
A Review of Delayed Delivery Models and the Analysis Method in Mice


Abstract
1. Introduction
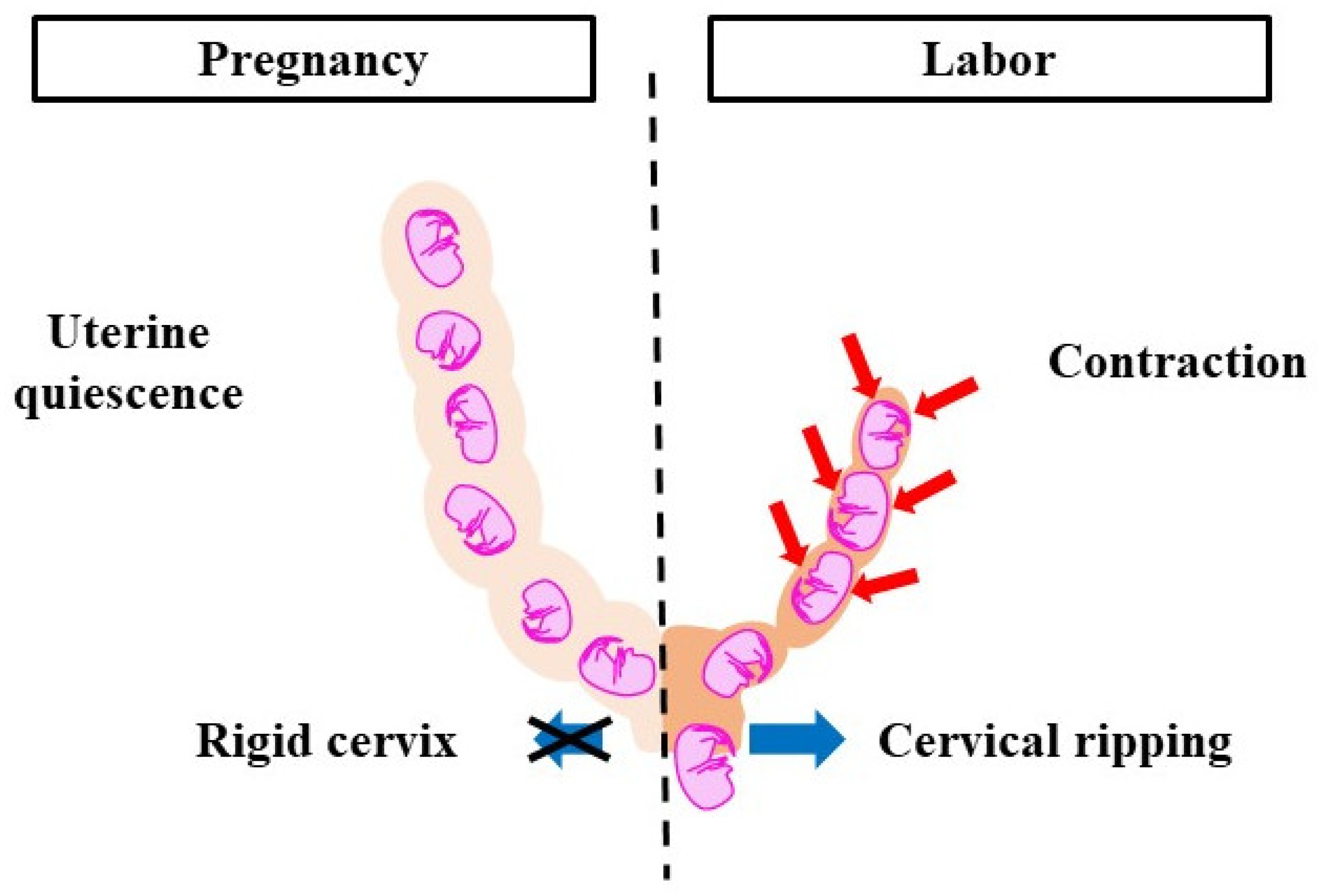
2. Normal Progression of Pregnancy and Labor Onset in Mice
3. Factors of Pregnancy Maintenance and Labor Induction
4. Classification by Cause of Labor Delay
4.1. Failure of Luteolysis
4.1.1. Prostaglandin F Receptor Knockout Mouse: Fp KO
4.1.2. Cox1-Deficient Mouse: Cox1 KO
4.1.3. 20α Hydroxysteroid Dehydrogenase-Deficient Mice: 20α Hsd KO
4.1.4. Gαqf/f;Gα11−/−;Cre+ Mice: Gq/11 cKO
4.1.5. Mastermind-Like Domain-Containing 1 Knockout Mouse: Mamld1 KO
4.2. Abnormalities in P₄ Metabolism and Receptors
4.2.1. Kruppel-Like Factor 9 Knockout Mouse: Klf 9 KO
4.2.2. Cytochrome p450 Family 11a1-Overexpression Mouse: Cyp11a1 Tg
4.3. Fetal Factors
4.3.1. Surfactant Protein A and D Double-Deficient Mice: Sp A/D dKO
4.3.2. Steroid Receptor Coactivator 1 and 2 Double-Deficient Mice: Src1/2 dKO
4.4. Placental Factors
4.4.1. Solute Carrier Organic Anion Transporter Family Member 2A1 Knockout Mouse: Slco2a1 KO
4.4.2. Sushi Ichi Retrotransposon Homolog 7/Leucine Zipper, Downregulated Cancer 1 Knockout Mice: Sirh7/Ldoc1 KO
4.4.3. Nik-Related Kinase Knockout Mouse: Nrk KO
4.5. Autoimmune Disorder
4.5.1. Toll-Like Receptor 2-Deficient Mice: Tlr2 KO
4.5.2. Toll-Like Receptor 4-Deficient Mice: Tlr4 KO
4.5.3. Interleukin 6 Null Mutant Mice: Il6 KO
4.6. Uterine Contractile Dysfunction
4.6.1. Connexin 43fl/fl:SM-CreERT2 Mice: Sm-CreERT2 KO
4.6.2. Kcnn3tm1Jpad/Kcnn3tm1jpad: Sk3T/T Mice
4.7. Poor Cervical Ripening
4.7.1. 5α-Reductase Type 1 Knockout Mice: 5αR1 KO
4.7.2. TgN(hApoB) 1102SY Line: Tg/Tg Mouse
4.7.3. Anthrax Toxin Receptor 2 Knockout Mice: Antxr2 KO
4.8. Delayed Implantation
4.8.1. Cytosolic Phosphatase A2 Mutant Mouse: Pla2g4a KO
4.8.2. Lysophosphatidic Acid Receptor 3-Deficient Mouse: LPA3 KO
4.9. Unknown Cause
4.9.1. Erk1 and Erk2 Conditional Knockout Mouse: Erk1/2 cKO
4.9.2. Leptin KO Mouse: Leptin KO
4.9.3. MT-mER Mouse
4.9.4. Activin/Inhibin βB Subunit-Deficient Mouse: Activin/Inhibin βB Subunit dKO
5. How to Identify the Cause of Delayed Labor in Mice
5.1. Measurement of the Landing Time
5.2. Investigation of the Following Factors: Dam, Fetus, and Placenta
5.3. Presence or Absence of P₄ Withdrawal during Late Pregnancy
5.4. Evaluation of Uterine Contractions and Cervical Ripening
5.5. Comparison of Wild-Type Mice
