Creation of Cybrid Cultures Containing mtDNA Mutations m.12315G>A and m.1555G>A, Associated with Atherosclerosis

 ,
, 

 and
and
Abstract
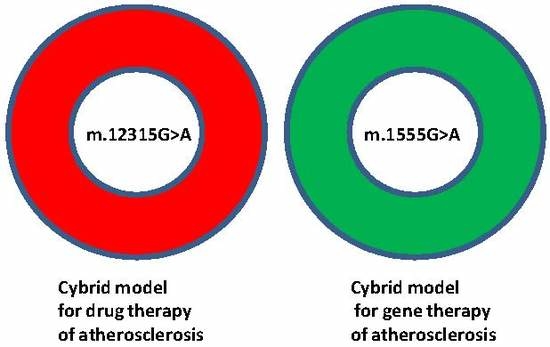
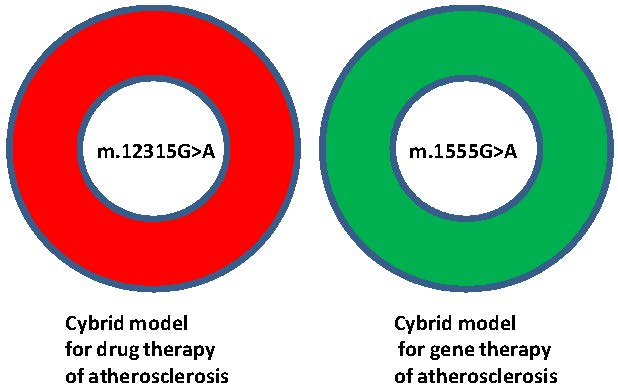
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Creation of rho0 Cell Cultures
- (1)
- With the addition of uridine and ethidium bromide;
- (2)
- With the addition of ethidium bromide only.
- (1)
- Time of cultivation;
- (2)
- Concentration of additives used in the medium.
3.2. Isolation of Platelets from Blood
3.3. Creation of Cybrid Cultures
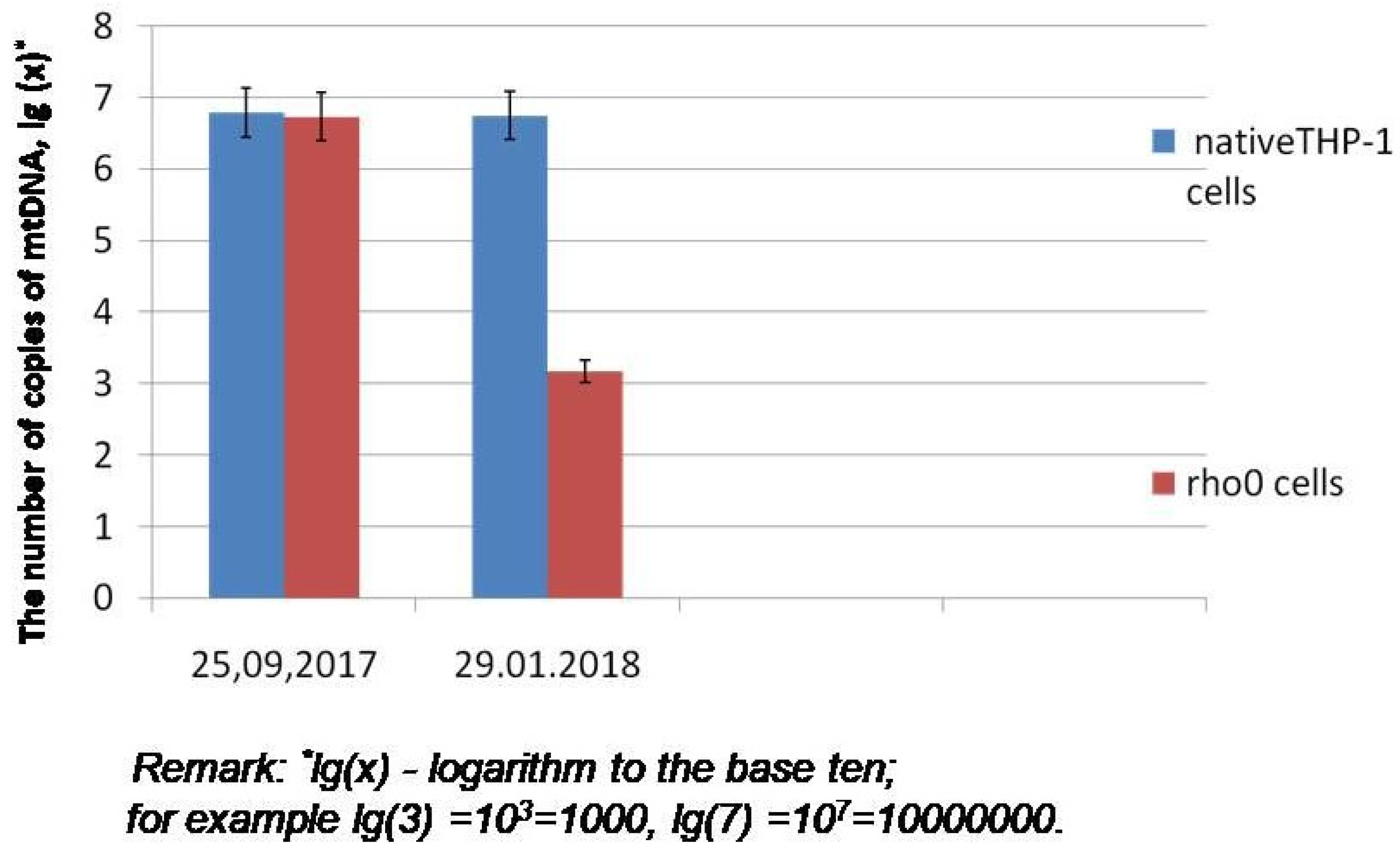
3.4. Verification of Mitochondrial Genome Copies Number in rho0 (mtDNA-less) and Cybrid Cell Cultures
3.5. Determination of the Heteroplasmy Level of Mitochondrial Genome Mutations m.12315G>A and m.1555G>A
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DNA | Deoxyribonucleic acid |
| mtDNA | mitochondrial DNA |
| DMEM | Dulbecco modified Eagle medium |
| HIV | Human immunodeficiency virus |
| FBS | Foetal bovine serum |
| MELAS | Mitochondrial encephalomyopathy, lactic acidosis, and stroke-like episodes |
| RNA | Ribonucleic acid |
| DMSO | dimethylsulphoxide |
| PEG | polyethylene glycol |
References
- Arduíno, D.M.; Esteves, A.R.; Swerdlow, R.H.; Cardoso, S.M. A cybrid cell model for the assessment of the link between mitochondrial deficits and sporadic Parkinson’s disease. Methods Mol. Biol. 2015, 1265, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Maraver, J.; Cordero, M.D.; Moñino, I.D.; Pereira-Arenas, S.; Lechuga-Vieco, A.V.; Cotán, D.; De la Mata, M.; Oropesa-Ávila, M.; De Miguel, M.; Bautista Lorite, J.; et al. Screening of effective pharmacological treatments for MELAS syndrome using yeasts, fibroblasts and cybrid models of the disease. Br. J. Pharmacol. 2012, 167, 1311–1328. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Liang, M.; Zhang, C.; Zhao, X.; He, Q.; Cui, L.; Liu, X.; Sun, Y.H.; Fu, Q.; Ji, Y.; et al. Biochemical evidence for a mitochondrial genetic modifier in the phenotypic manifestation of Leber’s hereditary optic neuropathy-associated mitochondrial DNA mutation. Hum. Mol. Genet. 2016, 25, 3613–3625. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.H.; Yum, K.S.; Chang, J.Y.; Kim, M.; Ahn, J.Y.; Kim, S.; Lapchak, P.A.; Han, M.K. Dose-specific effect of simvastatin on hypoxia-induced HIF-1α and BACE expression in Alzheimer’s disease cybrid cells. BMC Neurol. 2015, 15, 127. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, H.M.; Carl, S.M.; Swerdlow, R.H. Cytoplasmic hybrid (cybrid) cell lines as a practical model for mitochondriopathies. Redox. Biol. 2014, 2, 619–631. [Google Scholar] [CrossRef] [PubMed]
