
Phytochemicals as Biopesticides against the Pinewood Nematode Bursaphelenchus xylophilus: A Review on Essential Oils and Their Volatiles

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Pine Wilt Disease and the Pinewood Nematode
3. Pest Management
4. Research on Anti-Pinewood Nematode Essential Oils
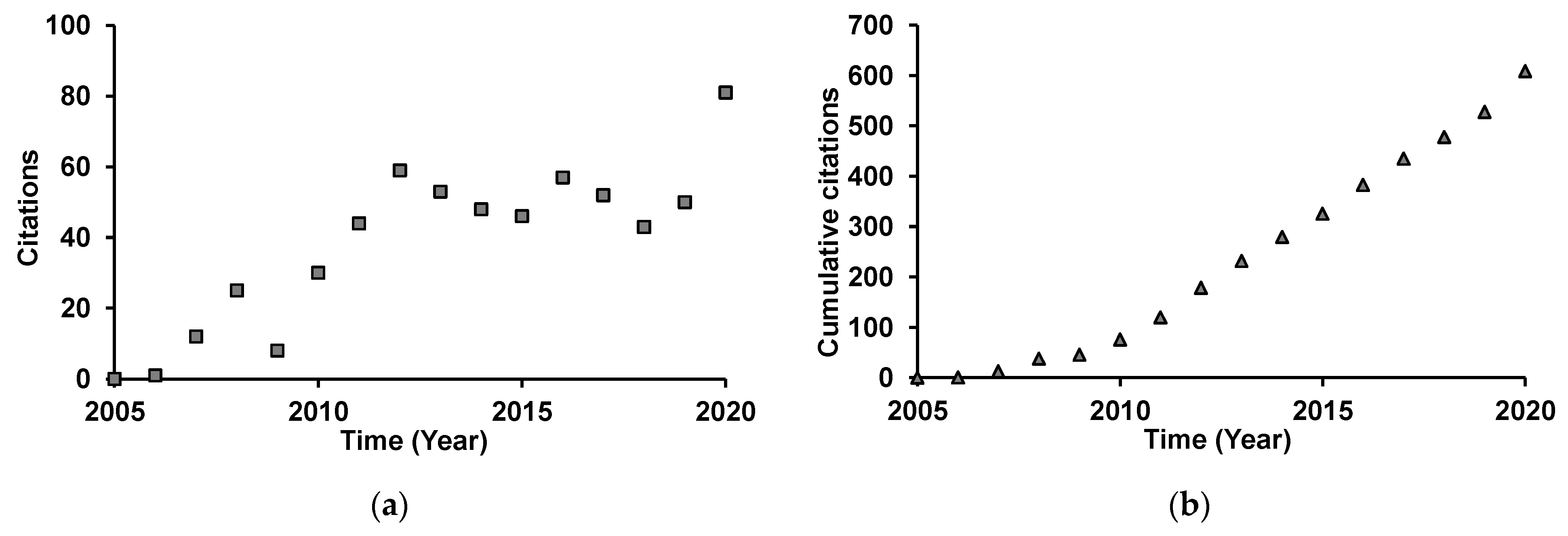
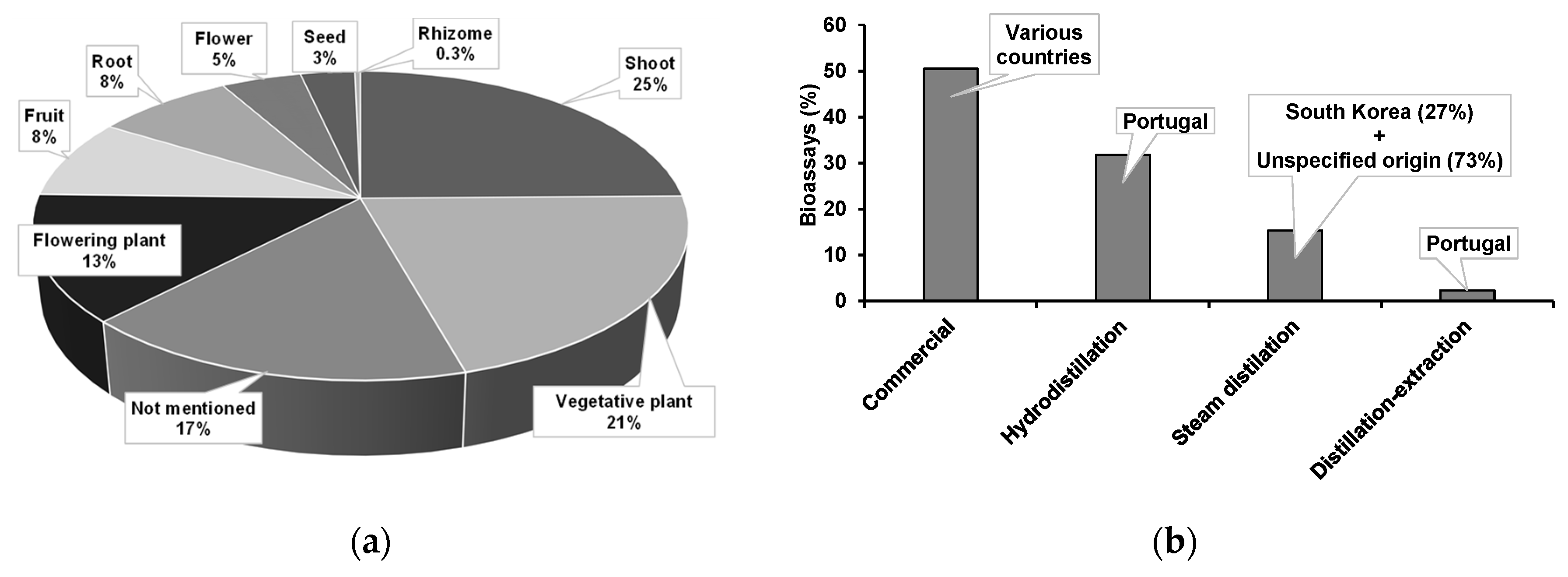
4.1. Bibliographic Sources
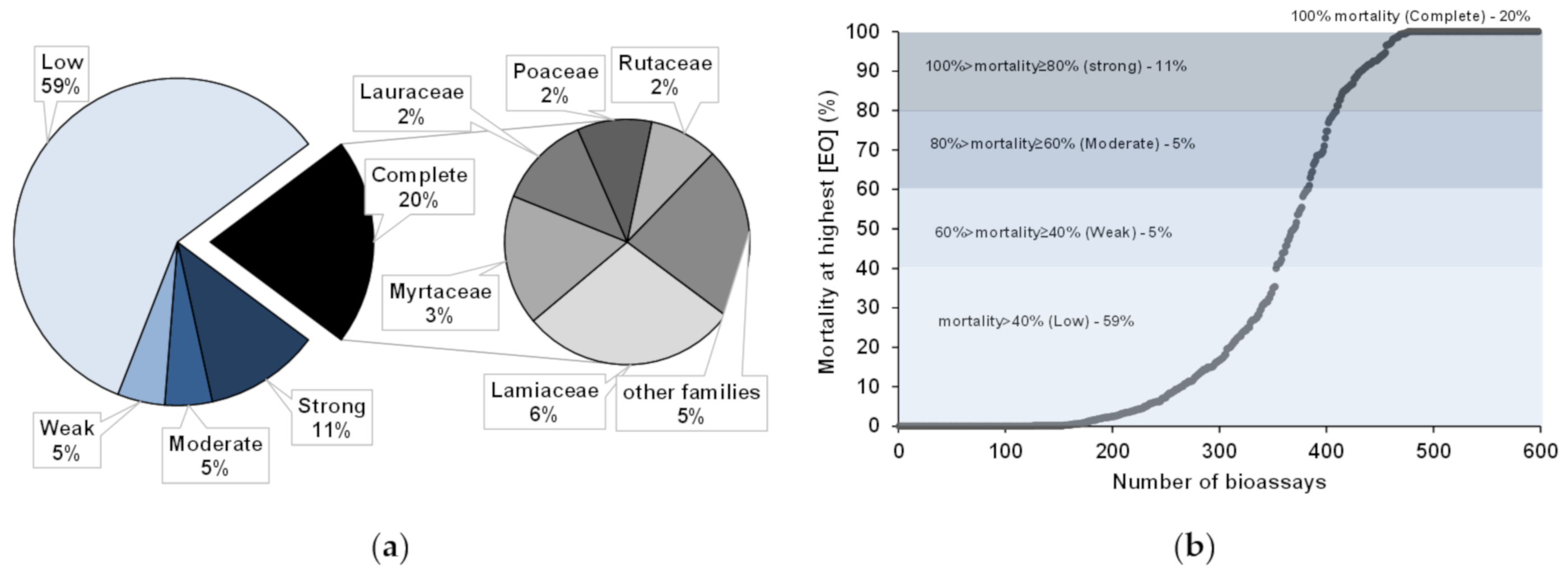
4.2. Anti-Pinewood Nematode Bioassays
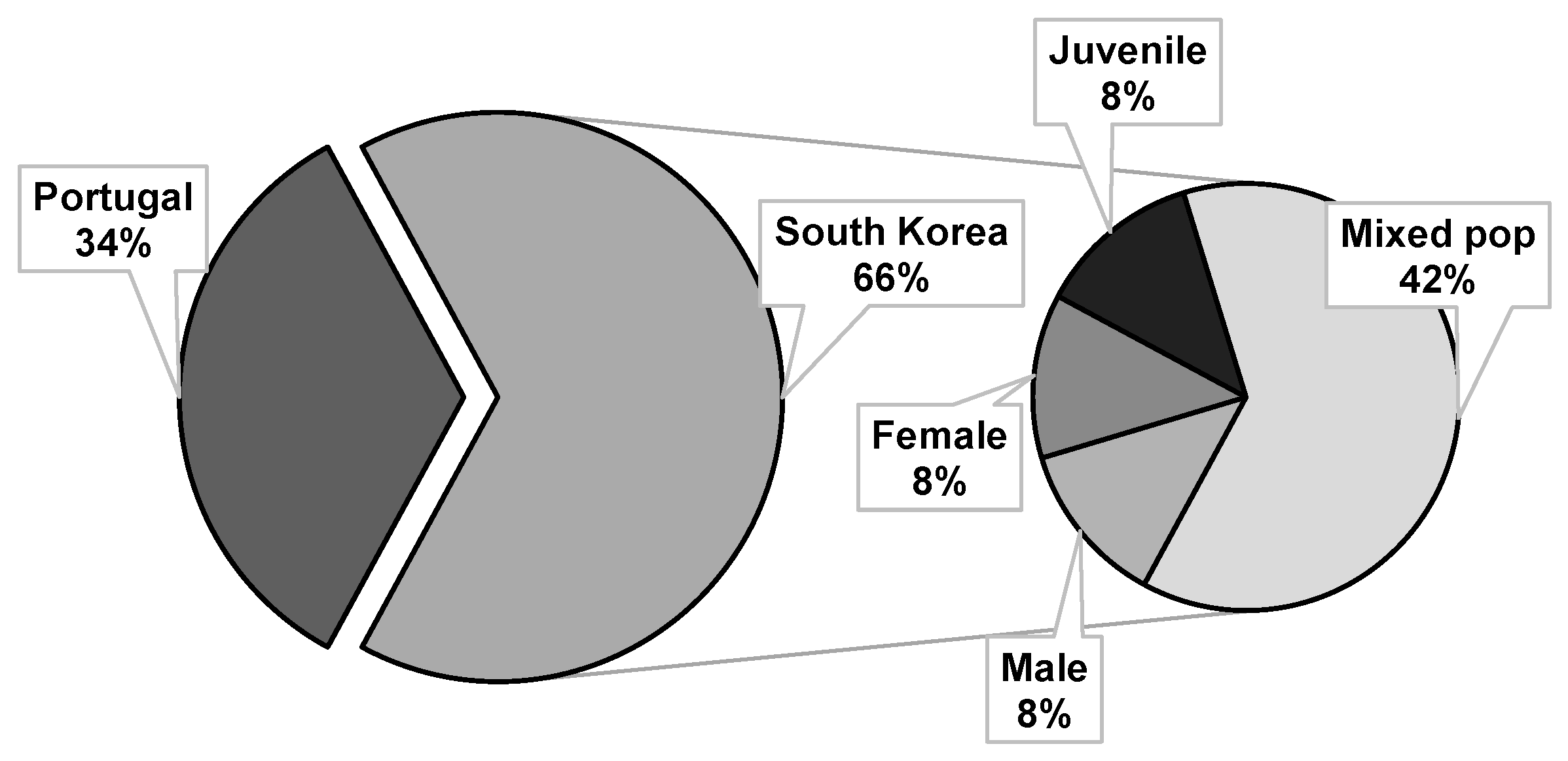
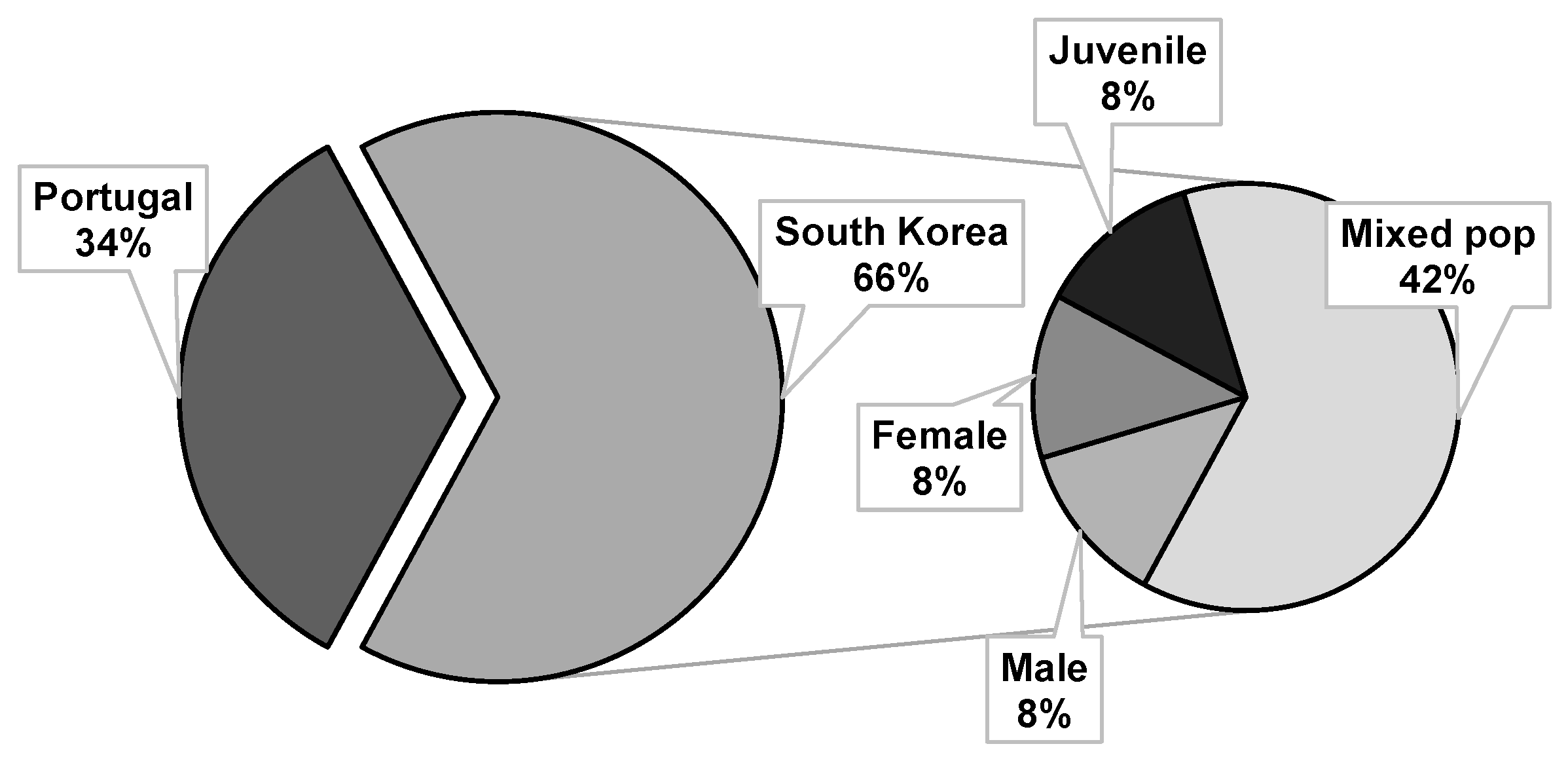
4.3. Bioassayed Pinewood Nematodes
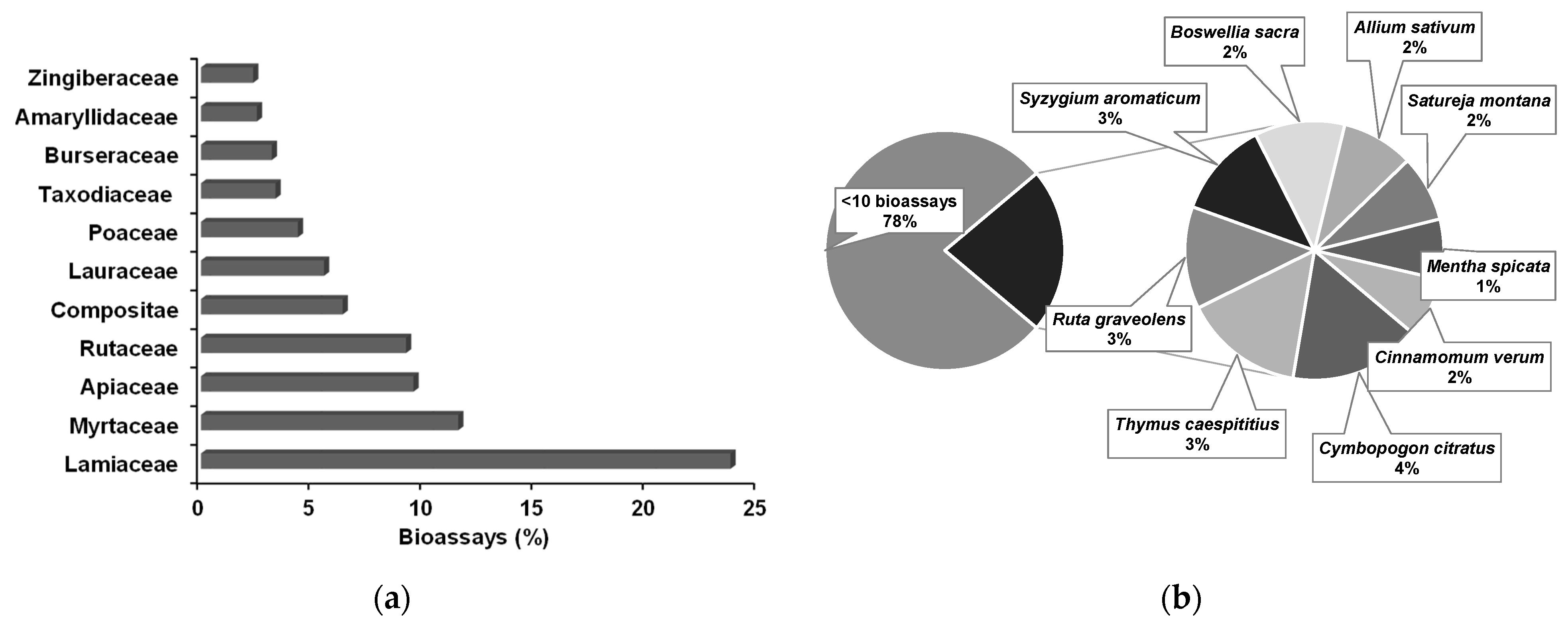
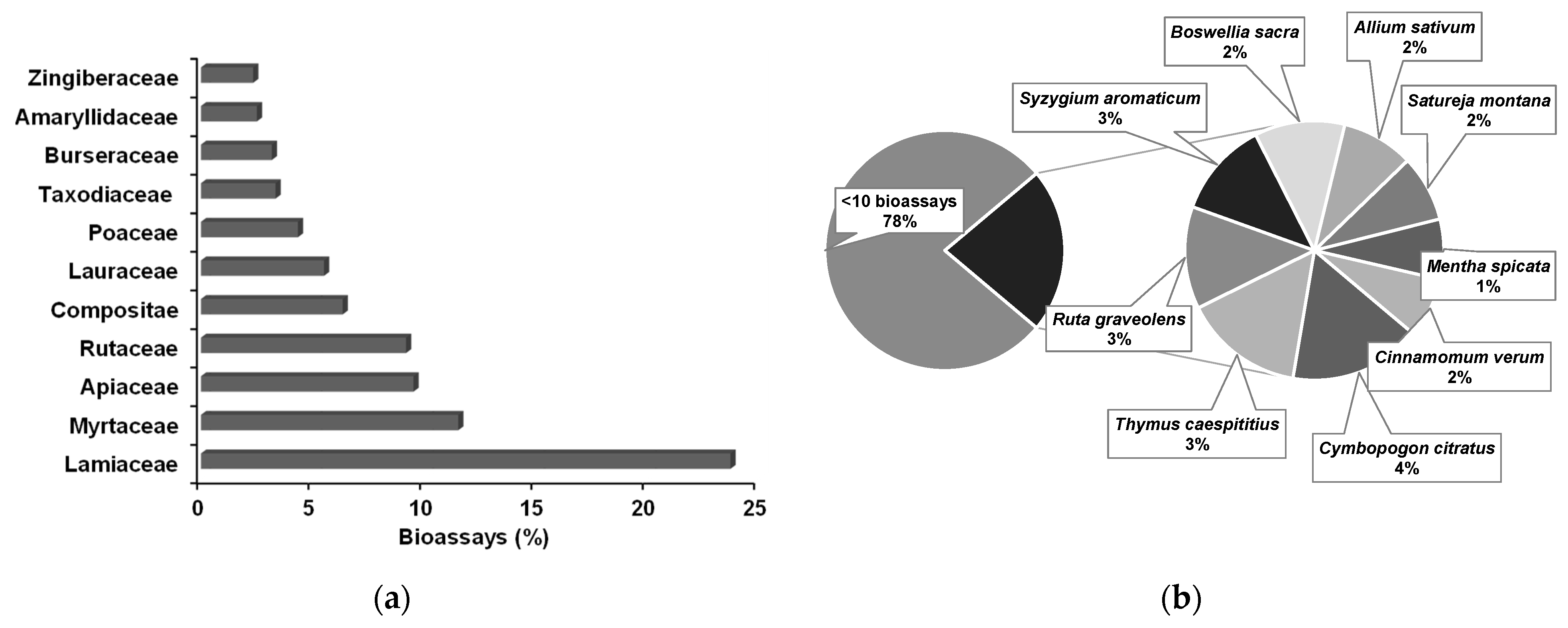
4.4. Essential Oils
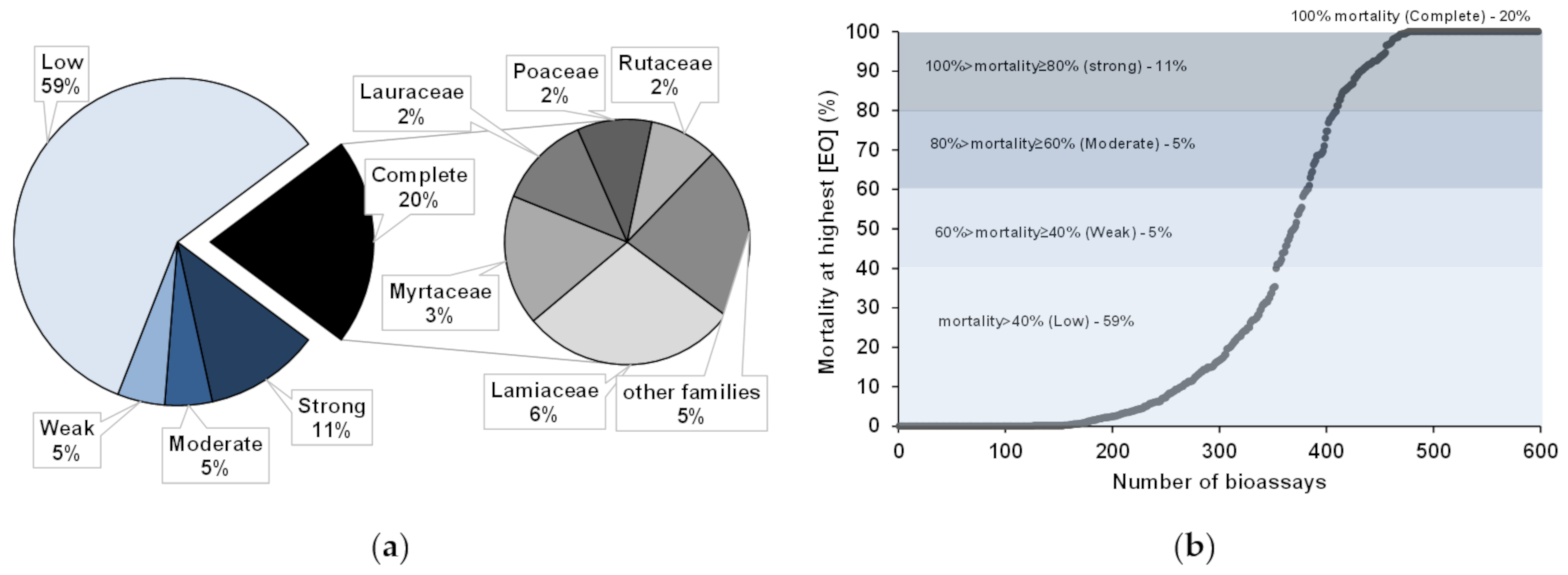
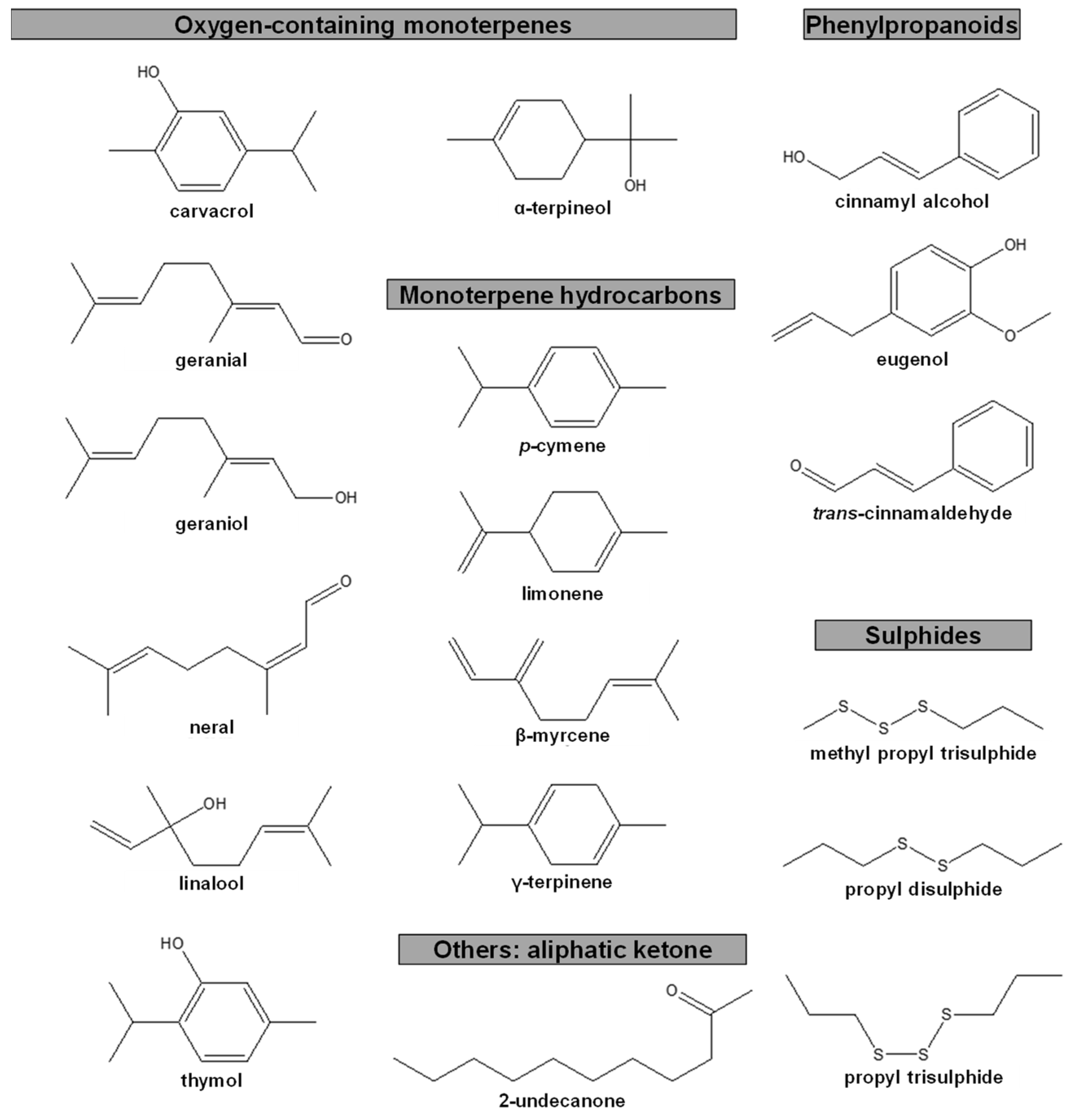
5. Anti-PWN EOs and Their Composition
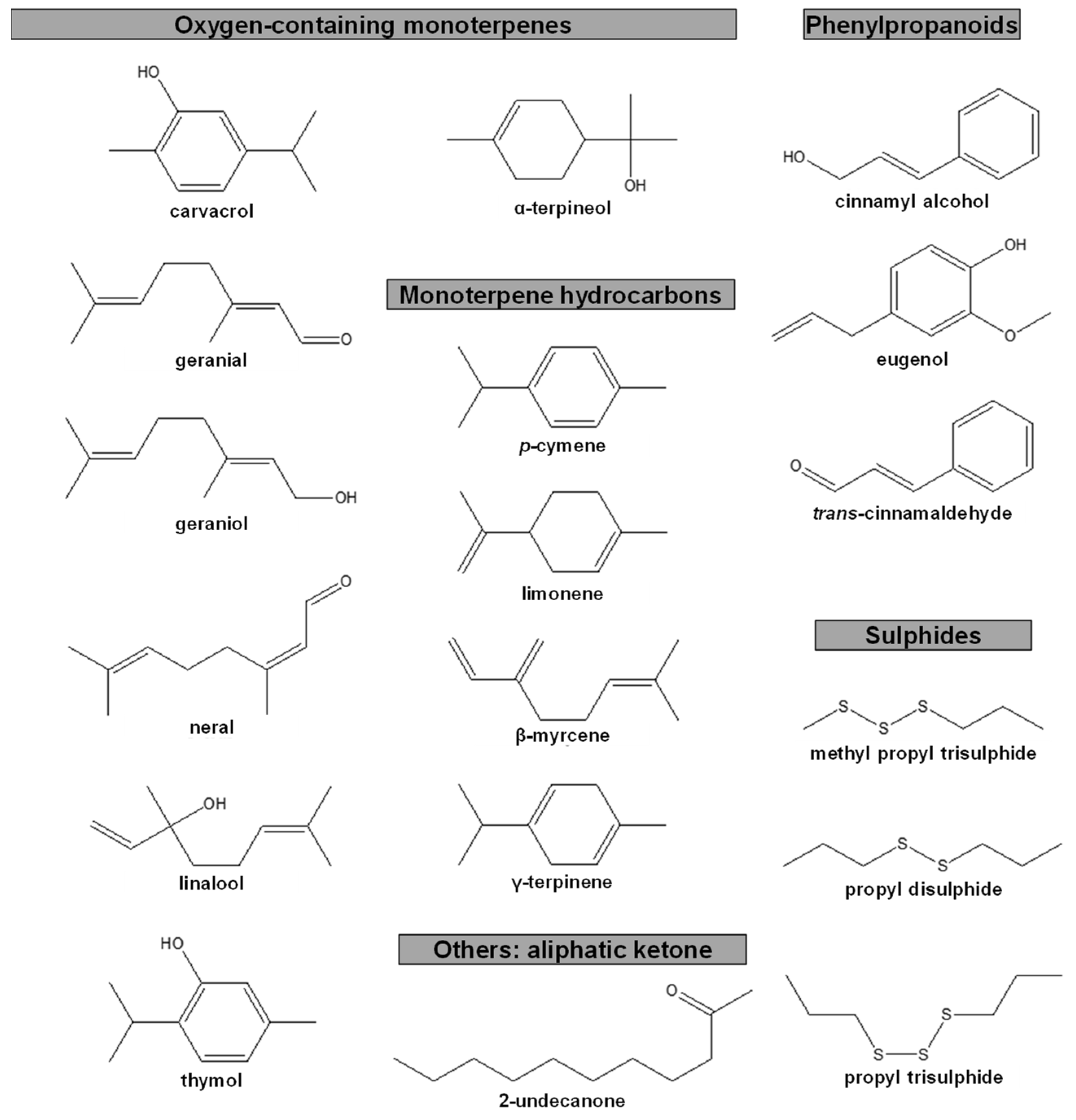
Anti-Pinewood Nematode Essential Oil Compounds
6. Future Challenges for Research on Anti-PWN EOs
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Boyd, I.L.; Freer-Smith, P.H.; Gilligan, C.A.; Godfray, H.C.J. The Consequence of Tree Pests and Diseases for Ecosystem Services. Science 2013, 342, 1235773. [Google Scholar] [CrossRef]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar] [CrossRef]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef]
- Mamiya, Y. History of pine wilt disease in Japan. J. Nematol. 1988, 20, 219–226. [Google Scholar]
- Kim, B.-N.; Kim, J.H.; Ahn, J.-Y.; Kim, S.; Cho, B.-K.; Kim, Y.-H.; Min, J. A short review of the pinewood nematode, Bursaphelenchus xylophilus. Toxicol. Environ. Health Sci. 2020, 12, 297–304. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, Z.; Chen, T.; Zhao, T.; Song, L.; Mei, L. Fertilization and clear-cutting effects on greenhouse gas emissions of pinewood nematode damaged Masson pine plantation. Ecosyst. Health Sustain. 2021, 7, 1868271. [Google Scholar] [CrossRef]
- Futai, K. Pine Wilt in Japan: From first incidence to the present. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 5–12. ISBN 978-4-431-75655-2. [Google Scholar]
- Mota, M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Rodrigues, J.M. National eradication programme for the pinewood nematode. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Mota, M.M., Vieira, P., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 5–14. ISBN 978-1-4020-8455-3. [Google Scholar]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus xylophilus, causal agent of Pine Wilt Disease on Pinus pinaster in Northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef]
- Fonseca, L.; Cardoso, J.M.S.; Lopes, A.; Pestana, M.; Abreu, F.; Nunes, N.; Mota, M.; Abrantes, I. The pinewood nematode, Bursaphelenchus xylophilus, in Madeira Island. Helminthologia 2012, 49, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Robertson, L.; Cobacho Arcos, S.; Escuer, M.; Santiago Merino, R.; Esparrago, G.; Abelleira, A.; Navas, A. Incidence of the pinewood nematode Bursaphelenchus xylophlius Steiner & Buhrer, 1934 (Nickle, 1970) in Spain. Nematology 2011, 13, 755–757. [Google Scholar] [CrossRef]
