
Introgression of Bacterial Blight Resistance Genes in the Rice Cultivar Ciherang: Response against Xanthomonas oryzae pv. oryzae in the F6 Generation

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Results
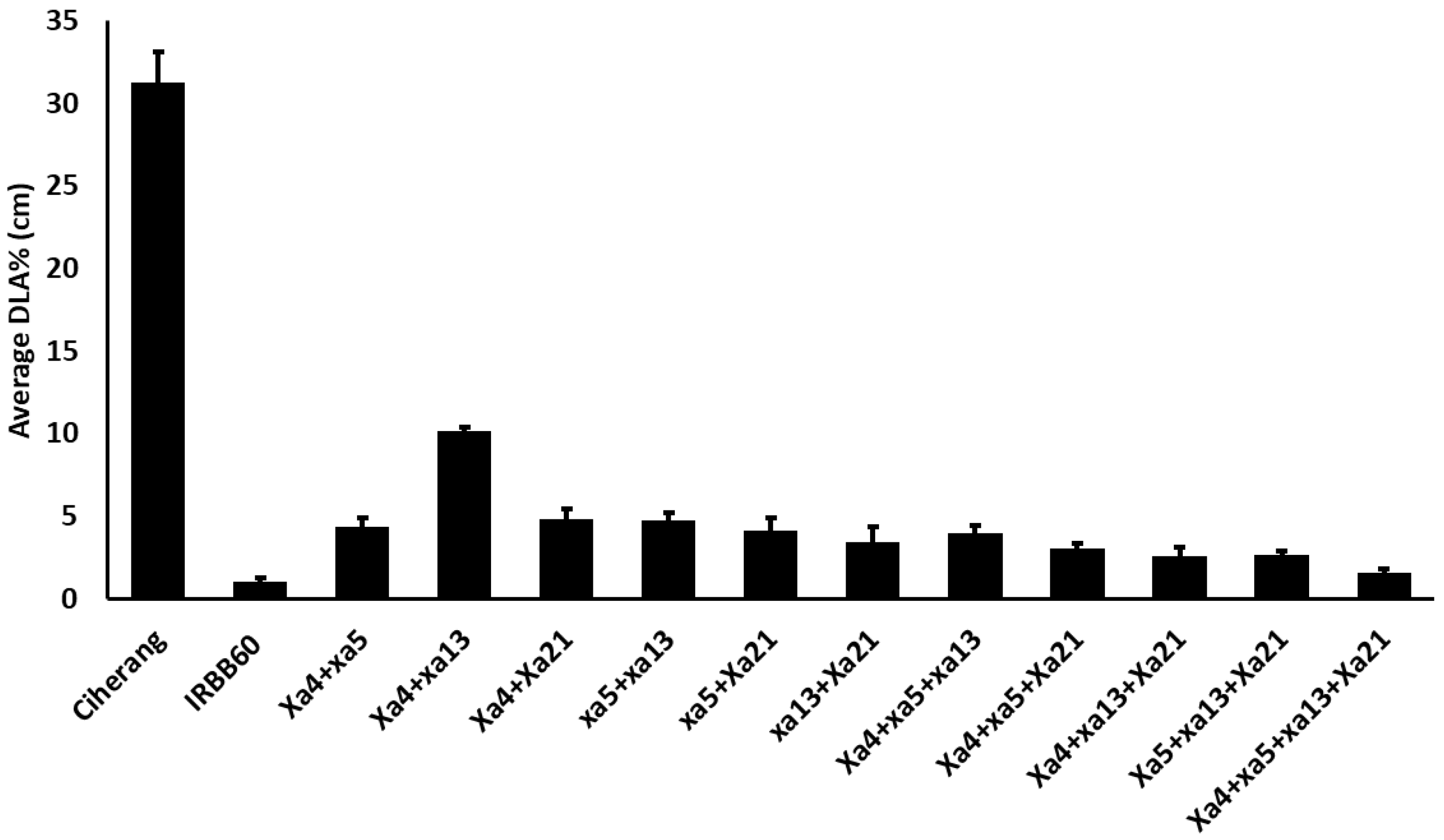
2.1. Response of RILs toward BB Races
2.2. Molecular Marker Analysis for Characterizing the RILs
2.3. Agronomic Performance of Pyramided Lines
2.4. Associations between the Number and Combinations of Pyramided Resistance Genes and Disease Reaction against Different BB Races
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Preparation of Inocula to Infect Rice Plants
4.3. Inoculation of Rice Plants with BB Races/Isolates
4.4. Assessment of Disease Response and Scoring
4.5. DNA Isolation and PCR Analysis
4.6. Data Collection and Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cottyn, B.; Debode, J.; Regalado, E.; Mew, T.; Swings, J. Phenotypic and genetic diversity of rice seed-associated bacteria and their role in pathogenicity and biological control. J. Appl. Microbiol. 2009, 107, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Virmani, S.S. Hybrid rice. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 1996; Volume 57, pp. 377–462. [Google Scholar]
- Fitzgerald, M.A.; McCouch, S.R.; Hall, R.D. Not just a grain of rice: The quest for quality. Trends Plant Sci. 2009, 14, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Ashikari, M.; Matsuoka, M. The role of QTLs in the breeding of high-yielding rice. Trends Plant Sci. 2011, 16, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Carriger, S.; Vallée, D. More crop per drop. Rice Today 2007, 6, 10–13. [Google Scholar]
- Peng, S.; Huang, J.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.; Centeno, G.S.; Khush, G.S.; Cassman, K.G. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971–9975. [Google Scholar] [CrossRef] [Green Version]
- Horino, O.; Siwi, B.H.; Miah, S.A.; Mew, T.W. Virulence of Xanthomonas campestris pv. oryzae isolated in Indonesia and Bangladesh. Jpn. J. Phytopathol. 1983, 49, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Mew, T.; Cruz, V.; Medalla, E. Changes in race frequency of Xanthomonas oryzae pv. oryzae in response to rice cultivars planted in the Philippines. Plant Dis. 1992, 76, 1029–1032. [Google Scholar] [CrossRef]
- Noh, T.-H.; Lee, D.-K.; Park, J.-C.; Shim, H.-K.; Choi, M.-Y.; Kang, M.-H.; Kim, J.-D. Effects of bacterial leaf blight occurrence on rice yield and grain quality in different rice growth stage. Res. Plant Dis. 2007, 13, 20–23. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A. Bacterial blight: Crop loss assessment and disease management. In Proceedings of the International Workshop on Bacterial Blight of Rice 1989; International Rice Research Institute: Manila, Philippines, 1989; pp. 79–88. [Google Scholar]
- Shin, M.; Shin, H.; Jun, B.; Choi, B. Effects of inoculation of compatible and incompatible bacterial blight races on grain yield and quality of two rice cultivars. Korean J. Breed. 1992, 24, 264–267. [Google Scholar]
