
From Past to Present: Biotechnology in Mexico Using Algae and Fungi



Abstract
:1. Introduction
2. The Use of Algae and Fungi in Pre-Hispanic Mexico
2.1. Algae
2.2. Fungi
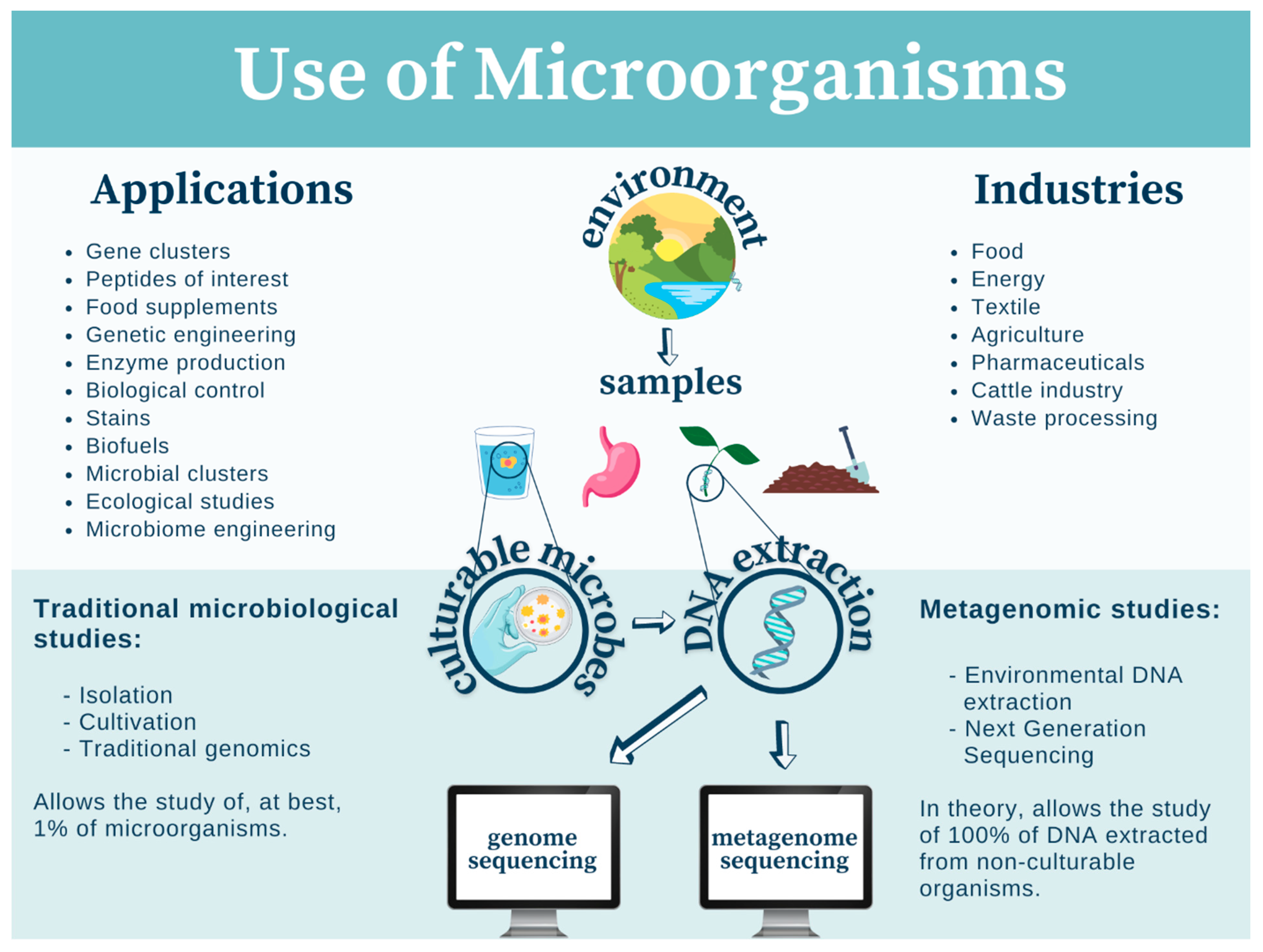
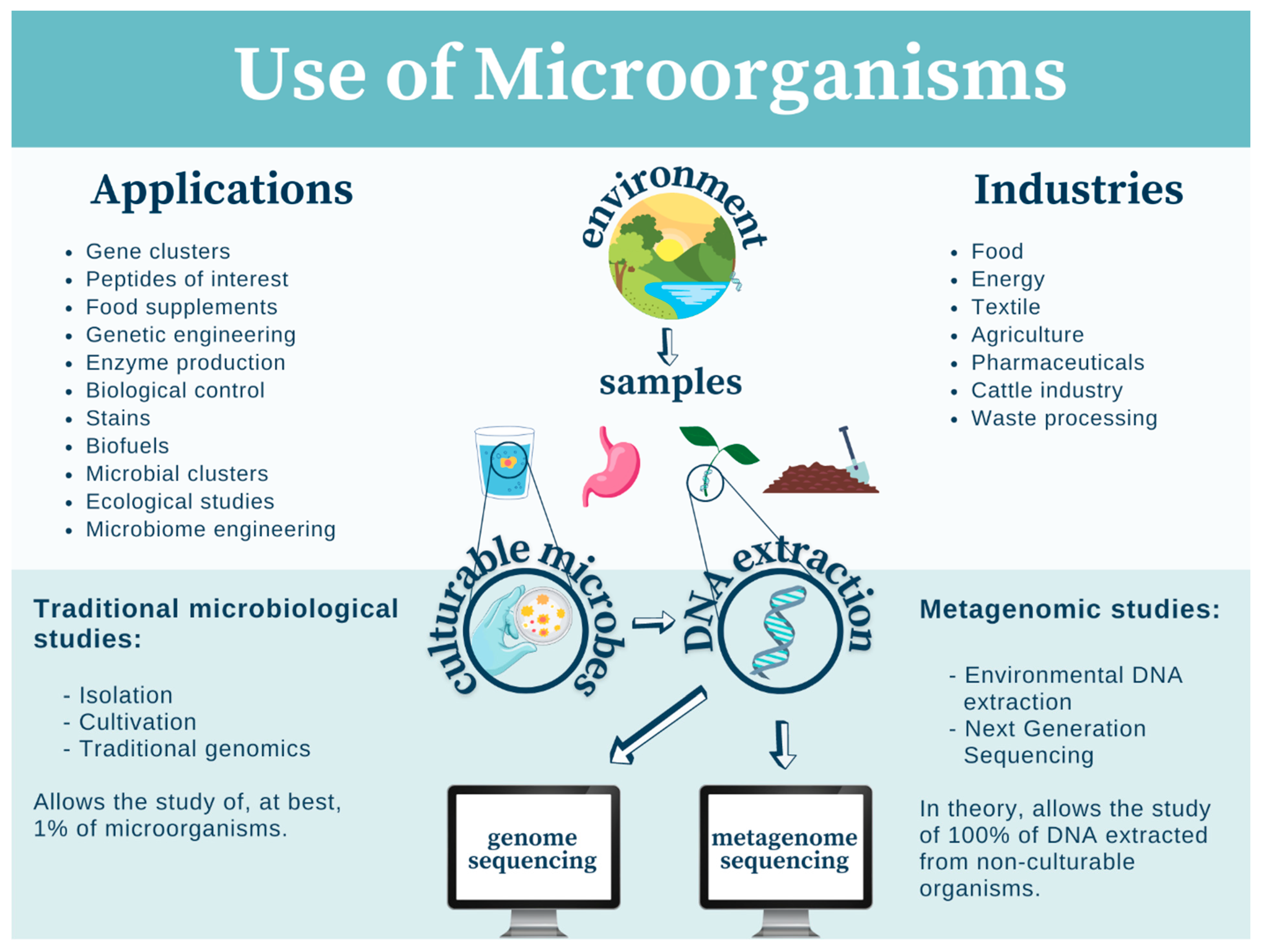
3. Mexican Microorganisms in Modern-Day Biotechnology
3.1. Algae
3.2. Fungi
3.2.1. Enzyme Production
3.2.2. Biocontrol of Pests/Pathogens
3.2.3. Production of Pharmaceuticals
3.2.4. Food Improvement
3.2.5. Waste Fermentation

{kind=link}
| Species | Field | Use | Reference |
|---|---|---|---|
| FUNGI | |||
| Amanita basii+ | nutrition | food (supplement) | [28] |
| Amanita laurae+ | nutrition | food (supplement) | [28] |
| Aspergillus awamori | biotechnology | pectinase production | [70] |
| Aspergillus niger | biotechnology | invertase production | [64,65,66] |
| Aspergillus sp. | biotechnology | xylanase production | [69] |
| Aspergillus sp. | biotechnology | pectinase production | [70] |
| Aspergillus spp. | biotechnology | tannase production | [68] |
| Calvatia cyathiformis | ludic use | projectile (toy) | [28] |
| Candida parapsilosis | bioprocessing | waste treatment | [93] |
| Cercospora piaropi | environment | pest control (water hyacinth) | [71] |
| Ganoderma lucidum | pharmaceuticals | medical | [85] |
| Ganoderma oerstedii | pharmaceuticals | ergosta-7,22-dien-3-one production | [82] |
| Ganoderma oerstedii | pharmaceuticals | medical | [28] |
| Metarhizium anisopliae | environment | pest control (Asian blue tick) | [74] |
| Nigrospora sp. | pharmaceuticals | paclitaxel production | [81] |
| Panaeolus spp. | ludic use | hallucinogen | [22] |
| Pichia anomala | bioprocessing | waste treatment | [93] |
| Pisolithus spp. | materials | projectile (toy) | [28] |
| Pleurotus agaves * | nutrition | food (supplement) | [87] |
| Pleurotus djamor | nutrition | food (supplement) | [28] |
| Pleurotus djamor | environment | pest control (nematode) | [78] |
| Pleurotus opuntiae * | nutrition | food (supplement) | [28] |
| Psilocybe spp. | ludic use | hallucinogen | [22] |
| Psilocybe cubensis | ludic use | hallucinogen | [22] |
| Pycnoporus sanguineus | pharmaceuticals | medical | [28] |
| Rhodotorula mucilaginosa | pharmaceuticals | bacterial biofilm inhibition | [77] |
| Saccharomyces cerevisiae | bioprocessing | waste treatment | [93] |
| Stropharia spp. | ludic use | hallucinogen | [22] |
| Trametes maxima+ | bioprocessing | waste treatment | [91] |
| Trametes sp. | bioprocessing | waste treatment | [91] |
| Ustilago maydis | nutrition | food (supplement) | [28] |
| Volvariella bombycina | nutrition | food (supplement) | [28] |
| ALGAE | |||
| Arthrospira platensis | nutrition | food (supplement) | [16] |
| Arthrospira platensis | pharmaceuticals | neuro and hepatoprotective | [56] |
