

Genetic and Epigenetic Regulation of Drosophila Oocyte Determination


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
Germ Line Cyst and Oocyte Determination
2. Drosophila Melanogaster Oogenesis
2.1. Cystoblast, Fusome, and Germ Line Cyst Formation
2.2. First among Equals: Oocyte Determination
2.3. Oocyte-Specific Factors
2.4. Shall I Stay or Shall I Go: Entry into Meiosis
3. Germ Cells Differentiation: Regulation of Gene Expression
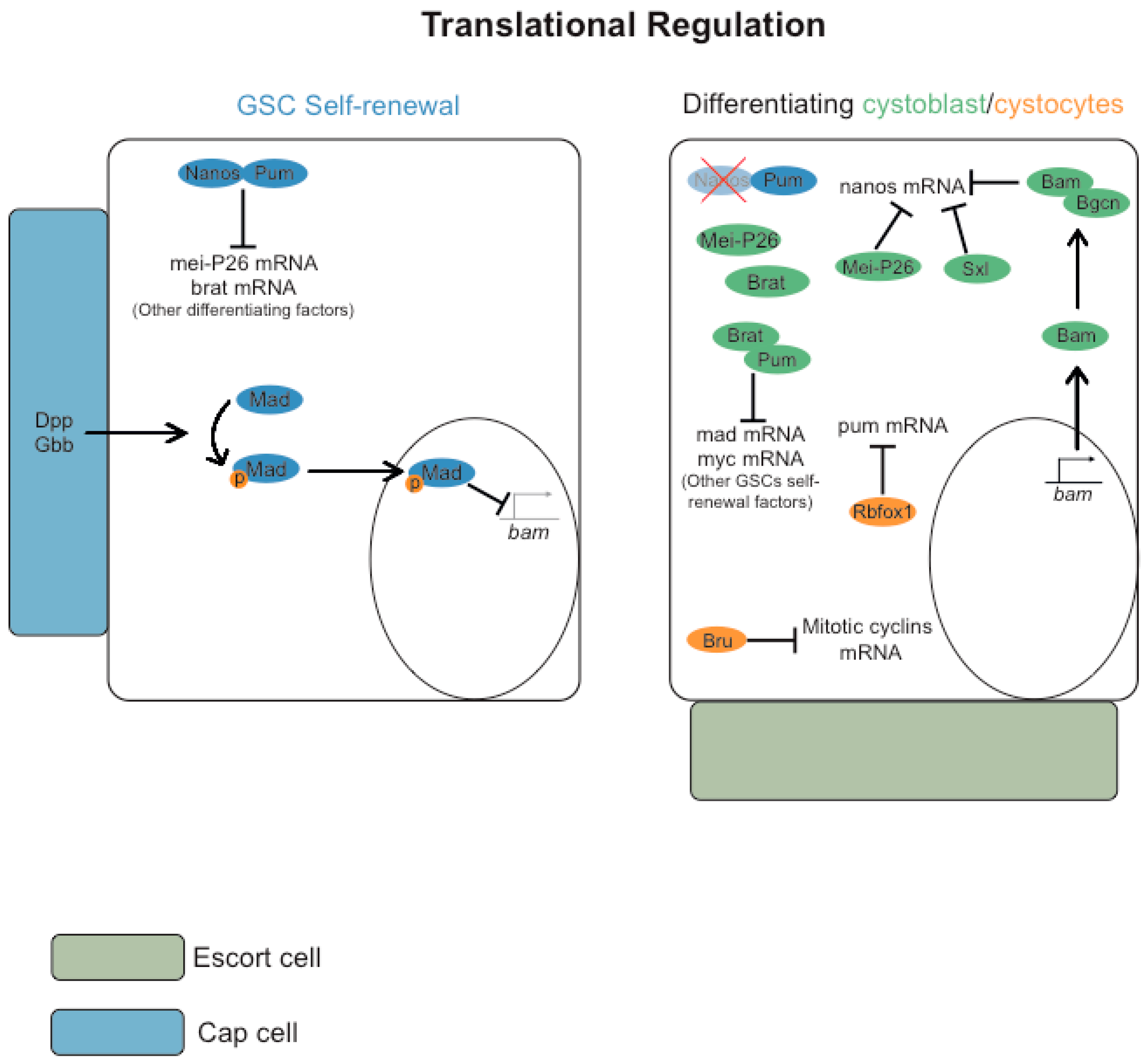
3.1. Post-Transcriptional Regulation
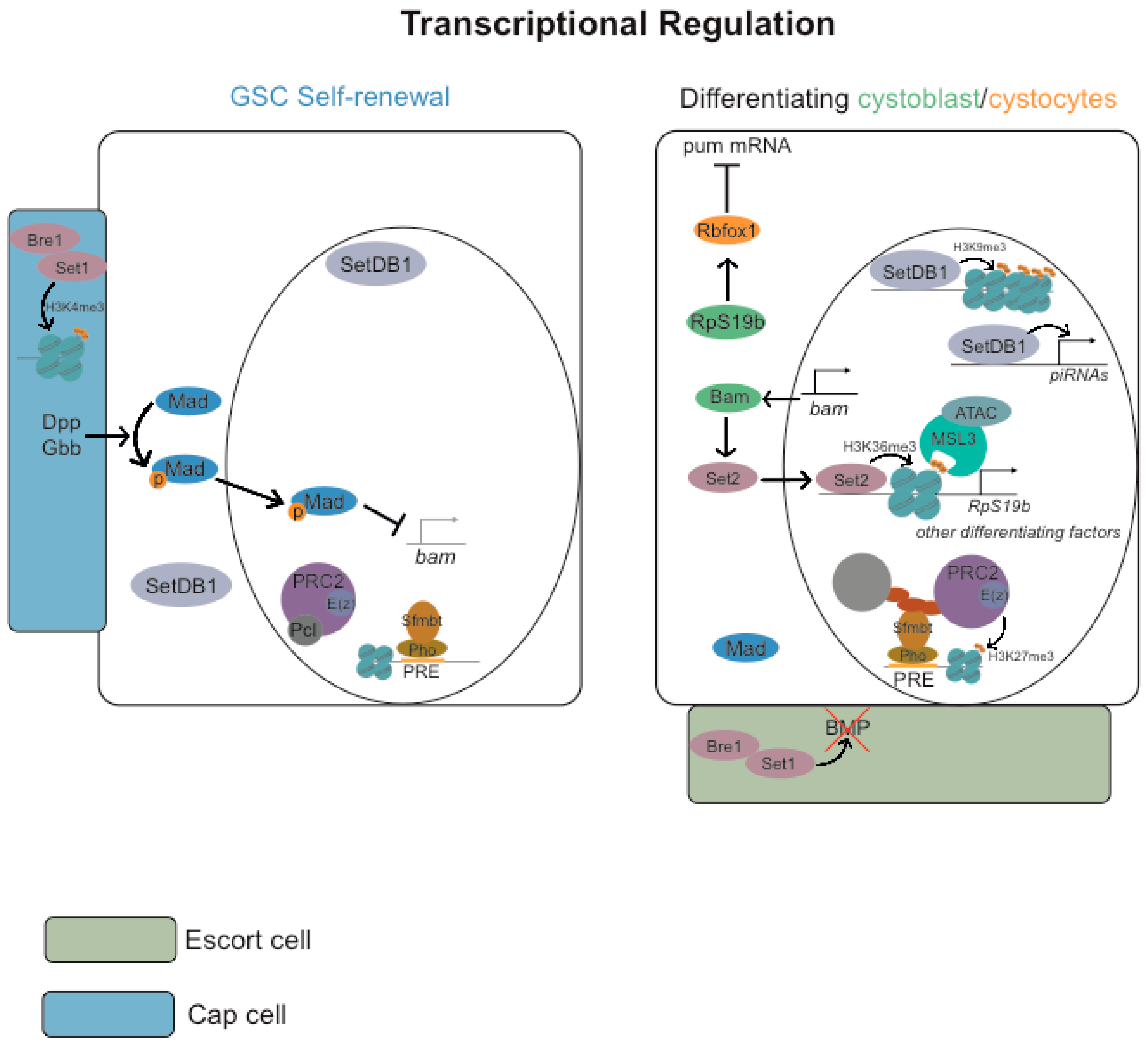
3.2. Transcriptional Regulation
3.2.1. Repressive Histone Marks (H3K9me3 and H3K27me3)
3.2.2. Active Histone Marks (H2B Monoubiquitylation H3K4me3, H3K36me3 and H3K79me3)
3.2.3. ATP-Dependent Chromatin-Remodeling
4. Drosophila and Mammalian Oogenesis
5. Final Remarks and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peterson, N.G.; Fox, D.T. Communal living: The role of polyploidy and syncytia in tissue biology. Chromosome Res. 2021, 29, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Spradling, A.C.; Niu, W.; Yin, Q.; Pathak, M.; Maurya, B. Conservation of oocyte development in germline cysts from Drosophila to mouse. eLife 2022, 11, e83230. [Google Scholar] [CrossRef] [PubMed]
- Spradling, A.C. Developmental genetics of oogenesis. In The Development of Drosophila Melanogaster; Bate, M., Martinez Arias, A., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1993; Volume 1. [Google Scholar]
- Mahajan-Miklos, S.; Cooley, L. Intercellular cytoplasm transport during Drosophila oogenesis. Dev. Biol. 1994, 165, 336–351. [Google Scholar] [CrossRef] [PubMed]
- Huynh, J.R.; St Johnston, D. The origin of asymmetry: Early polarisation of the Drosophila germline cyst and oocyte. Curr. Biol. 2004, 14, R438–R449. [Google Scholar] [CrossRef]
- Lei, L.; Spradling, A.C. Mouse oocytes differentiate through organelle enrichment from sister cyst germ cells. Science 2016, 352, 95–99. [Google Scholar] [CrossRef]
- Niu, W.; Spradling, A.C. Mouse oocytes develop in cysts with the help of nurse cells. Cell 2022, 185, 2576–2590.e12. [Google Scholar] [CrossRef]
- Matova, N.; Cooley, L. Comparative aspects of animal oogenesis. Dev. Biol. 2001, 231, 291–320. [Google Scholar] [CrossRef]
- Buning, J. Germ cell cluster formation in insect ovaries. Int. J. Insect Morphol. Embryol. 1993, 22, 237–253. [Google Scholar] [CrossRef]
- Jezierska, M.; Miernik, A.; Sojka, J.; Student, S.; Sliwinska, M.A.; Gross, V.; Poprawa, I. Oogenesis in the tardigrade Hypsibius exemplaris Gasiorek, Stec, Morek & Michalczyk, 2018 (Eutardigrada, Hypsibiidae). Micron 2021, 150, 103126. [Google Scholar] [CrossRef]
- Kloc, M.; Bilinski, S.; Dougherty, M.T.; Brey, E.M.; Etkin, L.D. Formation, architecture and polarity of female germline cyst in Xenopus. Dev. Biol. 2004, 266, 43–61. [Google Scholar] [CrossRef]
- Nakamura, S.; Kobayashi, K.; Nishimura, T.; Tanaka, M. Ovarian germline stem cells in the teleost fish, medaka (Oryzias latipes). Int. J. Biol. Sci. 2011, 7, 403–409. [Google Scholar] [CrossRef]
- Bertho, S.; Clapp, M.; Banisch, T.U.; Bandemer, J.; Raz, E.; Marlow, F.L. Zebrafish dazl regulates cystogenesis and germline stem cell specification during the primordial germ cell to germline stem cell transition. Development 2021, 148, dev187773. [Google Scholar] [CrossRef]
- Hughes, S.E.; Miller, D.E.; Miller, A.L.; Hawley, R.S. Female Meiosis: Synapsis, Recombination, and Segregation in Drosophila melanogaster. Genetics 2018, 208, 875–908. [Google Scholar] [CrossRef]
- Hinnant, T.D.; Merkle, J.A.; Ables, E.T. Coordinating Proliferation, Polarity, and Cell Fate in the Drosophila Female Germline. Front. Cell Dev. Biol. 2020, 8, 19. [Google Scholar] [CrossRef]
- Kahney, E.W.; Snedeker, J.C.; Chen, X. Regulation of Drosophila germline stem cells. Curr. Opin. Cell Biol. 2019, 60, 27–35. [Google Scholar] [CrossRef]
- Xie, T.; Spradling, A.C. Decapentaplegic is essential for the maintenance and division of germline stem cells in the Drosophila ovary. Cell 1998, 94, 251–260. [Google Scholar] [CrossRef]
