
Lizard Blastema Organoid Model Recapitulates Regenerated Tail Chondrogenesis

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lizard Maintenance and Handling
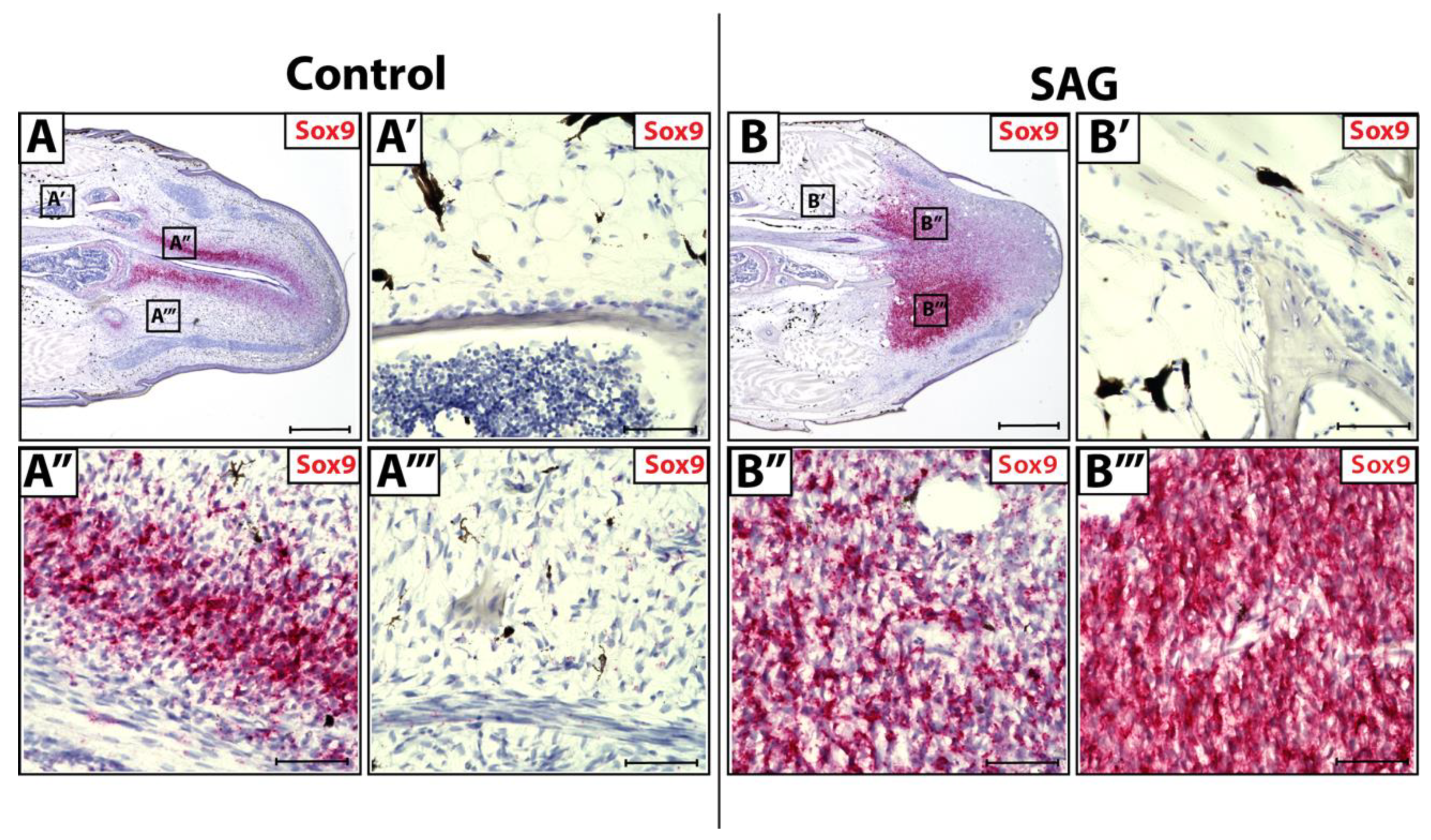
2.2. SAG Injections
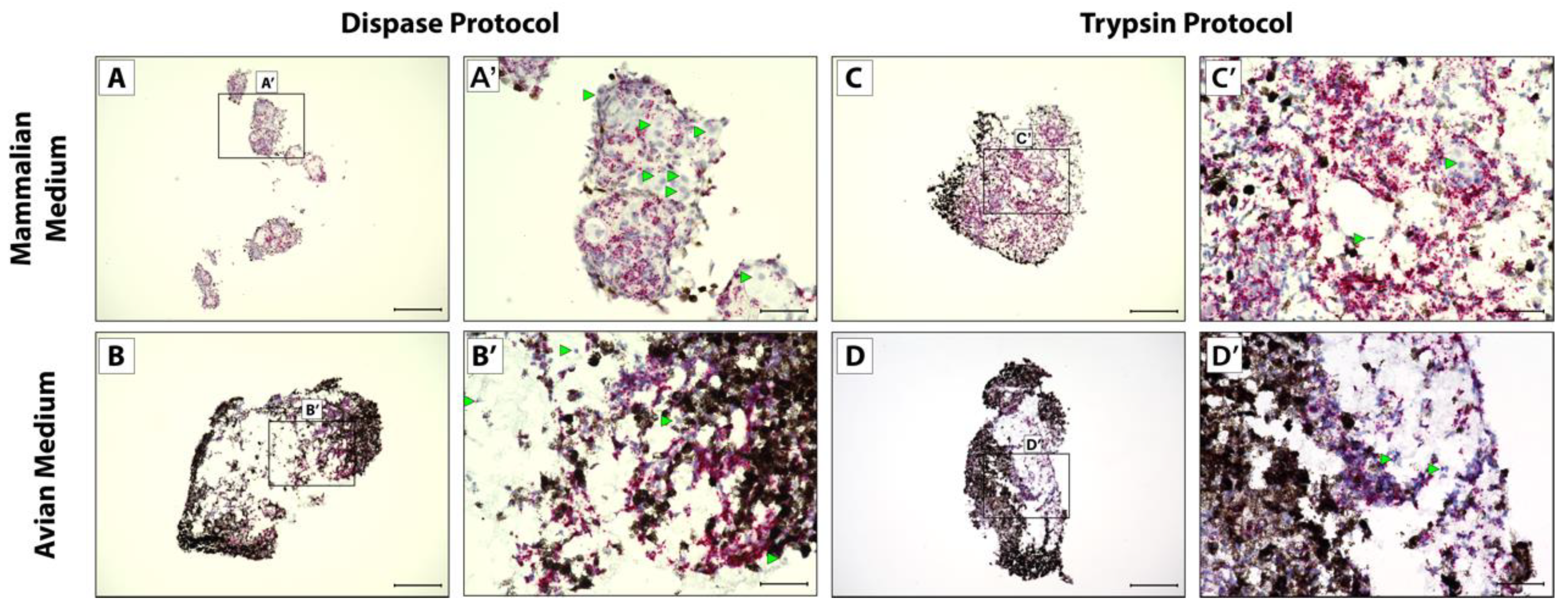
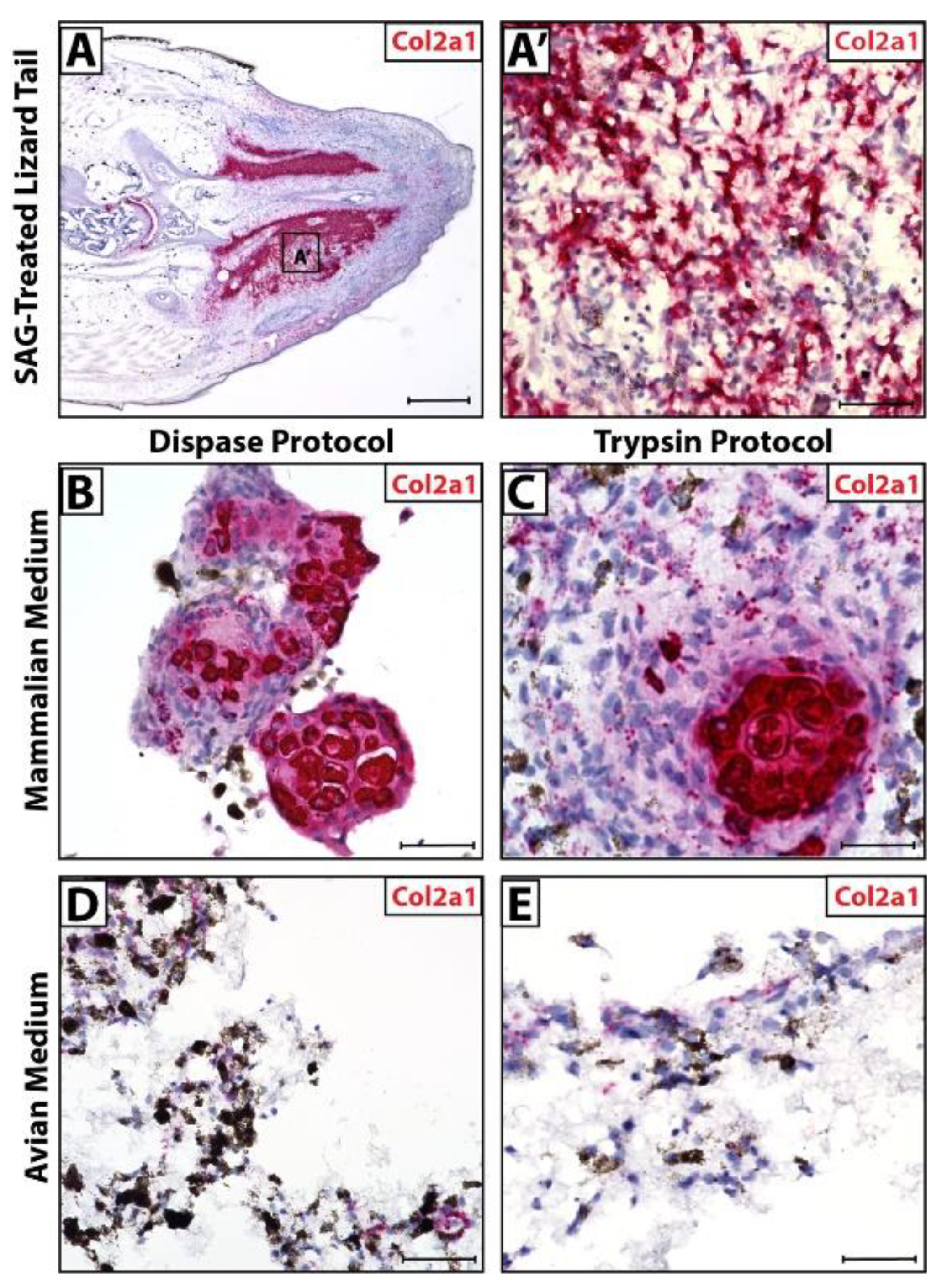
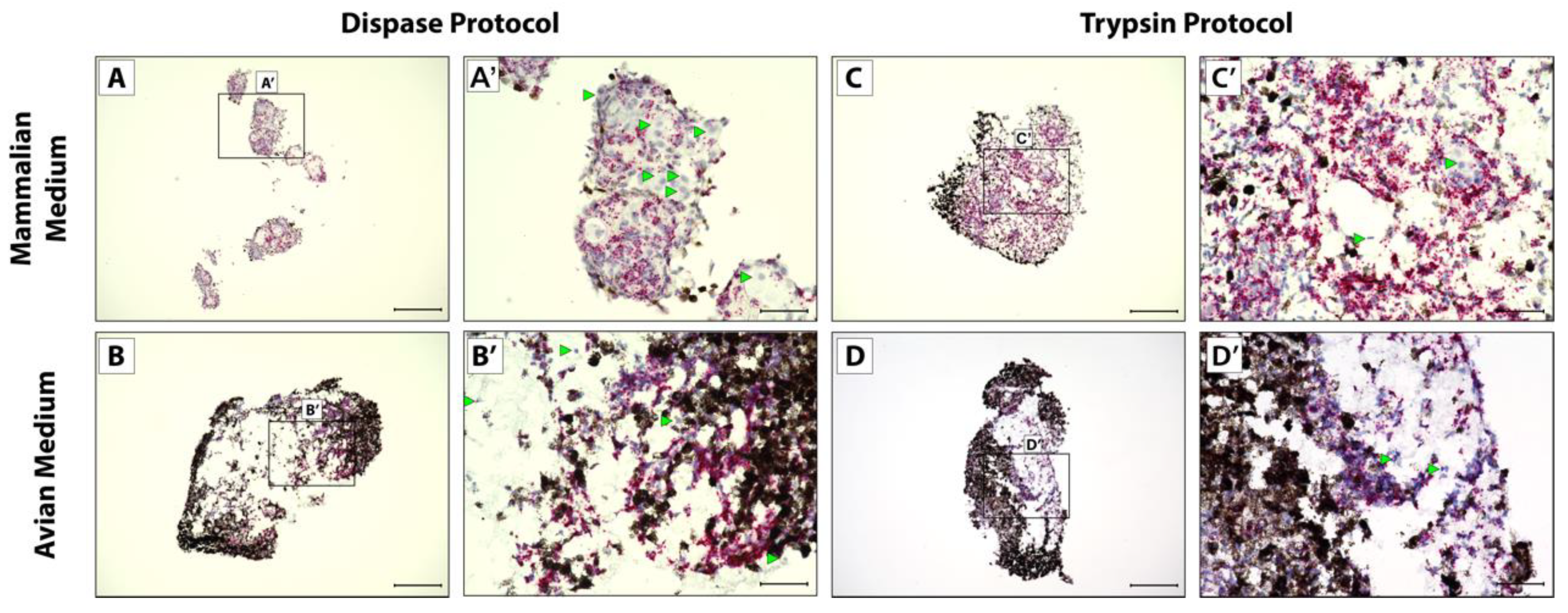
2.3. Lizard Tail Amputations and Dissociation Protocols
2.4. Cell Culture
2.5. Histology
2.6. In Situ Hybridization (ISH)
3. Results
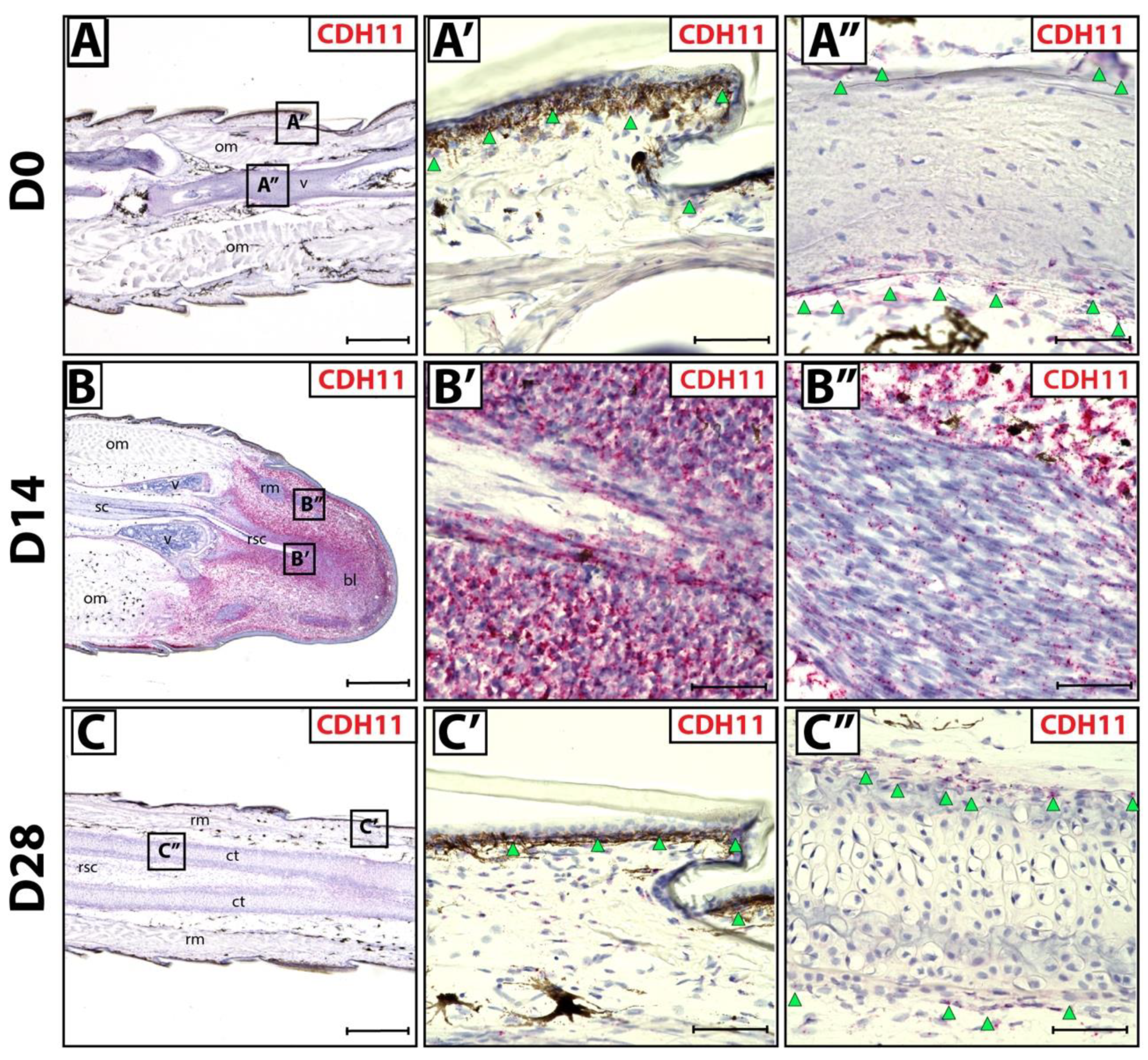
3.1. Fibroblastic Connective Tissue Cells Express CDH11 in Lizard Blastema
3.2. CDH11+ Blastema Cell Culture System Optimization
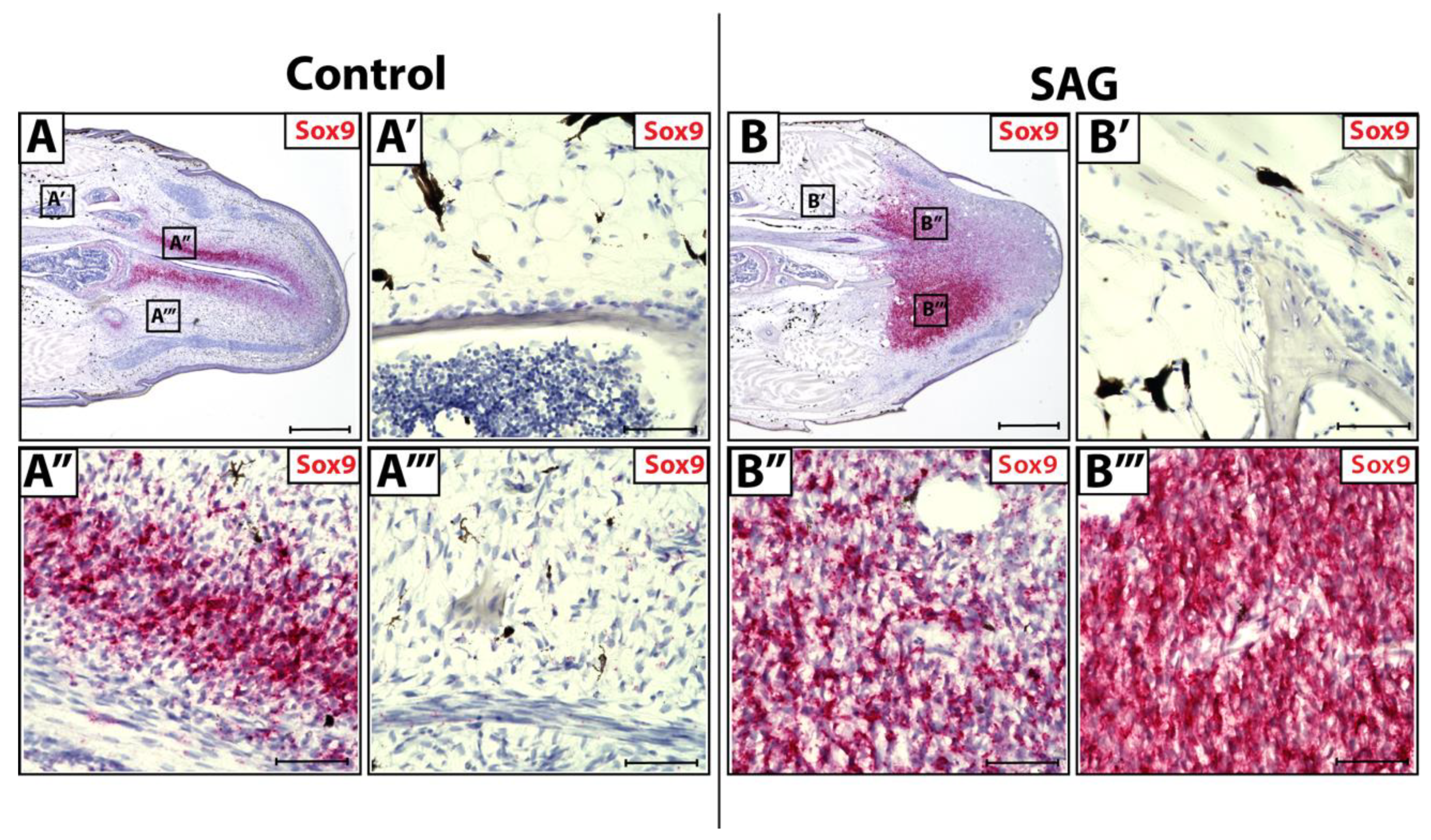
3.3. Sox9 Expression in Wild-Type and SAG-Treated Blastema
3.4. Micromass Blastema Organoid Cultures Mimic Regenerating Tail Cartilage Formation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lozito, T.P.; Tuan, R.S. Lizard Tail Regeneration as an Instructive Model of Enhanced Healing Capabilities in an Adult Amniote. Connect. Tissue Res. 2017, 58, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Londono, R.; Sun, A.X.; Tuan, R.S.; Lozito, T.P. Tissue repair and epimorphic regeneration: An overview. Curr. Pathobiol. Rep. 2018, 6, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Stocum, D.L. Mechanisms of Urodele Limb Regeneration. Regeneration 2017, 4, 159–200. [Google Scholar] [CrossRef] [PubMed]