| Biddulphia alternans | pisciculture | food (supplement) | [57] |
| Ceramium nitens+ | pharmaceuticals | antimicrobial activity | [58] |
| Eucheuma isiforme | pharmaceuticals | thickener and stabilizer | [60,61] |
| Eucheuma isiforme | nutrition | food (thickener and stabilizer) | [60,61] |
| Gelidium robustum | biotechnology | agar extraction | [31,32,33] |
| Macrocystis pyrifera | biotechnology | alginate extraction | [31,32,33] |
| Porphyra perforata | nutrition | food (supplement) | [31,32,33] |
3.2.6. Ludic Uses of Hallucinogenic Fungi
3.3. Other Microorganisms
4. Perspective: Utilizing Mexican Biodiversity for the Improvement of Society
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mittermeier, R.A.; Mittermeier, C.G.; Gil, P.R. Megadiversity: Earth’s Biologically Wealthiest Nations, 1st ed.; CEMEX: San Pedro Garza García, Mexico, 1997; ISBN 9789686397505. [Google Scholar]
- Gómez-Pompa, A.; Kaus, A. From pre-Hispanic to future conservation alternatives: Lessons from Mexico. Proc. Natl. Acad. Sci. USA 1999, 96, 5982–5986. [Google Scholar] [CrossRef] [Green Version]
- Chambers, P.J.; Pretorius, I.S. Fermenting knowledge: The history of winemaking, science and yeast research. EMBO Rep. 2010, 11, 914–920. [Google Scholar] [CrossRef] [Green Version]
- Lappe-Oliveras, P.; Moreno-Terrazas, R.; Arrizón-Gaviño, J.; Herrera-Suárez, T.; García-Mendoza, A.; Gschaedler-Mathis, A. Yeasts associated with the production of Mexican alcoholic nondistilled and distilled Agave beverages. FEMS Yeast Res. 2008, 8, 1037–1052. [Google Scholar] [CrossRef] [Green Version]
- Ruiz Oronoz, M. Estudios en México sobre levaduras. In Proceedings of the Memoria del Congreso Cientıfico Mexicano; Universidad Nacional Autonoma de Mexico: Ciudad de México, Mexico, 1953; pp. 127–149. [Google Scholar]
- Mc Govern, P.E.; Toro, F.H.; Hall, G.R.; Davidson, T.; Prigge, K.P. Pre-Hispanic distillation? A biomolecular archaeological investigation. J. Archaeol. Anthropol. 2019, 509, 2. [Google Scholar] [CrossRef]
- Needham, J.; Ping-Yu, H.; Gwei-djen, L.; Sivin, N. Spagyrical discovery and invention: Apparatus, theories and gifts. In Science and Civilisation in China; Cambridge University Press: Cambridge, UK, 1980; p. 804. [Google Scholar]
- Pérez-Armendáriz, B.; Cardoso-Ugarte, G.A. Traditional fermented beverages in Mexico: Biotechnological, nutritional, and functional approaches. Food Res. Int. 2020, 136, 109307. [Google Scholar] [CrossRef]
- Romero-Luna, H.E.; Hernández-Sánchez, H.; Dávila-Ortiz, G. Traditional fermented beverages from Mexico as a potential probiotic source. Ann. Microbiol. 2017, 67, 577–586. [Google Scholar] [CrossRef]