- Song, X.; Wong, M.D.; Kawase, E.; Xi, R.; Ding, B.C.; McCarthy, J.J.; Xie, T. Bmp signals from niche cells directly repress transcription of a differentiation-promoting gene, bag of marbles, in germline stem cells in the Drosophila ovary. Development 2004, 131, 1353–1364. [Google Scholar] [CrossRef]
- Wilcockson, S.G.; Ashe, H.L. Drosophila Ovarian Germline Stem Cell Cytocensor Projections Dynamically Receive and Attenuate BMP Signaling. Dev. Cell 2019, 50, 296–312.e5. [Google Scholar] [CrossRef]
- Chen, D.; McKearin, D. Dpp signaling silences bam transcription directly to establish asymmetric divisions of germline stem cells. Curr. Biol. 2003, 13, 1786–1791. [Google Scholar] [CrossRef]
- Morris, L.X.; Spradling, A.C. Steroid signaling within Drosophila ovarian epithelial cells sex-specifically modulates early germ cell development and meiotic entry. PLoS ONE 2012, 7, e46109. [Google Scholar] [CrossRef]
- Eliazer, S.; Palacios, V.; Wang, Z.; Kollipara, R.K.; Kittler, R.; Buszczak, M. Lsd1 restricts the number of germline stem cells by regulating multiple targets in escort cells. PLoS Genet. 2014, 10, e1004200. [Google Scholar] [CrossRef] [PubMed]
- Konig, A.; Shcherbata, H.R. Soma influences GSC progeny differentiation via the cell adhesion-mediated steroid-let-7-Wingless signaling cascade that regulates chromatin dynamics. Biol. Open 2015, 4, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhong, G.; Chai, P.C.; Luo, L.; Liu, S.; Yang, Y.; Baeg, G.H.; Cai, Y. Coordinated niche-associated signals promote germline homeostasis in the Drosophila ovary. J. Cell Biol. 2015, 211, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Wang, S.; Gao, Y.; Mao, Y.; Yang, Z.; Liu, L.; Song, X.; Ni, J.; Xie, T. COP9-Hedgehog axis regulates the function of the germline stem cell progeny differentiation niche in the Drosophila ovary. Development 2015, 142, 4242–4252. [Google Scholar] [CrossRef]
- Mottier-Pavie, V.I.; Palacios, V.; Eliazer, S.; Scoggin, S.; Buszczak, M. The Wnt pathway limits BMP signaling outside of the germline stem cell niche in Drosophila ovaries. Dev. Biol. 2016, 417, 50–62. [Google Scholar] [CrossRef]
- Su, Y.H.; Rastegri, E.; Kao, S.H.; Lai, C.M.; Lin, K.Y.; Liao, H.Y.; Wang, M.H.; Hsu, H.J. Diet regulates membrane extension and survival of niche escort cells for germline homeostasis via insulin signaling. Development 2018, 145, dev159186. [Google Scholar] [CrossRef]
- Wang, X.; Page-McCaw, A. Wnt6 maintains anterior escort cells as an integral component of the germline stem cell niche. Development 2018, 145, dev158527. [Google Scholar] [CrossRef]
- Morris, L.X.; Spradling, A.C. Long-term live imaging provides new insight into stem cell regulation and germline-soma coordination in the Drosophila ovary. Development 2011, 138, 2207–2215. [Google Scholar] [CrossRef]
- Hsu, H.J.; LaFever, L.; Drummond-Barbosa, D. Diet controls normal and tumorous germline stem cells via insulin-dependent and -independent mechanisms in Drosophila. Dev. Biol. 2008, 313, 700–712. [Google Scholar] [CrossRef]
- Ables, E.T.; Drummond-Barbosa, D. Cyclin E controls Drosophila female germline stem cell maintenance independently of its role in proliferation by modulating responsiveness to niche signals. Development 2013, 140, 530–540. [Google Scholar] [CrossRef]
- Wang, Z.; Lin, H. The division of Drosophila germline stem cells and their precursors requires a specific cyclin. Curr. Biol. 2005, 15, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, I.; Lilly, M.A. Bruno inhibits the expression of mitotic cyclins during the prophase I meiotic arrest of Drosophila oocytes. Dev. Cell 2006, 10, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Lilly, M.A.; de Cuevas, M.; Spradling, A.C. Cyclin A associates with the fusome during germline cyst formation in the Drosophila ovary. Dev. Biol. 2000, 218, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.; Tan, C. Germline cyst formation and incomplete cytokinesis during Drosophila melanogaster oogenesis. Dev. Biol. 2010, 337, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.N.; Cooley, L. Stable intercellular bridges in development: The cytoskeleton lining the tunnel. Trends Cell Biol. 1996, 6, 474–479. [Google Scholar] [CrossRef]
