
Effects of Low Temperature, Nitrogen Starvation and Their Combination on the Photosynthesis and Metabolites of Thermosynechococcus E542: A Comparison Study


 ,
, 

Abstract
:1. Introduction
2. Results
2.1. Effects of Temperature, Nitrogen Starvation, and Their Combination on the Growth
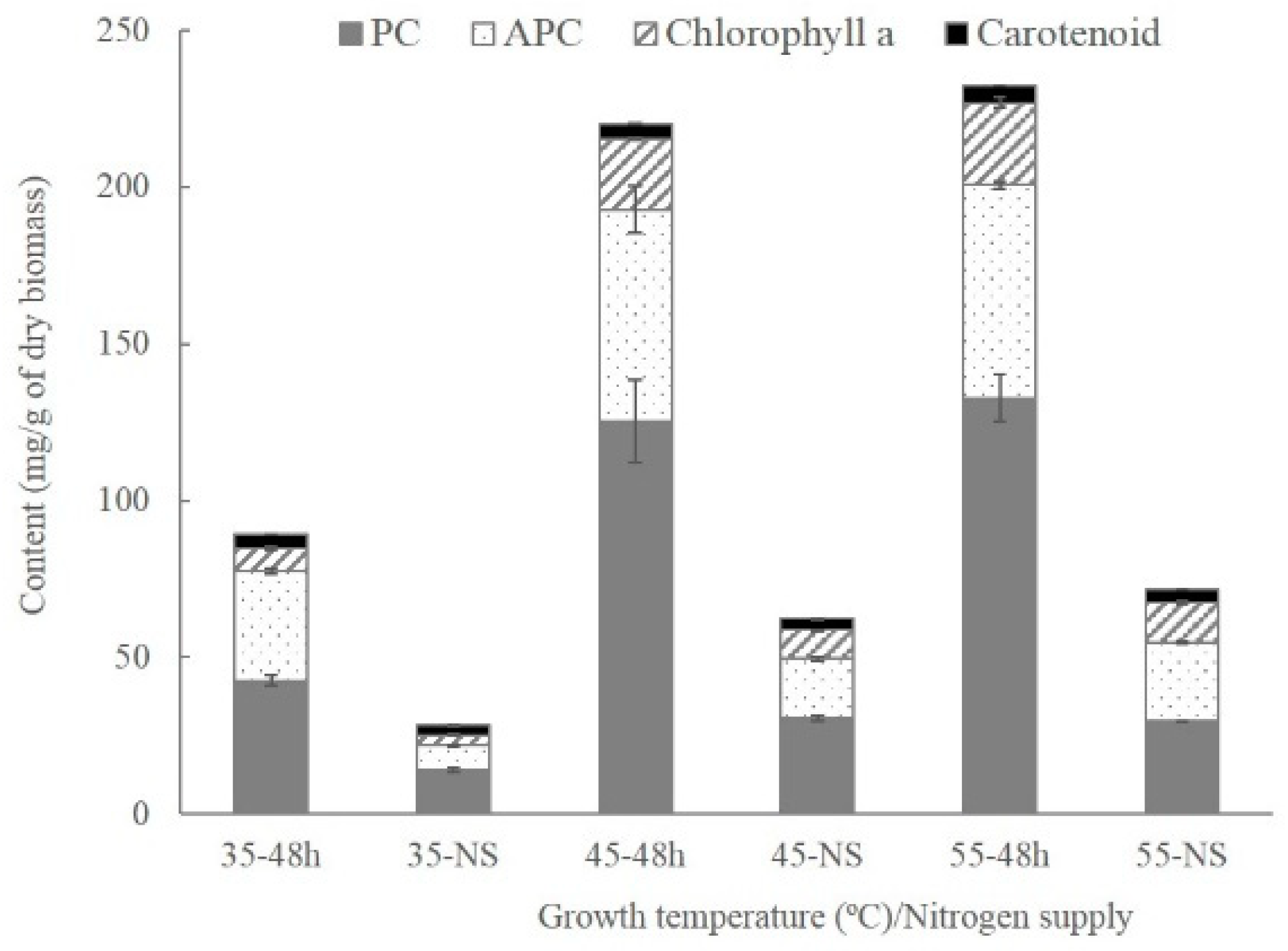
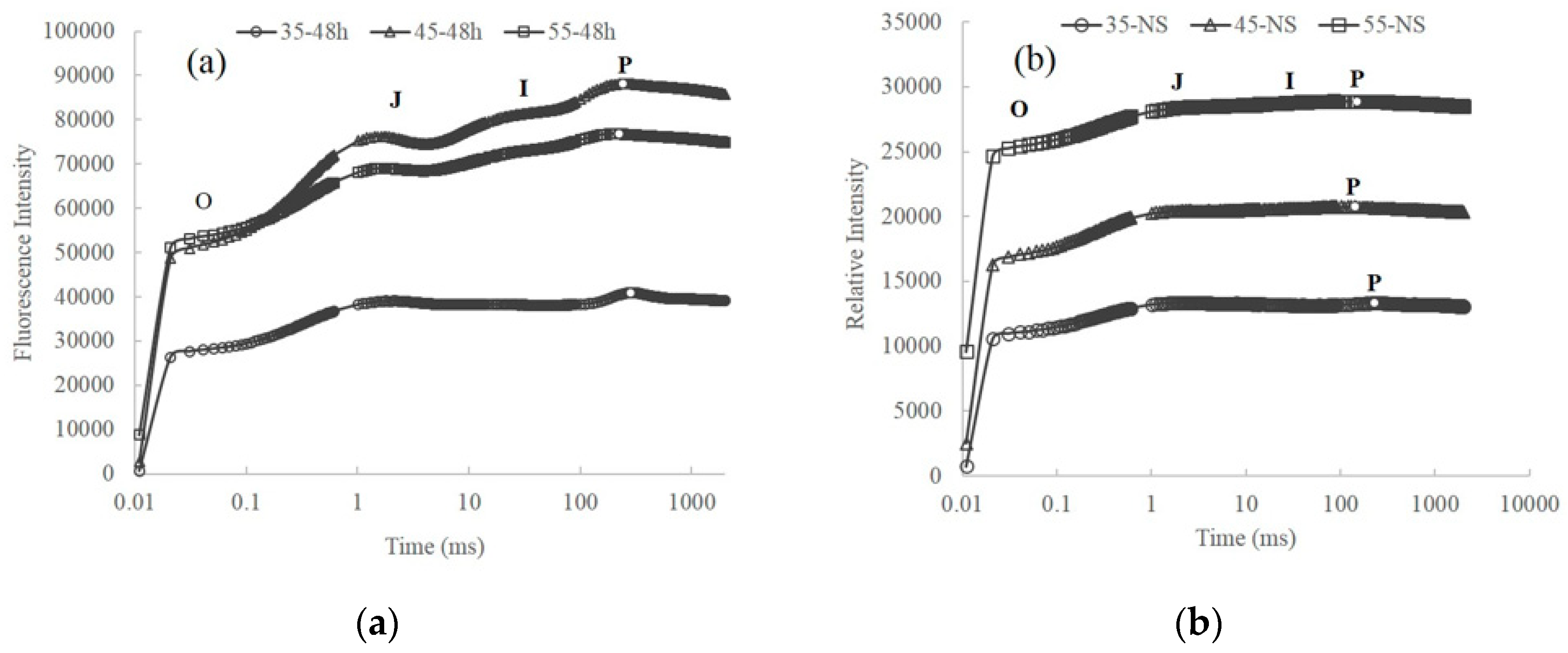
2.2. Variation of Pigment Contents and Chlorophyll Fluorescence
2.3. Variation of Alkanes and Fatty Acids
2.4. Lipidomic Analysis Based on LC-MS/MS
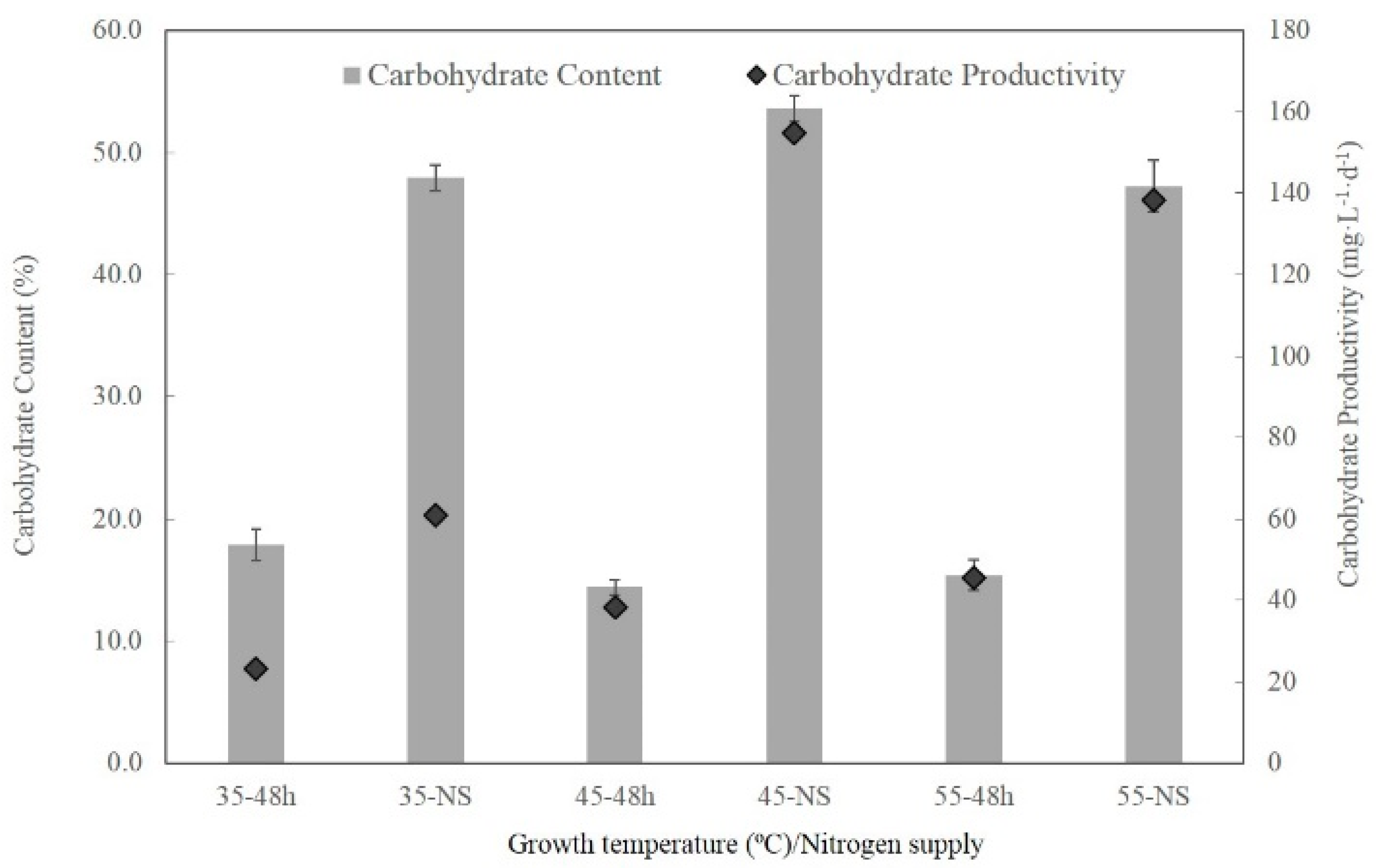
2.5. Content and Productivity of Carbohydrate and Elemental Composition of the Biomass
3. Discussion
4. Materials and Methods
4.1. Cultivation of Cyanobacterium
4.2. Chla Fluorescence Transient Analysis
4.3. Measurement of Pigments
4.4. Determination of FAMEs and Alkane
4.5. Lipidomic Analysis
4.6. Determination of Carbohydrates and Elemental Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, J.; Liang, Y.; Jiang, D.; Li, L.; Luo, Y.; Shah, M.M.R.; Daroch, M. Temperature-controlled Thermophilic Bacterial Communities in Hot Springs of Western Sichuan, China. BMC Microbiol. 2018, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.-H.; Liu, H.-L.; Chen, Y.-J.; Cheng, H.; Lin, W.-L.; Yeh, C.-H.; Chang, C.-H. Enhancing CO2 Bio-mitigation by Genetic Engineering of Cyanobacteria. Energy Environ. Sci. 2012, 5, 8318–8327. [Google Scholar] [CrossRef]
- Liang, Y.; Kaczmarek, M.B.; Kasprzak, A.K.; Tang, J.; Shah, M.M.R.; Jin, P.; Klepacz-Smolka, A.; Cheng, J.J.; Ledakowicz, S.; Daroch, M. Thermosynechococcaceae as a Source of Thermostable C-phycocyanins: Properties and Molecular Insights. Algal Res. Biomass Biofuels Bioprod. 2018, 35, 223–235. [Google Scholar] [CrossRef]
- Forchhammer, K.; Schwarz, R. Nitrogen Chlorosis in Unicellular Cyanobacteria—A Developmental Program for Surviving Nitrogen Deprivation. Environ. Microbiol. 2019, 21, 1173–1184. [Google Scholar] [CrossRef] [Green Version]
