
Micromanagement of Drosophila Post-Embryonic Development by Hox Genes


{kind=link}
Abstract
1. Introduction
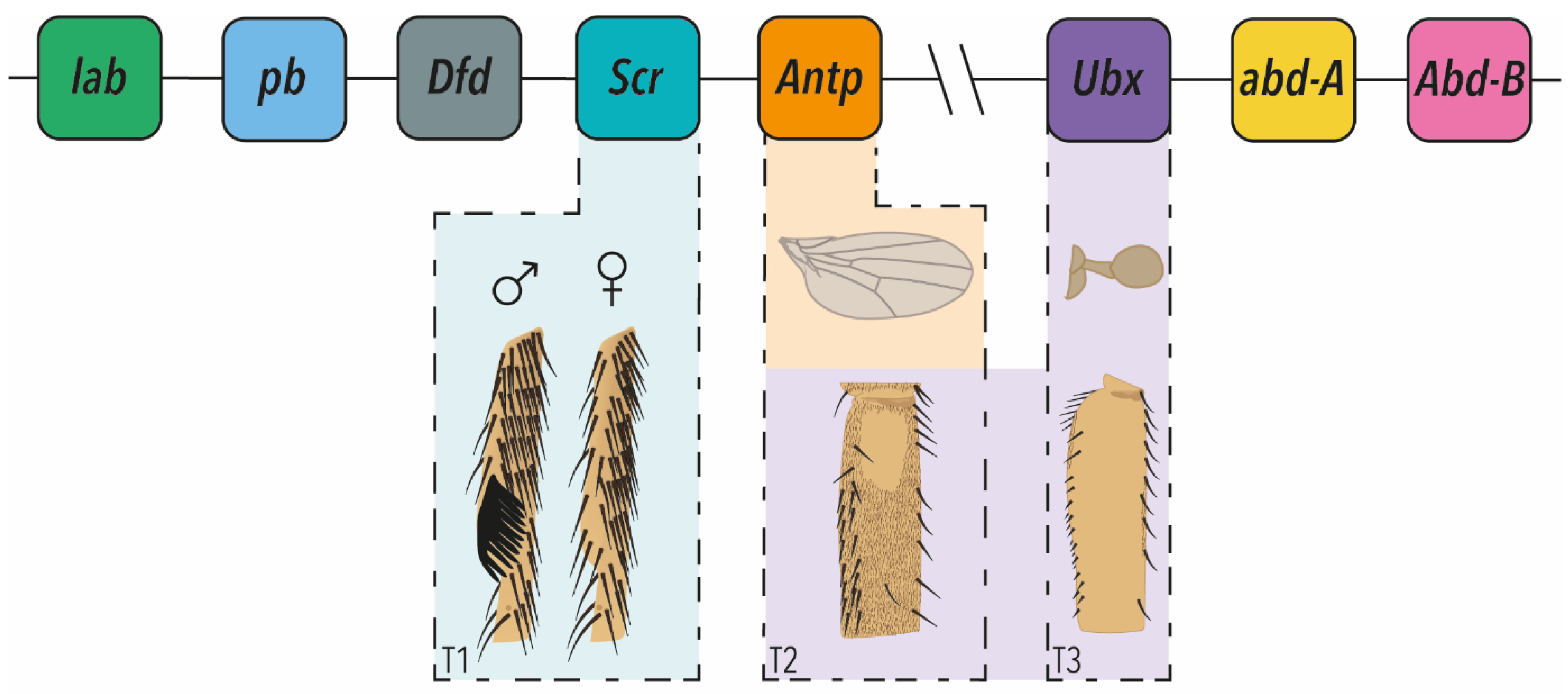
2. Hox Gene Regulation of Fine-Scale Phenotypes in Drosophila
2.1. Wings (T2) versus Halteres (T3)
2.2. Leg Morphology
2.3. Micromanagement of Other Aspects of Post-Embryonic Development
3. Hox Target Genes in Post-Embryonic Development
4. Future Work on Roles of Hox Genes in Post-Embryonic Gene Regulatory Networks
Funding
Acknowledgments
Conflicts of Interest
References
- Hughes, C.L.; Kaufman, T.C. Hox genes and the evolution of the arthropod body plan. Evol. Dev. 2002, 4, 459–499. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.B. A gene complex controlling segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Gellon, G.; McGinnis, W. Shaping animal body plans in development and evolution by modulation of Hox expression patterns. BioEssays 1998, 20, 116–125. [Google Scholar] [CrossRef]
- Gehring, W.J. Homeotic genes, the homeo box, and the genetic control of development. Cold Spring Harb. Symp. Quant. Biol. 1985, 50, 243–251. [Google Scholar] [CrossRef]
- Holland, P.W. Evolution of homeobox genes. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 31–45. [Google Scholar] [CrossRef]
- Akam, M. Hox and HOM: Homologous gene clusters in insects and vertebrates. Cell 1989, 57, 347–349. [Google Scholar] [CrossRef]
- Gaunt, S.J. Hox cluster genes and collinearities throughout the tree of animal life. Int. J. Dev. Biol. 2018, 62, 673–683. [Google Scholar] [CrossRef]
- Duboule, D. The rise and fall of Hox gene clusters. Development 2007, 134, 2549–2560. [Google Scholar] [CrossRef]
- Akam, M.; Averof, M.; Castelli-Gair, J.; Dawes, R.; Falciani, F.; Ferrier, D. The evolving role of Hox genes in arthropods. Dev. Suppl. 1994, 209–215. [Google Scholar] [CrossRef]
- Krumlauf, R. Hox genes, clusters and collinearity. Int. J. Dev. Biol. 2018, 62, 659–663. [Google Scholar] [CrossRef]
- Akam, M. Hox genes: From master genes to micromanagers. Curr. Biol. 1998, 8, R676–R678. [Google Scholar] [CrossRef]
- Bateson, W. Materials for the Study of Variation Treated with Especial Regard to Discontinuity in the Origin of Species; MacMillan: London, UK, 1894. [Google Scholar]
- Akam, M. Hox genes, homeosis and the evolution of segment identity: No need for hopeless monsters. Int. J. Dev. Biol. 1998, 42, 445–451. [Google Scholar] [PubMed]
- Barmina, O.; Kopp, A. Sex-specific expression of a HOX gene associated with rapid morphological evolution. Dev. Biol. 2007, 311, 277–286. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jeong, S.; Rokas, A.; Carroll, S.B. Regulation of body pigmentation by the Abdominal-B Hox protein and its gain and loss in Drosophila evolution. Cell 2006, 125, 1387–1399. [Google Scholar] [CrossRef] [PubMed]
- Stern, D.L. A role of Ultrabithorax in morphological differences between Drosophila species. Nature 1998, 396, 463–466. [Google Scholar] [CrossRef]
- Roch, F.; Akam, M. Ultrabithorax and the control of cell morphology in Drosophila halteres. Development 2000, 127, 97–107. [Google Scholar] [CrossRef]
- Carroll, S.B.; Weatherbee, S.D.; Langeland, J.A. Homeotic genes and the regulation and evolution of insect wing number. Nature 1995, 375, 58–61. [Google Scholar] [CrossRef]