- Carpenter, A.T. Egalitarian and the choice of cell fates in Drosophila melanogaster oogenesis. Ciba Found. Symp. 1994, 182, 223–246; discussion 246–254. [Google Scholar]
- Huynh, J.R.; St Johnston, D. The role of BicD, Egl, Orb and the microtubules in the restriction of meiosis to the Drosophila oocyte. Development 2000, 127, 2785–2794. [Google Scholar] [CrossRef]
- Huynh, J.R. Fusome as a Cell-Cell Communication Channel of Drosophila Ovarian Cyst. In Cell-Cell Channels; Baluska, F., Volkmann, D., Barlow, P.W., Eds.; Springer: New York, NY, USA, 2006. [Google Scholar]
- de Cuevas, M.; Spradling, A.C. Morphogenesis of the Drosophila fusome and its implications for oocyte specification. Development 1998, 125, 2781–2789. [Google Scholar] [CrossRef]
- Lin, H.; Yue, L.; Spradling, A.C. The Drosophila fusome, a germline-specific organelle, contains membrane skeletal proteins and functions in cyst formation. Development 1994, 120, 947–956. [Google Scholar] [CrossRef]
- Lin, H.; Spradling, A.C. Fusome asymmetry and oocyte determination in Drosophila. Dev. Genet. 1995, 16, 6–12. [Google Scholar] [CrossRef]
- Nashchekin, D.; Busby, L.; Jakobs, M.; Squires, I.; St Johnston, D. Symmetry breaking in the female germline cyst. Science 2021, 374, 874–879. [Google Scholar] [CrossRef]
- Röper, K.; Brown, N.H. A Spectraplakin Is Enriched on the Fusome and Organizes Microtubules during Oocyte Specification in Drosophila. Curr. Biol. 2004, 14, 99–110. [Google Scholar] [CrossRef]
- Roper, K. Rtnl1 is enriched in a specialized germline ER that associates with ribonucleoprotein granule components. J. Cell Sci. 2007, 120 Pt 6, 1081–1092. [Google Scholar] [CrossRef]
- Snapp, E.L.; Iida, T.; Frescas, D.; Lippincott-Schwartz, J.; Lilly, M.A. The fusome mediates intercellular endoplasmic reticulum connectivity in Drosophila ovarian cysts. Mol. Biol. Cell 2004, 15, 4512–4521. [Google Scholar] [CrossRef]
- Lighthouse, D.V.; Buszczak, M.; Spradling, A.C. New components of the Drosophila fusome suggest it plays novel roles in signaling and transport. Dev. Biol. 2008, 317, 59–71. [Google Scholar] [CrossRef]
- Pielage, J.; Bulat, V.; Zuchero, J.B.; Fetter, R.D.; Davis, G.W. Hts/Adducin controls synaptic elaboration and elimination. Neuron 2011, 69, 1114–1131. [Google Scholar] [CrossRef]
- Yue, L.; Spradling, A.C. hu-li tai shao, a gene required for ring canal formation during Drosophila oogenesis, encodes a homolog of adducin. Genes Dev. 1992, 6, 2443–2454. [Google Scholar] [CrossRef]
- Petrella, L.N.; Smith-Leiker, T.; Cooley, L. The Ovhts polyprotein is cleaved to produce fusome and ring canal proteins required for Drosophila oogenesis. Development 2007, 134, 703–712. [Google Scholar] [CrossRef]
- Huynh, J.R.; Shulman, J.M.; Benton, R.; St Johnston, D. PAR-1 is required for the maintenance of oocyte fate in Drosophila. Development 2001, 128, 1201–1209. [Google Scholar] [CrossRef]
- Bolivar, J.; Huynh, J.R.; Lopez-Schier, H.; Gonzalez, C.; St Johnston, D.; Gonzalez-Reyes, A. Centrosome migration into the Drosophila oocyte is independent of BicD and egl, and of the organisation of the microtubule cytoskeleton. Development 2001, 128, 1889–1897. [Google Scholar] [CrossRef]
- Cox, R.T.; Spradling, A.C. A Balbiani body and the fusome mediate mitochondrial inheritance during Drosophila oogenesis. Development 2003, 130, 1579–1590. [Google Scholar] [CrossRef] [PubMed]
- Wharton, R.P.; Struhl, G. Structure of the Drosophila BicaudalD protein and its role in localizing the posterior determinant nanos. Cell 1989, 59, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Suter, B.; Romberg, L.M.; Steward, R. Bicaudal-D, a Drosophila gene involved in developmental asymmetry: Localized transcript accumulation in ovaries and sequence similarity to myosin heavy chain tail domains. Genes Dev. 1989, 3, 1957–1968. [Google Scholar] [CrossRef] [PubMed]
- Mach, J.M.; Lehmann, R. An Egalitarian-BicaudalD complex is essential for oocyte specification and axis determination in Drosophila. Genes Dev. 1997, 11, 423–435. [Google Scholar] [CrossRef]