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Vayssiere, C.; Haumonte, J.B.; Chantry, A.; Coatleven, F.; Debord, M.P.; Gomez, C.; le Ray, C.; Lopez, E.; Salomon, L.J.; Senat, M.V.; et al. Prolonged and post-term pregnancies: Guidelines for clinical practice from the French College of Gynecologists and Obstetricians (CNGOF). Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 169, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Nishimori, K.; Young, L.J.; Guo, Q.; Wang, Z.; Insel, T.R.; Matzuk, M.M. Oxytocin is required for nursing but is not essential for parturition or reproductive behavior. Proc. Natl Acad. Sci. USA 1996, 93, 11699–11704. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Luedke, C.E.; Vogt, S.K.; Muglia, L.J. Oxytocin modulates the onset of murine parturition by competing ovarian and uterine effects. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R1061–R1067. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, M.; Nishimura, T.; Nakanishi, T.; Shimada, H.; Noguchi, S.; Akanuma, S.I.; Tachikawa, M.; Hosoya, K.I.; Tamai, I.; Nakashima, E.; et al. Contribution of prostaglandin transporter OATP2A1/SLCO2A1 to placenta-to-maternal hormone signaling and labor induction. iScience 2020, 23, 101098. [Google Scholar] [CrossRef]
- Sugimoto, Y.; Yamasaki, A.; Segi, E.; Tsuboi, K.; Aze, Y.; Nishimura, T.; Oida, H.; Yoshida, N.; Tanaka, T.; Katsuyama, M.; et al. Failure of parturition in mice lacking the prostaglandin F receptor. Science 1997, 277, 681–683. [Google Scholar] [CrossRef]
- Tsuboi, K.; Iwane, A.; Nakazawa, S.; Sugimoto, Y.; Ichikawa, A. Role of prostaglandin H2 synthase 2 in murine parturition: Study on ovariectomy-induced parturition in prostaglandin F receptor-deficient mice. Biol. Reprod. 2003, 69, 195–201. [Google Scholar] [CrossRef]
- Gross, G.A.; Imamura, T.; Luedke, C.; Vogt, S.K.; Olson, L.M.; Nelson, D.M.; Sadovsky, Y.; Muglia, L.J. Opposing actions of prostaglandins and oxytocin determine the onset of murine labor. Proc. Natl Acad. Sci. USA 1998, 95, 11875–11879. [Google Scholar] [CrossRef]
- Piekorz, R.P.; Gingras, S.; Hoffmeyer, A.; Ihle, J.N.; Weinstein, Y. Regulation of progesterone levels during pregnancy and parturition by signal transducer and activator of transcription 5 and 20α-hydroxysteroid dehydrogenase. Mol. Endocrinol. 2005, 19, 431–440. [Google Scholar] [CrossRef]
- Mejia, R.; Waite, C.; Ascoli, M. Activation of Gq/11 in the mouse corpus luteum is required for parturition. Mol. Endocrinol. 2015, 29, 238–246. [Google Scholar] [CrossRef]
- Miyado, M.; Miyado, K.; Katsumi, M.; Saito, K.; Nakamura, A.; Shihara, D.; Ogata, T.; Fukami, M. Parturition failure in mice lacking Mamld1. Sci. Rep. 2015, 5, 14705. [Google Scholar] [CrossRef]
- Zeng, Z.; Velarde, M.C.; Simmen, F.A.; Simmen, R.C. Delayed parturition and altered myometrial progesterone receptor isoform A expression in mice null for Kruppel-like factor 9. Biol. Reprod. 2008, 78, 1029–1037. [Google Scholar] [CrossRef]