- van Capelleveen, J.C.; Bochem, A.E.; Boekholdt, S.M.; Mora, S.; Hoogeveen, R.C.; Ballantyne, C.M.; Ridker, P.M.; Sun, W.; Barter, P.J.; Tall, A.R.; et al. Association of High-Density Lipoprotein-Cholesterol Versus Apolipoprotein A-I With Risk of Coronary Heart Disease: The European Prospective Investigation Into Cancer-Norfolk Prospective Population Study, the Atherosclerosis Risk in Communities Study, and the Women’s Health Study. J. Am. Heart Assoc. 2017, 6, e006636. [Google Scholar] [CrossRef]
- Colantonio, L.D.; Gamboa, C.M.; Richman, J.S.; Levitan, E.B.; Soliman, E.Z.; Howard, G.; Safford, M.M. Black-White Differences in Incident Fatal, Nonfatal, and Total Coronary Heart Disease. Circulation. 2017, 136, 152–166. [Google Scholar] [CrossRef] [PubMed]
- Halcox, J.P.; Banegas, J.R.; Roy, C.; Dallongeville, J.; De Backer, G.; Guallar, E.; Perk, J.; Hajage, D.; Henriksson, K.M.; Borghi, C. Prevalence and treatment of atherogenic dyslipidemia in the primary prevention of cardiovascular disease in Europe: EURIKA, a cross-sectional observational study. BMC Cardiovasc. Disord. 2017, 17, 160. [Google Scholar] [CrossRef]
- Kubota, Y.; Heiss, G.; MacLehose, R.F.; Roetker, N.S.; Folsom, A.R. Association of Educational Attainment with Lifetime Risk of Cardiovascular Disease: The Atherosclerosis Risk in Communities Study. JAMA Intern. Med. 2017, 177, 1165–1172. [Google Scholar] [CrossRef]
- Kalbaugh, C.A.; Kucharska-Newton, A.; Wruck, L.; Lund, J.L.; Selvin, E.; Matsushita, K.; Bengtson, L.G.S.; Heiss, G.; Loehr, L. Peripheral Artery Disease Prevalence and Incidence Estimated from Both Outpatient and Inpatient Settings Among Medicare Fee-for-Service Beneficiaries in the Atherosclerosis Risk in Communities (ARIC) Study. J. Am. Heart Assoc. 2017, 6, e003796. [Google Scholar] [CrossRef]
- Alipov, V.I.; Sukhorukov, V.N.; Karagodin, V.P.; Grechko, A.V.; Orekhov, A.N. Chemical composition of circulating native and desialylated low density lipoprotein: What is the difference? Vessel Plus. 2017, 1, 107–115. [Google Scholar] [CrossRef]