- Ramírez-Guzmán, K.N.; Torres-León, C.; Martinez-Medina, G.A.; de la Rosa, O.; Hernández-Almanza, A.; Alvarez-Perez, O.B.; Araujo, R.; González, L.R.; Londoño, L.; Ventura, J. Traditional fermented beverages in Mexico. In Fermented Beverages; Woodhead Publishing: London, UK, 2019; pp. 605–635. [Google Scholar]
- Escalante, A.; López Soto, D.R.; Velázquez Gutiérrez, J.E.; Giles-Gómez, M.; Bolívar, F.; López-Munguía, A. Pulque, a traditional Mexican alcoholic fermented beverage: Historical, microbiological, and technical aspects. Front. Microbiol. 2016, 7, 1026. [Google Scholar] [CrossRef] [Green Version]
- Lemus Fuentes, E. Los enemas prehispánicos como instrumentos para aplicar probióticos. Repos. Nac. CONACYT 2006, 10, 17–26. [Google Scholar]
- Wacher, C.; Cañas, A.; Bárzana, E.; Lappe, P.; Ulloa, M.; Owens, J.D. Microbiology of Indian and Mestizo pozol fermentations. Food Microbiol. 2000, 17, 251–256. [Google Scholar] [CrossRef]
- Godínez Ortega, J.L. Colectores de algas de México (1787–1954). Acta Bot. Mex. 2008, 85, 75–97. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R.K.; Guzman-Juarez, M. The nutritional potential of the algae. Plant Foods Man 1978, 2, 195–202. [Google Scholar] [CrossRef]
- Pirie, N.W. The Spirulina algae. Food Protein Sources 1975, 4, 33–39. [Google Scholar]
- Ortega, M.M. Study of the edible algae of the Valley of Mexico. Bot. Mar. 1972, 15, 162–166. [Google Scholar]
- Godínez, J.L.; Ortega, M.M.; Garduño, G.; Oliva, M.G.; Vilaclara, G. Traditional knowledge of Mexican continental algae. J. Ethnobiol. 2001, 21, 57–88. [Google Scholar]
- Dupey García, É. The materiality of color in the body ornamentation of Aztec gods. Res Anthropol. Aesthet. 2015, 65, 72–88. [Google Scholar] [CrossRef]
- Hernández-Santiago, F.; Moreno, J.P.; Xoconostle-Cázares, B.; Almaraz-Suárez, J.J.; Ojeda-Trejo, E.; Mata Montes de Oca, G.; Díaz-Aguilar, I. Traditional knowledge and use of wild mushrooms by Mixtecs or Ñuu savi, the people of the rain, from Southeastern Mexico. J. Ethnobiol. Ethnomed. 2016, 12, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, G. Diversity and use of traditional Mexican medicinal fungi. A review. Int. J. Med. Mushrooms 2008, 10, 209–217. [Google Scholar] [CrossRef]
- Carod-Artal, F.J. Alucinógenos en las culturas precolombinas mesoamericanas. Neurología 2011, 30, 42–49. [Google Scholar] [CrossRef]
- Passie, T.; Seifert, J.; Schneider, U.; Emrich, H.M. The pharmacology of psilocybin. Addict. Biol. 2002, 7, 357–364. [Google Scholar] [CrossRef]
- Boege, E.; Vidriales Chan, G.; Garcıa Coll, I.; Mondragón, M.; Rivas, A.J.; Lozada, M.P.; Soto, F. El Patrimonio Biocultural de los Pueblos Indıgenas de México: Hacia la Conservación In Situ de la Biodiversidad y Agrodiversidad en los Territorios Indıgenas, 1st ed.; Instituto Nacional de Antropología e Historia: Ciudad de México, Mexico, 2008.