- Chien, Y.; Cheng, W.C.; Wu, M.R.; Jiang, S.T.; Shen, C.K.; Chung, B.C. Misregulated progesterone secretion and impaired pregnancy in Cyp11a1 transgenic mice. Biol. Reprod. 2013, 89, 91. [Google Scholar] [CrossRef]
- Montalbano, A.P.; Hawgood, S.; Mendelson, C.R. Mice deficient in surfactant protein A (SP-A) and SP-D or in TLR2 manifest delayed parturition and decreased expression of inflammatory and contractile genes. Endocrinology 2013, 154, 483–498. [Google Scholar] [CrossRef]
- Gao, L.; Rabbitt, E.H.; Condon, J.C.; Renthal, N.E.; Johnston, J.M.; Mitsche, M.A.; Chambon, P.; Xu, J.; O’Malley, B.W.; Mendelson, C.R. Steroid receptor coactivators 1 and 2 mediate fetal-to-maternal signaling that initiates parturition. J. Clin. Investig. 2015, 125, 2808–2824. [Google Scholar] [CrossRef]
- Naruse, M.; Ono, R.; Irie, M.; Nakamura, K.; Furuse, T.; Hino, T.; Oda, K.; Kashimura, M.; Yamada, I.; Wakana, S.; et al. Sirh7/Ldoc1 knockout mice exhibit placental P4 overproduction and delayed parturition. Development 2014, 141, 4763–4771. [Google Scholar] [CrossRef]
- Denda, K.; Nakao-Wakabayashi, K.; Okamoto, N.; Kitamura, N.; Ryu, J.Y.; Tagawa, Y.I.; Ichisaka, T.; Yamanaka, S.; Komada, M. Nrk, an X-linked protein kinase in the germinal center kinase family, is required for placental development and fetoplacental induction of labor. J. Biol. Chem. 2011, 286, 28802–28810. [Google Scholar] [CrossRef]
- Wahid, H.H.; Dorian, C.L.; Chin, P.Y.; Hutchinson, M.R.; Rice, K.C.; Olson, D.M.; Moldenhauer, L.M.; Robertson, S.A. Toll-like receptor 4 is an essential upstream regulator of on-time parturition and perinatal viability in mice. Endocrinology 2015, 156, 3828–3841. [Google Scholar] [CrossRef]
- Robertson, S.A.; Christiaens, I.; Dorian, C.L.; Zaragoza, D.B.; Care, A.S.; Banks, A.M.; Olson, D.M. Interleukin-6 is an essential determinant of on-time parturition in the mouse. Endocrinology 2010, 151, 3996–4006. [Google Scholar] [CrossRef]
- Döring, B.; Shynlova, O.; Tsui, P.; Eckardt, D.; Janssen-Bienhold, U.; Hofmann, F.; Feil, S.; Feil, R.; Lye, S.J.; Willecke, K. Ablation of connexin43 in uterine smooth muscle cells of the mouse causes delayed parturition. J. Cell Sci. 2006, 119, 1715–1722. [Google Scholar] [CrossRef]
- Pierce, S.L.; Kresowik, J.D.; Lamping, K.G.; England, S.K. Overexpression of SK3 channels dampens uterine contractility to prevent preterm labor in mice. Biol. Reprod. 2008, 78, 1058–1063. [Google Scholar] [CrossRef]
- Reeves, C.V.; Wang, X.; Charles-Horvath, P.C.; Vink, J.Y.; Borisenko, V.Y.; Young, J.A.; Kitajewski, J.K. Anthrax toxin receptor 2 functions in ECM homeostasis of the murine reproductive tract and promotes MMP activity. PLoS ONE 2012, 7, e34862. [Google Scholar] [CrossRef]