- Sazonova, M.A.; Ryzhkova, A.I.; Sinyov, V.V.; Sazonova, M.D.; Khasanova, Z.B.; Nikitina, N.A.; Karagodin, V.P.; Orekhov, A.N.; Sobenin, I.A. Creation of Cultures Containing Mutations Linked with Cardiovascular Diseases using Transfection and Genome Editing. Curr. Pharm. Des. 2019, 25, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Sinyov, V.V.; Sazonova, M.A.; Ryzhkova, A.I.; Galitsyna, E.V.; Melnichenko, A.A.; Postnov, A.Y.; Orekhov, A.N.; Grechko, A.V.; Sobenin, I.A. Potential use of buccal epithelium for genetic diagnosis of atherosclerosis using mtDNA mutations. Vessel Plus. 2017, 1, 145–150. [Google Scholar] [CrossRef][Green Version]
- Orekhov, A.N.; Ivanova, E.A. Introduction of the special issue “Atherosclerosis and Related Diseases”. Vessel Plus. 2017, 1, 163–165. [Google Scholar] [CrossRef]
- Strassheim, D.; Karoor, V.; Stenmark, K.; Verin, A.; Gerasimovskaya, E. A current view of G protein-coupled receptor-mediated signaling in pulmonary hypertension: Finding opportunities for therapeutic intervention. Vessel Plus. 2018, 2, 29. [Google Scholar] [CrossRef] [PubMed]
- Sazonova, M.A.; Ryzhkova, A.I.; Sinyov, V.V.; Galitsyna, E.V.; Melnichenko, A.A.; Demakova, N.A.; Sobenin, I.A.; Shkurat, T.P.; Orekhov, A.N. Mitochondrial Genome Mutations Associated with Myocardial Infarction. Dis. Markers. 2018, 2018, 9749457. [Google Scholar] [CrossRef] [PubMed]
- Sazonova, M.A.; Ryzhkova, A.I.; Sinyov, V.V.; Sazonova, M.D.; Nikitina, N.A.; Shkurat, T.P.; Sobenin, I.A.; Orekhov, A.N. Mitochondrial mutations associated with cardiac angina. Vessel Plus. 2019, 3, 8. [Google Scholar] [CrossRef]
- Sazonova, M.A.; Sinyov, V.V.; Ryzhkova, A.I.; Sazonova, M.D.; Khasanova, Z.B.; Sobenin, I.A. MtDNA mutations linked with left ventricular hypertrophy. Vessel Plus. 2019, 3, 5. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, C.; Jin, Y.; Yang, Q.; Meng, Q.; Liu, Q.; Dai, Y.; Cai, L.; Liu, Z.; Liu, K.; et al. Activating the PGC-1α/TERT Pathway by Catalpol Ameliorates Atherosclerosis via Modulating ROS Production, DNA Damage, and Telomere Function: Implications on Mitochondria and Telomere Link. Oxid. Med. Cell Longev. 2018, 2018, 2876350. [Google Scholar] [CrossRef]
- Salazar, G. NADPH Oxidases and Mitochondria in Vascular Senescence. Int. J. Mol. Sci. 2018, 19, 1327. [Google Scholar] [CrossRef]
- Dikalov, S. Cross talk between mitochondria and NADPH oxidases. Free Radic. Biol. Med. 2011, 51, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
- McCully, K.S. Communication: Melatonin, Hyperhomocysteinemia, Thioretinaco Ozonide, Adenosylmethionine and Mitochondrial Dysfunction in Aging and Dementia. Ann. Clin. Lab. Sci. 2018, 48, 126–131. [Google Scholar] [PubMed]