- Navarro, C.; Puthalakath, H.; Adams, J.M.; Strasser, A.; Lehmann, R. Egalitarian binds dynein light chain to establish oocyte polarity and maintain oocyte fate. Nat. Cell Biol. 2004, 6, 427–435. [Google Scholar] [CrossRef]
- Ephrussi, A.; Dickinson, L.K.; Lehmann, R. Oskar organizes the germ plasm and directs localization of the posterior determinant nanos. Cell 1991, 66, 37–50. [Google Scholar] [CrossRef]
- Lantz, V.; Chang, J.S.; Horabin, J.I.; Bopp, D.; Schedl, P. The Drosophila orb RNA-binding protein is required for the formation of the egg chamber and establishment of polarity. Genes Dev. 1994, 8, 598–613. [Google Scholar] [CrossRef]
- Wong, L.C.; Schedl, P. Cup blocks the precocious activation of the orb autoregulatory loop. PLoS ONE 2011, 6, e28261. [Google Scholar] [CrossRef]
- Christerson, L.B.; McKearin, D.M. orb is required for anteroposterior and dorsoventral patterning during Drosophila oogenesis. Genes Dev. 1994, 8, 614–628. [Google Scholar] [CrossRef]
- Chang, J.S.; Tan, L.; Schedl, P. The Drosophila CPEB homolog, orb, is required for oskar protein expression in oocytes. Dev. Biol. 1999, 215, 91–106. [Google Scholar] [CrossRef]
- Chang, J.S.; Tan, L.; Wolf, M.R.; Schedl, P. Functioning of the Drosophila orb gene in gurken mRNA localization and translation. Development 2001, 128, 3169–3177. [Google Scholar] [CrossRef]
- Castagnetti, S.; Ephrussi, A. Orb and a long poly(A) tail are required for efficient oskar translation at the posterior pole of the Drosophila oocyte. Development 2003, 130, 835–843. [Google Scholar] [CrossRef]
- Tan, L.; Chang, J.S.; Costa, A.; Schedl, P. An autoregulatory feedback loop directs the localized expression of the Drosophila CPEB protein Orb in the developing oocyte. Development 2001, 128, 1159–1169. [Google Scholar] [CrossRef]
- Barr, J.; Gilmutdinov, R.; Wang, L.; Shidlovskii, Y.; Schedl, P. The Drosophila CPEB Protein Orb Specifies Oocyte Fate by a 3’UTR-Dependent Autoregulatory Loop. Genetics 2019, 213, 1431–1446. [Google Scholar] [CrossRef]
- Rubin, T.; Christophorou, N.; Huynh, J.R. How to pre-pair chromosomes for meiosis. Cell Cycle 2016, 15, 609–610. [Google Scholar] [CrossRef]
- Christophorou, N.; Rubin, T.; Huynh, J.R. Synaptonemal complex components promote centromere pairing in pre-meiotic germ cells. PLoS Genet. 2013, 9, e1004012. [Google Scholar] [CrossRef]
- Rust, K.; Byrnes, L.E.; Yu, K.S.; Park, J.S.; Sneddon, J.B.; Tward, A.D.; Nystul, T.G. A single-cell atlas and lineage analysis of the adult Drosophila ovary. Nat. Commun. 2020, 11, 5628. [Google Scholar] [CrossRef]
- Slaidina, M.; Lehmann, R. Translational control in germline stem cell development. J. Cell Biol. 2014, 207, 13–21. [Google Scholar] [CrossRef]
- Lin, H.; Spradling, A.C. A novel group of pumilio mutations affects the asymmetric division of germline stem cells in the Drosophila ovary. Development 1997, 124, 2463–2476. [Google Scholar] [CrossRef]
- Forbes, A.; Lehmann, R. Nanos and Pumilio have critical roles in the development and function of Drosophila germline stem cells. Development 1998, 125, 679–690. [Google Scholar] [CrossRef]
- Gilboa, L.; Lehmann, R. Repression of primordial germ cell differentiation parallels germ line stem cell maintenance. Curr. Biol. 2004, 14, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Joly, W.; Chartier, A.; Rojas-Rios, P.; Busseau, I.; Simonelig, M. The CCR4 deadenylase acts with Nanos and Pumilio in the fine-tuning of Mei-P26 expression to promote germline stem cell self-renewal. Stem Cell Rep. 2013, 1, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Neumuller, R.A.; Betschinger, J.; Fischer, A.; Bushati, N.; Poernbacher, I.; Mechtler, K.; Cohen, S.M.; Knoblich, J.A. Mei-P26 regulates microRNAs and cell growth in the Drosophila ovarian stem cell lineage. Nature 2008, 454, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.E.; Pargett, M.; Sutcliffe, C.; Umulis, D.; Ashe, H.L. Brat promotes stem cell differentiation via control of a bistable switch that restricts BMP signaling. Dev. Cell 2011, 20, 72–83. [Google Scholar] [CrossRef]