- Mahendroo, M.S.; Porter, A.; Russell, D.W.; Word, R.A. The parturition defect in steroid 5α-reductase type 1 knockout mice is due to impaired cervical ripening. Mol. Endocrinol. 1999, 13, 981–992. [Google Scholar] [CrossRef]
- Mahendroo, M.S.; Cala, K.M.; Russell, D.W. 5 Alpha-reduced androgens play a key role in murine parturition. Mol. Endocrinol. 1996, 10, 380–392. [Google Scholar] [CrossRef]
- Word, R.A.; Landrum, C.P.; Timmons, B.C.; Young, S.G.; Mahendroo, M.S. Transgene insertion on mouse chromosome 6 impairs function of the uterine cervix and causes failure of parturition. Biol. Reprod. 2005, 73, 1046–1056. [Google Scholar] [CrossRef][Green Version]
- Song, H.; Lim, H.; Paria, B.C.; Matsumoto, H.; Swift, L.L.; Morrow, J.; Bonventre, J.V.; Dey, S.K. Cytosolic phospholipase A2alpha is crucial [correction of A2alpha deficiency is crucial] for ‘on-time’ embryo implantation that directs subsequent development. Development 2002, 129, 2879–2889. [Google Scholar] [CrossRef]
- Uozumi, N.; Kume, K.; Nagase, T.; Nakatani, N.; Ishii, S.; Tashiro, F.; Komagata, Y.; Maki, K.; Ikuta, K.; Ouchi, Y.; et al. Role of cytosolic phospholipase A2 in allergic response and parturition. Nature 1997, 390, 618–622. [Google Scholar] [CrossRef]
- Brown, N.; Morrow, J.D.; Slaughter, J.C.; Paria, B.C.; Reese, J. Restoration of on-time embryo implantation corrects the timing of parturition in cytosolic phospholipase A2 group IVA deficient mice. Biol. Reprod. 2009, 81, 1131–1138. [Google Scholar] [CrossRef]
- Ye, X.; Hama, K.; Contos, J.J.; Anliker, B.; Inoue, A.; Skinner, M.K.; Suzuki, H.; Amano, T.; Kennedy, G.; Arai, H.; et al. LPA3-mediated lysophosphatidic acid signalling in embryo implantation and spacing. Nature 2005, 435, 104–108. [Google Scholar] [CrossRef]
- Brown, J.L.; Sones, J.L.; Angulo, C.N.; Abbott, K.; Miller, A.D.; Boehm, U.; Roberson, M.S. Conditional loss of ERK1 and ERK2 results in abnormal placentation and delayed parturition in the mouse. Sci. Rep. 2019, 9, 9641. [Google Scholar] [CrossRef]
- Mounzih, K.; Qiu, J.; Ewart-Toland, A.; Chehab, F.F. Leptin is not necessary for gestation and parturition but regulates maternal nutrition via a leptin resistance state. Endocrinology 1998, 139, 5259–5262. [Google Scholar] [CrossRef][Green Version]
