Ttc21b Is Required in Bergmann Glia for Proper Granule Cell Radial Migration


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Husbandry
2.2. Histology and Immunohistochemistry
2.3. Section In Situ Hybridization and LacZ Staining
3. Results
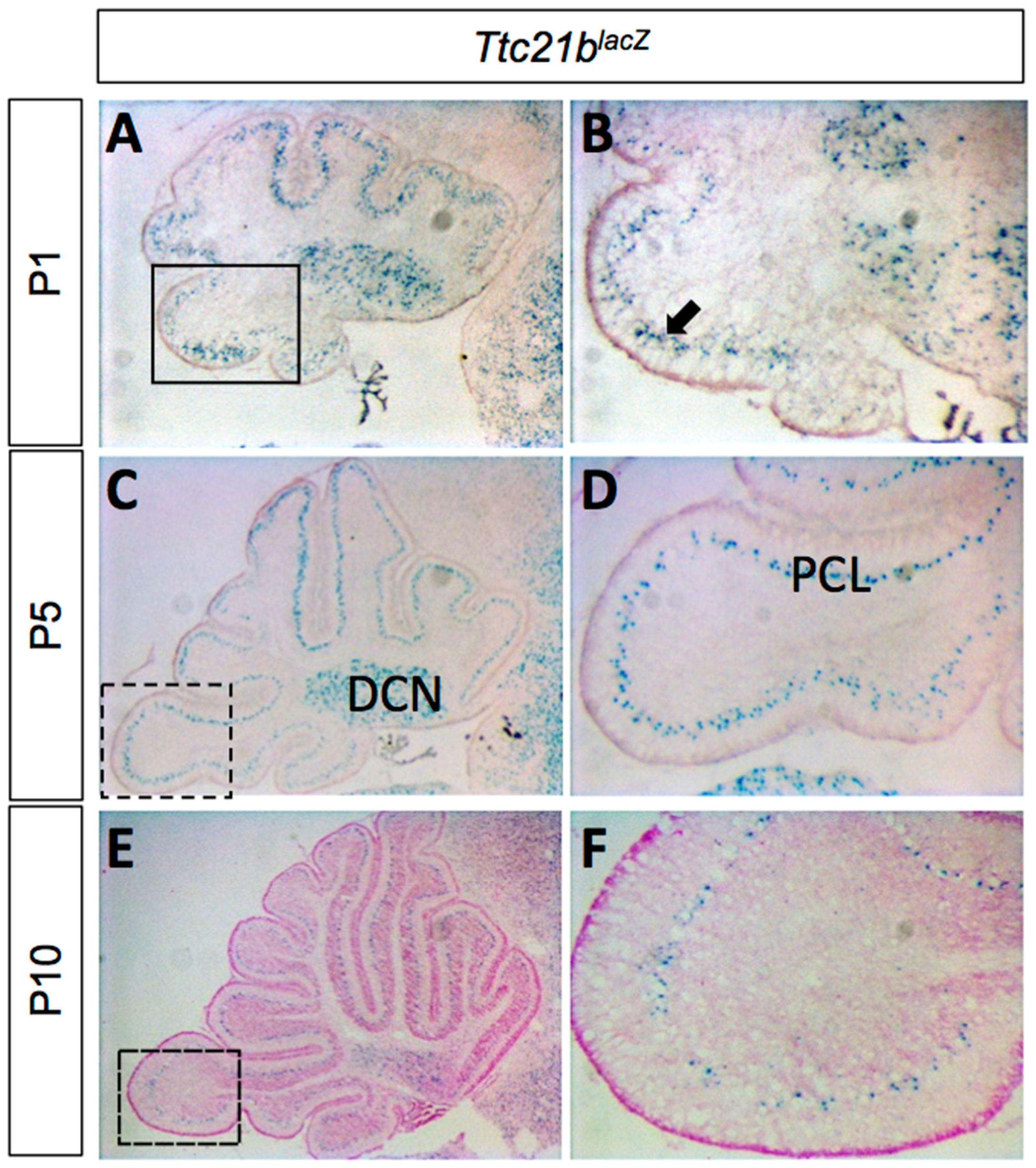
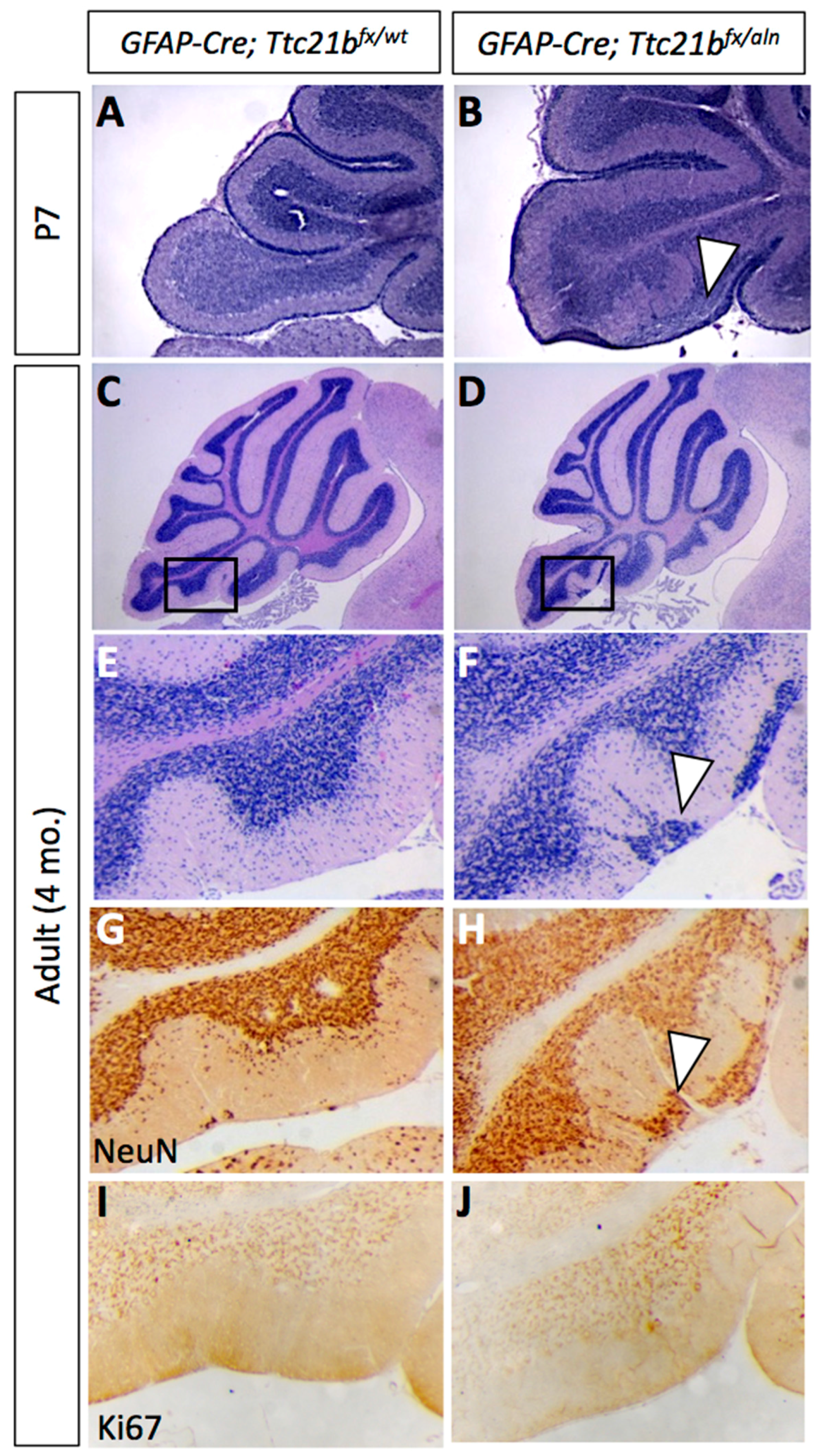
3.1. Ttc21b Is Enriched in the Purkinje Cell Layer of the Developing Cerebellum and GFAP-Cre Ablation of Ttc21b Results in Ectopic Granule Cells in the Cerebellum
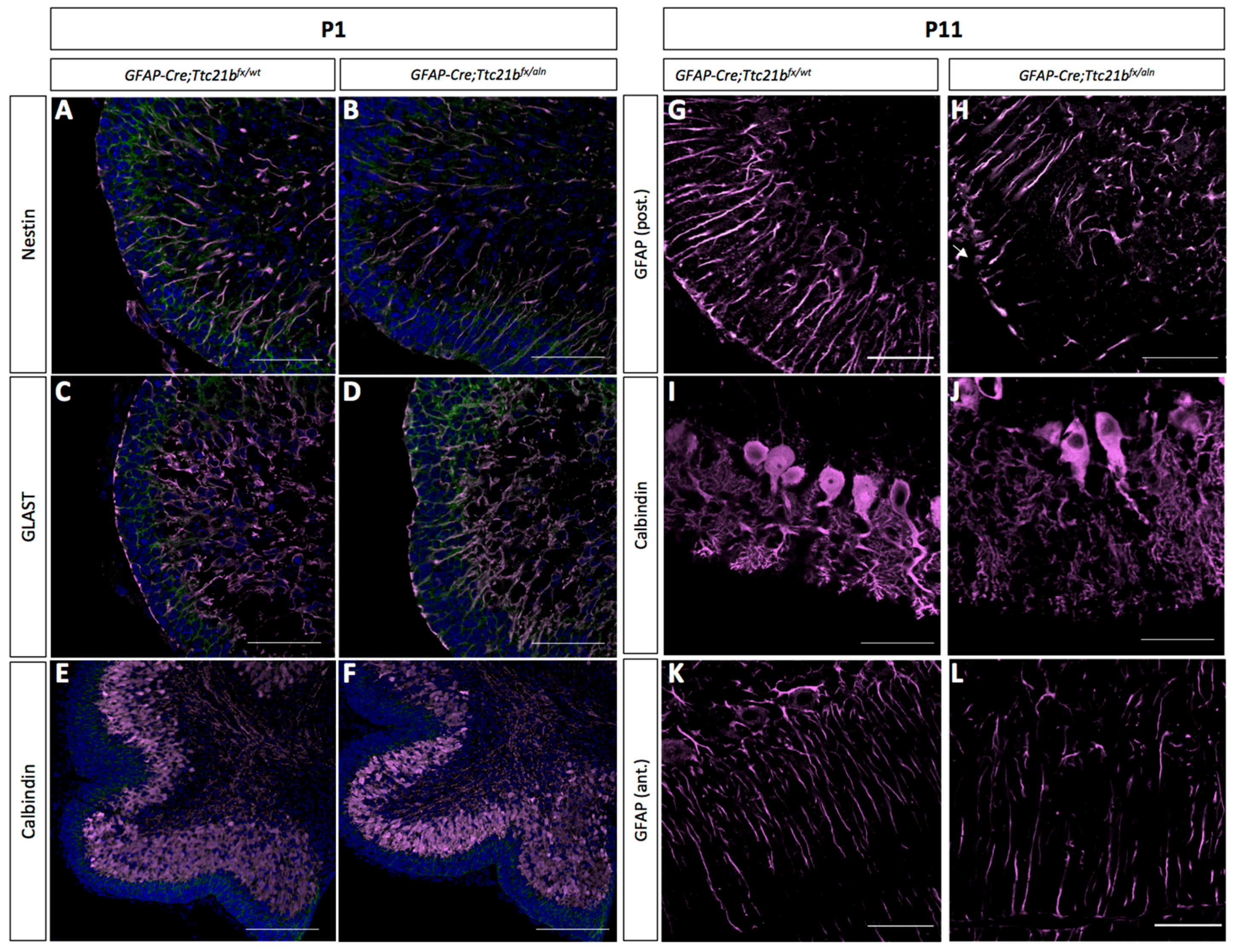
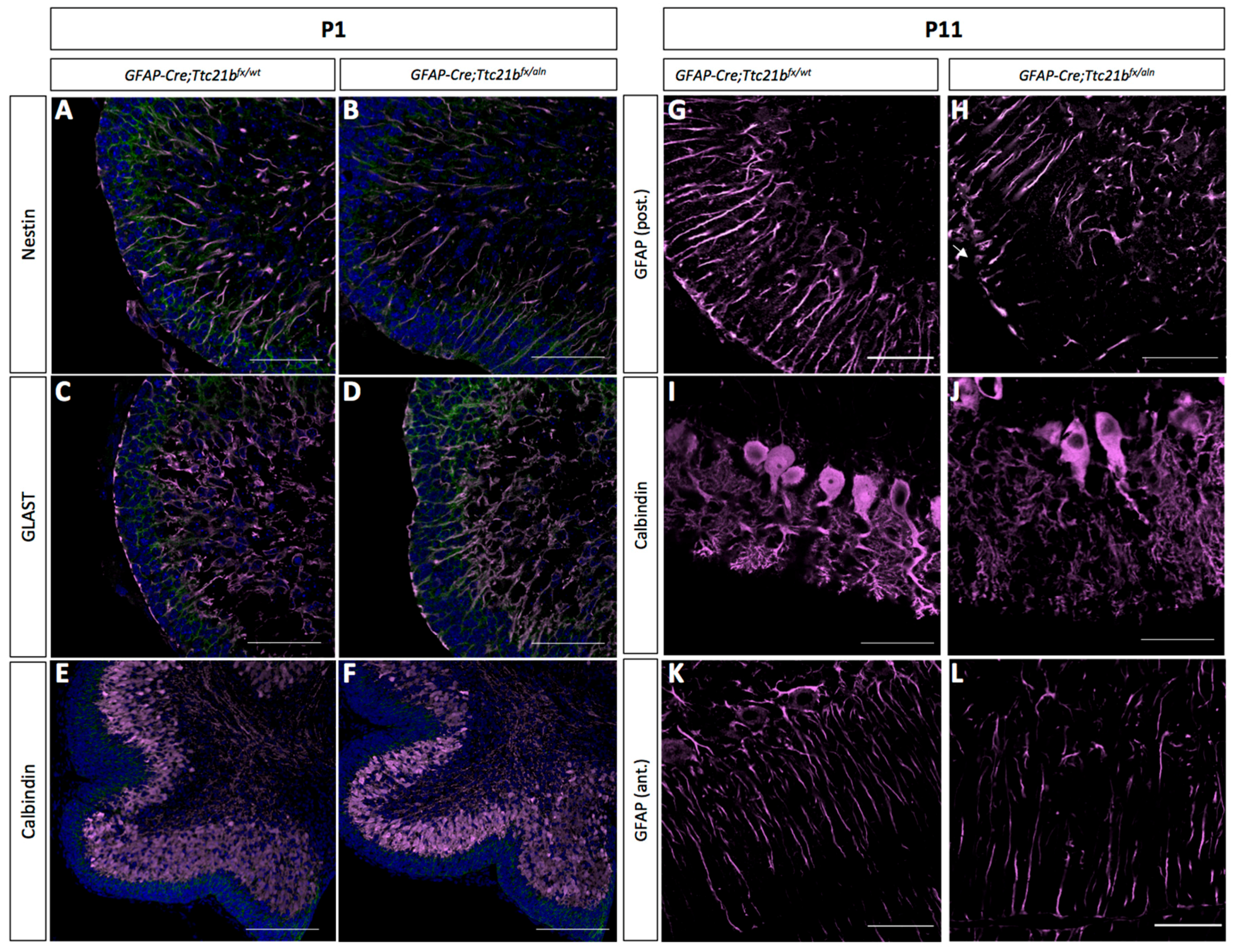
3.2. Purkinje Cell and Bergmann Glial Cell Development Are Disrupted upon Loss of Ttc21b
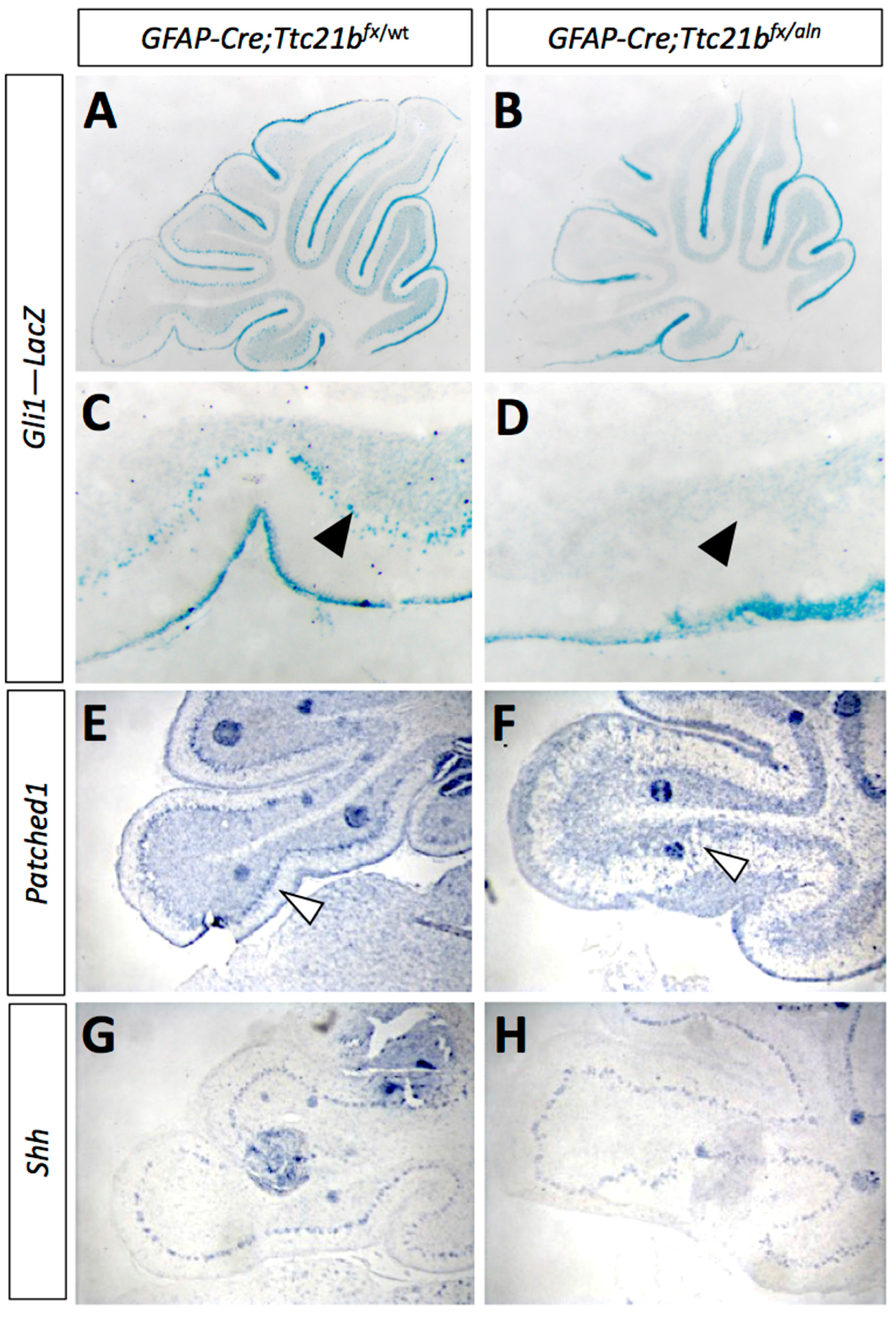
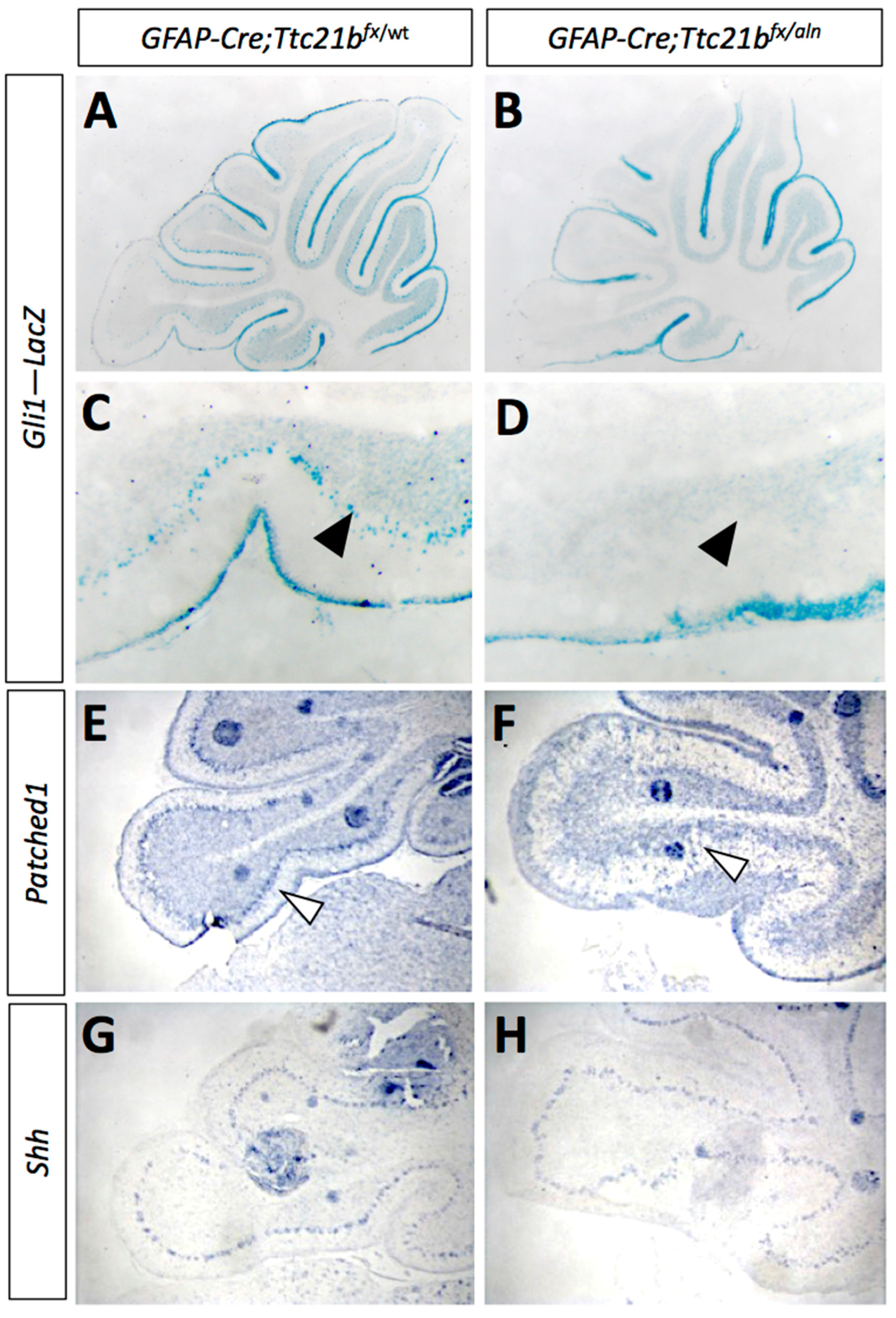
3.3. SHH Signaling Is Reduced in GFAP-Cre;Ttc21bflox/aln Cerebella
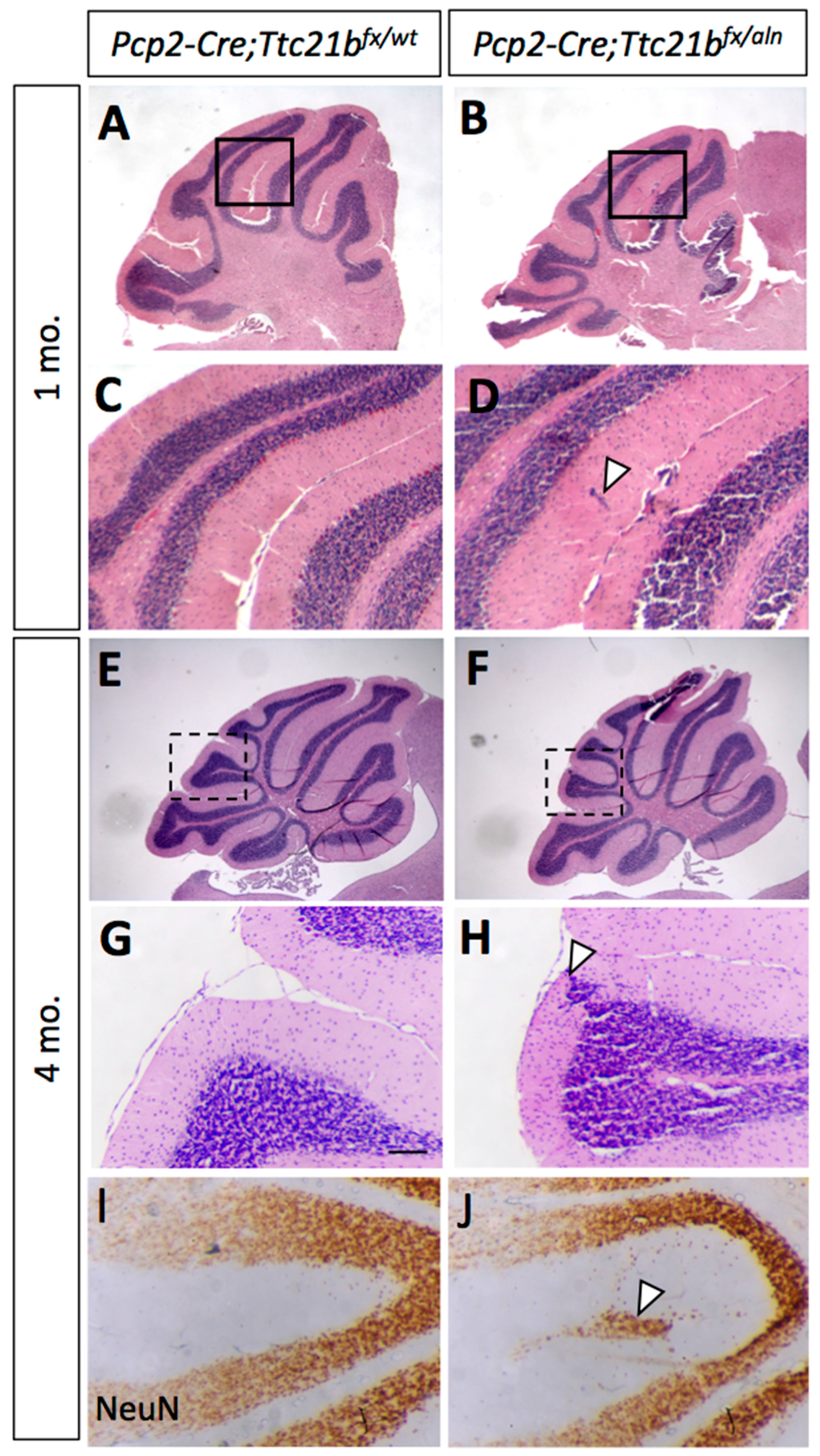
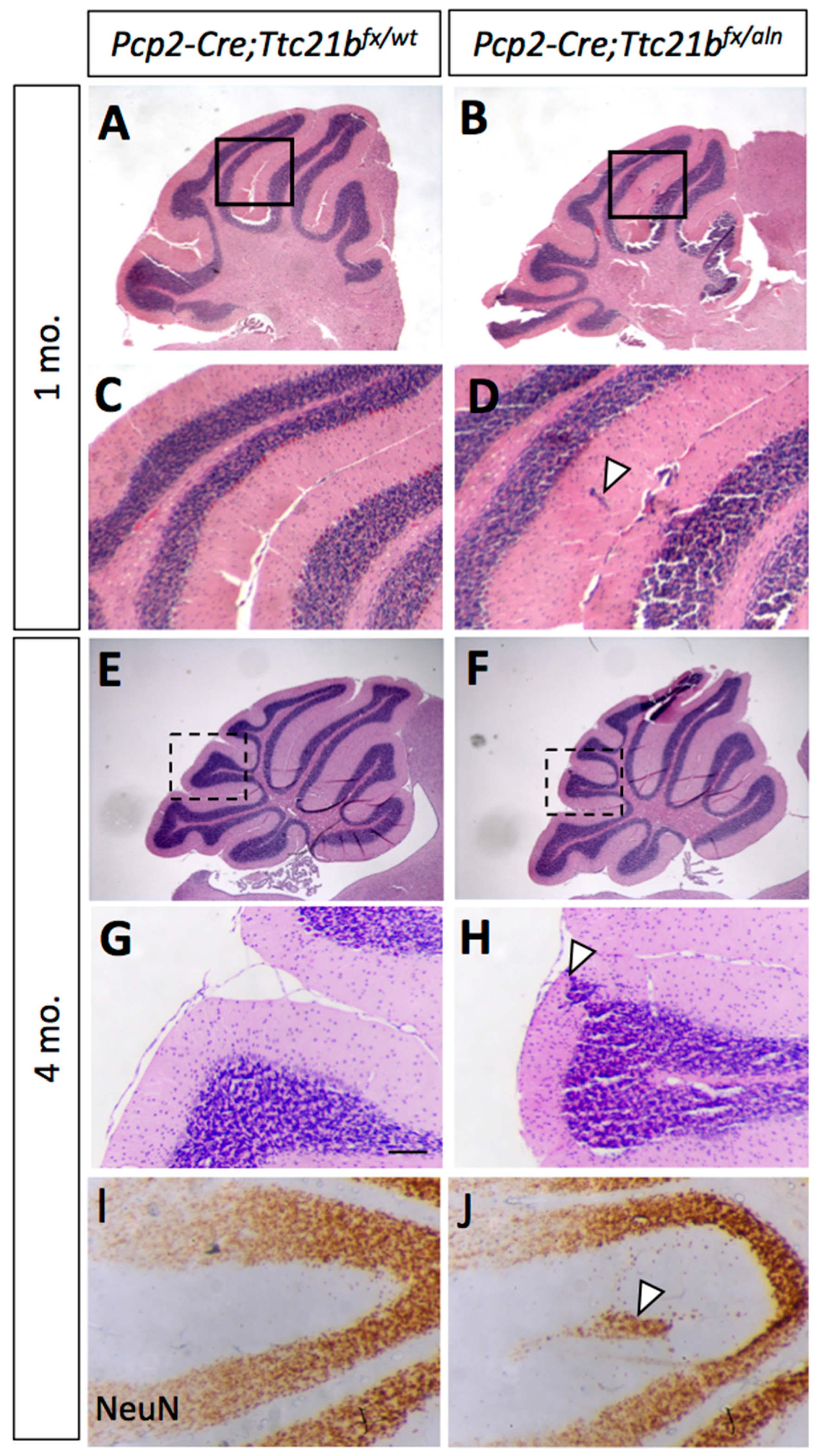
3.4. Purkinje Cell Ablation of Ttc21b Results in Milder Ectopic Granule Cell Phenotype
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Andersen, B.B.; Korbo, L.; Pakkenberg, B. A quantitative study of the human cerebellum with unbiased stereological techniques. J. Comp. Neurol. 1992, 326, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Ten Donkelaar, H.J.; Lammens, M. Development of the human cerebellum and its disorders. Clin. Perinatol. 2009, 36, 513–530. [Google Scholar] [CrossRef] [PubMed]