- Delgado-Sánchez, P.; Jiménez-Bremont, J.F.; de la Luz Guerrero-González, M.; Flores, J. Effect of fungi and light on seed germination of three Opuntia species from semiarid lands of central Mexico. J. Plant Res. 2013, 126, 643–649. [Google Scholar] [CrossRef]
- Jiménez Ruiz, M.; Pérez-Moreno, J.; Almaraz-Suárez, J.J.; Torres-Aquino, M. Hongos silvestres con potencial nutricional, medicinal y biotecnológico comercializados en Valles Centrales, Oaxaca. Rev. Mex. Cienc. Agrícolas 2013, 4, 199–213. [Google Scholar] [CrossRef] [Green Version]
- Garibay-Orijel, R.; Ruan-Soto, F. Listado de los hongos silvestres consumidos como alimento tradicional en México. In La Etnomicología en México, Estado del Arte; CONACYT, UAEH, UNAM: Ciudad de México, Mexico, 2014; pp. 95–115. [Google Scholar]
- Haro-Luna, M.X.; Ruan-Soto, F.; Guzmán-Dávalos, L. Traditional knowledge, uses, and perceptions of mushrooms among the Wixaritari and mestizos of Villa Guerrero, Jalisco, Mexico. IMA Fungus 2019, 10, 16. [Google Scholar] [CrossRef]
- Díaz-Larrea, J.; Pedroche, F.F.; Sentíes, A. Impacto de los estudios moleculares en la ficología comparada mexicana: El caso de las macroalgas marinas. Cymbella 2016, 2, 1–15. [Google Scholar]
- Novelo, E.; Tavera, R. Un panorama gráfico de las algas de agua dulce de México. Hidrobiológica 2011, 21, 333–341. [Google Scholar]
- Guzmán del Próo, S.A. Desarrollo y perspectivas de la explotación de algas marinas en México. Cienc. Pesq. 1993, 9, 129–136. [Google Scholar]
- Zertuche-Gonzalez, J.A. Situación actual del cultivo de algas agarofitas en América Latina y el Caribe. In Situación Actual de la Industria de Macroalgas Productoras Ficocoloides en America Latina y el Caribe; FAO: Mexico City, Mexico, 1993. [Google Scholar]
- Zertuche-González, J.A. Situación actual de la industria de las algas marinas productoras de ficocolides en México. In Situación Actual de la Industria de Macroalgas Productoras Ficocoloides en America Latina y el Caribe; FAO: Mexico City, Mexico, 1993. [Google Scholar]
- Aguilar-Rosas, R.; Espinoza-Avalos, J. Uso de las algas marinas en México. Cienc. Desarro. 1998, 143, 66–73. [Google Scholar]
- Ahmadi, A.; Zorofchian Moghadamtousi, S.; Abubakar, S.; Zandi, K. Antiviral potential of algae polysaccharides isolated from marine sources: A review. Biomed Res. Int. 2015, 2015, 825203. [Google Scholar] [CrossRef] [Green Version]
- Chitwood, D.J. Phytochemical based strategies for nematode control. Annu. Rev. Phytopathol. 2002, 40, 221–249. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.M.; Gloyna, E.F. Bactericidal Effects of Algae on Enteric Organisms, 1st ed.; California, U., Ed.; US Federal Water Quality Administration: San Diego, CA, USA, 1970.