- Weatherbee, S.D.; Halder, G.; Kim, J.; Hudson, A.; Carroll, S. Ultrabithorax regulates genes at several levels of the wing-patterning hierarchy to shape the development of the Drosophila haltere. Genes Dev. 1998, 12, 1474–1482. [Google Scholar] [CrossRef]
- Paul, R.; Giraud, G.; Domsch, K.; Duffraisse, M.; Marmigère, F.; Khan, S.; Vanderperre, S.; Lohmann, I.; Stoks, R.; Shashidhara, L.S.; et al. Hox dosage contributes to flight appendage morphology in Drosophila. Nat. Commun 2021, 12, 2892. [Google Scholar] [CrossRef]
- Casares, F.; Calleja, M.; Sánchez-Herrero, E. Functional similarity in appendage specification by the Ultrabithorax and abdominal-A Drosophila HOX genes. EMBO J. 1996, 15, 3934–3942. [Google Scholar] [CrossRef]
- De Las Heras, J.M.; García-Cortés, C.; Foronda, D.; Pastor-Pareja, J.C.; Shashidhara, L.S.; Sánchez-Herrero, E. The Drosophila Hox gene Ultrabithorax controls appendage shape by regulating extracellular matrix dynamics. Development 2018, 145, dev161844. [Google Scholar] [CrossRef] [PubMed]
- Diaz-de-la-Loza, M.; Loker, R.; Mann, R.S.; Thompson, B.J. Control of tissue morphogenesis by the HOX gene Ultrabithorax. Development 2020, 147, dev184564. [Google Scholar] [CrossRef] [PubMed]
- Hersh, B.B.; Nelson, C.E.; Stoll, S.J.; Norton, J.E.; Albert, T.J.; Carroll, S.B. The UBX-regulated network in the haltere imaginal disc of D. melanogaster. Dev. Biol. 2007, 302, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Pallavi, S.K.; Kannan, R.; Shashidhara, L.S. Negative regulation of Egfr/Ras pathway by Ultrabithorax during haltere development in Drosophila. Dev. Biol. 2006, 296, 340–352. [Google Scholar] [CrossRef]
- Crickmore, M.A.; Mann, R.S. Hox control of organ size by regulation of morphogen production and mobility. Science 2006, 313, 63–68. [Google Scholar] [CrossRef] [PubMed]
- de Navas, L.F.; Garaulet, D.L.; Sánchez-Herrero, E. The ultrabithorax Hox gene of Drosophila controls haltere size by regulating the Dpp pathway. Development 2006, 133, 4495–4506. [Google Scholar] [CrossRef] [PubMed]
- Crickmore, M.A.; Ranade, V.; Mann, R.S. Regulation of Ubx expression by epigenetic enhancer silencing in response to Ubx levels and genetic variation. PLoS Genet. 2009, 5, e1000633. [Google Scholar] [CrossRef]
- Delker, R.K.; Ranade, V.; Loker, R.; Voutev, R.; Mann, R.S. Low affinity binding sites in an activating CRM mediate negative autoregulation of the Drosophila Hox gene Ultrabithorax. PLoS Genet. 2019, 15, e1008444. [Google Scholar] [CrossRef]
- Little, J.W.; Byrd, C.A.; Brower, D.L. Effect of abx, bx and pbx mutations on expression of homeotic genes in Drosophila larvae. Genetics 1990, 124, 899–908. [Google Scholar] [CrossRef]
- Simon, J.; Peifer, M.; Bender, W.; O’Connor, M. Regulatory elements of the bithorax complex that control expression along the anterior-posterior axis. EMBO J. 1990, 9, 3945–3956. [Google Scholar] [CrossRef]
- Loker, R.; Sanner, J.E.; Mann, R.S. Cell-type-specific Hox regulatory strategies orchestrate tissue identity. Curr. Biol. 2021, 31, 4246–4255.e4244. [Google Scholar] [CrossRef] [PubMed]