- Davis, V.L.; Couse, J.F.; Goulding, E.H.; Power, S.G.; Eddy, E.M.; Korach, K.S. Aberrant reproductive phenotypes evident in transgenic mice expressing the wild-type mouse estrogen receptor. Endocrinology 1994, 135, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Vassalli, A.; Matzuk, M.M.; Gardner, H.A.; Lee, K.F.; Jaenisch, R. Activin/inhibin beta B subunit gene disruption leads to defects in eyelid development and female reproduction. Genes Dev. 1994, 8, 414–427. [Google Scholar] [CrossRef] [PubMed]
- Herington, J.L.; O’Brien, C.; Robuck, M.F.; Lei, W.; Brown, N.; Slaughter, J.C.; Paria, B.C.; Mahadevan-Jansen, A.; Reese, J. Prostaglandin-endoperoxide synthase 1 mediates the timing of parturition in mice despite unhindered uterine contractility. Endocrinology 2018, 159, 490–505. [Google Scholar] [CrossRef] [PubMed]
- Loftin, C.D.; Trivedi, D.B.; Langenbach, R. Cyclooxygenase-1-selective inhibition prolongs gestation in mice without adverse effects on the ductus arteriosus. J. Clin. Investig. 2002, 110, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Reinebrant, H.E.; Pileggi-Castro, C.; Romero, C.L.; Santos, R.A.D.; Kumar, S.; Souza, J.P.; Flenady, V. Cyclo-oxygenase (COX) inhibitors for treating preterm labour. Cochrane Database Syst. Rev. 2015, 2015, CD001992. [Google Scholar] [CrossRef] [PubMed]
- Bole-Feysot, C.; Goffin, V.; Edery, M.; Binart, N.; Kelly, P.A. Prolactin (PRL) and its receptor: Actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr. Rev. 1998, 19, 225–268. [Google Scholar] [CrossRef] [PubMed]
- Mulac-Jericevic, B.; Lydon, J.P.; DeMayo, F.J.; Conneely, O.M. Defective mammary gland morphogenesis in mice lacking the progesterone receptor B isoform. Proc. Natl Acad. Sci. USA 2003, 100, 9744–9749. [Google Scholar] [CrossRef] [PubMed]
- Mulac-Jericevic, B.; Mullinax, R.A.; DeMayo, F.J.; Lydon, J.P.; Conneely, O.M. Subgroup of reproductive functions of progesterone mediated by progesterone receptor-B isoform. Science 2000, 289, 1751–1754. [Google Scholar] [CrossRef]
- Nadeem, L.; Shynlova, O.; Matysiak-Zablocki, E.; Mesiano, S.; Dong, X.; Lye, S. Molecular evidence of functional progesterone withdrawal in human myometrium. Nat. Commun. 2016, 7, 11565. [Google Scholar] [CrossRef]
- Velarde, M.C.; Geng, Y.; Eason, R.R.; Simmen, F.A.; Simmen, R.C. Null mutation of Kruppel-like factor9/basic transcription element binding protein-1 alters peri-implantation uterine development in mice. Biol. Reprod. 2005, 73, 472–481. [Google Scholar] [CrossRef][Green Version]
- Pabona, J.M.; Zhang, D.; Ginsburg, D.S.; Simmen, F.A.; Simmen, R.C. Prolonged pregnancy in women is associated with attenuated myometrial expression of progesterone receptor co-regulator Kruppel-like factor 9. J. Clin. Endocrinol. Metab. 2015, 100, 166–174. [Google Scholar] [CrossRef][Green Version]