- Docherty, C.K.; Carswell, A.; Friel, E.; Mercer, J.R. Impaired mitochondrial respiration in human carotid plaque atherosclerosis: A potential role for Pink1 in vascular smooth muscle cell energetics. Atherosclerosis. 2018, 268, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.P.K.; Reinhold, J.; Yu, H.; Starks, L.; Uryga, A.K.; Foote, K.; Finigan, A.; Figg, N.; Pung, Y.F.; Logan, A.; et al. Mitochondrial Respiration Is Reduced in Atherosclerosis, Promoting Necrotic Core Formation and Reducing Relative Fibrous Cap Thickness. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 2322–2332. [Google Scholar] [CrossRef] [PubMed]
- Sazonova, M.; Budnikov, E.; Khasanova, Z.; Sobenin, I.; Postnov, A.; Orekhov, A. Studies of the human aortic intima by a direct quantitative assay of mutant alleles in the mitochondrial genome. Atherosclerosis. 2009, 204, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Sazonova, M.A.; Sinyov, V.V.; Barinova, V.A.; Ryzhkova, A.I.; Zhelankin, A.V.; Postnov, A.Y.; Sobenin, I.A.; Bobryshev, Y.V.; Orekhov, A.N. Mosaicism of Mitochondrial Genetic Variation in Atherosclerotic Lesions of the Human Aorta. Biomed. Res. Int. 2015, 2015, 825468. [Google Scholar] [CrossRef]
- Sazonova, M.A.; Sinyov, V.V.; Ryzhkova, A.I.; Galitsyna, E.V.; Khasanova, Z.B.; Postnov, A.Y.; Yarygina, E.I.; Orekhov, A.N.; Sobenin, I.A. Role of Mitochondrial Genome Mutations in Pathogenesis of Carotid Atherosclerosis. Oxid. Med. Cell Longev. 2017, 2017, 6934394. [Google Scholar] [CrossRef] [PubMed]
- Sazonova, M.A.; Ryzhkova, A.I.; Sinyov, V.V.; Galitsyna, E.V.; Orekhova, V.A.; Melnichenko, A.A.; Orekhov, A.N.; Ravani, A.L.; Sobenin, I.A. New markers of atherosclerosis: A threshold level of heteroplasmy in mtDNA mutations. Vessel Plus. 2017, 1, 182–191. [Google Scholar] [CrossRef]
- Sazonova, M.A.; Postnov, A.Y.; Orekhov, A.N.; Sobenin, I.A. A new method of quantitative estimation of mutant allele in mitochondrial genome. Patol. Fiziol. Eksp. Ter. 2011, 4, 81–84. (In Russian) [Google Scholar]
- King, M.P.; Attardi, G. Isolation of human cell lines lacking mitochondrial DNA. Methods Enzymol. 1996, 264, 304–313. [Google Scholar] [CrossRef]