- El-Ansary, M.S.M.; Hamouda, R.A. Biocontrol of root-knot nematode infected banana plants by some marine algae. Russ. J. Mar. Biol. 2014, 40, 140–146. [Google Scholar] [CrossRef]
- Lefranc, F.; Koutsaviti, A.; Ioannou, E.; Kornienko, A.; Roussis, V.; Kiss, R.; Newman, D. Algae metabolites: From in vitro growth inhibitory effects to promising anticancer activity. Nat. Prod. Rep. 2019, 36, 810–841. [Google Scholar] [CrossRef] [PubMed]
- Namvar, F.; Baharara, J.; Mahdi, A.A. Antioxidant and anticancer activities of selected Persian Gulf algae. Indian J. Clin. Biochem. 2014, 29, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Pesando, D.; Caram, B. Screening of marine algae from the French Mediterranean coast for antibacterial and antifungal activity. Bot. Mar. 1984, 27, 381–386. [Google Scholar] [CrossRef]
- Richards, J.T.; Kern, E.R.; Glasgow, L.A.; Overall Jr, J.C.; Deign, E.F.; Hatch, M.T. Antiviral activity of extracts from marine algae. Antimicrob. Agents Chemother. 1978, 14, 24–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjeewa, K.K.A.; Lee, J.-S.; Kim, W.-S.; Jeon, Y.-J. The potential of brown-algae polysaccharides for the development of anticancer agents: An update on anticancer effects reported for fucoidan and laminaran. Carbohydr. Polym. 2017, 177, 451–459. [Google Scholar] [CrossRef]
- Sheih, I.-C.; Fang, T.J.; Wu, T.-K.; Lin, P.-H. Anticancer and antioxidant activities of the peptide fraction from algae protein waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar] [CrossRef]
- Sunda, W.; Kieber, D.J.; Kiene, R.P.; Huntsman, S. An antioxidant function for DMSP and DMS in marine algae. Nature 2002, 418, 317–320. [Google Scholar] [CrossRef]
- Wan-Loy, C.; Siew-Moi, P. Marine algae as a potential source for anti-obesity agents. Mar. Drugs 2016, 14, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witvrouw, M.; De Clercq, E. Sulfated polysaccharides extracted from sea algae as potential antiviral drugs. Gen. Pharmacol. Vasc. Syst. 1997, 29, 497–511. [Google Scholar] [CrossRef]
- Nuño, T.I.; Enciso, I.; Hernández, R.M. Uso potencial de algas marinas presentes en el litoral de Jalisco. Méx. e-CUCBA 2017, 4, 5–17. [Google Scholar]
- Ianora, A.; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2010, 19, 493–511. [Google Scholar] [CrossRef]
- Saide, A.; Martínez, K.A.; Ianora, A.; Lauritano, C. Unlocking the health potential of microalgae as sustainable sources of bioactive compounds. Int. J. Mol. Sci. 2021, 22, 4383. [Google Scholar] [CrossRef] [PubMed]
- Basurto Peña, F. El Tecuítlatl o Espirulina (Arthrospira maxima Setchell & Gardner): Alimento Prehispánico con Potencial al Futuro. Etnoficología Apl Estud Casos en Relación a la Salud y la Aliment en Ambient Rural y Urbanos San Salvador Jujuy Cyted–Programa Iberoam Cienc y Tecnol para el Desarro; RISAPRET-CYTED: San Salvador de Jujuy, Argentina, 2009; pp. 43–68. [Google Scholar]
- Costa, J.A.V.; Freitas, B.C.B.; Rosa, G.M.; Moraes, L.; Morais, M.G.; Mitchell, B.G. Operational and economic aspects of Spirulina-based biorefinery. Bioresour. Technol. 2019, 292, 121946. [Google Scholar] [CrossRef] [PubMed]
- González, J. Las Algas de México; UNAM: Ciudad de México, Mexico, 1987; Volume 10. [Google Scholar]
- De Rivera, C.G.; Miranda-Zamora, R.; Diaz-Zagoya, J.C.; Juárez-Oropeza, M.A. Preventive effect of Spirulina maxima on the fatty liver induced by a fructose-rich diet in the rat, a preliminary report. Life Sci. 1993, 53, 57–61. [Google Scholar] [CrossRef]
- Torres-Durán, P.V.; Miranda-Zamora, R.; Paredes-Carbajal, M.C.; Mascher, D.; Díaz-Zagoya, J.C.; Juárez-Oropeza, M.A. Spirulina maxima prevents induction of fatty liver by carbon tetrachloride in the rat. IUBMB Life 1998, 44, 787–793. [Google Scholar] [CrossRef]
- Chamorro, G.; Pérez-Albiter, M.; Serrano-García, N.; Mares-Sámano, J.J.; Rojas, P. Spirulina maxima pretreatment partially protects against 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine neurotoxicity. Nutr. Neurosci. 2006, 9, 207–212. [Google Scholar] [CrossRef]