- Takai, K.; Soejima, T.; Suzuki, T.; Kawazu, K. Development of a water-soluble preparation of emamectin benzoate and its preventative effect against the wilting of pot-grown pine trees inoculated with the pine wood nematode, Bursaphelenchus xylophilus. Pest Manag. Sci. 2001, 57, 463–466. [Google Scholar] [CrossRef]
- Chitwood, D.J. Nematicides. In Encyclopedia of Agrochemicals; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003; Volume 17, pp. 473–474. ISBN 9780123864543. [Google Scholar]
- Chitwood, D.J. Research on plant-parasitic nematode biology conducted by the United States Department of Agriculture-Agricultural Research Service. Pest Manag. Sci. 2003, 59, 748–753. [Google Scholar] [CrossRef]
- Seabright, K.; Davila-Flores, A.; Myers, S.; Taylor, A. Efficacy of methyl bromide and alternative fumigants against pinewood nematode in pine wood samples. J. Plant Dis. Prot. 2020, 127, 393–400. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J.C. Factors affecting secondary metabolite production in plants: Volatile components and essential oils. Flavour Fragr. J. 2008, 23, 213–226. [Google Scholar] [CrossRef]
- Kong, J.O.; Lee, S.M.; Moon, Y.S.; Lee, S.G.; Ahn, Y.J. Nematicidal Activity of Plant Essential Oils against Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae). J. Asia Pac. Entomol. 2006, 9, 173–178. [Google Scholar] [CrossRef]
- Vicente, C.; Espada, M.; Vieira, P.; Mota, M. Pine Wilt Disease: A threat to European forestry. Eur. J. Plant Pathol. 2012, 133, 89–99. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Hasegawa, K.; Mota, M.; Vicente, C.S.L. Bacterial role in pine wilt disease development-review and future perspectives. Environ. Microbiol. Rep. 2015, 7, 51–63. [Google Scholar] [CrossRef]
- Proença, D.N.; Francisco, R.; Kublik, S.; Schöler, A.; Vestergaard, G.; Schloter, M.; Morais, P.V. The Microbiome of Endophytic, Wood Colonizing Bacteria from Pine Trees as Affected by Pine Wilt Disease. Sci. Rep. 2017, 7, 4205. [Google Scholar] [CrossRef] [Green Version]
- Vicente, C.S.L.; Soares, M.; Faria, J.M.S.; Ramos, A.P.; Inácio, M.L. Insights into the Role of Fungi in Pine Wilt Disease. J. Fungi 2021, 7, 780. [Google Scholar] [CrossRef]
- Kuroda, K. Physiological incidences related to symptom development and wilting mechanism. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 204–222. ISBN 978-4-431-75655-2. [Google Scholar]
- Yazaki, K.; Takanashi, T.; Kanzaki, N.; Komatsu, M.; Levia, D.F.; Kabeya, D.; Tobita, H.; Kitao, M.; Ishida, A. Pine wilt disease causes cavitation around the resin canals and irrecoverable xylem conduit dysfunction. J. Exp. Bot. 2018, 69, 589–602. [Google Scholar] [CrossRef]
- Moens, M.; Perry, R.N. Migratory plant endoparasitic nematodes: A group rich in contrasts and divergence. Annu. Rev. Phytopathol. 2009, 47, 313–332. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Mota, M.; Vieira, P.; Butcher, R.A.; Sun, J. Interspecific communication between pinewood nematode, its insect vector, and associated microbes. Trends Parasitol. 2014, 30, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Mamiya, Y. Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef]
- Wang, Y.; Yamada, T.; Sakaue, D.; Suzuki, K. Influence of Fungi on Multiplication and Distribution of the Pinewood Nematode. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Mota, M.M., Vieira, P., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 115–128. ISBN 978-1-4020-8455-3. [Google Scholar]
