
Cyanidiophyceae (Rhodophyta) Tolerance to Precious Metals: Metabolic Response to Palladium and Gold

 ,
,  , ,
, ,  ,
, 
{kind=link}
{kind=link}
Abstract
:1. Introduction
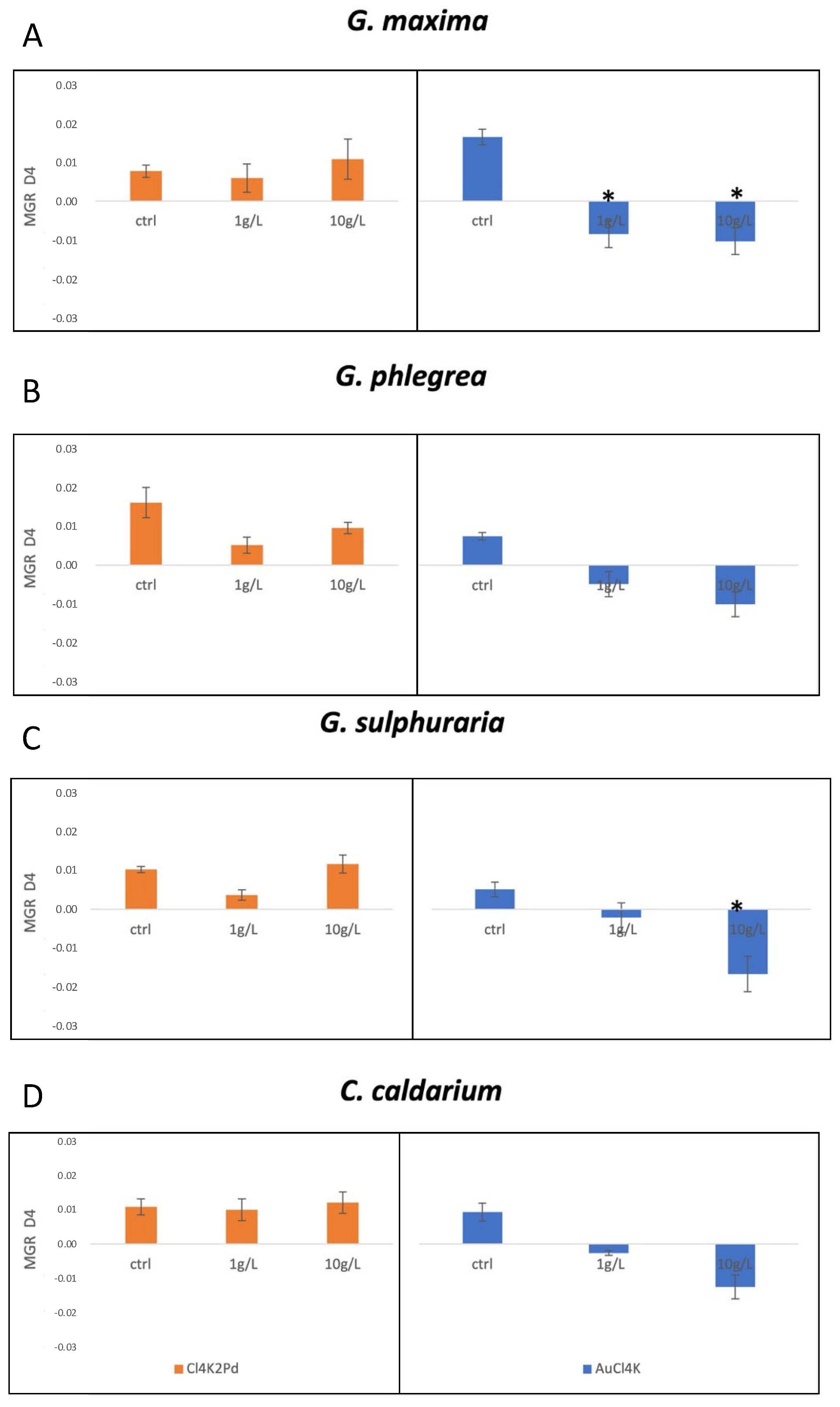
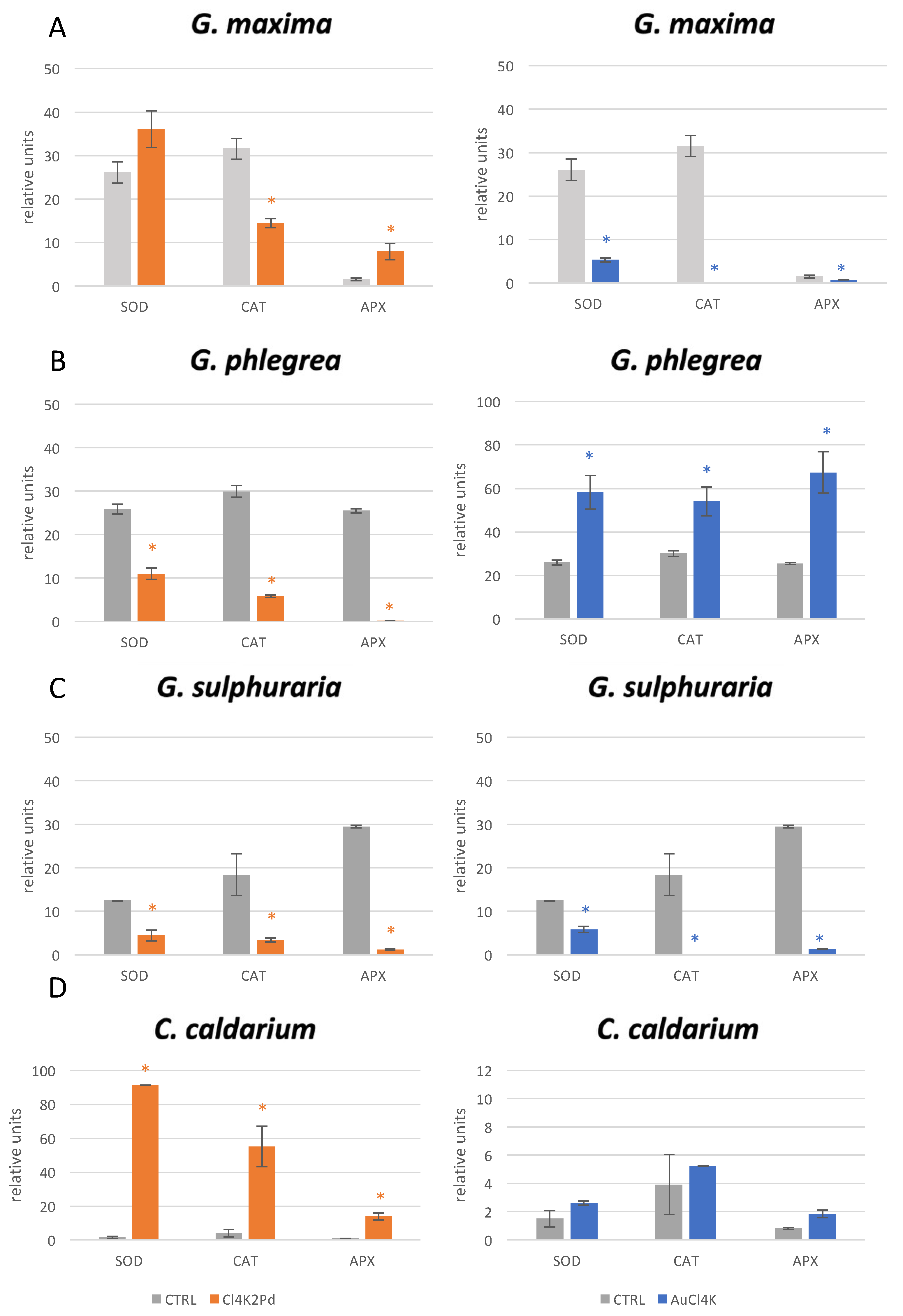
2. Results
3. Discussion
4. Materials and Methods
4.1. Strain Cultivations
4.2. Experimental Procedure
4.3. Enzyme Extraction and Assays
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Cicco, M.R.; Spagnuolo, A.; Masiello, A.; Vetromile, C.; Nappa, M.; Lubritto, C. Energetic and environmental analysis of a wastewater treatment plant through static and dynamic monitoring activities. Int. J. Environ. Sci. Technol. 2020, 17, 4299–4312. [Google Scholar] [CrossRef]
- Güzel, Y.; Rainer, M.; Mirza, M.R.; Messner, C.B.; Bonn, G.K. Highly selective recovery of phosphopeptides using trypsin-assisted digestion of precipitated lanthanide–phosphoprotein complexes. Analyst 2013, 138, 2897–2905. [Google Scholar] [CrossRef]
- Hosomomi, Y.; Baba, Y.; Kubota, F.; Kamiya, N.; Goto, M. Biosorption of Rare Earth Elements by E. coli. J. Chem. Eng. Jpn. 2013, 46, 13we031. [Google Scholar] [CrossRef]
- Jiang, M.; Ohnuki, T.; Kozai, N.; Tanaka, K.; Suzuki, Y.; Sakamoto, F.; Kamiishi, E.; Utsunomiya, S. Biological nano-mineralization of Ce phosphate by Saccharomyces cerevisiae. Chem. Geol. 2010, 277, 61–69. [Google Scholar] [CrossRef]
- Ju, X.; Igarashi, K.; Miyashita, S.; Mitsuhashi, H.; Inagaki, K.; Fujii, S.; Sawada, H.; Kuwabara, T.; Minoda, A. Effective and selective recovery of gold and palladium ions from metal wastewater using a sulfothermophilic red alga, Galdieria sulphuraria. Bioresour. Technol. 2016, 211, 759–764. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, K.; Ueda, M. Engineering of microorganisms towards recovery of rare metal ions. Appl. Microbiol. Biotechnol. 2010, 87, 53–60. [Google Scholar] [CrossRef]