- Dahmane, N.; Ruiz i Altaba, A. Sonic hedgehog regulates the growth and patterning of the cerebellum. Development 1999, 126, 3089–3100. [Google Scholar] [PubMed]
- Wallace, V.A. Purkinje-cell-derived Sonic hedgehog regulates granule neuron precursor cell proliferation in the developing mouse cerebellum. Curr. Biol. 1999, 9, 445–448. [Google Scholar] [CrossRef]
- Wechsler-Reya, R.J.; Scott, M.P. Control of neuronal precursor proliferation in the cerebellum by Sonic Hedgehog. Neuron 1999, 22, 103–114. [Google Scholar] [CrossRef]
- Xu, H.; Yang, Y.; Tang, X.; Zhao, M.; Liang, F.; Xu, P.; Hou, B.; Xing, Y.; Bao, X.; Fan, X. Bergmann glia function in granule cell migration during cerebellum development. Mol. Neurobiol. 2013, 47, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Corrales, J.D.; Rocco, G.L.; Blaess, S.; Guo, Q.; Joyner, A.L. Spatial pattern of sonic hedgehog signaling through Gli genes during cerebellum development. Development 2004, 131, 5581–5590. [Google Scholar] [CrossRef] [PubMed]
- Corrales, J.D.; Blaess, S.; Mahoney, E.M.; Joyner, A.L. The level of sonic hedgehog signaling regulates the complexity of cerebellar foliation. Development 2006, 133, 1811–1821. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.M.; Gritli-Linde, A.; Smeyne, R.; Kottmann, A.; McMahon, A.P. Sonic hedgehog signaling is required for expansion of granule neuron precursors and patterning of the mouse cerebellum. Dev. Biol. 2004, 270, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Spassky, N.; Han, Y.G.; Aguilar, A.; Strehl, L.; Besse, L.; Laclef, C.; Ros, M.R.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Primary cilia are required for cerebellar development and Shh-dependent expansion of progenitor pool. Dev. Biol. 2008, 317, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, A.; Meunier, A.; Strehl, L.; Martinovic, J.; Bonniere, M.; Attie-Bitach, T.; Encha-Razavi, F.; Spassky, N. Analysis of human samples reveals impaired SHH-dependent cerebellar development in Joubert syndrome/Meckel syndrome. Proc. Natl. Acad. Sci. USA 2012, 109, 16951–16956. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, L.V.; Scott, M.P. Hedgehog and patched in neural development and disease. Neuron 1998, 21, 1243–1257. [Google Scholar] [CrossRef]
- Hallahan, A.R.; Pritchard, J.I.; Hansen, S.; Benson, M.; Stoeck, J.; Hatton, B.A.; Russell, T.L.; Ellenbogen, R.G.; Bernstein, I.D.; Beachy, P.A.; et al. The SmoA1 mouse model reveals that notch signaling is critical for the growth and survival of sonic hedgehog-induced medulloblastomas. Cancer Res. 2004, 64, 7794–7800. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.G.; Kim, H.J.; Dlugosz, A.A.; Ellison, D.W.; Gilbertson, R.J.; Alvarez-Buylla, A. Dual and opposing roles of primary cilia in medulloblastoma development. Nat. Med. 2009, 15, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Oliver, T.G.; Read, T.A.; Kessler, J.D.; Mehmeti, A.; Wells, J.F.; Huynh, T.T.; Lin, S.M.; Wechsler-Reya, R.J. Loss of patched and disruption of granule cell development in a pre-neoplastic stage of medulloblastoma. Development 2005, 132, 2425–2439. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.J.; Ellis, T.; Markant, S.L.; Read, T.A.; Kessler, J.D.; Bourboulas, M.; Schuller, U.; Machold, R.; Fishell, G.; Rowitch, D.H.; et al. Medulloblastoma can be initiated by deletion of Patched in lineage-restricted progenitors or stem cells. Cancer Cell 2008, 14, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Bangs, F.; Anderson, K.V. Primary Cilia and Mammalian Hedgehog Signaling. Cold Spring Harb. Perspect. Biol. 2017, 9, a028175. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Wang, B.; Niswander, L.A. Mouse intraflagellar transport proteins regulate both the activator and repressor functions of Gli transcription factors. Development 2005, 132, 3103–3111. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.V.; Haycraft, C.J.; Besschetnova, T.Y.; Turbe-Doan, A.; Stottmann, R.W.; Herron, B.J.; Chesebro, A.L.; Qiu, H.; Scherz, P.J.; Shah, J.V.; et al. THM1 negatively modulates mouse sonic hedgehog signal transduction and affects retrograde intraflagellar transport in cilia. Nat. Genet. 