- Schantz, S.L.; Gasior, D.M.; Polverejan, E.; McCaffrey, R.J.; Sweeney, A.M.; Humphrey, H.; Gardiner, J.C. Impairments of memory and learning in older adults exposed to polychlorinated biphenyls via consumption of Great Lakes fish. Environ. Health Perspect. 2001, 109, 605–611. [Google Scholar] [CrossRef]
- Huang, N.; Angeles, E.; Domingo, J.; Magpantay, G.; Singh, S.; Zhang, G.; Kumaravadivel, N.; Bennett, J.; Khush, G. Pyramiding of bacterial blight resistance genes in rice: Marker-assisted selection using RFLP and PCR. Theor. Appl. Genet. 1997, 95, 313–320. [Google Scholar] [CrossRef]
- Jena, K.; Mackill, D. Molecular markers and their use in marker-assisted selection in rice. Crop. Sci. 2008, 48, 1266–1276. [Google Scholar] [CrossRef]
- Singh, S.; Sidhu, J.; Huang, N.; Vikal, Y.; Li, Z.; Brar, D.; Dhaliwal, H.; Khush, G. Pyramiding three bacterial blight resistance genes (xa5, xa13 and Xa21) using marker-assisted selection into indica rice cultivar PR106. Theor. Appl. Genet. 2001, 102, 1011–1015. [Google Scholar] [CrossRef]
- Rao, S.R.; Priyanka, M.; Kumar, M.A.; Ramanaiah, C.; Yashwanth, B.; Mohan, K.M.; Chandra, B.V.; Venkateshwarlu, V.; Gandhi, D.; Rao, N.J.M. Marker-assisted breeding for bacterial blight resistance in parental lines of hybrid rice. J. Plant Pathol. 2017, 99, 691–701. [Google Scholar]
- Neelam, K.; Mahajan, R.; Gupta, V.; Bhatia, D.; Gill, B.K.; Komal, R.; Lore, J.S.; Mangat, G.S.; Singh, K. High-resolution genetic mapping of a novel bacterial blight resistance gene xa-45 (t) identified from Oryza glaberrima and transferred to Oryza sativa. Theor. Appl. Genet. 2020, 133, 689–705. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-M.; Reinke, R.F. A novel resistance gene for bacterial blight in rice, Xa43 (t) identified by GWAS, confirmed by QTL mapping using a bi-parental population. PLoS ONE 2019, 14, e0211775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-M. Identification of novel recessive gene xa44 (t) conferring resistance to bacterial blight races in rice by QTL linkage analysis using an SNP chip. Theor. Appl. Genet. 2018, 131, 2733–2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-M.; Suh, J.-P.; Qin, Y.; Noh, T.-H.; Reinke, R.F.; Jena, K.K. Identification and fine-mapping of a new resistance gene, Xa40, conferring resistance to bacterial blight races in rice (Oryza sativa L.). Theor. Appl. Genet. 2015, 128, 1933–1943. [Google Scholar] [CrossRef] [PubMed]
- Bhasin, H.; Bhatia, D.; Raghuvanshi, S.; Lore, J.S.; Sahi, G.K.; Kaur, B.; Vikal, Y.; Singh, K. New PCR-based sequence-tagged site marker for bacterial blight resistance gene Xa38 of rice. Mol. Breed. 2012, 30, 607–611. [Google Scholar] [CrossRef]