- Stamps, W.T.; Linit, M.J. Interaction of intrinsic and extrinsic chemical cues in the behaviour of Bursaphelenchus xylophilus (Aphelenchida: Aphelenchoididae) in relation to its beetle vectors. Nematology 2001, 3, 295–301. [Google Scholar] [CrossRef]
- Futai, K. Pine Wood Nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Zhang, R.; Li, D.; Wang, F.; Jiang, S.; Wang, J. Trehalose in pine wood nematode participates in DJ3 formation and confers resistance to low-temperature stress. BMC Genom. 2021, 22, 524. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Langer, S.; Carrasquinho, I.; Bergström, E.; Larson, T.; Thomas-Oates, J.; António, C. Pinus pinaster Early Hormonal Defence Responses to Pinewood Nematode (Bursaphelenchus xylophilus) Infection. Metabolites 2021, 11, 227. [Google Scholar] [CrossRef]
- Jones, J.T.; Moens, M.; Mota, M.; Li, H.; Kikuchi, T. Bursaphelenchus xylophilus: Opportunities in comparative genomics and molecular host-parasite interactions. Mol. Plant Pathol. 2008, 9, 357–368. [Google Scholar] [CrossRef]
- Nose, M.; Shiraishi, S. Breeding for resistance to pine wilt disease. In Pine Wilt Disease; Springer: Tokyo, Japan, 2008; pp. 334–350. ISBN 9784431756545. [Google Scholar]
- Carrasquinho, I.; Lisboa, A.; Inácio, M.L.; Gonçalves, E. Genetic variation in susceptibility to pine wilt disease of maritime pine (Pinus pinaster Aiton) half-sib families. Ann. For. Sci. 2018, 75, 85. [Google Scholar] [CrossRef] [Green Version]
- Menéndez-Gutiérrez, M.; Alonso, M.; Toval, G.; Díaz, R. Testing of selected Pinus pinaster half-sib families for tolerance to pinewood nematode (Bursaphelenchus xylophilus). Forestry 2018, 91, 38–48. [Google Scholar] [CrossRef]
- Kamata, N. Integrated pest management of pine wilt disease in Japan: Tactics and strategies. In Pine Wilt Disease; Springer: Tokyo, Japan, 2008; pp. 304–322. ISBN 9784431756545. [Google Scholar]
- Xu, F. Recent advances in the integrated management of the pine wood nematode in China. In Pine Wilt Disease; Springer: Tokyo, Japan, 2008; pp. 323–333. ISBN 9784431756545. [Google Scholar]
- Bi, Z.; Gong, Y.; Huang, X.; Yu, H.; Bai, L.; Hu, J. Efficacy of four nematicides against the reproduction and development of pinewood Nematode, Bursaphelenchus xylophilus. J. Nematol. 2015, 47, 126–132. [Google Scholar]
- Shimazu, M. Biological control of the Japanese pine sawyer beetle, monochamus alternatus. In Pine Wilt Disease; Springer: Tokyo, Japan, 2008; pp. 351–370. ISBN 9784431756545. [Google Scholar]
- Nakamura, K. Attraction trap for monitoring monochamus alternatus adults-its usefulness and limitations. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Springer: Dordrecht, The Netherlands, 2008; pp. 369–378. ISBN 9784431756545. [Google Scholar]
- Kim, J.; Lee, S.M.; Park, C.G. Bursaphelenchus xylophilus is killed by homologues of 2-(1-undecyloxy)-1-ethanol. Sci. Rep. 2016, 6, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; McVeigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.A.; et al. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef] [Green Version]