2008, 40, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Snedeker, J.; Schock, E.N.; Struve, J.N.; Chang, C.F.; Cionni, M.; Tran, P.V.; Brugmann, S.A.; Stottmann, R.W. Unique spatiotemporal requirements for intraflagellar transport genes during forebrain development. PLoS ONE 2017, 12, e0173258. [Google Scholar] [CrossRef] [PubMed]
- Stottmann, R.W.; Tran, P.V.; Turbe-Doan, A.; Beier, D.R. Ttc21b is required to restrict sonic hedgehog activity in the developing mouse forebrain. Dev. Biol. 2009, 335, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.E.; Zhang, Q.; Liu, Q.; Diplas, B.H.; Davey, L.M.; Hartley, J.; Stoetzel, C.; Szymanska, K.; Ramaswami, G.; Logan, C.V.; et al. TTC21B contributes both causal and modifying alleles across the ciliopathy spectrum. Nat. Genet. 2011, 43, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Niwa, S. The nephronophthisis-related gene ift-139 is required for ciliogenesis in Caenorhabditis elegans. Sci. Rep. 2016, 6, 31544. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.V.; Talbott, G.C.; Turbe-Doan, A.; Jacobs, D.T.; Schonfeld, M.P.; Silva, L.M.; Chatterjee, A.; Prysak, M.; Allard, B.A.; Beier, D.R. Downregulating hedgehog signaling reduces renal cystogenic potential of mouse models. J. Am. Soc. Nephrol. 