- Kumar, P.N.; Sujatha, K.; Laha, G.; Rao, K.S.; Mishra, B.; Viraktamath, B.; Hari, Y.; Reddy, C.; Balachandran, S.; Ram, T. Identification and fine-mapping of Xa33, a novel gene for resistance to Xanthomonas oryzae pv. oryzae. Phytopathology 2012, 102, 222–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.-Y.; Pi, L.-Y.; Wang, G.-L.; Gardner, J.; Holsten, T.; Ronald, P.C. Evolution of the rice Xa21 disease resistance gene family. Plant Cell 1997, 9, 1279–1287. [Google Scholar] [PubMed]
- Yang, D.; Sanchez, A.; Khush, G.; Zhu, Y.; Huang, N. Construction of a BAC contig containing the xa5 locus in rice. Theor. Appl. Genet. 1998, 97, 1120–1124. [Google Scholar] [CrossRef]
- Sun, X.; Yang, Z.; Wang, S.; Zhang, Q. Identification of a 47-kb DNA fragment containing Xa4, a locus for bacterial blight resistance in rice. Theor. Appl. Genet. 2003, 106, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.; Yang, B.; Tian, D.; Wu, L.; Wang, D.; Sreekala, C.; Yang, F.; Chu, Z.; Wang, G.-L.; White, F.F. R gene expression induced by a type-III effector triggers disease resistance in rice. Nature 2005, 435, 1122. [Google Scholar] [CrossRef]
- Nino-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef]
- Cheema, K.K.; Grewal, N.K.; Vikal, Y.; Sharma, R.; Lore, J.S.; Das, A.; Bhatia, D.; Mahajan, R.; Gupta, V.; Bharaj, T.S. A novel bacterial blight resistance gene from Oryza nivara mapped to 38 kb region on chromosome 4L and transferred to Oryza sativa L. Genet. Res. 2008, 90, 397–407. [Google Scholar] [CrossRef]
- Zhang, F.; Zhuo, D.L.; Huang, L.Y.; Wang, W.S.; Xu, J.L.; Vera Cruz, C.; Li, Z.K.; Zhou, Y.L. Xa39, a novel dominant gene conferring broad-spectrum resistance to Xanthomonas oryzae pv. oryzae in rice. Plant Pathol. 2015, 64, 568–575. [Google Scholar] [CrossRef]
- Sundaram, R.M.; Vishnupriya, M.R.; Biradar, S.K.; Laha, G.S.; Reddy, G.A.; Rani, N.S.; Sarma, N.P.; Sonti, R.V. Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 2008, 160, 411–422. [Google Scholar] [CrossRef]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Adhikari, T.B.; Cruz, C.; Zhang, Q.; Nelson, R.; Skinner, D.; Mew, T.; Leach, J. Genetic Diversity of Xanthomonas oryzae pv. oryzae in Asia. Appl. Environ. Microbiol. 1995, 61, 966–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arif, M.; Jaffar, M.; Babar, M.; Sheikh, M.A.; Kousar, S.; Arif, A.; Zafar, Y. Identification of bacterial blight resistance genes Xa4 in Pakistani rice germplasm using PCR. Afr. J. Biotechnol. 2008, 7, 541–545. [Google Scholar]
- Garris, A.J.; McCOUCH, S.R.; Kresovich, S. Population structure and its effect on haplotype diversity and linkage disequilibrium surrounding the xa5 locus of rice (Oryza sativa L.). Genetics 2003, 165, 759–769. [Google Scholar] [CrossRef]
- Sidhu, G.; Khush, G.S.; Mew, T. Genetic analysis of bacterial blight resistance in seventy-four cultivars of rice, Oryza sativa L. Theor. Appl. Genet. 1978, 53, 105–111. [Google Scholar] [CrossRef]
- Zhang, Q.; Ling, S.; Zhao, B.; Wang, C.; Yang, W.; Zhao, K.; Zhu, L.; Li, D.; Chen, C. A new gene for resistance to bacterial blight from Oryza rufipogon. In Advances in Rice Genetics: (In 2 Parts); World Scientific: Singapore, 2003; pp. 143–145. [Google Scholar]