- Di Cicco, M.R.; Spagnuolo, A.; Masiello, A.; Vetromile, C.; Lubritto, C.; Nappa, M.; Corbo, G. Energy monitoring of a wastewater treatment plant in salerno, campania region (southern italy). In Advances in Science, Technology and Innovation; Springer: Cham, Switzerland, 2019; pp. 107–111. [Google Scholar]
- Vetromile, C.; Spagnuolo, A.; Petraglia, A.; Masiello, A.; di Cicco, M.R.; Lubritto, C. Pre- and post-operam comparison of the energy consumption of a radio base station under energy efficiency actions. Energy Build. 2021, 236, 110772. [Google Scholar] [CrossRef]
- Arshadi, M.; Mousavi, S.M.; Rasoulnia, P. Enhancement of simultaneous gold and copper recovery from discarded mobile phone PCBs using Bacillus megaterium: RSM based optimization of effective factors and evaluation of their interactions. Waste Manag. 2016, 57, 158–167. [Google Scholar] [CrossRef]
- Latorre, M.; Cortés, M.P.; Travisany, D.; Di Genova, A.; Budinich, M.; Reyes-Jara, A.; Hödar, C.; González, M.; Parada, P.; Bobadilla-Fazzini, R.A.; et al. The bioleaching potential of a bacterial consortium. Bioresour. Technol. 2016, 218, 659–666. [Google Scholar] [CrossRef]
- Wang, M.; Tan, Q.; Chiang, J.F.; Li, J. Recovery of rare and precious metals from urban mines—A review. Front. Environ. Sci. Eng. 2017, 11, 1–17. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, H.; Liu, Y.; Bian, L.; Wang, X.; Zhou, Z.; Huang, Y. Bioleaching of rare earth elements from bastnaesite-bearing rock by actinobacteria. Chem. Geol. 2018, 483, 544–557. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.J.; Mimura, T. Vacuolar compartmentalization as indispensable component of heavy metal detoxification in plants. Plant Cell Environ. 2016, 39, 1112–1126. [Google Scholar] [CrossRef]
- Di Cicco, M.R.; Palmieri, M.; Altieri, S.; Ciniglia, C.; Lubritto, C. Cultivation of the Acidophilic Microalgae Galdieria phlegrea with Wastewater: Process Yields. Int. J. Environ. Res. Public Health 2021, 18, 2291. [Google Scholar] [CrossRef]