- Gross, M. Regeneration Lessons from the Axolotl. Curr. Biol. 2018, 28, R187–R189. [Google Scholar] [CrossRef]
- Fisher, R.E.; Geiger, L.A.; Stroik, L.K.; Hutchins, E.D.; George, R.M.; Denardo, D.F.; Kusumi, K.; Rawls, J.A.; Wilson-Rawls, J. A Histological Comparison of the Original and Regenerated Tail in the Green Anole, Anolis Carolinensis. Anat. Rec. 2012, 295, 1609–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alibardi, L. Regeneration in Reptiles and Its Position Among Vertebrates. In Morphological and Cellular Aspects of Tail and Limb Regeneration in Lizards: A Model System With Implications for Tissue Regeneration in Mammals; Alibardi, L., Ed.; Advances in Anatomy, Embryology and Cell Biology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1–49. ISBN 978-3-642-03733-7. [Google Scholar]
- Lozito, T.P.; Tuan, R.S. Lizard Tail Skeletal Regeneration Combines Aspects of Fracture Healing and Blastema-Based Regeneration. Development 2016, 143, 2946–2957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londono, R.; Tighe, S.; Milnes, B.; DeMoya, C.; Quijano, L.M.; Hudnall, M.L.; Nguyen, J.; Tran, E.; Badylak, S.; Lozito, T.P. Single Cell Sequencing Analysis of Lizard Phagocytic Cell Populations and Their Role in Tail Regeneration. J. Immunol. Regen. Med. 2020, 8, 100029. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Histochemical, Biochemical and Cell Biological Aspects of Tail Regeneration in Lizard, an Amniote Model for Studies on Tissue Regeneration. Prog. Histochem. Cytochem. 2014, 48, 143–244. [Google Scholar] [CrossRef] [PubMed]
- Lozito, T.P.; Tuan, R.S. Lizard Tail Regeneration: Regulation of Two Distinct Cartilage Regions by Indian Hedgehog. Dev. Biol. 2015, 399, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Londono, R.; Wenzhong, W.; Wang, B.; Tuan, R.S.; Lozito, T.P. Cartilage and Muscle Cell Fate and Origins during Lizard Tail Regeneration. Front. Bioeng. Biotechnol. 2017, 5, 70. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, V.; Dvir-Ginzberg, M. SOX9 and the Many Facets of Its Regulation in the Chondrocyte Lineage. Connect. Tissue Res. 2017, 58, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Stimulation of Regenerative Blastema Formation in Lizards as a Model to Analyze Limb Regeneration in Amniotes. Histol. Histopathol. 2019, 34, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L.; Meyer-Rochow, V.B. Comparative Fine Structure of the Axial Skeleton inside the Regenerated Tail of Some Lizard Species and the Tuatara (Sphenodon Punctatus). Gegenbaurs Morphol. Jahrb. 1989, 135, 705–716. [Google Scholar] [PubMed]
- Pritchard, J.J.; Ruzicka, A.J. Comparison of Fracture Repair in the Frog, Lizard and Rat. J. Anat. 1950, 84, 236–261. [Google Scholar] [PubMed]
- Gerber, T.; Murawala, P.; Knapp, D.; Masselink, W.; Schuez, M.; Hermann, S.; Gac-Santel, M.; Nowoshilow, S.; Kageyama, J.; Khattak, S.; et al. Single-Cell Analysis Uncovers Convergence of Cell Identities during Axolotl Limb Regeneration. Science 2018, 362, eaaq0681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, N.D.; Dunlap, G.S.; Johnson, K.; Mariano, R.; Oshiro, R.; Wong, A.Y.; Bryant, D.M.; Miller, B.M.; Ratner, A.; Chen, A.; et al. Transcriptomic Landscape of the Blastema Niche in Regenerating Adult Axolotl Limbs at Single-Cell Resolution. Nat. Commun. 2018, 9, 5153. [Google Scholar] [CrossRef] [Green Version]
- Ahrens, P.B.; Solursh, M.; Reiter, R.S. Stage-Related Capacity for Limb Chondrogenesis in Cell Culture. Dev. Biol. 1977, 60, 69–82. [Google Scholar] [CrossRef]