- Jeung, J.; Heu, S.; Shin, M.; Vera Cruz, C.; Jena, K. Dynamics of Xanthomonas oryzae pv. oryzae populations in Korea and their relationship to known bacterial blight resistance genes. Phytopathology 2006, 96, 867–875. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Shin, M.; Kim, W.; Mo, Y.; Nam, J.; Noh, T.; Kim, B.; Ko, J. Effective combination of resistance genes against rice bacterial blight pathogen. Korean J. Breed. Sci. 2009, 41, 244–251. [Google Scholar]
- Suh, J.-P.; Noh, T.-H.; Kim, K.-Y.; Kim, J.-J.; Kim, Y.-G.; Jena, K.K. Expression levels of three bacterial blight resistance genes against K3a race of Korea by molecular and phenotype analysis in japonica rice (O. sativa L.). J. Crop. Sci. Biotechnol. 2009, 12, 103–108. [Google Scholar] [CrossRef]
- Joseph, M.; Gopalakrishnan, S.; Sharma, R.; Singh, V.; Singh, A.; Singh, N.; Mohapatra, T. Combining bacterial blight resistance and Basmati quality characteristics by phenotypic and molecular marker-assisted selection in rice. Mol. Breed. 2004, 13, 377–387. [Google Scholar] [CrossRef]
- Rajpurohit, D.; Kumar, R.; Kumar, M.; Paul, P.; Awasthi, A.; Basha, P.O.; Puri, A.; Jhang, T.; Singh, K.; Dhaliwal, H.S. Pyramiding of two bacterial blight resistance and a semidwarfing gene in Type 3 Basmati using marker-assisted selection. Euphytica 2011, 178, 111–126. [Google Scholar] [CrossRef]
- Sundaram, R.M.; Vishnupriya, M.; Laha, G.S.; Rani, N.S.; Rao, P.S.; Balachandran, S.M.; Reddy, G.A.; Sarma, N.P.; Sonti, R.V. Introduction of bacterial blight resistance into Triguna, a high yielding, mid-early duration rice variety. Biotechnol. J. Healthc. Nutr. Technol. 2009, 4, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Shanti, M.; Shenoy, V.; Devi, G.L.; Kumar, V.M.; Premalatha, P.; Kumar, G.N.; Shashidhar, H.; Zehr, U.; Freeman, W. Marker-assisted breeding for resistance to bacterial leaf blight in popular cultivar and parental lines of hybrid rice. J. Plant Pathol. 2010, 495–501. [Google Scholar]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [Green Version]
- Das, G.; Rao, G.J.; Varier, M.; Prakash, A.; Prasad, D. Improved Tapaswini having four BB resistance genes pyramided with six genes/QTLs, resistance/tolerance to biotic and abiotic stresses in rice. Sci. Rep. 2018, 8, 2413. [Google Scholar]
- Luo, Y.; Sangha, J.S.; Wang, S.; Li, Z.; Yang, J.; Yin, Z. Marker-assisted breeding of Xa4, Xa21 and Xa27 in the restorer lines of hybrid rice for broad-spectrum and enhanced disease resistance to bacterial blight. Mol. Breed. 2012, 30, 1601–1610. [Google Scholar] [CrossRef]
- Suh, J.-P.; Jeung, J.-U.; Noh, T.-H.; Cho, Y.-C.; Park, S.-H.; Park, H.-S.; Shin, M.-S.; Kim, C.-K.; Jena, K.K. Development of breeding lines with three pyramided resistance genes that confer broad-spectrum bacterial blight resistance and their molecular analysis in rice. Rice 2013, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.S.; Kumari, K.A.; Rani, C.V.D.; Sundaram, R.; Vanisree, S.; Jamaloddin, M.; Swathi, G. Study of simple sequence repeat (SSR) polymorphism for biotic stress resistance in elite rice variety JGL 1798. Afr. J. Biotechnol. 2013, 12, 5833–5838. [Google Scholar]
