
Nuclear Translocation of Soybean MPK6, GmMPK6, Is Mediated by Hydrogen Peroxide in Salt Stress


 ,
,  ,
, 
{kind=link}
Abstract
1. Introduction
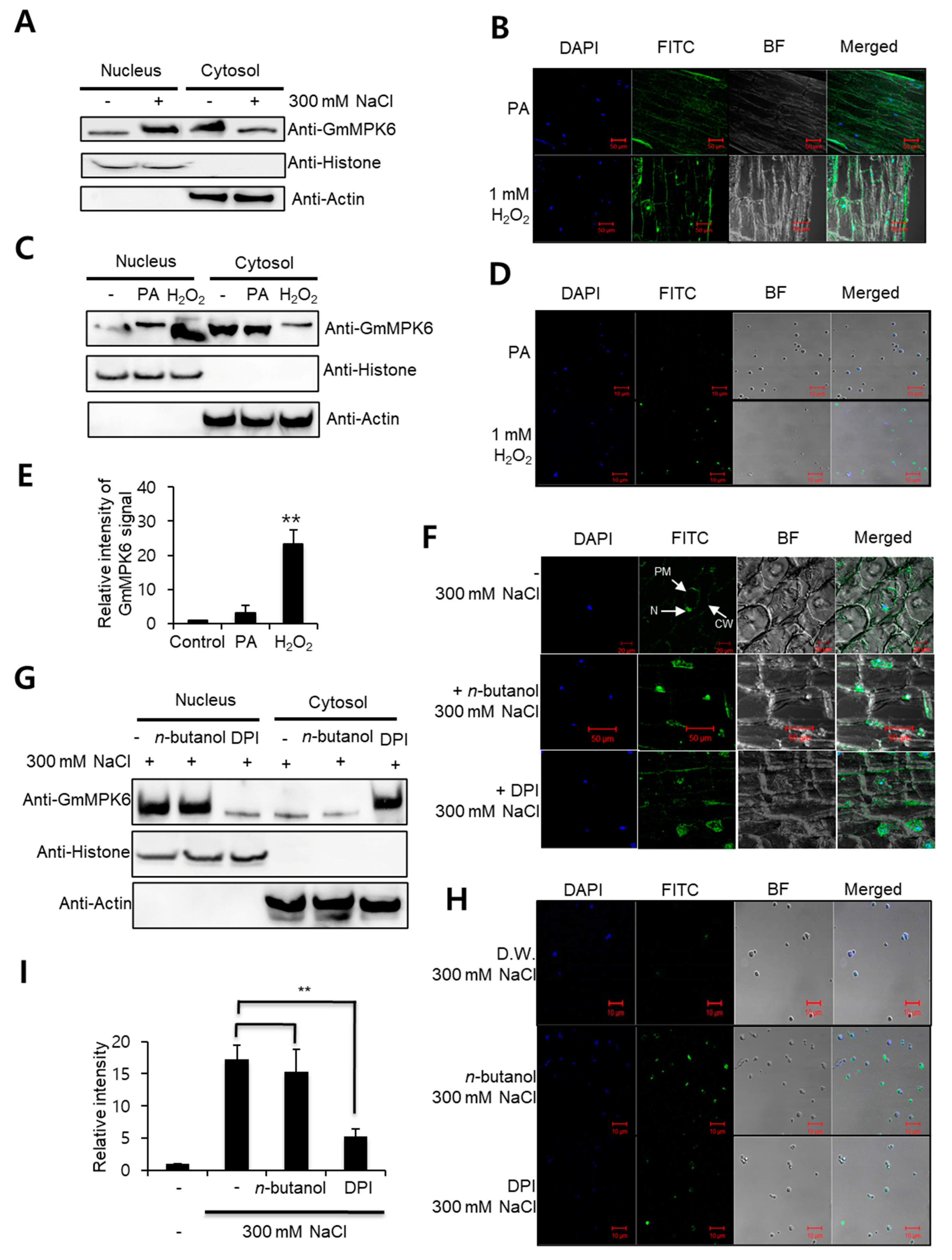
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Subcellular Immunoblot Analysis
4.3. Immunolocalization Assay
4.4. Nuclei Isolation and GmMPK6 Detection
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Ichimura, K.; Mizoguchi, T.; Yoshida, R.; Yuasa, T.; Shinozaki, K. Various abiotic stresses vapidly activate arabidopsis map kinases atmpk4 and atmpk6. Plant J. 2000, 24, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Nie, J.; Cao, C.; Jin, Y.; Yan, M.; Wang, F.; Liu, J.; Xiao, Y.; Liang, Y.; Zhang, W. Phosphatidic acid mediates salt stress response by regulation of mpk6 in arabidopsis thaliana. New Phytol. 2010, 188, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Pitzschke, A.; Datta, S.; Persak, H. Salt stress in arabidopsis: Lipid transfer protein azi1 and its control by mitogen-activated protein kinase mpk3. Mol. Plant 2014, 7, 722–738. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.F.; Chou, W.C.; Huang, D.D.; Huang, H.J. Transcriptional regulation of a rice mitogen-activated protein kinase gene, osmapk4, in response to environmental stresses. Plant Cell Physiol. 2002, 43, 958–963. [Google Scholar] [CrossRef]
- Munnik, T.; Ligterink, W.; Meskiene, I.; Calderini, O.; Beyerly, J.; Musgrave, A.; Hirt, H. Distinct osmo-sensing protein kinase pathways are involved in signalling moderate and severe hyper-osmotic stress. Plant J. 1999, 20, 381–388. [Google Scholar] [CrossRef]
- Im, J.H.; Lee, H.; Kim, J.; Kim, H.B.; An, C.S. Soybean mapk, gmk1 is dually regulated by phosphatidic acid and hydrogen peroxide and translocated to nucleus during salt stress. Mol. Cells 2012, 34, 271–278. [Google Scholar] [CrossRef]
- Munnik, T.; Meijer, H.J.; Ter Riet, B.; Hirt, H.; Frank, W.; Bartels, D.; Musgrave, A. Hyperosmotic stress stimulates phospholipase d activity and elevates the levels of phosphatidic acid and diacylglycerol pyrophosphate. Plant J. 2000, 22, 147–154. [Google Scholar] [CrossRef]
- Hong, Y.; Zhang, W.; Wang, X. Phospholipase d and phosphatidic acid signalling in plant response to drought and salinity. Plant Cell Environ. 2010, 33, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Li, J.; Bao, Q.; Zhang, F.; Tulaxi, G.; Wang, Z. Salt-induced hydrogen peroxide is involved in modulation of antioxidant enzymes in cotton. Crop J. 2016, 4, 490–498. [Google Scholar] [CrossRef][Green Version]