- Daniels, K.; Reiter, R.; Solursh, M. Chapter 12 Micromass Cultures of Limb and Other Mesenchyme. In Methods in Cell Biology; Bronner-Fraser, M., Ed.; Methods in Avian Embryology; Academic Press: Cambridge, MA, USA, 1996; Volume 51, pp. 237–247. [Google Scholar]
- Mello, M.A.; Tuan, R.S. High Density Micromass Cultures of Embryonic Limb Bud Mesenchymal Cells: An in Vitro Model of Endochondral Skeletal Development. In Vitro Cell. Dev. Biol.-Anim. 1999, 35, 262–269. [Google Scholar] [CrossRef]
- De Lise, A.M.; Stringa, E.; Woodward, W.A.; Mello, M.A.; Tuan, R.S. Embryonic Limb Mesenchyme Micromass Culture as an In Vitro Model for Chondrogenesis and Cartilage Maturation. In Developmental Biology Protocols; Tuan, R.S., Lo, C.W., Eds.; Methods in Molecular BiologyTM; Humana Press: Totowa, NJ, USA, 2000; pp. 359–375. ISBN 978-1-59259-066-7. [Google Scholar]
- Handschel, J.G.K.; Depprich, R.A.; Kübler, N.R.; Wiesmann, H.-P.; Ommerborn, M.; Meyer, U. Prospects of Micromass Culture Technology in Tissue Engineering. Head Face Med. 2007, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Koo, B.-K.; Knoblich, J.A. Human Organoids: Model Systems for Human Biology and Medicine. Nat. Rev. Mol. Cell Biol. 2020, 21, 571–584. [Google Scholar] [CrossRef]
- Farmer, D.T.; Mlcochova, H.; Zhou, Y.; Koelling, N.; Wang, G.; Ashley, N.; Bugacov, H.; Chen, H.-J.; Parvez, R.; Tseng, K.-C.; et al. The Developing Mouse Coronal Suture at Single-Cell Resolution. Nat. Commun. 2021, 12, 4797. [Google Scholar] [CrossRef] [PubMed]