- Matsunaga, T.; Takeyama, H.; Nakao, T.; Yamazawa, A. Screening of marine microalgae for bioremediation of cadmium-polluted seawater. J. Biotechnol. 1999, 70, 33–38. [Google Scholar] [CrossRef]
- Mehta, S.K.; Tripathi, B.N.; Gaur, J.P. Enhanced sorption of Cu2+ and Ni2+ by acid-pretreated Chlorella vulgaris from single and binary metal solutions. J. Appl. Phycol. 2002, 14, 267–273. [Google Scholar] [CrossRef]
- Wang, J.; Chen, C. Biosorbents for heavy metals removal and their future. Biotechnol. Adv. 2009, 27, 195–226. [Google Scholar] [CrossRef]
- Bourcier, W.L.; Lin, M.; Nix, G. Recovery of Minerals and Metals from Geothermal Fluids. In Proceedings of the 2003 SME Annual Meeting, Cincinnati, OH, USA, 24–26 February 2003. [Google Scholar]
- Doemel, W.N.; Brock, T.D. The Physiological ecology of Cyanidium caldarium. J. Gen. Microbiol. 1971, 67, 17–32. [Google Scholar] [CrossRef] [Green Version]
- De Luca, P.; Taddei, R.; Varano, L. “Cyanidioschyzon merolae”: A new alga of thermal acidic environments. Webbia 1978, 33, 37–44. [Google Scholar] [CrossRef]
- Albertano, P.; Ciniglia, C.; Pinto, G.; Pollio, A. The taxonomic position of Cyanidium, Cyanidioschyzon and Galdieria: An update. Hydrobiologia 2000, 433, 137–143. [Google Scholar] [CrossRef]
- Minoda, A.; Sawada, H.; Suzuki, S.; Miyashita, S.; Inagaki, K.; Yamamoto, T.; Tsuzuki, M. Recovery of rare earth elements from the sulfothermophilic red alga Galdieria sulphuraria using aqueous acid. Appl. Microbiol. Biotechnol. 2015, 99, 1513–1519. [Google Scholar] [CrossRef]
- Minoda, A. Agent for Selective Metal Recovery, Metal Recovery Method, and Metal Elution Method. U.S. Patent US20190024209A1, 24 January 2019. [Google Scholar]
- Okamoto, O.K.; Pinto, E.; Latorre, L.R.; Bechara, E.J.H.; Colepicolo, P. Antioxidant Modulation in Response to Metal-Induced Oxidative Stress in Algal Chloroplasts. Arch. Environ. Contam. Toxicol. 2001, 40, 18–24. [Google Scholar]
- Miazek, K.; Iwanek, W.; Remacle, C.; Richel, A.; Goffin, D. Effect of metals, metalloids and metallic nanoparticles on microalgae growth and industrial product biosynthesis: A review. Int. J. Mol. Sci. 2015, 16, 23929–23969. [Google Scholar] [CrossRef] [Green Version]