- Casares, F.; Mann, R.S. A dual role for homothorax in inhibiting wing blade development and specifying proximal wing identities in Drosophila. Development 2000, 127, 1499–1508. [Google Scholar] [CrossRef]
- Choo, S.W.; White, R.; Russell, S. Genome-wide analysis of the binding of the Hox protein Ultrabithorax and the Hox cofactor Homothorax in Drosophila. PLoS ONE 2011, 6, e14778. [Google Scholar] [CrossRef] [PubMed]
- Galant, R.; Walsh, C.M.; Carroll, S.B. Hox repression of a target gene: Extradenticle-independent, additive action through multiple monomer binding sites. Development 2002, 129, 3115–3126. [Google Scholar] [CrossRef]
- Porcelli, D.; Fischer, B.; Russell, S.; White, R. Chromatin accessibility plays a key role in selective targeting of Hox proteins. Genome Biol. 2019, 20, 115. [Google Scholar] [CrossRef] [PubMed]
- Bulajić, M.; Srivastava, D.; Dasen, J.S.; Wichterle, H.; Mahony, S.; Mazzoni, E.O. Differential abilities to engage inaccessible chromatin diversify vertebrate Hox binding patterns. Development 2020, 147, dev194761. [Google Scholar] [CrossRef] [PubMed]
- Desanlis, I.; Kherdjemil, Y.; Mayran, A.; Bouklouch, Y.; Gentile, C.; Sheth, R.; Zeller, R.; Drouin, J.; Kmita, M. HOX13-dependent chromatin accessibility underlies the transition towards the digit development program. Nat. Commun. 2020, 11, 2491. [Google Scholar] [CrossRef] [PubMed]
- Schubiger, G.; Schubiger, M.; Sustar, A. The three leg imaginal discs of Drosophila: “Vive la difference”. Dev. Biol. 2012, 369, 76–90. [Google Scholar] [CrossRef][Green Version]
- Tokunaga, C. Cell lineage and differentiation on the male foreleg of Drosophila melanogaster. Dev. Biol. 1962, 4, 489–516. [Google Scholar] [CrossRef]
- Hannah-Alava, A. Morphology and chaetotaxy of the legs of Drosophila melanogaster. J. Morphol. 1958, 103, 281–310. [Google Scholar] [CrossRef]
- Eksi, S.E.; Barmina, O.; McCallough, C.L.; Kopp, A.; Orenic, T.V. A Distalless-responsive enhancer of the Hox gene Sex combs reduced is required for segment- and sex-specific sensory organ development in Drosophila. PLoS Genet. 2018, 14, e1007320. [Google Scholar] [CrossRef]
- Shroff, S.; Joshi, M.; Orenic, T.V. Differential Delta expression underlies the diversity of sensory organ patterns among the legs of the Drosophila adult. Mech. Dev. 2007, 124, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Spieth, H.T. Mating behavior within the genus Drosophila (Diptera). Bull. Am. J. Nat. Hist. 1952, 99, 395–474. [Google Scholar]
- Massey, J.H.; Chung, D.; Siwanowicz, I.; Stern, D.L.; Wittkopp, P.J. The yellow gene influences Drosophila male mating success through sex comb melanization. eLife 2019, 8, e49388. [Google Scholar] [CrossRef] [PubMed]
- Rogers, B.T.; Peterson, M.D.; Kaufman, T.C. Evolution of the insect body plan as revealed by the Sex combs reduced expression pattern. Development 1997, 124, 149–157. [Google Scholar] [CrossRef]
- Glicksman, M.A.; Brower, D.L. Expression of the Sex combs reduced protein in Drosophila larvae. Dev. Biol. 1988, 127, 113–118. [Google Scholar] [CrossRef]
- Tanaka, K.; Barmina, O.; Sanders, L.E.; Arbeitman, M.N.; Kopp, A. Evolution of sex-specific traits through changes in HOX-dependent doublesex expression. PLoS Biol. 2011, 9, e1001131. [Google Scholar] [CrossRef]