- Davierwala, A.; Reddy, A.; Lagu, M.; Ranjekar, P.; Gupta, V. Marker assisted selection of bacterial blight resistance genes in rice. Biochem. Genet. 2001, 39, 261–278. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Jiang, J.; Dong, X.; Ali, J.; Mou, T. Introgression of bacterial blight (BB) resistance genes Xa7 and Xa21 into popular restorer line and their hybrids by molecular marker-assisted backcross (MABC) selection scheme. Afr. J. Biotechnol. 2012, 11, 8225–8233. [Google Scholar]
- Dokku, P.; Das, K.; Rao, G. Pyramiding of four resistance genes of bacterial blight in Tapaswini, an elite rice cultivar, through marker-assisted selection. Euphytica 2013, 192, 87–96. [Google Scholar] [CrossRef]
- Mundt, C. Probability of mutation to multiple virulence and durability of resistance gene pyramids. Phytopathology 1990, 80, 221–223. [Google Scholar] [CrossRef]
- Ramalingam, J.; Raveendra, C.; Savitha, P.; Vidya, V.; Chaithra, T.L.; Velprabakaran, S.; Saraswathi, R.; Ramanathan, A.; Pillai, M.P.A.; Arumugachamy, S. Gene pyramiding for achieving enhanced resistance to bacterial blight, blast, and sheath blight diseases in rice. Front. Plant Sci. 2020, 11, 1662. [Google Scholar] [CrossRef] [PubMed]
- Finatto, T.; de Oliveira, A.C.; Chaparro, C.; Da Maia, L.C.; Farias, D.R.; Woyann, L.G.; Mistura, C.C.; Soares-Bresolin, A.P.; Llauro, C.; Panaud, O. Abiotic stress and genome dynamics: Specific genes and transposable elements response to iron excess in rice. Rice 2015, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Mew, T. Current status and future prospects of research on bacterial blight of rice. Annu. Rev. Phytopathol. 1987, 25, 359–382. [Google Scholar] [CrossRef]
- Zhai, W.; Zhu, L. Rice bacterial blight resistance genes and their utilization in molecular breeding. Adv. Biotechnol. 1999, 19, 9–15. [Google Scholar]
- Dokku, P.; Das, K.; Rao, G. Genetic enhancement of host plant-resistance of the Lalat cultivar of rice against bacterial blight employing marker-assisted selection. Biotechnol. Lett. 2013, 35, 1339–1348. [Google Scholar] [CrossRef] [PubMed]
- Dash, A.K.; Rao, R.N.; Rao, G.; Verma, R.L.; Katara, J.L.; Mukherjee, A.K.; Singh, O.N.; Bagchi, T.B. Phenotypic and marker-assisted genetic enhancement of parental lines of Rajalaxmi, an elite rice hybrid. Front. Plant Sci. 2016, 7, 1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, G.; Rao, G. Molecular marker assisted gene stacking for biotic and abiotic stress resistance genes in an elite rice cultivar. Front. Plant Sci. 2015, 6, 698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, S.K.; Nayak, D.K.; Mohanty, S.; Behera, L.; Barik, S.R.; Pandit, E.; Lenka, S.; Anandan, A. Pyramiding of three bacterial blight resistance genes for broad-spectrum resistance in deepwater rice variety, Jalmagna. Rice 2015, 8, 19. [Google Scholar] [CrossRef]