- Cervantes-Urieta, V.A.; Pérez-Castro, D.; Galeana-Parra, M.A.; Ramírez-Fuentes, E.; Trujillo-Tapia, M.N. Cultivation and biochemical composition of marine diatoms (Bacillariophyta) of Santa Lucía Bay, Acapulco, Mexico. Gayana Bot. 2020, 77, 11–22. [Google Scholar] [CrossRef]
- Freile-Pelegrín, Y.; Morales, J.L. Antibacterial activity in marine algae from the coast of Yucatan, Mexico. Bot. Mar. 2004, 47, 140–146. [Google Scholar] [CrossRef]
- Nuñez Resendiz, M.L.; Dreckmann, K.M.; Senties, A.; Wynne, M.J.; Tejera, H.L. Marine red algae (Rhodophyta) of economic use in the algal drifts from the Yucatan Peninsula, Mexico. Phytotaxa 2019, 387, 219–240. [Google Scholar] [CrossRef]
- Freile-Pelegrín, Y.; Robledo, D.; Azamar, J.A. Carrageenan of Eucheuma isiforme (Solieriaceae, Rhodophyta) from Yucatán, Mexico. I. Effect of extraction conditions. Bot. Mar. 2006, 49, 65–71. [Google Scholar] [CrossRef]
- Robledo, D.; Freile-Pelegrín, Y. Prospects for the cultivation of economically important carrageenophytes in Southeast Mexico. J. Appl. Phycol. 2011, 23, 415–419. [Google Scholar] [CrossRef]
- Huerta, L.; Chávez, M.L. Presencia de vitamina B12 en algunas algas marinas de las costas de México. An. Esc. Nac. Ciencias Biológicas 1968, 15, 9–22. [Google Scholar]
- Canales López, B. Enzimas-algas: Posibilidades de su uso para estimular la producción agrícola y mejorar los suelos. Terra Latinoam. 1999, 17, 271–276. [Google Scholar]
- Flores-Gallegos, A.C.; Castillo-Reyes, F.; Lafuente, C.B.; Loyola-Licea, J.C.; Reyes-Valdés, M.H.; Aguilar, C.N.; Herrera, R.R. Invertase production by Aspergillus and Penicillium and sequencing of an inv gene fragment. Micol. Apl. Int. 2012, 24, 1–10. [Google Scholar]
- Veana, F.; Fuentes-Garibay, J.A.; Aguilar, C.N.; Rodríguez-Herrera, R.; Guerrero-Olazarán, M.; Viader-Salvadó, J.M. Gene encoding a novel invertase from a xerophilic Aspergillus niger strain and production of the enzyme in Pichia pastoris. Enzyme Microb. Technol. 2014, 63, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Veana, F.; Martínez-Hernández, J.L.; Aguilar, C.N.; Rodríguez-Herrera, R.; Michelena, G. Utilization of molasses and sugar cane bagasse for production of fungal invertase in solid state fermentation using Aspergillus niger GH1. Braz. J. Microbiol. 2014, 45, 373–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belmares, R.; Contreras-Esquivel, J.C.; Rodríguez-Herrera, R.; Coronel, A.R.; Aguilar, C.N. Microbial production of tannase: An enzyme with potential use in food industry. LWT—Food Sci. Technol. 2004, 37, 857–864. [Google Scholar] [CrossRef]
- Borrego-Terrazas, J.A.; Lara-Victoriano, F.; Flores-Gallegos, A.C.; Veana, F.; Aguilar, C.N.; Rodríguez-Herrera, R. Nucleotide and amino acid variations of tannase gene from different Aspergillus strains. Can. J. Microbiol. 2014, 60, 509–516. [Google Scholar] [CrossRef]
- Camacho, N.A.; Aguilar, O.G. Production, purification, and characterization of a low-molecular-mass xylanase from Aspergillus sp. And its application in baking. Appl. Biochem. Biotechnol.—Part A Enzym. Eng. Biotechnol. 2003, 104, 159–171. [Google Scholar] [CrossRef]
- Peraza, L.; Ortiz, M.A.; Peberdy, J.F.; Aguilar, G. Growth and pectinase production by Aspergillus Mexican strain protoplast regenerated under acidic stress. Appl. Biochem. Biotechnol. 2003, 111, 15–27. [Google Scholar] [CrossRef]
- Martínez Jiménez, M.; Bahena, S.M.; Espinoza, C.; Trigos, Á. Isolation, characterization, and production of red pigment from Cercospora piaropi a biocontrol agent for waterhyacinth. Mycopathologia 2010, 169, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Kuyama, S.; Tamura, T. Cercosporin. A pigment of Cercosporina kikuchii Matsumoto et Tomoyasu. I. Cultivation of fungus, isolation and purification of pigment. J. Am. Chem. Soc. 1957, 79, 5725–5726. [Google Scholar] [CrossRef]
- Lousberg, R.J.J.C.; Weiss, U.; Salemink, C.A.; Arnone, A.; Merlini, L.; Nasini, G. The structure of cercosporin, a naturally occurring quinone. J. Chem. Soc. Chem. Commun. 1971, 22, 1463–1464. [Google Scholar] [CrossRef]