- Vannuccini, S.; Bocchi, C.; Severi, F.M.; Challis, J.R.; Petraglia, F. Endocrinology of human parturition. Ann. Endocrinol. 2016, 77, 105–113. [Google Scholar] [CrossRef]
- Huang, C.C.; Shih, M.C.; Hsu, N.C.; Chien, Y.; Chung, B.C. Fetal glucocorticoid synthesis is required for development of fetal adrenal medulla and hypothalamus feedback suppression. Endocrinology 2012, 153, 4749–4756. [Google Scholar] [CrossRef]
- Toyoshima, K.; Narahara, H.; Furukawa, M.; Frenkel, R.A.; Johnston, J.M. Platelet-activating factor. Role in fetal lung development and relationship to normal and premature labor. Clin. Perinatol. 1995, 22, 263–280. [Google Scholar] [CrossRef]
- Okada, Y.; Ueshin, Y.; Isotani, A.; Saito-Fujita, T.; Nakashima, H.; Kimura, K.; Mizoguchi, A.; Oh-Hora, M.; Mori, Y.; Ogata, M.; et al. Complementation of placental defects and embryonic lethality by trophoblast-specific lentiviral gene transfer. Nat. Biotechnol. 2007, 25, 233–237. [Google Scholar] [CrossRef]
- Miki, H.; Wakisaka, N.; Inoue, K.; Ogonuki, N.; Mori, M.; Kim, J.M.; Ohta, A.; Ogura, A. Embryonic rather than extraembryonic tissues have more impact on the development of placental hyperplasia in cloned mice. Placenta 2009, 30, 543–546. [Google Scholar] [CrossRef]
- Kanai-Azuma, M.; Kanai, Y.; Okamoto, M.; Hayashi, Y.; Yonekawa, H.; Yazaki, K. Nrk: A murine X-linked NIK (Nck-interacting kinase)-related kinase gene expressed in skeletal muscle. Mech. Dev. 1999, 89, 155–159. [Google Scholar] [CrossRef]
- Doyle, S.L.; O’Neill, L.A. Toll-like receptors: From the discovery of NFκB to new insights into transcriptional regulations in innate immunity. Biochem. Pharmacol. 2006, 72, 1102–1113. [Google Scholar] [CrossRef]
- Piccinini, A.M.; Midwood, K.S. DAMPening inflammation by modulating TLR signalling. Mediat. Inflam. 2010, 2010, 672395. [Google Scholar] [CrossRef]
- Rada, C.C.; Pierce, S.L.; Grotegut, C.A.; England, S.K. Intrauterine telemetry to measure mouse contractile pressure in vivo. J. Vis. Exp. 2015, 98, 52541. [Google Scholar] [CrossRef]
- Robuck, M.F.; O’Brien, C.M.; Knapp, K.M.; Shay, S.D.; West, J.D.; Newton, J.M.; Slaughter, J.C.; Paria, B.C.; Reese, J.; Herington, J.L. Monitoring uterine contractility in mice using a transcervical intrauterine pressure catheter. Reproduction 2018, 155, 447–456. [Google Scholar] [CrossRef]
- Mazzone, J.N.; Kaiser, R.A.; Buxton, I.L. Calcium-activated potassium channel expression in human myometrium: Effect of pregnancy. Proc. West. Pharmacol. Soc. 2002, 45, 184–186. [Google Scholar]
- Woclawek-Potocka, I.; Komiyama, J.; Saulnier-Blache, J.S.; Brzezicka, E.; Bah, M.M.; Okuda, K.; Skarzynski, D.J. Lysophosphatic acid modulates prostaglandin secretion in the bovine uterus. Reproduction 2009, 137, 95–105. [Google Scholar] [CrossRef]