- Li, Y.; Minor, N.T.; Park, J.K.; McKearin, D.M.; Maines, J.Z. Bam and Bgcn antagonize Nanos-dependent germ-line stem cell maintenance. Proc. Natl. Acad. Sci. USA 2009, 106, 9304–9309. [Google Scholar] [CrossRef]
- Chau, J.; Kulnane, L.S.; Salz, H.K. Sex-lethal enables germline stem cell differentiation by down-regulating Nanos protein levels during Drosophila oogenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 9465–9470. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Q.; Carreira-Rosario, A.; Maines, J.Z.; McKearin, D.M.; Buszczak, M. Mei-p26 cooperates with Bam, Bgcn and Sxl to promote early germline development in the Drosophila ovary. PLoS ONE 2013, 8, e58301. [Google Scholar] [CrossRef]
- Carreira-Rosario, A.; Bhargava, V.; Hillebrand, J.; Kollipara, R.K.; Ramaswami, M.; Buszczak, M. Repression of Pumilio Protein Expression by Rbfox1 Promotes Germ Cell Differentiation. Dev. Cell 2016, 36, 562–571. [Google Scholar] [CrossRef]
- Navarro-Costa, P.; McCarthy, A.; Prudencio, P.; Greer, C.; Guilgur, L.G.; Becker, J.D.; Secombe, J.; Rangan, P.; Martinho, R.G. Early programming of the oocyte epigenome temporally controls late prophase I transcription and chromatin remodelling. Nat. Commun. 2016, 7, 12331. [Google Scholar] [CrossRef]
- Vidaurre, V.; Chen, X. Epigenetic regulation of Drosophila germline stem cell maintenance and differentiation. Dev. Biol. 2021, 473, 105–118. [Google Scholar] [CrossRef]
- Schuettengruber, B.; Bourbon, H.M.; Di Croce, L.; Cavalli, G. Genome Regulation by Polycomb and Trithorax: 70 Years and Counting. Cell 2017, 171, 34–57. [Google Scholar] [CrossRef]
- Kassis, J.A.; Kennison, J.A.; Tamkun, J.W. Polycomb and Trithorax Group Genes in Drosophila. Genetics 2017, 206, 1699–1725. [Google Scholar] [CrossRef]
- Blackledge, N.P.; Klose, R.J. The molecular principles of gene regulation by Polycomb repressive complexes. Nat. Rev. Mol. Cell Biol. 2021, 22, 815–833. [Google Scholar] [CrossRef]
- Li, X.Y.; Harrison, M.M.; Villalta, J.E.; Kaplan, T.; Eisen, M.B. Establishment of regions of genomic activity during the Drosophila maternal to zygotic transition. eLife 2014, 3, e03737. [Google Scholar] [CrossRef]
- Liu, X.; Wang, C.; Liu, W.; Li, J.; Li, C.; Kou, X.; Chen, J.; Zhao, Y.; Gao, H.; Wang, H.; et al. Distinct features of H3K4me3 and H3K27me3 chromatin domains in pre-implantation embryos. Nature 2016, 537, 558–562. [Google Scholar] [CrossRef]
- DeLuca, S.Z.; Ghildiyal, M.; Pang, L.Y.; Spradling, A.C. Differentiating Drosophila female germ cells initiate Polycomb silencing by regulating PRC2-interacting proteins. eLife 2020, 9, e56922. [Google Scholar] [CrossRef] [PubMed]
- Klymenko, T.; Papp, B.; Fischle, W.; Kocher, T.; Schelder, M.; Fritsch, C.; Wild, B.; Wilm, M.; Muller, J. A Polycomb group protein complex with sequence-specific DNA-binding and selective methyl-lysine-binding activities. Genes Dev. 2006, 20, 1110–1122. [Google Scholar] [CrossRef]
- Alfieri, C.; Gambetta, M.C.; Matos, R.; Glatt, S.; Sehr, P.; Fraterman, S.; Wilm, M.; Muller, J.; Muller, C.W. Structural basis for targeting the chromatin repressor Sfmbt to Polycomb response elements. Genes Dev. 2013, 27, 2367–2379. [Google Scholar] [CrossRef]
- Kang, H.; McElroy, K.A.; Jung, Y.L.; Alekseyenko, A.A.; Zee, B.M.; Park, P.J.; Kuroda, M.I. Sex comb on midleg (Scm) is a functional link between PcG-repressive complexes in Drosophila. Genes Dev. 2015, 29, 1136–1150. [Google Scholar] [CrossRef]
- Frey, F.; Sheahan, T.; Finkl, K.; Stoehr, G.; Mann, M.; Benda, C.; Muller, J. Molecular basis of PRC1 targeting to Polycomb response elements by PhoRC. Genes Dev. 2016, 30, 1116–1127. [Google Scholar] [CrossRef]
- Birve, A.; Sengupta, A.K.; Beuchle, D.; Larsson, J.; Kennison, J.A.; Rasmuson-Lestander, A.; Muller, J. Su(z)12, a novel Drosophila Polycomb group gene that is conserved in vertebrates and plants. Development 2001, 128, 3371–3379. [Google Scholar] [CrossRef] [PubMed]