- Ojeda-Chi, M.M.; Rodriguez-Vivas, R.I.; Galindo-Velasco, E.; Lezama-Gutiérrrez, R. Laboratory and field evaluation of Metarhizium anisopliae (Deuteromycotina: Hyphomycetes) for the control of Rhipicephalus microplus (Acari: Ixodidae) in the Mexican tropics. Vet. Parasitol. 2010, 170, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Polar, P.; Moore, D.; Kairo, M.T.K.; Ramsubhag, A. Topically applied myco-acaricides for the control of cattle ticks: Overcoming the challenges. Exp. Appl. Acarol. 2008, 46, 119–148. [Google Scholar] [CrossRef]
- Fernández-Salas, A.; Alonso-Díaz, M.A.; Alonso-Morales, R.A.; Lezama-Gutiérrez, R.; Rodríguez-Rodríguez, J.C.; Cervantes-Chávez, J.A. Acaricidal activity of Metarhizium anisopliae isolated from paddocks in the Mexican tropics against two populations of the cattle tick Rhipicephalus microplus. Med. Vet. Entomol. 2017, 31, 36–43. [Google Scholar] [CrossRef]
- Vazquez-Rodriguez, A.; Vasto-Anzaldo, X.G.; Barboza Perez, D.; Vázquez-Garza, E.; Chapoy-Villanueva, H.; García-Rivas, G.; Garza-Cervantes, J.A.; Gómez-Lugo, J.J.; Gomez-Loredo, A.E.; Garza Gonzalez, M.T.; et al. Microbial competition of Rhodotorula mucilaginosa UANL-001L and E. coli increase biosynthesis of non-toxic exopolysaccharide with applications as a wide-spectrum antimicrobial. Sci. Rep. 2018, 8, 798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Cortázar, M.; Sánchez, J.E.; Huicochea-Medina, M.; Hernández-Velázquez, V.M.; Mendoza-De-Gives, P.; Zamilpa, A.; López-Arellano, M.E.; Pineda-Alegriá, J.A.; Aguilar-Marcelino, L. In vitro and in vivo nematicide effect of extract fractions of Pleurotus djamor against Haemonchus contortus. J. Med. Food 2021, 24, 310–318. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Yang, Z.; Liu, J.J.; Ueno, H.; Nantermet, P.G.; Guy, R.K.; Claiborne, C.F.; Renaud, J.; Couladouros, E.A.; Paulvannan, K.; et al. Total synthesis of taxol. Nature 1994, 367, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Holton, R.A.; Somoza, C.; Kim, H.B.; Liang, F.; Biediger, R.J.; Boatman, P.D.; Shindo, M.; Smith, C.C.; Kim, S.; Nadizadeh, H.; et al. First total synthesis of taxol. 1. Functionalization of the B ring. J. Am. Chem. Soc. 1994, 116, 1597–1598. [Google Scholar] [CrossRef]
- Ruiz-Sanchez, J.; Flores-Bustamante, Z.R.; Dendooven, L.; Favela-Torres, E.; Soca-Chafre, G.; Galindez-Mayer, J.; Flores-Cotera, L.B. A comparative study of Taxol production in liquid and solid-state fermentation with Nigrospora sp. a fungus isolated from Taxus globosa. J. Appl. Microbiol. 2010, 109, 2144–2150. [Google Scholar] [CrossRef]
- Ramos-Ligonio, A.; López-Monteon, A.; de la Soledad Lagunes-Castro, M.; Suárez-Medellín, J.; Espinoza, C.; Mendoza, G.; Trigos, Á. In vitro expression of toll-like receptors and proinflammatory molecules induced by ergosta-7,22-dien-3-one isolated from a wild Mexican strain of Ganoderma oerstedii (Agaricomycetes). Int. J. Med. Mushrooms 2017, 19, 203–211. [Google Scholar] [CrossRef]
- Sanodiya, B.; Thakur, G.; Baghel, R.; Prasad, G.; Bisen, P. Ganoderma lucidum: A potent pharmacological macrofungus. Curr. Pharm. Biotechnol. 2009, 10, 717–742. [Google Scholar] [CrossRef]
- Pan, D.; Zhang, D.; Wu, J.; Chen, C.; Xu, Z.; Yang, H.; Zhou, P. Antidiabetic, antihyperlipidemic and antioxidant activities of a novel proteoglycan from Ganoderma lucidum Fruiting Bodies on db/db mice and the possible mechanism. PLoS ONE 2013, 8, e0068332. [Google Scholar] [CrossRef]
- Romero-Córdoba, S.L.; Salido-Guadarrama, I.; Meneses, M.E.; Cosentino, G.; Iorio, M.V.; Tagliabue, E.; Torres, N.; Sánchez-Tapia, M.; Bonilla, M.; Castillo, I.; et al. Mexican Ganoderma lucidum extracts decrease lipogenesis modulating transcriptional metabolic networks and gut microbiota in C57BL/6 mice fed with a high-cholesterol diet. Nutrients 2021, 13, 38. [Google Scholar] [CrossRef] [PubMed]
- Meneses, M.E.; Martínez-Carrera, D.; Torres, N.; Sánchez-Tapia, M.; Aguilar-López, M.; Morales, P.; Sobal, M.; Bernabé, T.; Escudero, H.; Granados-Portillo, O.; et al. Hypocholesterolemic properties and prebiotic effects of Mexican Ganoderma lucidum in C57BL/6 mice. PLoS ONE 2016, 11, e0159631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Rojas, D.A.; Meneses, M.E.; Martínez-Carrera, D.; Figueroa-Cárdenas, J.D.; Sánchez-Medina, M.A.; Bonilla-Quintero, M.; Petlacalco-Sánchez, B.; Martínez-Gutiérrez, G.A.; Pérez-Herrera, A. Effect of Pleurotus agaves mushroom addition on the physicochemical and sensory properties of blue maize tortillas produced with traditional and ecological nixtamalization. Food Funct. 2020, 11, 8768–8779. [Google Scholar] [CrossRef]