- Espada, M.; Silva, A.C.; Van Den Akker, S.E.; Cock, P.J.A.; Mota, M.; Jones, J.T. Identification and characterization of parasitism genes from the pinewood nematode Bursaphelenchus xylophilus reveals a multilayered detoxification strategy. Mol. Plant Pathol. 2016, 17, 286–295. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Hong, H.; Xie, R.; Ji, J.; Guo, K.; Bai, L.; Tang, J.; Yu, H.; Ye, J.; Hu, J. Molecular characterization and functional analysis of daf-8 in the pinewood nematode, Bursaphelenchus xylophilus. J. For. Res. 2021. [Google Scholar] [CrossRef]
- Hao, X.; Wang, B.; Chen, J.; Wang, B.; Xu, J.; Pan, J.; Ma, L. Molecular characterization and functional analysis of multidrug resistance-associated genes of Pinewood nematode (Bursaphelenchus xylophilus) for nematicides. Pestic. Biochem. Physiol. 2021, 177, 104902. [Google Scholar] [CrossRef]
- Hu, L.-J.; Wu, X.-Q.; Ding, X.-L.; Ye, J.-R. Comparative transcriptomic analysis of candidate effectors to explore the infection and survival strategy of Bursaphelenchus xylophilus during different interaction stages with pine trees. BMC Plant Biol. 2021, 21, 224. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, X.; Guo, K.; Chen, S.-N.; Su, X. Transcriptomic insights into the effects of CytCo, a novel nematotoxic protein, on the pine wood nematode Bursaphelenchus xylophilus. BMC Genom. 2021, 22, 394. [Google Scholar] [CrossRef]
- Kim, J.-C.; Lee, M.-R.; Kim, S.; Park, S.-E.; Lee, S.-J.; Shin, T.-Y.; Kim, W.-J.; Kim, J. Transcriptome Analysis of the Japanese Pine Sawyer Beetle, Monochamus alternatus, Infected with the Entomopathogenic Fungus Metarhizium anisopliae JEF-197. J. Fungi 2021, 7, 373. [Google Scholar] [CrossRef]
- Berger, C.; Laurent, F. Trunk injection of plant protection products to protect trees from pests and diseases. Crop Prot. 2019, 124, 104831. [Google Scholar] [CrossRef]
- Stejskal, V.; Douda, O.; Zouhar, M.; Manasova, M.; Dlouhy, M.; Simbera, J.; Aulicky, R. Wood penetration ability of hydrogen cyanide and its efficacy for fumigation of Anoplophora glabripennis, Hylotrupes bajulus (Coleoptera), and Bursaphelenchus xylophilus (Nematoda). Int. Biodeterior. Biodegrad. 2014, 86, 189–195. [Google Scholar] [CrossRef]
- Yang, Z.; Yu, Z.; Lei, L.; Xia, Z.; Shao, L.; Zhang, K.; Li, G. Nematicidal effect of volatiles produced by Trichoderma sp. J. Asia Pac. Entomol. 2012, 15, 647–650. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Y.; Wang, C.; Wang, Y.; Sung, C. Esteya vermicola controls the pinewood nematode, Bursaphelenchus xylophilus, in pine seedlings. J. Nematol. 2017, 49, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Sasanelli, N.; Konrat, A.; Migunova, V.; Toderas, I.; Iurcu-Straistaru, E.; Rusu, S.; Bivol, A.; Andoni, C.; Veronico, P. Review on Control Methods against Plant Parasitic Nematodes Applied in Southern Member States (C Zone) of the European Union. Agriculture 2021, 11, 602. [Google Scholar] [CrossRef]
- Sporleder, M.; Lacey, L.A. Biopesticides. In Insect Pests of Potato; Elsevier: San Diego, CA, USA, 2013; pp. 463–497. [Google Scholar]