- Iovino, N.; Ciabrelli, F.; Cavalli, G. PRC2 controls Drosophila oocyte cell fate by repressing cell cycle genes. Dev. Cell 2013, 26, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Feijao, T.; Marques, B.; Silva, R.D.; Carvalho, C.; Sobral, D.; Matos, R.; Tan, T.; Pereira, A.; Morais-de-Sa, E.; Maiato, H.; et al. Polycomb group (PcG) proteins prevent the assembly of abnormal synaptonemal complex structures during meiosis. Proc. Natl. Acad. Sci. USA 2022, 119, e2204701119. [Google Scholar] [CrossRef] [PubMed]
- Clough, E.; Moon, W.; Wang, S.; Smith, K.; Hazelrigg, T. Histone methylation is required for oogenesis in Drosophila. Development 2007, 134, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Lee, K.S.; Park, J.S.; Yu, K.; Paik, S.G.; Kang, Y.K. dSETDB1 and SU(VAR)3-9 sequentially function during germline-stem cell differentiation in Drosophila melanogaster. PLoS ONE 2008, 3, e2234. [Google Scholar] [CrossRef]
- Rangan, P.; Malone, C.D.; Navarro, C.; Newbold, S.P.; Hayes, P.S.; Sachidanandam, R.; Hannon, G.J.; Lehmann, R. piRNA production requires heterochromatin formation in Drosophila. Curr. Biol. 2011, 21, 1373–1379. [Google Scholar] [CrossRef]
- Shokri, L.; Inukai, S.; Hafner, A.; Weinand, K.; Hens, K.; Vedenko, A.; Gisselbrecht, S.S.; Dainese, R.; Bischof, J.; Furger, E.; et al. A Comprehensive Drosophila melanogaster Transcription Factor Interactome. Cell Rep. 2019, 27, 955–970.e7. [Google Scholar] [CrossRef]
- Yi, X.; de Vries, H.I.; Siudeja, K.; Rana, A.; Lemstra, W.; Brunsting, J.F.; Kok, R.M.; Smulders, Y.M.; Schaefer, M.; Dijk, F.; et al. Stwl modifies chromatin compaction and is required to maintain DNA integrity in the presence of perturbed DNA replication. Mol. Biol. Cell 2009, 20, 983–994. [Google Scholar] [CrossRef]
- Maines, J.Z.; Park, J.K.; Williams, M.; McKearin, D.M. Stonewalling Drosophila stem cell differentiation by epigenetic controls. Development 2007, 134, 1471–1479. [Google Scholar] [CrossRef]
- Zinshteyn, D.; Barbash, D.A. Stonewall prevents expression of ectopic genes in the ovary and accumulates at insulator elements in D. melanogaster. PLoS Genet. 2022, 18, e1010110. [Google Scholar] [CrossRef]
- Wood, A.; Krogan, N.J.; Dover, J.; Schneider, J.; Heidt, J.; Boateng, M.A.; Dean, K.; Golshani, A.; Zhang, Y.; Greenblatt, J.F.; et al. Bre1, an E3 ubiquitin ligase required for recruitment and substrate selection of Rad6 at a promoter. Mol. Cell 2003, 11, 267–274. [Google Scholar] [CrossRef]
- Bray, S.; Musisi, H.; Bienz, M. Bre1 is required for Notch signaling and histone modification. Dev. Cell 2005, 8, 279–286. [Google Scholar] [CrossRef]
- Mohan, M.; Herz, H.M.; Smith, E.R.; Zhang, Y.; Jackson, J.; Washburn, M.P.; Florens, L.; Eissenberg, J.C.; Shilatifard, A. The COMPASS family of H3K4 methylases in Drosophila. Mol. Cell. Biol. 2011, 31, 4310–4318. [Google Scholar] [CrossRef]
- Xuan, T.; Xin, T.; He, J.; Tan, J.; Gao, Y.; Feng, S.; He, L.; Zhao, G.; Li, M. dBre1/dSet1-dependent pathway for histone H3K4 trimethylation has essential roles in controlling germline stem cell maintenance and germ cell differentiation in the Drosophila ovary. Dev. Biol. 2013, 379, 167–181. [Google Scholar] [CrossRef]
- Prudencio, P.; Guilgur, L.G.; Sobral, J.; Becker, J.D.; Martinho, R.G.; Navarro-Costa, P. The Trithorax group protein dMLL3/4 instructs the assembly of the zygotic genome at fertilization. EMBO Rep. 2018, 19, e45728. [Google Scholar] [CrossRef]
- Navarro-Costa, P.; Martinho, R.G. The emerging role of transcriptional regulation in the oocyte-to-zygote transition. PLoS Genet. 2020, 16, e1008602. [Google Scholar] [CrossRef]
- Mukai, M.; Hira, S.; Nakamura, K.; Nakamura, S.; Kimura, H.; Sato, M.; Kobayashi, S. H3K36 Trimethylation-Mediated Epigenetic Regulation is Activated by Bam and Promotes Germ Cell Differentiation During Early Oogenesis in Drosophila. Biol. Open 2015, 4, 119–124. [Google Scholar] [CrossRef]
- McCarthy, A.; Sarkar, K.; Martin, E.T.; Upadhyay, M.; Jang, S.; Williams, N.D.; Forni, P.E.; Buszczak, M.; Rangan, P. Msl3 promotes germline stem cell differentiation in female Drosophila. Development 2022, 149, dev199625. [Google Scholar] [CrossRef]
- Stabell, M.; Larsson, J.; Aalen, R.B.; Lambertsson, A. Drosophila dSet2 functions in H3-K36 methylation and is required for development. Biochem. Biophys. Res. Commun. 2007, 359, 784–789. [Google Scholar] [CrossRef]