- Huner, N.P.A.; Oquist, G.; Sarhan, F. Energy Balance and Acclimation to Light and Cold. Trends Plant Sci. 1998, 3, 224–230. [Google Scholar] [CrossRef]
- Mackey, K.R.M.; Paytan, A.; Caldeira, K.; Grossman, A.R.; Moran, D.; McIlvin, M.; Saito, M.A. Effect of Temperature on Photosynthesis and Growth in Marine Synechococcus spp. Plant Physiol. 2013, 163, 815–829. [Google Scholar] [CrossRef] [Green Version]
- Nikolova, D.; Weber, D.; Scholz, M.; Bald, T.; Scharsack, J.P.; Hippler, M. Temperature-Induced Remodeling of the Photosynthetic Machinery Tunes Photosynthesis in the Thermophilic Alga Cyanidioschyzon merolae. Plant Physiol. 2017, 174, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikami, K.; Murata, N. Membrane Fluidity and the Perception of Environmental Signals in Cyanobacteria and Plants. Prog. Lipid Res. 2003, 42, 527–543. [Google Scholar] [CrossRef]
- Schwarz, R.; Forchhammer, K. Acclimation of Unicellular Cyanobacteria to Macronutrient Deficiency: Emergence of a Complex Network of Cellular Responses. Microbiology 2005, 151, 2503–2514. [Google Scholar] [CrossRef] [Green Version]
- Maranon, E.; Lorenzo, M.P.; Cermeno, P.; Mourino-Carballido, B. Nutrient Limitation Suppresses the Temperature Dependence of Phytoplankton Metabolic Rates. ISME J. 2018, 12, 1836–1845. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Tang, J.; Luo, Y.; Kaczmarek, M.B.; Li, X.; Daroch, M. Thermosynechococcus as a Thermophilic Photosynthetic Microbial Cell Factory for CO2 Utilisation. Bioresour. Technol. 2019, 278, 255–265. [Google Scholar] [CrossRef]
- Tang, J.; Jiang, D.; Luo, Y.; Liang, Y.; Li, L.; Shah, M.M.R.; Daroch, M. Potential New Genera of Cyanobacterial Strains Isolated from Thermal Springs of Western Sichuan, China. Algal Res. Biomass Biofuels Bioprod. 2018, 31, 14–20. [Google Scholar] [CrossRef]