- Faria, J.M.S.; Rodrigues, A.M.; Sena, I.; Moiteiro, C.; Bennett, R.N.; Mota, M.; Figueiredo, A.C. Bioactivity of Ruta graveolens and Satureja Montana Essential Oils on Solanum tuberosum Hairy Roots and Solanum tuberosum Hairy Roots with Meloidogyne chitwoodi Co-cultures. J. Agric. Food Chem. 2016, 64, 7452–7458. [Google Scholar] [CrossRef]
- Sangwan, N.K.; Verma, K.K.; Verma, B.S.; Malik, M.S. Nematicidal Activity of Essential Oils of Cymbopogon Grasses. Nematologica 1985, 31, 93–99. [Google Scholar] [CrossRef]
- Park, I.K.; Park, J.Y.; Kim, K.H.; Choi, K.S.; Choi, I.H.; Kim, C.S.; Shin, S.C. Nematicidal activity of plant essential oils and components from garlic (Allium sativum) and cinnamon (Cinnamomum verum) oils against the pine wood nematode (Bursaphelenchus xylophilus). Nematology 2005, 7, 767–774. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Vicente, C. Essential oils and volatiles as nematodicides against the cyst nematodes Globodera and Heterodera. Biol. Life Sci. Forum 2021, 2–6. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Rodrigues, A.M. Essential oils as potential biopesticides in the control of the genus Meloidogyne: A review. Biol. Life Sci. Forum 2021, 2–6. [Google Scholar] [CrossRef]
- Turek, C.; Stintzing, F.C. Stability of essential oils: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 40–53. [Google Scholar] [CrossRef]
- Andrés, M.F.; González-Coloma, A.; Sanz, J.; Burillo, J.; Sainz, P. Nematicidal activity of essential oils: A review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef] [Green Version]
- Web of Science. Available online: www.webofscience.com (accessed on 29 March 2021).
- Kim, J.; Seo, S.M.; Park, I.K. Nematicidal activity of plant essential oils and components from Gaultheria fragrantissima and Zanthoxylum alatum against the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2011, 13, 87–93. [Google Scholar] [CrossRef]
- Choi, I.H.; Park, J.Y.; Shin, S.C.; Kim, J.; Park, I.K. Nematicidal activity of medicinal plant essential oils against the pinewood nematode (Bursaphelenchus xylophilus). Appl. Entomol. Zool. 2007, 42, 397–401. [Google Scholar] [CrossRef]
- Elbadri, G.A.A.; Lee, D.W.; Park, J.C.; Yu, H.B.; Choo, H.Y.; Lee, S.M.; Lim, T.H. Nematocidal screening of essential oils and herbal extracts against Bursaphelenchus xylophilus. Plant Pathol. J. 2008, 24, 178–182. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.O.; Lee, S.M.; Moon, Y.S.; Lee, S.G.; Ahn, Y.J. Nematicidal activity of cassia and cinnamon oil compounds and related compounds toward Bursaphelenchus xylophilus (Nematoda: Parasitaphelenchidae). J. Nematol. 2007, 39, 31–36. [Google Scholar] [PubMed]
- Choi, I.H.; Shin, S.C.; Park, I.K. Nematicidal activity of onion (Allium cepa) oil and its components against the pine wood nematode (Bursaphelenchus xylophilus). Nematology 2007, 9, 231–235. [Google Scholar] [CrossRef]
- Barbosa, P.; Lima, A.S.; Vieira, P.; Dias, L.S.; Tinoco, M.T.; Barroso, J.G.; Pedro, L.G.; Figueiredo, A.C.; Mota, M. Nematicidal activity of essential oils and volatiles derived from Portuguese aromatic flora against the pinewood nematode, Bursaphelenchus xylophilus. J. Nematol. 2010, 42, 8–16. [Google Scholar]