- Tuli, R.; Tuli, S.; Nandi, S.; Wang, M.L.; Alexander, P.G.; Haleem-Smith, H.; Hozack, W.J.; Manner, P.A.; Danielson, K.G.; Tuan, R.S. Characterization of Multipotential Mesenchymal Progenitor Cells Derived from Human Trabecular Bone. Stem Cells 2003, 21, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.-C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.-T.; Ma, X.-J.; Luo, Y. RNAscope. J. Mol. Diagn. 2012, 14, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Alimperti, S.; Andreadis, S.T. CDH2 and CDH11 Act as Regulators of Stem Cell Fate Decisions. Stem Cell Res. 2015, 14, 270–282. [Google Scholar] [CrossRef] [Green Version]
- Sun, A.X.; Londono, R.; Hudnall, M.L.; Tuan, R.S.; Lozito, T.P. Differences in Neural Stem Cell Identity and Differentiation Capacity Drive Divergent Regenerative Outcomes in Lizards and Salamanders. Proc. Natl. Acad. Sci. USA 2018, 115, E8256–E8265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozito, T.P.; Londono, R.; Sun, A.X.; Hudnall, M.L. Introducing Dorsoventral Patterning in Adult Regenerating Lizard Tails with Gene-Edited Embryonic Neural Stem Cells. Nat. Commun. 2021, 12, 6010. [Google Scholar] [CrossRef] [PubMed]