- Vonshak, A.; Novoplansky, N. Acclimation to Low Temperature of Two Arthrospira platensis (Cyanobacteria) Strains Involves Down-regulation of PSII and Improved Resistance to Photoinhibition. J. Phycol. 2008, 44, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Tsimilli-Michael, M.; Stamatakis, K.; Papageorgiou, G.C. Dark-to-light Transition in Synechococcus sp PCC 7942 Cells Studied by Fluorescence Kinetics Assesses Plastoquinone Redox Poise in the Dark and Photosystem II Fluorescence Component and Dynamics during State 2 to State 1 Transition. Photosynth. Res. 2009, 99, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.; Hurry, V.; Clarke, A.K.; Gustafsson, P.; Oquist, G. Chlorophyll Fluorescence Analysis of Cyanobacterial Photosynthesis and Acclimation. Microbiol. Mol. Biol. Rev. 1998, 62, 667–683. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Vonshak, A. Effects of Salinity Stress on Photosystem II Function in Cyanobacterial Spirulina platensis Cells. Physiol. Plant. 2002, 114, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Gong, H.; Wen, X.; Lu, C. Salt Stress Induces a Decrease in Excitation Energy Transfer from Phycobilisomes to Photosystem II but an Increase to Photosystem I in the Cyanobacterium Spirulina platensis. J. Plant Physiol. 2010, 167, 951–958. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, J.; Gong, H.; Wen, X.; Ren, H.; Lu, C. Effects of Heat Stress on PSII Photochemistry in a Cyanobacterium Spirulina platensis. Plant Sci. 2008, 175, 556–564. [Google Scholar] [CrossRef]
- Wada, H.; Murata, N. Temperature-induced changes in the Fatty-acid Composition of the Cyanobacterium, Synechocystis PCC6803. Plant Physiol. 1990, 92, 1062–1069. [Google Scholar] [CrossRef] [Green Version]