- Park, J.; Gu, Y.; Lee, Y.; Yang, Z.; Lee, Y. Phosphatidic acid induces leaf cell death in arabidopsis by activating the rho-related small g protein gtpase-mediated pathway of reactive oxygen species generation. Plant Physiol. 2004, 134, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hirt, H.; Lee, Y. Phosphatidic acid activates a wound-activated mapk in glycine max. Plant J. 2001, 26, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.Y.; Xue, H.W. Phosphatidic acid plays key roles regulating plant development and stress responses. J. Integr. Plant Biol. 2018, 60, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Palladina, T.O.; Kurylenko, I.M.; Chyzhykova, O.A. Plant salt-tolerance can be increased by synthetic compounds with antioxidant and antiradical properties. Acta Physiol. Plant 2005, 27, 22. [Google Scholar]
- O’Reilly, M.C.; Scott, S.A.; Daniels, J.S.; Morrison, R.; Engers, J.L.; Oguin, T.; Thomas, P.; Brown, H.A.; Lindsley, C.W. A next generation pld2 inhibitor with improved physiochemical properties and dmpk profile for translational in vivo. In Probe Reports from the Nih Molecular Libraries Program; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2010. [Google Scholar]
- Bolomini-Vittori, M.; Mennens, S.F.B.; Joosten, B.; Fransen, J.; Du, G.; van den Dries, K.; Cambi, A. Pld-dependent phosphatidic acid microdomains are signaling platforms for podosome formation. Sci. Rep. 2019, 9, 3556. [Google Scholar] [CrossRef] [PubMed]
- Hung, S.H.; Yu, C.W.; Lin, C.H. Hydrogen peroxide functions as a stress signal in plants. Bot. Bull. Acad. Sin. 2005, 46, 1–10. [Google Scholar]
- Torres, M.A.; Dangl, J.L.; Jones, J.D.G. Arabidopsis gp91(phox) homologues atrbohd and atrbohf are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Sagi, M.; Fluhr, R. Superoxide production by plant homologues of the gp91(phox) nadph oxidase. Modulation of activity by calcium and by tobacco mosaic virus infection. Plant Physiol. 2001, 126, 1281–1290. [Google Scholar] [CrossRef]
- Mazel, A.; Leshem, Y.; Tiwari, B.S.; Levine, A. Induction of salt and osmotic stress tolerance by overexpression of an intracellular vesicle trafficking protein atrab7 (atrabg3e). Plant Physiol. 2004, 134, 118–128. [Google Scholar] [CrossRef]
- Orman-Ligeza, B.; Parizot, B.; de Rycke, R.; Fernandez, A.; Himschoot, E.; Van Breusegem, F.; Bennett, M.J.; Perilleux, C.; Beeckman, T.; Draye, X. Rboh-mediated ros production facilitates lateral root emergence in arabidopsis. Development 2016, 143, 3328–3339. [Google Scholar] [CrossRef]