- Currie, J.D.; Kawaguchi, A.; Traspas, R.M.; Schuez, M.; Chara, O.; Tanaka, E.M. Live Imaging of Axolotl Digit Regeneration Reveals Spatiotemporal Choreography of Diverse Connective Tissue Progenitor Pools. Dev. Cell 2016, 39, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Rasys, A.M.; Park, S.; Ball, R.E.; Alcala, A.J.; Lauderdale, J.D.; Menke, D.B. CRISPR-Cas9 Gene Editing in Lizards through Microinjection of Unfertilized Oocytes. Cell Rep. 2019, 28, 2288–2292.e3. [Google Scholar] [CrossRef] [Green Version]
- Palade, J.; Djordjevic, D.; Hutchins, E.D.; George, R.M.; Cornelius, J.A.; Rawls, A.; Ho, J.W.K.; Kusumi, K.; Wilson-Rawls, J. Identification of Satellite Cells from Anole Lizard Skeletal Muscle and Demonstration of Expanded Musculoskeletal Potential. Dev. Biol. 2018, 433, 344–356. [Google Scholar] [CrossRef]
- Zuk, P. Adipose-Derived Stem Cells in Tissue Regeneration: A Review. ISRN Stem Cells 2013, 2013, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Tickle, C.; Towers, M. Sonic Hedgehog Signaling in Limb Development. Front. Cell Dev. Biol. 2017, 5, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, M.B.; Fischer, M.; Zellner, J.; Berner, A.; Dienstknecht, T.; Kujat, R.; Prantl, L.; Nerlich, M.; Tuan, R.S.; Angele, P. Effect of Parathyroid Hormone-Related Protein in an in Vitro Hypertrophy Model for Mesenchymal Stem Cell Chondrogenesis. Int. Orthop. 