- Berla, B.M.; Saha, R.; Maranas, C.D.; Pakrasi, H.B. Cyanobacterial Alkanes Modulate Photosynthetic Cyclic Electron Flow to Assist Growth under Cold Stress. Sci. Rep. 2015, 5, 14894. [Google Scholar] [CrossRef]
- Shan, Y.; Liu, Y.; Yang, L.; Nie, H.; Shen, S.; Dong, C.; Bai, Y.; Sun, Q.; Zhao, J.; Liu, H. Lipid Profiling of Cyanobacteria Synechococcus sp PCC 7002 Using Two-dimensional Liquid Chromatography with Quadrupole Time-of-flight Mass Spectrometry. J. Sep. Sci. 2016, 39, 3745–3753. [Google Scholar] [CrossRef]
- Mavroudakis, L.; Valsami, E.-A.; Grafanaki, S.; Andreadaki, T.-P.; Ghanotakis, D.F.; Pergantis, S.A. The Effect of Nitrogen Starvation on Membrane Lipids of Synechocystis sp. PCC 6803 Investigated by Using Easy Ambient Sonic-spray Ionization Mass Spectrometry. Biochim. Biophys. Acta-Biomembr. 2019, 1861, 183027. [Google Scholar] [CrossRef]
- Pittera, J.; Jouhet, J.; Breton, S.; Garczarek, L.; Partensky, F.; Marechal, E.; Nguyen, N.A.; Dore, H.; Ratin, M.; Pitt, F.D.; et al. Thermoacclimation and Genome Adaptation of the Membrane Lipidome in Marine Synechococcus. Environ. Microbiol. 2018, 20, 612–631. [Google Scholar] [CrossRef] [Green Version]
- Collier, J.L.; Grossman, A.R. A Small Polypeptide Triggers Complete Degradation of Light-harvesting Phycobiliproteins in Nutrient-deprived Cyanobacteria. EMBO J. 1994, 13, 1039–1047. [Google Scholar] [CrossRef]
- Collier, J.L.; Grossman, A.R. Chlorosis Induced by Nutrient Deprivation in Synechococcus sp Strain PCC-7942-Not All Bleaching is the Same. J. Bacteriol. 1992, 174, 4718–4726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasunuma, T.; Kikuyama, F.; Matsuda, M.; Aikawa, S.; Izumi, Y.; Kondo, A. Dynamic Metabolic Profiling of Cyanobacterial Glycogen Biosynthesis under Conditions of Nitrate Depletion. J. Exp. Bot. 2013, 64, 2943–2954. [Google Scholar] [CrossRef] [Green Version]