2014, 25, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Muzumdar, M.D.; Tasic, B.; Miyamichi, K.; Li, L.; Luo, L. A global double-fluorescent Cre reporter mouse. Genesis 2007, 45, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, L.; Theis, M.; Alvarez-Maya, I.; Brenner, M.; Willecke, K.; Messing, A. hGFAP-cre transgenic mice for manipulation of glial and neuronal function in vivo. Genesis 2001, 31, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Barski, J.J.; Dethleffsen, K.; Meyer, M. Cre recombinase expression in cerebellar Purkinje cells. Genesis 2000, 28, 93–98. [Google Scholar] [CrossRef]
- Bai, C.B.; Auerbach, W.; Lee, J.S.; Stephen, D.; Joyner, A.L. Gli2, but not Gli1, is required for initial Shh signaling and ectopic activation of the Shh pathway. Development 2002, 129, 4753–4761. [Google Scholar] [PubMed]
- Yamada, K.; Watanabe, M. Cytodifferentiation of Bergmann glia and its relationship with Purkinje cells. Anat. Sci. Int. 2002, 77, 94–108. [Google Scholar] [CrossRef] [PubMed]
- White, J.J.; Sillitoe, R.V. Development of the cerebellum: From gene expression patterns to circuit maps. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Platt, K.A.; Censullo, P.; Ruiz i Altaba, A. Gli1 is a target of Sonic hedgehog that induces ventral neural tube development. Development 1997, 124, 2537–2552. [Google Scholar] [PubMed]