- Sanchez, A.; Brar, D.; Huang, N.; Li, Z.; Khush, G. Sequence tagged site marker-assisted selection for three bacterial blight resistance genes in rice. Crop. Sci. 2000, 40, 792–797. [Google Scholar] [CrossRef]
- Jialing, L.; Chunchao, W.; Dan, Z.; Jianmin, L.; Xiaorong, S.; Yingyao, S.; Yongli, Z. Genome-wide association study dissects resistance loci against bacterial blight in a diverse rice panel from the 3000 rice genomes project. Rice 2021, 14, 22. [Google Scholar]
- Sugio, A.; Yang, B.; Zhu, T.; White, F.F. Two type III effector genes of Xanthomonas oryzae pv. oryzae control the induction of the host genes OsTFII Agamma1 and OsTFX1 during bacterial blight of rice. Proc. Natl. Acad. Sci. USA 2007, 104, 10720–10725. [Google Scholar] [CrossRef] [Green Version]
- Ou, S.H. Rice Diseases; IRRI: Los Banos, Philippines, 1985. [Google Scholar]
- Chien, C.; Shieh, L. Pathogenic Groups of Xanthomonas campestris pv. oryzae in Taiwan. J. Agric. Res. 1989, 38, 216–228. [Google Scholar]
- Hsieh, L.; Chang, Y.; Hsieh, T. Improvement of resistant screening techniques for bacterial leaf blight of rice caused by Xanthomonas oryzae pv. oryzae. J. Taiwan Agric. Res. 2005, 54, 15–22. [Google Scholar]
- Kauffman, H. An improved technique for evaluat-ing resistance of rice varieties to Xanthomonas oryzae. Plant Dis. Rep. 1973, 57, 537–541. [Google Scholar]
- IRRI. Training Module: Stress and Disease Tolerance; IRRI: Los Banos, Philippines, 2006; Available online: http://www.knowledgebank.irri.org (accessed on 12 May 2021).
- Xu, G.-W.; Magill, C.; Schertz, K.; Hart, G. A RFLP linkage map of Sorghum bicolor (L.) Moench. Theor. Appl. Genet. 1994, 89, 139–145. [Google Scholar] [CrossRef] [PubMed]
- IRRI. A Report on Bacterial Blight Resistance Genes Xa4, xa5, Xa7, xa13, Xa21, Xa23, and Xa27 Detection Protocol Using Gene-Based and Linked Markers; IRRI: Los Banos, Philippines, 2013; pp. 1–14. [Google Scholar]
- Kottapalli, K.R.; Narasu, M.L.; Jena, K.K. Effective strategy for pyramiding three bacterial blight resistance genes into fine grain rice cultivar, Samba Mahsuri, using sequence tagged site markers. Biotechnol. Lett. 2010, 32, 989–996. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Races | Phenotypes | |||
|---|---|---|---|---|
| R | MR | MS | S | |
| C5 | 38 | 4 | 21 | 0 |
| P6 | 8 | 17 | 0 | 0 |
| V | 7 | 6 | 0 | 0 |
| C5 + P6 | 27 | 5 | 10 | 0 |
| C5 + V | 29 | 7 | 4 | 0 |
| P6 + V | 3 | 3 | 0 | 0 |
| C5 + P6 + V | 55 | 8 | 11 | 2 |
| Total | 217 | 48 | ||
| Phenotypic Status | Genic Status * | RILs with Different Genes | |||
|---|---|---|---|---|---|
| Xa4 | xa5 | xa13 | Xa21 | ||
| Single locus | 41 | 23 | 29 | 25 | |
| Resistant | Double loci | 19 | 20 | 19 | 22 |
| Triple loci | 29 | 18 | 27 | 28 | |
| Quadruple loci | 11 | 11 | 11 | 11 | |
| Susceptible | No amplification | 165 | 193 | 179 | 179 |
| Total | 265 | 265 | 265 | 265 | |