- Chow, T.-J.; Su, H.-Y.; Tsai, T.-Y.; Chou, H.-H.; Lee, T.-M.; Chang, J.-S. Using Recombinant Cyanobacterium (Synechococcus elongatus) with Increased Carbohydrate Productivity as Feedstock for Bioethanol Production via Separate Hydrolysis and Fermentation Process. Bioresour. Technol. 2015, 184, 33–41. [Google Scholar] [CrossRef]
- Li, X.; Li, W.; Zhai, J.; Wei, H.; Wang, Q. Effect of Ammonium Nitrogen on Microalgal Growth, Biochemical Composition and Photosynthetic Performance in Mixotrophic Cultivation. Bioresour. Technol. 2019, 273, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Zavrel, T.; Sinetova, M.A.; Červený, J. Measurement of Chlorophyll a and Carotenoids Concentration in Cyanobacteria. Bio-Protocol 2015, 5, e1467. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, R.J. Consistent Sets of Spectrophotometric Chlorophyll Equations for Acetone, Methanol and Ethanol Solvents. Photosynth. Res. 2006, 89, 27–41. [Google Scholar] [CrossRef]
- Zavrel, T.; Chmelik, D.; Sinetova, M.A.; Cerveny, J. Spectrophotometric Determination of Phycobiliprotein Content in Cyanobacterium Synechocystis. Jove-J. Vis. Exp. 2018. [Google Scholar] [CrossRef] [Green Version]
- Van Wychen, S.; Laurens, L. Determination of Total Lipids as Fatty Acid Methyl Esters (FAME) by in Situ Transesterification. Contract 2013, 303, 275–3000. [Google Scholar]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, B. Determination of Structural Carbohydrates and Lignin in Biomass; Document # NREL/TP-510-42618; National Renewable Energy Laboratory: Golden, CO, USA, 2008.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomass –1st Stage (mgL−1) | Growth Rate μ –1st Stage (1 d−1) | Biomass –2nd Stage (mgL−1) | Growth Rate μ –2nd Stage (1 d−1) | |
|---|---|---|---|---|
| 35–48 h | 0.117 ± 0.008 | 1.29 ± 0.11 | 0.265 ± 0.01 | 0.82 ± 0.07 |
| 35–NS | 0.102 ± 0.005 | 1.32 ± 0.21 | 0.196 ± 0.009 | 0.65 ± 0.05 |