- Barbosa, P.; Faria, J.M.S.; Mendes, M.D.; Dias, L.S.; Tinoco, M.T.; Barroso, J.G.; Pedro, L.G.; Figueiredo, A.C.; Mota, M. Bioassays Against Pinewood Nematode: Assessment of a Suitable Dilution Agent and Screening for Bioactive Essential Oils. Molecules 2012, 17, 12312–12329. [Google Scholar] [CrossRef] [Green Version]
- Faria, J.M.S.; Barbosa, P.; Bennett, R.N.; Mota, M.; Figueiredo, A.C. Bioactivity against Bursaphelenchus xylophilus: Nematotoxics from essential oils, essential oils fractions and decoction waters. Phytochemistry 2013, 94, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, J.-O.; Park, I.; Choi, K.-S.; Shin, S.-C.; Ahn, Y.-J. Nematicidal and propagation activities of thyme red and white oil compounds toward Bursaphelenchus xylophilus (Nematoda: Parasitaphelenchidae). J. Nematol. 2007, 39, 237–242. [Google Scholar]
- Park, I.-K.K.; Kim, J.; Lee, S.-G.G.; Shin, S.-C.C. Nematicidal activity of plant essential oils and components from Ajowan (Trachyspermum ammi), Allspice (Pimenta dioica) and Litsea (Litsea cubeba) essential oils against pine wood nematode (Bursaphelenchus xylophilus). J. Nematol. 2007, 39, 275–279. [Google Scholar] [PubMed]
- Kim, J.; Seo, S.-M.M.; Lee, S.-G.G.; Shin, S.-C.C.; Park, I.-K.K. Nematicidal Activity of Plant Essential Oils and Components from Coriander (Coriandrum sativum), Oriental Sweetgum (Liquidambar orientalis), and Valerian (Valeriana wallichii) Essential Oils against Pine Wood Nematode (Bursaphelenchus xylophilus). J. Agric. Food Chem. 2008, 56, 7316–7320. [Google Scholar] [CrossRef]
- Martins, M.A.R.; Silva, L.P.; Ferreira, O.; Schröder, B.; Coutinho, J.A.P.; Pinho, S.P. Terpenes solubility in water and their environmental distribution. J. Mol. Liq. 2017, 241, 996–1002. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, J.M.; Fonseca, L.; Abrantes, I. Genetic diversity of ITS sequences of Bursaphelenchus xylophilus. Genet. Mol. Res. 2012, 11, 4508–4515. [Google Scholar] [CrossRef] [PubMed]
- Palomares-Rius, J.E.; Tsai, I.J.; Karim, N.; Akiba, M.; Kato, T.; Maruyama, H.; Takeuchi, Y.; Kikuchi, T. Genome-wide variation in the pinewood nematode Bursaphelenchus xylophilus and its relationship with pathogenic traits. BMC Genom. 2015, 16, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.; Han, H.; Ryu, S.H.; Kim, W. Microsatellite variation in the pinewood nematode, Bursaphelenchus xylophilus (Steiner and Buhrer) Nickle in South Korea. Genes Genom. 2010, 32, 151–158. [Google Scholar] [CrossRef]
- Metge, K.; Bürgermeister, W. Intraspecific Variation in isolates of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) revealed by ISSR and RAPD fingerprints. J. Plant Dis. Prot. 2006, 113, 275–282. [Google Scholar] [CrossRef]
- Vieira, P.; Burgermeister, W.; Mota, M.; Metge, K.; Silva, G. Lack of genetic variation of Bursaphelenchus xylophilus in Portugal revealed by RAPD-PCR analyses. J. Nematol. 2007, 39, 118–126. [Google Scholar]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Hanif, M.A.; Nisar, S.; Khan, G.S.; Mushtaq, Z.; Zubair, M. Essential Oils. In Essential Oil Research; Malik, S., Ed.; Springer International Publishing: Cham. Switzerland, 2019; pp. 3–17. [Google Scholar]
- ISO 9235 Aromatic Natural Raw Materials—Vocabulary. Available online: https://www.iso.org/obp/ui/#iso:std:iso:9235:ed-2:v1:en (accessed on 29 March 2021).