- Tokumura, A.; Kanaya, Y.; Miyake, M.; Yamano, S.; Irahara, M.; Fukuzawa, K. Increased production of bioactive lysophosphatidic acid by serum lysophospholipase D in human pregnancy. Biol. Reprod. 2002, 67, 1386–1392. [Google Scholar] [CrossRef]
- Saba-El-Leil, M.K.; Vella, F.D.; Vernay, B.; Voisin, L.; Chen, L.; Labrecque, N.; Ang, S.L.; Meloche, S. An essential function of the mitogen-activated protein kinase Erk2 in mouse trophoblast development. EMBO Rep. 2003, 4, 964–968. [Google Scholar] [CrossRef]
- Barash, I.A.; Cheung, C.C.; Weigle, D.S.; Ren, H.; Kabigting, E.B.; Kuijper, J.L.; Clifton, D.K.; Steiner, R.A. Leptin is a metabolic signal to the reproductive system. Endocrinology 1996, 137, 3144–3147. [Google Scholar] [CrossRef]
- Biggers, J.D.; Curnow, R.N.; Finn, C.A.; McLaren, A. Regulation of the gestation period in mice. J. Reprod. Fertil. 1963, 6, 125–138. [Google Scholar] [CrossRef]
- Murray, S.A.; Morgan, J.L.; Kane, C.; Sharma, Y.; Heffner, C.S.; Lake, J.; Donahue, L.R. Mouse gestation length is genetically determined. PLoS ONE 2010, 5, e12418. [Google Scholar] [CrossRef]
- Reaume, A.G.; de Sousa, P.A.; Kulkarni, S.; Langille, B.L.; Zhu, D.; Davies, T.C.; Juneja, S.C.; Kidder, G.M.; Rossant, J. Cardiac malformation in neonatal mice lacking connexin43. Science 1995, 267, 1831–1834. [Google Scholar] [CrossRef]
- Romero, R.; Nicolaides, K.; Conde-Agudelo, A.; Tabor, A.; O’Brien, J.M.; Cetingoz, E.; da Fonseca, E.; Creasy, G.W.; Klein, K.; Rode, L.; et al. Vaginal progesterone in women with an asymptomatic sonographic short cervix in the midtrimester decreases preterm delivery and neonatal morbidity: A systematic review and metaanalysis of individual patient data. Am. J. Obstet. Gynecol. 2012, 206, 124.e1–124.e19. [Google Scholar] [CrossRef]
- Ben-Jonathan, N.; Mershon, J.L.; Allen, D.L.; Steinmetz, R.W. Extrapituitary prolactin: Distribution, regulation, functions, and clinical aspects. Endocr. Rev. 1996, 17, 639–669. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1. Failure of luteolysis Prostaglandin F Receptor knockout mouse: Fp KO [5,6] Cyclooxygenase-1-deficient mouse: Cox1 KO [7] 20α hydroxysteroid dehydrogenase-deficient mice: 20α Hsd KO [8] Gαqf/f;Gα11-/-;Cre+ mice: Gq/11 cKO [9] Mastermind-like domain-containing 1 knockout mouse: Mamld1 KO [10] |
| 2. Abnormalities in progesterone metabolism and receptors Kruppel-like factor 9 knockout mouse: Klf 9 KO [11] Cytochrome p450 family 11a1-overexpression mouse: Cyp11a1 Tg [12] |
| 3. Fetal factors Surfactant protein A and D double-deficient mice: Sp A/D dKO [13] Steroid receptor coactivator 1 and 2 double-deficient mice: Src 1/2 dKO [14] |