| 45–48 h | 0.254 ± 0.003 | 2.26 ± 0.08 | 0.591 ± 0.012 | 0.84 ± 0.02 |
| 45–NS | 0.253 ± 0.007 | 2.23 ± 0.17 | 0.605 ± 0.015 | 0.87 ± 0.03 |
| 55–48 h | 0.244 ± 0.004 | 2.16 ± 0.13 | 0.678 ± 0.017 | 1.02 ± 0.07 |
| 55–NS | 0.240 ± 0.004 | 2.19 ± 0.04 | 0.654 ± 0.018 | 1.00 ± 0.04 |
| FO | VJ | VI | FV/FM | |
|---|---|---|---|---|
| 35–48 h | 27,089 ± 186 1 | 0.853 ± 0.016 | 0.794 ± 0.013 | 0.321 ± 0.007 |
| 45–48 h | 48,923 ± 306 | 0.665 ± 0.007 | 0.811 ± 0.008 | 0.422 ± 0.008 |
| 55–48 h | 44,738 ± 510 | 0.661 ± 0.008 | 0.835 ± 0.010 | 0.344 ± 0.006 |
| 35–NS | 10,348 ± 181 | 0.992 ± 0.007 | 0.910 ± 0.006 | 0.180 ± 0.008 |
| 45–NS | 14,796 ± 170 | 0.912 ± 0.007 | 0.956 ± 0.009 | 0.203 ± 0.006 |
| 55–NS | 15,527 ± 421 | 0.853 ± 0.007 | 0.959 ± 0.006 | 0.181 ± 0.002 |
| 35 °C | 35 °C–NS | 55 °C | 55 °C–NS | |
|---|---|---|---|---|
| MGDG content 1 | 17.8 ± 0.64 3 | 12.2 ± 0.10 | 18.6 ± 1.4 | 13.8 ± 0.48 |
| MGDG (16:0/16:1) | 63.8 ± 2.2 | 47.1 ± 1.1 | 27.8 ± 0.9 | 21.2 ± 0.2 |
| MGDG (16:0/18:1) | 21.4 ± 0.2 | 35.7 ± 0.3 | 35.1 ± 1.3 | 34.6 ± 0.2 |
| Others 2 | 14.8 | 17.2 | 37.1 | 44.2 |
| DGDG content | 26.7 ± 0.26 | 19.1 ± 1.0 | 25.1 ± 0.66 | 21.1 ± 1.1 |
| DGDG (16:0/18:1) | 17.4 ± 0.3 | 25.4 ± 1.6 | 32.0 ± 0.5 | 36.3 ± 1.3 |
| DGDG (16:0/16:1) | 60.1 ± 1.5 | 50.9 ± 4.3 | 25.0 ± 0.5 | 19.2 ± 0.3 |
| DGDG (16:0/16:0) | 6.2 ± 0.1 | 5.9 ± 0.5 | 20.1 ± 0.3 | 14.7 ± 0.1 |
| Others | 16.3 | 17.9 | 22.9 | 29.9 |
| SQDG content | 3.70 ± 0.11 | 1.75 ± 0.01 | 2.32 ± 0.05 | 1.42 ± 0.07 |
| SQDG (16:0/16:0) | > 99 | |||
| PG content | 2.20 ± 0.07 | 1.44 ± 0.01 | 1.58 ± 0.01 | 0.94 ± 0.05 |
| PG (16:0/16:0) | 8.3 ± 0.1 | 7.2 ± 0.1 | 19.1 ± 0.5 | 17.5 ± 0.1 |
| PG (16:0/16:1) | 49.9 ± 1.2 | 42.5 ± 0.1 | 61.9 ± 0.4 | 66.2 ± 0.2 |
| PG (16:0/18:1) | 41.8 ± 1.1 | 50.3 ± 0.2 | 0 | 0 |