| Group | R Genes | Number of RILs with Tag Number within Parenthesis |
|---|---|---|
| G1 | Xa4 + xa5 | 4 (68, 78, 163, 219) |
| G2 | Xa4 + xa13 | 3 (107, 121, 128) |
| G3 | Xa4 + Xa21 | 12 (16, 28, 63, 66, 69, 91, 126, 164, 180, 189, 212, 255) |
| G4 | xa5 + xa13 | 11 (39, 43, 52, 61, 71, 117, 144, 182, 202, 210, 253) |
| G5 | xa5 + Xa21 | 5 (109, 134, 143, 213, 252) |
| G6 | xa13 + Xa21 | 5 (11, 77, 191, 211, 248) |
| G7 | Xa4 + xa5 + xa13 | 6 (47, 58, 64, 206, 235, 262) |
| G8 | Xa4 + xa5 + Xa21 | 7 (7, 34, 74, 94, 186, 207, 199) |
| G9 | Xa4 + xa13 + Xa21 | 16 (3, 31, 35, 40, 42, 49, 57, 79, 118, 122, 127, 141, 161, 162, 194, 247) |
| G10 | xa5 + xa13 + Xa21 | 5 (133, 174, 221, 222, 223) |
| G11 | Xa4 + xa5 + xa13 + Xa21 | 11 (12, 15, 32, 44, 51, 53, 155, 156, 166, 215, 232) |
| Genotype | HD | PH | TN | PL | S/P | SF% | L/W | TGW | Y |
|---|---|---|---|---|---|---|---|---|---|
| Ciherang | 108.00i | 83.33f | 9.67ab | 20.09i | 114.33f | 79.83c–e | 3.25b–d | 24.15c–f | 16.10i |
| IRBB60 | 117.00c–e | 82.13f | 9.27a–c | 21.85h | 125.27ef | 86.57ab | 2.76e | 23.51e–g | 17.24gh |
| G1 | 119.17a–c | 90.57de | 7.00gh | 23.75d–f | 189.30ab | 77.32e | 3.41ab | 23.41f–h | 19.38bc |
| G2 | 120.78a | 90.42de | 8.01c–g | 23.06fg | 117.70ef | 83.49a–d | 3.42ab | 23.56e–g | 22.64a |
| G3 | 120.44ab | 94.58cd | 6.85gh | 24.22c–e | 163.05b–d | 78.89de | 3.47a | 25.29ab | 19.92b |
| G4 | 114.91e–g | 100.82ab | 8.66b–f | 25.26a | 186.13ab | 85.48ab | 3.43a | 24.48b–f | 18.56c–e |
| G5 | 116.47d–f | 96.12b–d | 7.77d–g | 24.52a–d | 193.21a | 85.45ab | 3.45a | 25.27a–c | 18.68fg |
| G6 | 116.60de | 95.37b–d | 8.83b–e | 24.20c–e | 172.03ab | 79.71de | 3.49a | 23.87d–f | 16.63hi |
| G7 | 118.11cd | 103.59a | 7.91c–g | 25.31a | 187.23ab | 83.55a–d | 3.43a | 25.15a–c | 18.16ef |
| G8 | 113.10gh | 99.91a–c | 7.94c–g | 23.56ef | 190.78a | 81.54b–e | 3. 41ab | 24.58a–e | 19.08cd |
| G9 | 116.67de | 100.18a–c | 8.85b–e | 25.04ab | 191.44a | 85.21a–c | 3.38a–c | 24.72a–d | 19.29bc |
| G10 | 116.80de | 96.33b–d | 10.36a | 24.72a–c | 142.67de | 87.46a | 3.53a | 25.64a | 18.29d–f |
| G11 | 116.55d–f | 101.36ab | 9.16a–d | 25.20ab | 171.65a–c | 88.26a | 3.42ab | 24.76a–d | 18.57c–e |
| IRBB4 | 114.33fg | 88.00ef | 7.67e–h | 23.21fg | 140.00df | 79.67de | 3.17d | 22.36hi | 18.45d–f |
| IRBB5 | 115.67ef | 85.44ef | 7.00gh | 23.32fg | 128.33ef | 83.28a–d | 3.08d | 22.13i | 17.63fg |
| IRBB13 | 111.00h | 86.00ef | 6.67gh | 23.50e–f | 128.67ef | 83.00a–d | 3.19d | 22.70g–i | 16.89g–i |
| IRBB21 | 118.33b–d | 86.56ef | 7.33f–h | 24.45b–d | 120.00ef | 83.53a–d | 3.20cd | 22.60g–i | 18.26d–f |
| IR24 | 115.33ef | 99.00a–c | 6.33h | 22.58gh | 144.33c–e | 69.67f | 3.08d | 22.28hi | 16.11i |
| p (≤0.05) | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| St dev (±) | 0.72 | 2.00 | 0.46 | 0.26 | 9.00 | 1.78 | 0.06 | 0.37 | 0.27 |
| Variables | PC1 | PC2 | PC3 |
|---|---|---|---|
| C5 | 0.189 | −0.145 | 0.11 |
| P6 | −0.084 | −0.041 | 0.644 |
| V | −0.123 | 0.21 | 0.514 |