- Randsholt, N.B.; Santamaria, P. How Drosophila change their combs: The Hox gene Sex combs reduced and sex comb variation among Sophophora species. Evol. Dev. 2008, 10, 121–133. [Google Scholar] [CrossRef]
- Stern, D.L. The Hox gene Ultrabithorax modulates the shape and size of the third leg of Drosophila by influencing diverse mechanisms. Dev. Biol. 2003, 256, 355–366. [Google Scholar] [CrossRef]
- Davis, G.K.; Srinivasan, D.G.; Wittkopp, P.J.; Stern, D.L. The function and regulation of Ultrabithorax in the legs of Drosophila melanogaster. Dev. Biol. 2007, 308, 621–631. [Google Scholar] [CrossRef]
- Arif, S.; Kittelmann, S.; McGregor, A.P. From shavenbaby to the naked valley: Trichome formation as a model for evolutionary developmental biology. Evol. Dev. 2015, 17, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Arif, S.; Murat, S.; Almudi, I.; Nunes, M.D.; Bortolamiol-Becet, D.; McGregor, N.S.; Currie, J.M.; Hughes, H.; Ronshaugen, M.; Sucena, E.; et al. Evolution of mir-92a Underlies Natural Morphological Variation in Drosophila melanogaster. Curr. Biol. 2013, 23, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Buffry, A.D.; Kittelmann, S.; McGregor, A.P. Characterisation of the role and regulation of Ultrabithorax in sculpting fine-scale leg morphology. bioRxiv 2020, biorxiv:152918. [Google Scholar] [CrossRef]
- Kittelmann, S.; Buffry, A.D.; Franke, F.A.; Almudi, I.; Yoth, M.; Sabaris, G.; Couso, J.P.; Nunes, M.D.S.; Frankel, N.; Gomez-Skarmeta, J.L.; et al. Gene regulatory network architecture in different developmental contexts influences the genetic basis of morphological evolution. PLoS Genet. 2018, 14, e1007375. [Google Scholar] [CrossRef]
- Singh, N.P.; Mishra, R.K. Role of abd-A and Abd-B in development of abdominal epithelia breaks posterior prevalence rule. PLoS Genet. 2014, 10, e1004717. [Google Scholar] [CrossRef]
- Pinto, P.B.; Espinosa-Vázquez, J.M.; Rivas, M.L.; Hombría, J.C. JAK/STAT and Hox Dynamic Interactions in an Organogenetic Gene Cascade. PLoS Genet. 2015, 11, e1005412. [Google Scholar] [CrossRef]
- Makki, R.; Cinnamon, E.; Gould, A.P. The development and functions of oenocytes. Annu. Rev. Entomol. 2014, 59, 405–425. [Google Scholar] [CrossRef]
- Li-Kroeger, D.; Witt, L.M.; Grimes, H.L.; Cook, T.A.; Gebelein, B. Hox and senseless antagonism functions as a molecular switch to regulate EGF secretion in the Drosophila PNS. Dev. Cell 2008, 15, 298–308. [Google Scholar] [CrossRef]
- Brodu, V.; Elstob, P.R.; Gould, A.P. abdominal A specifies one cell type in Drosophila by regulating one principal target gene. Development 2002, 129, 2957–2963. [Google Scholar] [CrossRef]
- Li-Kroeger, D.; Cook, T.A.; Gebelein, B. Integration of an abdominal Hox complex with Pax2 yields cell-specific EGF secretion from Drosophila sensory precursor cells. Development 2012, 139, 1611–1619. [Google Scholar] [CrossRef]
- Monier, B.; Astier, M.; Semeriva, M.; Perrin, L. Steroid-dependent modification of Hox function drives myocyte reprogramming in the Drosophila heart. Development 2005, 132, 5283–5293. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.; Sipani, R.; Bakshi, A. Roles of Drosophila Hox Genes in the Assembly of Neuromuscular Networks and Behavior. Front. Cell Dev. Biol. 2022, 9, 786993. [Google Scholar] [CrossRef] [PubMed]