| PG (18:0/16:0) | 0 | 0 | 19.0 ± 0.2 | 16.4 ± 0.2 |
| Carbon | Hydrogen | Nitrogen | Sulphur | |
|---|---|---|---|---|
| 35–48 h | 39.40 ± 0.30 1 | 7.28 ± 0.29 | 8.09 ± 0.04 | 0.68 ± 0.01 |
| 35–NS | 38.77 ± 0.09 | 8.22 ± 0.29 | 3.92 ± 0.12 | 0.49 ± 0.01 |
| 45–48 h | 42.55 ± 0.55 | 8.86 ± 0.25 | 9.36 ± 0.18 | 1.03 ± 0.11 |
| 45–NS | 41.03 ± 0.76 | 10.20 ± 0.02 | 3.93 ± 0.09 | 0.66 ± 0.02 |
| 55–48 h | 44.05 ± 1.30 | 11.07 ± 0.01 | 8.95 ± 0.33 | 1.02 ± 0.02 |
| 55–NS | 42.92 ± 0.01 | 10.54 ± 0.11 | 4.28 ± 0.02 | 0.81 ± 0.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Liang, Y.; Li, K.; Jin, P.; Tang, J.; Klepacz-Smółka, A.; Ledakowicz, S.; Daroch, M. Effects of Low Temperature, Nitrogen Starvation and Their Combination on the Photosynthesis and Metabolites of Thermosynechococcus E542: A Comparison Study. Plants 2021, 10, 2101. https://doi.org/10.3390/plants10102101
Li X, Liang Y, Li K, Jin P, Tang J, Klepacz-Smółka A, Ledakowicz S, Daroch M. Effects of Low Temperature, Nitrogen Starvation and Their Combination on the Photosynthesis and Metabolites of Thermosynechococcus E542: A Comparison Study. Plants. 2021; 10(10):2101. https://doi.org/10.3390/plants10102101
Chicago/Turabian StyleLi, Xingkang, Yuanmei Liang, Kai Li, Peng Jin, Jie Tang, Anna Klepacz-Smółka, Stanislaw Ledakowicz, and Maurycy Daroch. 2021. "Effects of Low Temperature, Nitrogen Starvation and Their Combination on the Photosynthesis and Metabolites of Thermosynechococcus E542: A Comparison Study" Plants 10, no. 10: 2101. https://doi.org/10.3390/plants10102101
APA StyleLi, X., Liang, Y., Li, K., Jin, P., Tang, J., Klepacz-Smółka, A., Ledakowicz, S., & Daroch, M. (2021). Effects of Low Temperature, Nitrogen Starvation and Their Combination on the Photosynthesis and Metabolites of Thermosynechococcus E542: A Comparison Study. Plants, 10(10), 2101. https://doi.org/10.3390/plants10102101