| 4. Placental factors Solute carrier organic anion transporter family member 2A1 knockout mouse: Slco2a1 KO [4] Sushi-ichi retrotransposon homolog 7/Leucine zipper, downregulated cancer 1 knockout mouse: Sirh7/Ldoc1 KO [15] Nik-related kinase knockout mouse: Nrk KO [16] |
| 5. Autoimmune disorder Toll-like receptor 2-deficient mice: Tlr 2 KO [13] Toll-like receptor 4-deficient mice: Tlr 4 KO [17] Interleukin 6 null mutant mice: Il6 KO [18] |
| 6. Uterine contractile dysfunction Connexin 43fl/fl:SM-CreERT2 mice: Sm-CreERT2 KO [19] Kcnn3tm1Jpad/Kcnn3tm1jpad: Sk3T/T mice [20] Anthrax toxin receptor 2 knockout mice: Antxr 2 KO [21] |
| 7. Poor cervical ripening 5α reductase type 1 knockout mice: 5αR1 KO [22,23] TgN(hApoB)1102SY line: Tg/Tg mouse [24] Anthrax toxin receptor 2 knockout mice: Antxr 2 KO [21] |
| 8. Delayed implantation Cytosolic phosphatase A2 mutant mouse: Pla2g4a KO [25,26,27] Lysophosphatidic acid receptor 3-deficient mouse: LPA3 KO [28] |
| 9. Unknown cause Erk 1 and Erk 2 conditional knockout mouse: Erk1/2 cKO [29] Leptin KO mouse: Leptin KO [30] Mt mER mouse [31] Activin/inhibin βB subunit-deficient mouse: Activin/inhibin βB subunit dKO [32] |
| Before pregnancy | Transplanting KO embryos into wild-type mice Tetraploid chimera Administration of LV |
| 5.5–6.5 dpc | Delay implantation |
| 17.5–19.5 dpc | Measurement of P4 Evaluation of cervical ripening (biometric test, pathological test) Evaluation of uterine contraction (in vivo and vitro) |
| 19.5 dpc- | HE staining and/or tunnel staining of corpus luteum Administration of RU486 Ovariectomy |
| After labor | Investigation of the maternal and fetal genotype with delayed labor Screening of placenta and fetus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yomogita, H.; Miyasaka, N.; Kanai-Azuma, M. A Review of Delayed Delivery Models and the Analysis Method in Mice. J. Dev. Biol. 2022, 10, 20. https://doi.org/10.3390/jdb10020020
Yomogita H, Miyasaka N, Kanai-Azuma M. A Review of Delayed Delivery Models and the Analysis Method in Mice. Journal of Developmental Biology. 2022; 10(2):20. https://doi.org/10.3390/jdb10020020
Chicago/Turabian StyleYomogita, Hiroshi, Naoyuki Miyasaka, and Masami Kanai-Azuma. 2022. "A Review of Delayed Delivery Models and the Analysis Method in Mice" Journal of Developmental Biology 10, no. 2: 20. https://doi.org/10.3390/jdb10020020
APA StyleYomogita, H., Miyasaka, N., & Kanai-Azuma, M. (2022). A Review of Delayed Delivery Models and the Analysis Method in Mice. Journal of Developmental Biology, 10(2), 20. https://doi.org/10.3390/jdb10020020