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Shaaban, H.A.E.; El-Ghorab, A.H.; Shibamoto, T. Bioactivity of essential oils and their volatile aroma components: Review. J. Essent. Oil Res. 2012, 24, 203–212. [Google Scholar] [CrossRef]
- León-Méndez, G.; Pájaro-Castro, N.; Pájaro-Castro, E.; Torrenegra-Alarcón, M.; Herrera-Barros, A. Essential oils as a source of bioactive molecules. Rev. Colomb. Cienc. Químico-Farm. 2019, 48, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Faria, J.M.S.; Sena, I.; Ribeiro, B.; Rodrigues, A.M.; Maleita, C.M.N.; Abrantes, I.; Bennett, R.; Mota, M.; Figueiredo, A.C. da Silva Figueiredo, A.C. First report on Meloidogyne chitwoodi hatching inhibition activity of essential oils and essential oils fractions. J. Pest Sci. 2016, 89, 207–217. [Google Scholar] [CrossRef]
- Soares, C.; Morales, H.; Faria, J.; Figueiredo, A.C.; Pedro, L.G.; Venâncio, A. Inhibitory effect of essential oils on growth and on aflatoxins production by Aspergillus parasiticus. World Mycotoxin J. 2016, 9, 525–534. [Google Scholar] [CrossRef]
- Chitwood, D.J. Phytochemical based strategies for nematode control. Annu. Rev. Phytopathol. 2002, 40, 221–249. [Google Scholar] [CrossRef] [Green Version]
- Ntalli, N.G.; Ferrari, F.; Giannakou, I.; Menkissoglu-Spiroudi, U. Phytochemistry and nematicidal activity of the essential oils from 8 greek lamiaceae aromatic plants and 13 terpene components. J. Agric. Food Chem. 2010, 58, 7856–7863. [Google Scholar] [CrossRef] [PubMed]
- Vicente, C.S.L.; Nascimento, F.; Espada, M.; Barbosa, P.; Mota, M.; Glick, B.R.; Oliveira, S. Characterization of Bacteria Associated with Pinewood Nematode Bursaphelenchus xylophilus. PLoS ONE 2012, 7, e46661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Flora Online. Available online: http://www.worldfloraonline.org/ (accessed on 29 March 2021).
- Trindade, H.; Pedro, L.G.; Figueiredo, A.C.; Barroso, J.G. Chemotypes and terpene synthase genes in Thymus genus: State of the art. Ind. Crops Prod. 2018, 124, 530–547. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Mendes, M.D.; Lima, A.S.; Barbosa, P.M.; Ascensão, L.; Barroso, J.G.; Pedro, L.G.; Mota, M.M.; Figueiredo, A.C. Pinus halepensis, Pinus pinaster, Pinus pinea and Pinus sylvestris essential oils chemotypes and monoterpene hydrocarbon enantiomers, before and after inoculation with the pinewood nematode Bursaphelenchus xylophilus. Chem. Biodivers. 2017, 14, e1600153. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Rodrigues, A.M. Metabolomic Variability in the Volatile Composition of Essential Oils from Pinus pinea and P. pinaster. Biol. Life Sci. Forum 2021, 2, 14. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Ferrari, F.; Giannakou, I.; Menkissoglu-Spiroudi, U. Synergistic and antagonistic interactions of terpenes against Meloidogyne incognita and the nematicidal activity of essential oils from seven plants indigenous to Greece. Pest Manag. Sci. 2011, 67, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.H.; Kim, J.; Shin, S.C.; Park, I.K. Nematicidal activity of monoterpenoids against the pine wood nematode (Bursaphelenchus xylophilus). Russ. J. Nematol. 2007, 15, 35–40. [Google Scholar]
- Bashyal, P.; Pandey, R.P.; Thapa, S.B.; Kang, M.-K.; Kim, C.-J.; Sohng, J.K. Biocatalytic Synthesis of Non-Natural Monoterpene O -Glycosides Exhibiting Superior Antibacterial and Antinematodal Properties. ACS Omega 2019, 4, 9367–9375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, I.S.; Faria, J.M.S.; Figueiredo, A.C.; Pedro, L.G.; Trindade, H.; Barroso, J.G. Menthol and geraniol biotransformation and glycosylation capacity of Levisticum officinale hairy roots. Planta Med. 2009, 75, 387–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, J.M.S.; Sena, I.; da Silva, I.V.; Ribeiro, B.; Barbosa, P.; Ascensão, L.; Bennett, R.N.; Mota, M.; Figueiredo, A.C. In vitro co-cultures of Pinus pinaster with Bursaphelenchus xylophilus: A biotechnological approach to study pine wilt disease. Planta 2015, 241, 1325–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, J.M.S.; Sena, I.; Maleita, C.M.; da Silva, I.V.; Ascensão, L.; Abrantes, I.; Bennett, R.N.; Mota, M.; Figueiredo, A.C. In vitro co-culture of Solanum tuberosum hairy roots with Meloidogyne chitwoodi: Structure, growth and production of volatiles. Plant Cell Tissue Organ Cult. 2014, 118, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Faria, J.M.S.; Barbosa, P.; Teixeira, D.M.; Mota, M. A Review on the Nematicidal Activity of Volatile Allelochemicals against the Pinewood Nematode. Environ. Sci. Proc. 2020, 3, 1. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Rodrigues, A.M.; Barbosa, P.; Mota, M. In Silico Screening of Agonist and Antagonist Natural Compounds from Reported Essential Oils against Bursaphelenchus xylophilus. Chem. Proc. 2020, 3, 31. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, S.; Lu, H.; Zhang, D.; Liu, F.; Lin, J.; Zhou, C.; Mu, W. Effects of the plant volatile trans-2-hexenal on the dispersal ability, nutrient metabolism and enzymatic activities of Bursaphelenchus xylophilus. Pestic. Biochem. Physiol. 2017, 143, 147–153. [Google Scholar] [CrossRef]
- Cheng, L.; Xu, S.; Xu, C.; Lu, H.; Zhang, Z.; Zhang, D.; Mu, W.; Liu, F. Effects of trans-2-hexenal on reproduction, growth and behaviour and efficacy against the pinewood nematode, Bursaphelenchus xylophilus. Pest Manag. Sci. 2017, 73, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.M.; Kim, J.; Koh, S.H.; Ahn, Y.J.; Park, I.K. Nematicidal activity of natural ester compounds and their analogues against pine wood nematode, Bursaphelenchus xylophilus. J. Agric. Food Chem. 2014, 62, 9103–9108. [Google Scholar] [CrossRef]
- Seo, S.M.I.; Junheon, K.; Eunae, K.; Park, H.M.I.; Kim, Y.J.; Park, I.L.K. Structure-Activity relationship of aliphatic compounds for nematicidal activity against pine wood nematode (Bursaphelenchus xylophilus). J. Agric. Food Chem. 2010, 58, 1823–1827. [Google Scholar] [CrossRef] [PubMed]
- Faria, J.M.S.; Sena, I.; Moiteiro, C.; Bennett, R.N.; Mota, M.; Cristina Figueiredo, A. Nematotoxic and phytotoxic activity of Satureja montana and Ruta graveolens essential oils on Pinus pinaster shoot cultures and P. pinaster with Bursaphelenchus xylophilus in vitro co-cultures. Ind. Crops Prod. 2015, 77, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Oh, W.-S.; Jeong, P.-Y.; Joo, H.-J.; Lee, J.-E.; Moon, Y.-S.; Cheon, H.-M.; Kim, J.-H.; Lee, Y.-U.; Shim, Y.-H.; Paik, Y.-K. Identification and Characterization of a Dual-Acting Antinematodal Agent against the Pinewood Nematode, Bursaphelenchus xylophilus. PLoS ONE 2009, 4, e7593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, E.; Lee, J.; Chang, P.; Park, I. Development of chitosan-coated nanoemulsions of two sulfides present in onion (Allium cepa) essential oil and their nematicidal activities against the pine wood nematode, Bursaphelenchus xylophilus. Environ. Sci. Pollut. Res. 2021. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EC50 [mg/mL (or µL/mL *)] | |||||