- Plotnikov, A.; Zehorai, E.; Procaccia, S.; Seger, R. The mapk cascades: Signaling components, nuclear roles and mechanisms of nuclear translocation. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2011, 1813, 1619–1633. [Google Scholar] [CrossRef] [PubMed]
- Im, J.H.; Lee, H.; Kim, J.; Kim, H.B.; Seyoung, K.; Kim, B.M.; An, C.S. A salt stress-activated mitogen-activated protein kinase in soybean is regulated by phosphatidic acid in early stages of the stress response. J. Plant Biol. 2012, 55, 303–309. [Google Scholar] [CrossRef]
- Liu, J.Z.; Braun, E.; Qiu, W.L.; Shi, Y.F.; Marcelino-Guimaraes, F.C.; Navarre, D.; Hill, J.H.; Whitham, S.A. Positive and negative roles for soybean mpk6 in regulating defense responses. Mol. Plant Microbe Interact. 2014, 27, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.R.; Srinivasan, T.; Kirti, P.B. A mitogen-activated protein kinase gene, ahmpk3 of peanut: Molecular cloning, genomic organization, and heterologous expression conferring resistance against spodoptera litura in tobacco. Mol. Genet. Genom. 2009, 282, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.C.; Yoon, H.W.; Kim, C.Y.; Moon, B.C.; Cheong, Y.H.; Han, H.J.; Lee, S.M.; Kang, K.Y.; Kim, M.C.; Lee, S.Y.; et al. Alternative splicing of the osbwmk1 gene generates three transcript variants showing differential subcellular localizations. Biochem. Biophys. Res. Commun. 2007, 360, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Broughton, W.J.; Dilworth, M.J. Control of leghaemoglobin synthesis in snake beans. Biochem. J. 1971, 125, 1075–1080. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Im, J.H.; Son, S.; Ko, J.-H.; Kim, K.-H.; An, C.S.; Han, K.-H. Nuclear Translocation of Soybean MPK6, GmMPK6, Is Mediated by Hydrogen Peroxide in Salt Stress. Plants 2021, 10, 2611. https://doi.org/10.3390/plants10122611
Im JH, Son S, Ko J-H, Kim K-H, An CS, Han K-H. Nuclear Translocation of Soybean MPK6, GmMPK6, Is Mediated by Hydrogen Peroxide in Salt Stress. Plants. 2021; 10(12):2611. https://doi.org/10.3390/plants10122611
Chicago/Turabian StyleIm, Jong Hee, Seungmin Son, Jae-Heung Ko, Kyung-Hwan Kim, Chung Sun An, and Kyung-Hwan Han. 2021. "Nuclear Translocation of Soybean MPK6, GmMPK6, Is Mediated by Hydrogen Peroxide in Salt Stress" Plants 10, no. 12: 2611. https://doi.org/10.3390/plants10122611
APA StyleIm, J. H., Son, S., Ko, J.-H., Kim, K.-H., An, C. S., & Han, K.-H. (2021). Nuclear Translocation of Soybean MPK6, GmMPK6, Is Mediated by Hydrogen Peroxide in Salt Stress. Plants, 10(12), 2611. https://doi.org/10.3390/plants10122611