- Castaneda de Leon, V.T.; Martínez Carrera, D.; Morales Almora, P.; Sobal Cruz, M.; Gil Munoz, A.; Leal Lara, H. El cuitlacoche, fruto de la interaccion Ustilago maydis-maíz, una aportacion de Mexico al mundo, en el sistema agroalimentario microbiano. In Ciencia, Tecnología e Innovación en el Sistema Agroalimentario de México: Hacia un Enfoque Integral de la Producción, la Dieta, la Salud y la Cultura en Beneficio de la Sociedad; Biblioteca Básica de Agricultura: Texcoco, Mexico, 2016; p. 856. [Google Scholar]
- Castañeda de León, V.; Martínez-Carrera, D.; Morales, P.; Sobal, M.; Gil-Muñoz, A.; Severiano-Pérez, P.; Leal-Lara, H. Productivity and flavor of diverse genotypes of Ustilago maydis “cuitlacoche” for human consumption. Fungal Biol. 2019, 123, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Huitron, C.; Perez, R.; Sanchez, A.E.; Lappe, P.; Rocha Zavaleta, L. Agricultural waste from the tequila industry as substrate for the production of commercially important enzymes. J. Environ. Biol. 2008, 29, 37–41. [Google Scholar]
- Levin, L.N.; Hernández-Luna, C.E.; Niño-Medina, G.; García-Rodríguez, J.P.; López-Sadin, I.; Méndez-Zamora, G.; Gutiérrez-Soto, G. Decolorization and detoxification of synthetic dyes by mexican strains of Trametes sp. Int. J. Environ. Res. Public Health 2019, 16, 4610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, C.F.; Arcuri, S.L.; Campos, C.R.; Vilela, D.M.; Alves, J.G.L.F.; Schwan, R.F. Using the residue of spirit production and bio-ethanol for protein production by yeasts. Waste Manag. 2011, 31, 108–114. [Google Scholar] [CrossRef]
- dos Reis, K.C.; Arrizon, J.; Amaya-Delgado, L.; Gschaedler, A.; Schwan, R.F.; Silva, C.F. Volatile compounds flavoring obtained from Brazilian and Mexican spirit wastes by yeasts. World J. Microbiol. Biotechnol. 2018, 34, 152. [Google Scholar] [CrossRef]
- Bogenschutz, M.P.; Ross, S. Therapeutic applications of classic hallucinogens. Curr. Top. Behav. Neurosci. 2018, 36, 361–391. [Google Scholar]
- Laure Vidriales, A.; Hannon Ovies, D. Psychedelic tourism in Mexico, a thriving trend. PASOS. Rev. Tur. Patrim. Cult. 2018, 16, 1037–1050. [Google Scholar] [CrossRef]
- Caron, D.A.; Worden, A.Z.; Countway, P.D.; Demir, E.; Heidelberg, K.B. Protists are microbes too: A perspective. ISME J. 2009, 3, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Vallesi, A.; Pucciarelli, S.; Buonanno, F.; Fontana, A.; Mangiagalli, M. Bioactive molecules from protists: Perspectives in biotechnology. Eur. J. Protistol. 2020, 75, 125720. [Google Scholar] [CrossRef] [PubMed]
- Rabe, P.; Rinkel, J.; Nubbemeyer, B.; Köllner, T.G.; Chen, F.; Dickschat, J.S. Terpene cyclases from social amoebae. Angew. Chem. Int. Ed. 2016, 55, 15420–15423. [Google Scholar] [CrossRef] [PubMed]
- Wohlleben, W.; Mast, Y.; Stegmann, E.; Ziemert, N. Antibiotic drug discovery. Microb. Biotechnol. 2016, 9, 541–548. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Obeso Fernandez Del Valle, A.; Scheckhuber, C.Q. From Past to Present: Biotechnology in Mexico Using Algae and Fungi. Plants 2021, 10, 2530. https://doi.org/10.3390/plants10112530
De Obeso Fernandez Del Valle A, Scheckhuber CQ. From Past to Present: Biotechnology in Mexico Using Algae and Fungi. Plants. 2021; 10(11):2530. https://doi.org/10.3390/plants10112530
Chicago/Turabian StyleDe Obeso Fernandez Del Valle, Alvaro, and Christian Quintus Scheckhuber. 2021. "From Past to Present: Biotechnology in Mexico Using Algae and Fungi" Plants 10, no. 11: 2530. https://doi.org/10.3390/plants10112530
APA StyleDe Obeso Fernandez Del Valle, A., & Scheckhuber, C. Q. (2021). From Past to Present: Biotechnology in Mexico Using Algae and Fungi. Plants, 10(11), 2530. https://doi.org/10.3390/plants10112530