2013, 37, 945–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vonk, A.C.; Hasel-Kolossa, S.C.; Lopez, G.A.; Hudnall, M.L.; Gamble, D.J.; Lozito, T.P. Lizard Blastema Organoid Model Recapitulates Regenerated Tail Chondrogenesis. J. Dev. Biol. 2022, 10, 12. https://doi.org/10.3390/jdb10010012
Vonk AC, Hasel-Kolossa SC, Lopez GA, Hudnall ML, Gamble DJ, Lozito TP. Lizard Blastema Organoid Model Recapitulates Regenerated Tail Chondrogenesis. Journal of Developmental Biology. 2022; 10(1):12. https://doi.org/10.3390/jdb10010012
Chicago/Turabian StyleVonk, Ariel C., Sarah C. Hasel-Kolossa, Gabriela A. Lopez, Megan L. Hudnall, Darian J. Gamble, and Thomas P. Lozito. 2022. "Lizard Blastema Organoid Model Recapitulates Regenerated Tail Chondrogenesis" Journal of Developmental Biology 10, no. 1: 12. https://doi.org/10.3390/jdb10010012
APA StyleVonk, A. C., Hasel-Kolossa, S. C., Lopez, G. A., Hudnall, M. L., Gamble, D. J., & Lozito, T. P. (2022). Lizard Blastema Organoid Model Recapitulates Regenerated Tail Chondrogenesis. Journal of Developmental Biology, 10(1), 12. https://doi.org/10.3390/jdb10010012