- Priyadarshini, E.; Priyadarshini, S.S.; Pradhan, N. Heavy metal resistance in algae and its application for metal nanoparticle synthesis. Appl. Microbiol. Biotechnol. 2019, 103, 3297–3316. [Google Scholar] [CrossRef]
- Schönknecht, G.; Chen, W.; Ternes, C.M.; Barbier, G.G.; Shrestha, R.P.; Stanke, M.; Bräutigam, A.; Baker, B.J.; Banfield, J.F.; Garavito, R.M.; et al. Gene Transfer from Bacteria and Archaea Facilitated Evolution of an Extremophilic Eukaryote. Science 2013, 339, 1207–1210. [Google Scholar] [CrossRef]
- Allen, M.B. Studies with cyanidium caldarium, an anomalously pigmented chlorophyte. Arch. Mikrobiol. 1959, 32, 270–277. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirakov, M.; Palmieri, M.; Iovinella, M.; Davis, S.J.; Petriccione, M.; di Cicco, M.R.; De Stefano, M.; Ciniglia, C. Cyanidiophyceae (Rhodophyta) Tolerance to Precious Metals: Metabolic Response to Palladium and Gold. Plants 2021, 10, 2367. https://doi.org/10.3390/plants10112367
Sirakov M, Palmieri M, Iovinella M, Davis SJ, Petriccione M, di Cicco MR, De Stefano M, Ciniglia C. Cyanidiophyceae (Rhodophyta) Tolerance to Precious Metals: Metabolic Response to Palladium and Gold. Plants. 2021; 10(11):2367. https://doi.org/10.3390/plants10112367
Chicago/Turabian StyleSirakov, Maria, Maria Palmieri, Manuela Iovinella, Seth J. Davis, Milena Petriccione, Maria Rosa di Cicco, Mario De Stefano, and Claudia Ciniglia. 2021. "Cyanidiophyceae (Rhodophyta) Tolerance to Precious Metals: Metabolic Response to Palladium and Gold" Plants 10, no. 11: 2367. https://doi.org/10.3390/plants10112367
APA StyleSirakov, M., Palmieri, M., Iovinella, M., Davis, S. J., Petriccione, M., di Cicco, M. R., De Stefano, M., & Ciniglia, C. (2021). Cyanidiophyceae (Rhodophyta) Tolerance to Precious Metals: Metabolic Response to Palladium and Gold. Plants, 10(11), 2367. https://doi.org/10.3390/plants10112367