| C5 P6 | 0.234 | −0.096 | 0.201 |
| C5V | 0.242 | −0.063 | 0.191 |
| P6 V | −0.117 | 0.283 | 0.436 |
| C5 P6 V | 0.26 | 0.021 | 0.091 |
| Xa4 | 0.009 | 0.454 | −0.09 |
| xa5 | −0.06 | 0.44 | −0.092 |
| xa13 | −0.047 | 0.445 | −0.092 |
| Xa21 | −0.036 | 0.448 | −0.092 |
| Xa4 + xa5 (A) | 0.259 | 0.061 | 0.006 |
| Xa4 + xa13 (B) | 0.259 | 0.061 | 0.006 |
| Xa4 + Xa21 (C) | 0.259 | 0.061 | 0.006 |
| xa5 + xa13 (D) | 0.259 | 0.061 | 0.006 |
| xa5 + Xa21 (E) | 0.259 | 0.061 | 0.006 |
| xa13 + Xa21 (F) | 0.259 | 0.061 | 0.006 |
| Xa4 + xa5 + xa13 (G) | 0.259 | 0.061 | 0.006 |
| Xa4 + xa5 + Xa21 (H) | 0.259 | 0.061 | 0.006 |
| Xa4 + xa13 + Xa21 (I) | 0.259 | 0.061 | 0.006 |
| xa5 + xa13 + Xa21 (J) | 0.259 | 0.061 | 0.006 |
| Xa4 + xa5 + xa13 + Xa21 (K) | 0.259 | 0.061 | 0.006 |
| Eigenvalue | 14.474 | 4.689 | 2.145 |
| Proportion | 0.658 | 0.213 | 0.097 |
| Cumulative | 0.658 | 0.871 | 0.969 |
| Gene | Chr No. | Marker Name | Primer Sequences | Expected Size (bp) | References | |
|---|---|---|---|---|---|---|
| Xa4 | 11 | MP1 | Forward | ATCGATCGATCTTCACGAGG | 150 | [69] |
| MP2 | Reverse | TGCTATAAAAGGCATTCGGG | ||||
| xa5 | 5 | xa5 | Forward | GCTCGCCATTCAAGTTCTTGAG | 198 | [70] |
| Reverse | CCTTGATAGAAACCT TGCCTTGAC | |||||
| xa13 | 8 | xa13 prom | Forward | CCTGATATGTGAGGTAGT | 500 | [22] |
| Reverse | GAGAAAGGCTTAAGTGC | |||||
| Xa21 | 11 | pTA248 | Forward | CGATCGGTATAACAGCAAAAC | 1000 | [71] |
| Reverse | AGACGCGGTAATCGAAAGATGAAA | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biswas, P.L.; Nath, U.K.; Ghosal, S.; Goswami, G.; Uddin, M.S.; Ali, O.M.; Latef, A.A.H.A.; Laing, A.M.; Gao, Y.-M.; Hossain, A. Introgression of Bacterial Blight Resistance Genes in the Rice Cultivar Ciherang: Response against Xanthomonas oryzae pv. oryzae in the F6 Generation. Plants 2021, 10, 2048. https://doi.org/10.3390/plants10102048
Biswas PL, Nath UK, Ghosal S, Goswami G, Uddin MS, Ali OM, Latef AAHA, Laing AM, Gao Y-M, Hossain A. Introgression of Bacterial Blight Resistance Genes in the Rice Cultivar Ciherang: Response against Xanthomonas oryzae pv. oryzae in the F6 Generation. Plants. 2021; 10(10):2048. https://doi.org/10.3390/plants10102048
Chicago/Turabian StyleBiswas, Priya Lal, Ujjal Kumar Nath, Sharmistha Ghosal, Gayatri Goswami, Md. Shalim Uddin, Omar M. Ali, Arafat Abdel Hamed Abdel Latef, Alison M. Laing, Yong-Ming Gao, and Akbar Hossain. 2021. "Introgression of Bacterial Blight Resistance Genes in the Rice Cultivar Ciherang: Response against Xanthomonas oryzae pv. oryzae in the F6 Generation" Plants 10, no. 10: 2048. https://doi.org/10.3390/plants10102048
APA StyleBiswas, P. L., Nath, U. K., Ghosal, S., Goswami, G., Uddin, M. S., Ali, O. M., Latef, A. A. H. A., Laing, A. M., Gao, Y.-M., & Hossain, A. (2021). Introgression of Bacterial Blight Resistance Genes in the Rice Cultivar Ciherang: Response against Xanthomonas oryzae pv. oryzae in the F6 Generation. Plants, 10(10), 2048. https://doi.org/10.3390/plants10102048