- Graba, Y.; Aragnol, D.; Pradel, J. Drosophila Hox complex downstream targets and the function of homeotic genes. BioEssays: News Rev. Mol. Cell. Dev. Biol. 1997, 19, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, P.; Habib, F.; Yelagandula, R.; Shashidhara, L.S. Genome-level identification of targets of Hox protein Ultrabithorax in Drosophila: Novel mechanisms for target selection. Sci. Rep. 2011, 1, 205. [Google Scholar] [CrossRef] [PubMed]
- Domsch, K.; Carnesecchi, J.; Disela, V.; Friedrich, J.; Trost, N.; Ermakova, O.; Polychronidou, M.; Lohmann, I. The Hox transcription factor Ubx stabilizes lineage commitment by suppressing cellular plasticity in Drosophila. eLife 2019, 8, e42675. [Google Scholar] [CrossRef]
- Pavlopoulos, A.; Akam, M. Hox gene Ultrabithorax regulates distinct sets of target genes at successive stages of Drosophila haltere morphogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 2855–2860. [Google Scholar] [CrossRef]
- Skene, P.J.; Henikoff, S. An efficient targeted nuclease strategy for high-resolution mapping of DNA binding sites. eLife 2017, 6, e21856. [Google Scholar] [CrossRef]
- Skene, P.J.; Henikoff, J.G.; Henikoff, S. Targeted in situ genome-wide profiling with high efficiency for low cell numbers. Nat. Protoc. 2018, 13, 1006–1019. [Google Scholar] [CrossRef]
- Cusanovich, D.A.; Reddington, J.P.; Garfield, D.A.; Daza, R.M.; Aghamirzaie, D.; Marco-Ferreres, R.; Pliner, H.A.; Christiansen, L.; Qiu, X.; Steemers, F.J.; et al. The cis-regulatory dynamics of embryonic development at single-cell resolution. Nature 2018, 555, 538–542. [Google Scholar] [CrossRef]
- Li, H. Single-cell RNA sequencing in Drosophila: Technologies and applications. Wiley Interdiscip. Rev. Dev. Biol. 2021, 10, e396. [Google Scholar] [CrossRef]
- Zirin, J.; Bosch, J.; Viswana, R.; Mohr, S.E.; Perrimon, N. State-of-the-art CRISPR for in vivo and cell-based studies in Drosophila. Trends Genet. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Port, F.; Strein, C.; Stricker, M.; Rauscher, B.; Heigwer, F.; Zhou, J.; Beyersdörffer, C.; Frei, J.; Hess, A.; Kern, K.; et al. A large-scale resource for tissue-specific CRISPR mutagenesis in Drosophila. eLife 2020, 9, e53865. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buffry, A.D.; McGregor, A.P. Micromanagement of Drosophila Post-Embryonic Development by Hox Genes. J. Dev. Biol. 2022, 10, 13. https://doi.org/10.3390/jdb10010013
Buffry AD, McGregor AP. Micromanagement of Drosophila Post-Embryonic Development by Hox Genes. Journal of Developmental Biology. 2022; 10(1):13. https://doi.org/10.3390/jdb10010013
Chicago/Turabian StyleBuffry, Alexandra D., and Alistair P. McGregor. 2022. "Micromanagement of Drosophila Post-Embryonic Development by Hox Genes" Journal of Developmental Biology 10, no. 1: 13. https://doi.org/10.3390/jdb10010013
APA StyleBuffry, A. D., & McGregor, A. P. (2022). Micromanagement of Drosophila Post-Embryonic Development by Hox Genes. Journal of Developmental Biology, 10(1), 13. https://doi.org/10.3390/jdb10010013