- Larschan, E.; Alekseyenko, A.A.; Gortchakov, A.A.; Peng, S.; Li, B.; Yang, P.; Workman, J.L.; Park, P.J.; Kuroda, M.I. MSL complex is attracted to genes marked by H3K36 trimethylation using a sequence-independent mechanism. Mol. Cell 2007, 28, 121–133. [Google Scholar] [CrossRef]
- Drelon, C.; Belalcazar, H.M.; Secombe, J. The Histone Demethylase KDM5 Is Essential for Larval Growth in Drosophila. Genetics 2018, 209, 773–787. [Google Scholar] [CrossRef] [PubMed]
- Drelon, C.; Rogers, M.F.; Belalcazar, H.M.; Secombe, J. The histone demethylase KDM5 controls developmental timing in Drosophila by promoting prothoracic gland endocycles. Development 2019, 146, dev182568. [Google Scholar] [CrossRef] [PubMed]
- Lloret-Llinares, M.; Perez-Lluch, S.; Rossell, D.; Moran, T.; Ponsa-Cobas, J.; Auer, H.; Corominas, M.; Azorin, F. dKDM5/LID regulates H3K4me3 dynamics at the transcription-start site (TSS) of actively transcribed developmental genes. Nucleic Acids Res. 2012, 40, 9493–9505. [Google Scholar] [CrossRef] [PubMed]
- Zhaunova, L.; Ohkura, H.; Breuer, M. Kdm5/Lid Regulates Chromosome Architecture in Meiotic Prophase I Independently of Its Histone Demethylase Activity. PLoS Genet. 2016, 12, e1006241. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Secombe, J. The Histone Demethylase KDM5 Activates Gene Expression by Recognizing Chromatin Context through Its PHD Reader Motif. Cell Rep. 2015, 13, 2219–2231. [Google Scholar] [CrossRef]
- Liu, X.; Greer, C.; Secombe, J. KDM5 interacts with Foxo to modulate cellular levels of oxidative stress. PLoS Genet. 2014, 10, e1004676. [Google Scholar] [CrossRef]
- Gajan, A.; Barnes, V.L.; Liu, M.; Saha, N.; Pile, L.A. The histone demethylase dKDM5/LID interacts with the SIN3 histone deacetylase complex and shares functional similarities with SIN3. Epigenet. Chromatin 2016, 9, 4. [Google Scholar] [CrossRef]
- Torres-Campana, D.; Kimura, S.; Orsi, G.A.; Horard, B.; Benoit, G.; Loppin, B. The Lid/KDM5 histone demethylase complex activates a critical effector of the oocyte-to-zygote transition. PLoS Genet. 2020, 16, e1008543. [Google Scholar] [CrossRef]
- Nieken, K.J.; O’Brien, K.; McDonnell, A.; Zhaunova, L.; Ohkura, H. A large-scale RNAi screen reveals that mitochondrial function is important for meiotic chromosome organization in oocytes. Chromosoma 2023, 132, 1–18. [Google Scholar] [CrossRef]
- Xi, R.; Xie, T. Stem cell self-renewal controlled by chromatin remodeling factors. Science 2005, 310, 1487–1489. [Google Scholar] [CrossRef]
- Buszczak, M.; Paterno, S.; Spradling, A.C. Drosophila stem cells share a common requirement for the histone H2B ubiquitin protease scrawny. Science 2009, 323, 248–251. [Google Scholar] [CrossRef]
- Fuchs, G.; Hollander, D.; Voichek, Y.; Ast, G.; Oren, M. Cotranscriptional histone H2B monoubiquitylation is tightly coupled with RNA polymerase II elongation rate. Genome Res. 2014, 24, 1572–1583. [Google Scholar] [CrossRef]
- Neumuller, R.A.; Wirtz-Peitz, F.; Lee, S.; Kwon, Y.; Buckner, M.; Hoskins, R.A.; Venken, K.J.; Bellen, H.J.; Mohr, S.E.; Perrimon, N. Stringent analysis of gene function and protein-protein interactions using fluorescently tagged genes. Genetics 2012, 190, 931–940. [Google Scholar] [CrossRef]
- Ardehali, M.B.; Yao, J.; Adelman, K.; Fuda, N.J.; Petesch, S.J.; Webb, W.W.; Lis, J.T. Spt6 enhances the elongation rate of RNA polymerase II in vivo. EMBO J. 2009, 28, 1067–1077. [Google Scholar] [CrossRef]
- He, J.; Xuan, T.; Xin, T.; An, H.; Wang, J.; Zhao, G.; Li, M. Evidence for chromatin-remodeling complex PBAP-controlled maintenance of the Drosophila ovarian germline stem cells. PLoS ONE 2014, 9, e103473. [Google Scholar] [CrossRef]
- Mondragon, A.A.; Yalonetskaya, A.; Ortega, A.J.; Zhang, Y.; Naranjo, O.; Elguero, J.; Chung, W.S.; McCall, K. Lysosomal Machinery Drives Extracellular Acidification to Direct Non-apoptotic Cell Death. Cell Rep. 2019, 27, 11–19.e3. [Google Scholar] [CrossRef]
- Seki, Y.; Yamaji, M.; Yabuta, Y.; Sano, M.; Shigeta, M.; Matsui, Y.; Saga, Y.; Tachibana, M.; Shinkai, Y.; Saitou, M. Cellular dynamics associated with the genome-wide epigenetic reprogramming in migrating primordial germ cells in mice. Development 2007, 134, 2627–2638. [Google Scholar] [CrossRef]
