J.; Pérez, L.M. Prospects in Cadmium-Contaminated Water Management Using Free-Living Cyanobacteria (Oscillatoria sp.). Water 2021, 13, 542. [Google Scholar] [CrossRef]
- Ramel, F.; Mialoundama, A.S.; Havaux, M. Nomenzymic Carotenoid Oxidation and Photooxidative Stress Signalling in Plants. J. Exp. Bot. 2013, 64, 799–805. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Queval, G.; Foyer, C.H. The Impact of Global Change Factors on Redox Signaling Underpinning Stress Tolerance. Plant Physiol. 2013, 161, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Caretto, S.; Linsalata, V.; Colella, G.; Lattanzio, V. Carbon Fluxes Between Primary Metabolism and Phenolic Pathway in Plant Tissues Under Stress. Int. J. Mol. Sci. 2015, 16, 26378–26394. [Google Scholar] [CrossRef] [PubMed]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-Induced Flavonoid Biosynthesis and the Antioxidant Machinery of Plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef]
- Estrella-Gómez, N.E.; Sauri-Duch, E.; Zapata-Pérez, O.; Santamaría, J.M. Glutathione Plays a Role in Protecting Leaves of Salvinia minima from Pb2+ Damage Associated with Changes in the Expression of SmGS Genes an Increased Activity of GS. Environ. Exp. Bot. 2012, 75, 188–194. [Google Scholar] [CrossRef]
- Leal-Alvarado, D.A.; Estrella-Maldonado, H.; Sáenz-Carbonell, L.; Ramírez-Prado, J.H.; Zapata-Pérez, O.; Santamaría, J.M. Genes Coding for Transporters Showed a Rapid and Sharp Increase in Their Expression in Response to Lead, in the Aquatic Fern (Salvinia minima Baker). Ecotoxicol. Environ. Saf. 2018, 147, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, A.; Prado, F. Soluble Sugars—Metabolism, Sensing and Abiotic Stress. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emiliani, J.; Oyarce, W.G.L.; Salvatierra, L.M.; Novo, L.A.B.; Pérez, L.M. Evaluation of Cadmium Bioaccumulation-Related Physiological Effects in Salvinia biloba: An Insight towards Its Use as Pollutant Bioindicator in Water Reservoirs. Plants 2021, 10, 2679. https://doi.org/10.3390/plants10122679
Emiliani J, Oyarce WGL, Salvatierra LM, Novo LAB, Pérez LM. Evaluation of Cadmium Bioaccumulation-Related Physiological Effects in Salvinia biloba: An Insight towards Its Use as Pollutant Bioindicator in Water Reservoirs. Plants. 2021; 10(12):2679. https://doi.org/10.3390/plants10122679
Chicago/Turabian StyleEmiliani, Julia, Wendi G. Llatance Oyarce, Lucas M. Salvatierra, Luís A. B. Novo, and Leonardo M. Pérez. 2021. "Evaluation of Cadmium Bioaccumulation-Related Physiological Effects in Salvinia biloba: An Insight towards Its Use as Pollutant Bioindicator in Water Reservoirs" Plants 10, no. 12: 2679. https://doi.org/10.3390/plants10122679
APA StyleEmiliani, J., Oyarce, W. G. L., Salvatierra, L. M., Novo, L. A. B., & Pérez, L. M. (2021). Evaluation of Cadmium Bioaccumulation-Related Physiological Effects in Salvinia biloba: An Insight towards Its Use as Pollutant Bioindicator in Water Reservoirs. Plants, 10(12), 2679. https://doi.org/10.3390/plants10122679