|---|---|---|---|---|---|
| Plant Species/Common Name | Mixed Population | Male | Female | Juvenile | Major Compounds (≥1%) |
| Acorus calamus [18] [Sweet flag/Calamus] | 2.850 | ||||
| Allium cepa [68] [Onion] | 0.018 | 0.014 | 0.012 | Propyl trisulphide 47, propyl disulphide 34, methyl propyl trisulphide 15, methyl propyl disulphide 3 | |
| Aniba rosaeodora [18] [Rosewood] | 2.990 | ||||
| Boswellia sacra [65] [Frankincense] | 0.290 | 0.260 | 0.210 | ||
| Cinnamomum cassia [67] [Cassia/Chinese cinnamon] | 0.084 | trans-Cinnamaldehyde 80, 2-methoxy cinnamaldehyde 13, cinnamyl acetate 4, α-copaene 2, benzaldehyde 1 | |||
| Cinnamomum cassia [67] [Cassia/Chinese cinnamon] | 0.085 | trans-Cinnamaldehyde 91, 2-methoxy cinnamaldehyde 5, cinnamyl acetate 2, trans-cinnamic acid 1 | |||
| Cinnamomum verum [18] [Cinnamon] | 0.120 | ||||
| Cinnamomum zeylanicum [67] [Cinnamon] | 0.064 | trans-Cinnamaldehyde 99, benzaldehyde 1 | |||
| Cinnamomum zeylanicum [67] [Cinnamon] | 0.097 | trans-Cinnamaldehyde 82, cinnamyl acetate 10, 2-methoxycinnamaldehyde 6, benzaldehyde 2 | |||
| Cinnamomum zeylanicum [67] [Cinnamon] | 0.107 | trans-Cinnamaldehyde 61, eugenol 13, α-terpineol 10, p-cymene 8, linalool 3 | |||
| Cinnamomum zeylanicum [67] [Cinnamon] | 0.113 | trans-Cinnamaldehyde 53, limonene 17, cinnamyl alcohol 16, eugenol 13 | |||
| Coriandrum sativum [18] [Coriander] | 0.140 | ||||
| Coriandrum sativum [18] [Coriander] | 2.760 | ||||
| Cymbopogon citratus [69] [Lemongrass] | 0.350 | Geranial 43, neral 29, β-myrcene 25 | |||
| Cymbopogon citratus [71] [Lemongrass] | 0.456* | Geranial 34, neral 22, β-myrcene 20, geraniol 18 | |||
| Cymbopogon citratus [18] [Lemongrass] | 0.570 | ||||
| Cymbopogon nardus [18] [Citronella grass] | 2.110 | ||||
| Genista tridentata [69] [Carqueja #] | 1.060 | 1-Octen-3-ol 9, n-nonanal 7, linalool 7, trans-anethole 5, dodecanoid acid 5, cis-theaspirane 3, 2-undecanone 2 | |||
| Litsea cubeba [73] [Aromatic litsea] | 0.504 | Geranial 39, neral 30, limonene 15 | |||
| Litsea cubeba [18] [Aromatic litsea] | 3.650 | ||||
| Melissa officinalis [18] [Lemon balm] | 4.110 | ||||
| Nepeta tenuifolia [65] [Jing Jie] | 0.470 | 0.490 | 0.410 | ||
| Origanum vulgare [71] [Oregano] | 0.754* | Carvacrol 14, cis-sabinene hydrate 14, γ-terpinene 10 | |||
| Origanum vulgare [71] [Oregano] | 0.850* | α-Terpineol 40, linalool 16, thymol 12 | |||
| Origanum vulgare [69] [Oregano] | 1.210 | Carvacrol 36, carvacrol methyl ether 8, β-caryophyllene 2 | |||
| Origanum vulgare [18] [Oregano] | 1.420 | ||||
| Paeonia × suffruticosa [65] [Tree peony] | 0.320 | 0.340 | 0.260 | ||
| Perilla frutescens [65] [Beefsteak plant/Perilla] | 0.530 | 0.570 | 0.410 | ||
| Pimenta dioica [73] [Allspice] | 0.609 | Eugenol 86, β-caryophyllene 8, methyl eugenol 4, α-humulene 1 | |||
| Pimenta dioica [18] [Allspice] | 1.800 | ||||
| Pimenta racemosa [18] [Bay rum tree] | 2.270 | ||||
| Rosa x damascena [18] [Damask rose] | 4.470 | ||||
| Ruta graveolens [71] [Rue] | 0.184 * | 2-Undecanone 93 | |||
| Ruta graveolens [70] [Rue] | 0.200 | 2-Undecanone 93 | |||
| Ruta graveolens [71] [Rue] | 0.230 * | 2-Undecanone 91 | |||
| Ruta graveolens [70] [Rue] | 0.230 | 2-Undecanone 94 | |||
| Ruta graveolens [71] [Rue] | 0.232 * | 2-Undecanone 94 | |||
| Satureja hortensis [18] [Summer savory] | 1.150 | ||||
| Satureja montana [71] [Winter savory] | 0.261 * | Carvacrol 64, γ-terpinene 18 | |||
| Satureja montana [70] [Winter savory] | 0.340 | γ-Terpinene 41, carvacrol 35, p-cymene 8, α-terpinene 4, β-myrcene 3, α-pinene 2, α-thujene 2 | |||
| Satureja montana [70] [Winter savory] | 0.350 | Carvacrol 40, p-cymene 20, thymol 15, γ-terpinene 4, borneol 4, terpinen-4-ol 4 | |||
| Satureja montana [69] [Winter savory] | 0.380 | Carvacrol 39, γ-terpinene 40, p-cymene 7, β-myrcene 3, α-pinene 2 | |||
| Syzygium aromaticum [18] [Clove] | 0.880 | ||||
| Thymbra capitata [71] [Conehead thyme] | 0.265 * | Carvacrol 68, γ-terpinene 11 | |||
| Thymbra capitata [69] [Conehead thyme] | 0.500 | Carvacrol 75 | |||
| Thymbra capitata [18] [Conehead thyme] | 0.820 | ||||
| Thymus caespititius [69] [Tormentelo #] | 0.390 | Carvacrol 65, carvacrol acetate 11 | |||
| Thymus caespititius [71] [Tormentelo #] | 0.972 * | Carvacrol 54, carvacrol acetate 10 | |||
| Thymus vulgaris [18] [Thyme] | 0.820 | ||||
| Thymus vulgaris [72] [Thyme] | 1.390 | Thymol 58, p-cymene 18, γ-terpinene 9, linalool 4, carvacrol 3 | |||
| Thymus vulgaris [72] [Thyme] | 1.640 | Thymol 48, p-cymene 18, linalool 11, γ-terpinene 7, limonene 4, camphor 4, terpinen-4-ol 2, carvacrol 2 | |||
| Trachyspermum ammi [73] [Ajwain] | 0.431 | Thymol 42, γ-terpinene 28, p-cymene 24, β-pinene 1 | |||
| EO Compounds | EC50 (mg/mL) |
|---|---|
| Diallyl trisulphide | 0.003–0.004 |
| Propyl sulphide | 0.004–0.005 |
| Methyl propyl trisulphide | 0.017–0.023 |
| Cinnamyl acetate | 0.033–2.766 |
| Diallyl disulphide | 0.037–0.047 |
| trans-Cinnamaldehyde | 0.057 |
| Geranial | 0.120 |
| Isoeugenol | 0.200 |
| Methyl isoeugenol | 0.210 |
| Geraniol | 0.430 |
| Eugenol | 0.480–1.212 |
| Methyl eugenol | 0.517 |
| Neral | 0.525 |
| trans-Cinnamic acid | 0.750 |
| EO Compound | Lowest Concentration (mg/mL) |
|---|---|
| Methyl trans-cinnamate | 0.063 |
| Decanol | 0.200 |
| trans-2-Decenal | 0.200 |
| Ethyl trans-cinnamate | 0.250 |
| Methyl propyl trisulphide | 0.250 |
| Propyl sulphide | 0.250 |
| trans-2-Decen-1-ol | 0.400 |
| cis-Asarone | 0.800 |
| trans-Cinnamyl alcohol | 0.800 |
| Decanal | 1.000 |
| Eugenol | 1.000 |
| Geranial | 1.000 |
| Isoeugenol | 1.000 |
| Methyl isoeugenol | 1.000 |
| Benzaldehyde | 2.000 |
| Dodecanal | 2.000 |
| Nonanal | 2.000 |
| Octanal | 2.000 |
| Undecanal | 2.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faria, J.M.S.; Barbosa, P.; Vieira, P.; Vicente, C.S.L.; Figueiredo, A.C.; Mota, M. Phytochemicals as Biopesticides against the Pinewood Nematode Bursaphelenchus xylophilus: A Review on Essential Oils and Their Volatiles. Plants 2021, 10, 2614. https://doi.org/10.3390/plants10122614
Faria JMS, Barbosa P, Vieira P, Vicente CSL, Figueiredo AC, Mota M. Phytochemicals as Biopesticides against the Pinewood Nematode Bursaphelenchus xylophilus: A Review on Essential Oils and Their Volatiles. Plants. 2021; 10(12):2614. https://doi.org/10.3390/plants10122614
Chicago/Turabian StyleFaria, Jorge M. S., Pedro Barbosa, Paulo Vieira, Cláudia S. L. Vicente, Ana Cristina Figueiredo, and Manuel Mota. 2021. "Phytochemicals as Biopesticides against the Pinewood Nematode Bursaphelenchus xylophilus: A Review on Essential Oils and Their Volatiles" Plants 10, no. 12: 2614. https://doi.org/10.3390/plants10122614
APA StyleFaria, J. M. S., Barbosa, P., Vieira, P., Vicente, C. S. L., Figueiredo, A. C., & Mota, M. (2021). Phytochemicals as Biopesticides against the Pinewood Nematode Bursaphelenchus xylophilus: A Review on Essential Oils and Their Volatiles. Plants, 10(12), 2614. https://doi.org/10.3390/plants10122614