- Frick, A.; Grammel, D.; Schmidt, F.; Poschl, J.; Priller, M.; Pagella, P.; von Bueren, A.O.; Peraud, A.; Tonn, J.C.; Herms, J.; et al. Proper cerebellar development requires expression of beta1-integrin in Bergmann glia, but not in granule neurons. Glia 2012, 60, 820–832. [Google Scholar] [CrossRef] [PubMed]
- Inouye, M.; Hayasaka, S.; Funahashi, A.; Yamamura, H. Gamma-radiation produces abnormal Bergmann fibers and ectopic granule cells in mouse cerebellar cortex. J. Radiat. Res. 1992, 33, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, T.C. Interactions between Purkinje neurones and Bergmann glia. Cerebellum 2006, 5, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Lordkipanidze, T.; Dunaevsky, A. Purkinje cell dendrites grow in alignment with Bergmann glia. Glia 2005, 51, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xu, Q.; Wang, W.; Takano, T.; Nedergaard, M. Bergmann glia modulate cerebellar Purkinje cell bistability via Ca2+-dependent K+ uptake. Proc. Natl. Acad. Sci. USA 2012, 109, 7911–7916. [Google Scholar] [CrossRef] [PubMed]
- Huynh Cong, E.; Bizet, A.A.; Boyer, O.; Woerner, S.; Gribouval, O.; Filhol, E.; Arrondel, C.; Thomas, S.; Silbermann, F.; Canaud, G.; et al. A homozygous missense mutation in the ciliary gene TTC21B causes familial FSGS. J. Am. Soc. Nephrol. 2014, 25, 2435–2443. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Longo, C.M.; Ackerman, S.L. Genetic and physical mapping of the cerebellar deficient folia (cdf) locus on mouse chromosome 6. Genomics 2000, 69, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Blaess, S.; Stephen, D.; Joyner, A.L. Gli3 coordinates three-dimensional patterning and growth of the tectum and cerebellum by integrating Shh and Fgf8 signaling. Development 2008, 135, 2093–2103. [Google Scholar] [CrossRef] [PubMed]
- Romani, M.; Micalizzi, A.; Valente, E.M. Joubert syndrome: Congenital cerebellar ataxia with the molar tooth. Lancet Neurol. 2013, 12, 894–905. [Google Scholar] [CrossRef]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Driver, A.M.; Shumrick, C.; Stottmann, R.W. Ttc21b Is Required in Bergmann Glia for Proper Granule Cell Radial Migration. J. Dev. Biol. 2017, 5, 18. https://doi.org/10.3390/jdb5040018
Driver AM, Shumrick C, Stottmann RW. Ttc21b Is Required in Bergmann Glia for Proper Granule Cell Radial Migration. Journal of Developmental Biology. 2017; 5(4):18. https://doi.org/10.3390/jdb5040018
Chicago/Turabian StyleDriver, Ashley M., Christopher Shumrick, and Rolf W. Stottmann. 2017. "Ttc21b Is Required in Bergmann Glia for Proper Granule Cell Radial Migration" Journal of Developmental Biology 5, no. 4: 18. https://doi.org/10.3390/jdb5040018
APA StyleDriver, A. M., Shumrick, C., & Stottmann, R. W. (2017). Ttc21b Is Required in Bergmann Glia for Proper Granule Cell Radial Migration. Journal of Developmental Biology, 5(4), 18. https://doi.org/10.3390/jdb5040018