- Seisenberger, S.; Andrews, S.; Krueger, F.; Arand, J.; Walter, J.; Santos, F.; Popp, C.; Thienpont, B.; Dean, W.; Reik, W. The dynamics of genome-wide DNA methylation reprogramming in mouse primordial germ cells. Mol. Cell 2012, 48, 849–862. [Google Scholar] [CrossRef]
- Seki, Y.; Hayashi, K.; Itoh, K.; Mizugaki, M.; Saitou, M.; Matsui, Y. Extensive and orderly reprogramming of genome-wide chromatin modifications associated with specification and early development of germ cells in mice. Dev. Biol. 2005, 278, 440–458. [Google Scholar] [CrossRef]
- Staubli, A.; Peters, A.H. Mechanisms of maternal intergenerational epigenetic inheritance. Curr. Opin. Genet. Dev. 2021, 67, 151–162. [Google Scholar] [CrossRef]
- Inoue, A.; Jiang, L.; Lu, F.; Suzuki, T.; Zhang, Y. Maternal H3K27me3 controls DNA methylation-independent imprinting. Nature 2017, 547, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yin, Q.; Inoue, A.; Zhang, C.; Zhang, Y. Allelic H3K27me3 to allelic DNA methylation switch maintains noncanonical imprinting in extraembryonic cells. Sci. Adv. 2019, 5, eaay7246. [Google Scholar] [CrossRef] [PubMed]
- Mei, H.; Kozuka, C.; Hayashi, R.; Kumon, M.; Koseki, H.; Inoue, A. H2AK119ub1 guides maternal inheritance and zygotic deposition of H3K27me3 in mouse embryos. Nat. Genet. 2021, 53, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Zenk, F.; Loeser, E.; Schiavo, R.; Kilpert, F.; Bogdanovic, O.; Iovino, N. Germ line-inherited H3K27me3 restricts enhancer function during maternal-to-zygotic transition. Science 2017, 357, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Samata, M.; Alexiadis, A.; Richard, G.; Georgiev, P.; Nuebler, J.; Kulkarni, T.; Renschler, G.; Basilicata, M.F.; Zenk, F.L.; Shvedunova, M.; et al. Intergenerationally Maintained Histone H4 Lysine 16 Acetylation Is Instructive for Future Gene Activation. Cell 2020, 182, 127–144.e23. [Google Scholar] [CrossRef]
- Lyko, F.; Ramsahoye, B.H.; Jaenisch, R. DNA methylation in Drosophila melanogaster. Nature 2000, 408, 538–540. [Google Scholar] [CrossRef]
- Church, S.H.; de Medeiros, B.A.S.; Donoughe, S.; Márquez Reyes, N.L.; Extavour, C.G. Repeated loss of variation in insect ovary morphology highlights the role of development in life-history evolution. Proc. R. Soc. B Biol. Sci. 2021, 288, 20210150. [Google Scholar] [CrossRef]
- Munro, C.; Cadis, H.; Pagnotta, S.; Houliston, E.; Huynh, J.R. Conserved meiotic mechanisms in the cnidarian Clytia hemisphaerica revealed by Spo11 knockout. Sci. Adv. 2023, 9, eadd2873. [Google Scholar] [CrossRef]
- Miramon-Puertolas, P.; Steinmetz, P.R.H. An adult stem-like cell population generates germline and neurons in the sea anemone Nematostella vectensis. bioRxiv 2023. [Google Scholar] [CrossRef]
- Saitou, M.; Hayashi, K. Mammalian in vitro gametogenesis. Science 2021, 374, eaaz6830. [Google Scholar] [CrossRef]
- Aizawa, E.; Ozonov, E.A.; Kawamura, Y.K.; Dumeau, C.-E.; Nagaoka, S.; Saitou, M.; Peters, A.H.F.M.; Wutz, A. Comprehensive comparison of female germ cell development in vitro and in vivo identifies epigenetic gene regulation crucial for oocyte development and embryonic competence. bioRxiv 2023. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabrita, B.; Martinho, R.G. Genetic and Epigenetic Regulation of Drosophila Oocyte Determination. J. Dev. Biol. 2023, 11, 21. https://doi.org/10.3390/jdb11020021
Cabrita B, Martinho RG. Genetic and Epigenetic Regulation of Drosophila Oocyte Determination. Journal of Developmental Biology. 2023; 11(2):21. https://doi.org/10.3390/jdb11020021
Chicago/Turabian StyleCabrita, Brigite, and Rui Gonçalo Martinho. 2023. "Genetic and Epigenetic Regulation of Drosophila Oocyte Determination" Journal of Developmental Biology 11, no. 2: 21. https://doi.org/10.3390/jdb11020021
APA StyleCabrita, B., & Martinho, R. G. (2023). Genetic and Epigenetic Regulation of Drosophila Oocyte Determination. Journal of Developmental Biology, 11(2), 21. https://doi.org/10.3390/jdb11020021