- Sazonova, M.A.; Chicheva, M.M.; Zhelankin, A.V.; Sobenin, I.A.; Bobryshev, Y.V.; Orekhov, A.N. Association of mutations in the mitochondrial genome with the subclinical carotid atherosclerosis in women. Exp. Mol. Pathol. 2015, 99, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Sobenin, I.A.; Sazonova, M.A.; Sinyov, V.V.; Ryzhkova, A.I.; Galitsyna, E.V.; Khazanova, Z.B.; Postnov, A.Y.; Yarygina, E.I.; Shkurat, T.P.; Orekhov, A.N. Response to: Comment on “Role of Mitochondrial Genome Mutations in Pathogenesis of Carotid Atherosclerosis”. Oxid. Med. Cell Longev. 2018, 2018, 7620234. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Growth Medium | ||
| Reagent | Concentration | Company |
| RPMI-1640 | - | Gibco |
| l-glutamine | 300 mg/L | Paneco |
| 2-mercaptoethanol | 2 × 10−5 M | Acros |
| Foetal bovine serum (FBS) | 10% of the total volume | HyClone |
| Penicillin-streptomycin, 100x solution | penicillin—50 U/mL, streptomycin—50 µg/ml | Paneco |
| Dextroglucose | 2500 mg/L | Amresco |
| Sodium pyruvate | 110 µg/L | Paneco |
| 10X Sodium Citrate in Physiological Solution | ||
| Reagents | Concentration | Company |
| Physiological solution | 0.15 M NaCl | Paneco |
| Sodium citrate (Na3C6H5O7·2H2O) trisodium salt, dihydrate) | 0.10 M | Amresco |
| Polyethylene glycol solution | ||
| Polyethylene glycol 1500 | 42% of the total volume | Acros |
| Dimethylsulphoxide(DMSO) | 2 mL | Amresco |
| DMEM [-Ca2+] | 9.5 mL | Gibco |
| Ready-to-Use Solutions and Media | ||
| Reagents | Concentration | Company |
| Uridine | 50 mg/mL solution | Sigma-Aldrich |
| Ethidium bromide | 1% solution | AppliChem |
| Ficoll-urografin | density 1.077 | Paneco |
| DMSO | Amresco | |
| Physiological solution | 0.15 M NaCl | Paneco |
| DMEM [-Ca2+] | Gibco | |
| Reagent kit for conducting real-time PCR in the presence of SYBR Green I | Synthol | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sazonova, M.A.; Sinyov, V.V.; Ryzhkova, A.I.; Sazonova, M.D.; Khasanova, Z.B.; Shkurat, T.P.; Karagodin, V.P.; Orekhov, A.N.; Sobenin, I.A. Creation of Cybrid Cultures Containing mtDNA Mutations m.12315G>A and m.1555G>A, Associated with Atherosclerosis. Biomolecules 2019, 9, 499. https://doi.org/10.3390/biom9090499
Sazonova MA, Sinyov VV, Ryzhkova AI, Sazonova MD, Khasanova ZB, Shkurat TP, Karagodin VP, Orekhov AN, Sobenin IA. Creation of Cybrid Cultures Containing mtDNA Mutations m.12315G>A and m.1555G>A, Associated with Atherosclerosis. Biomolecules. 2019; 9(9):499. https://doi.org/10.3390/biom9090499
Chicago/Turabian StyleSazonova, Margarita A., Vasily V. Sinyov, Anastasia I. Ryzhkova, Marina D. Sazonova, Zukhra B. Khasanova, Tatiana P. Shkurat, Vasily P. Karagodin, Alexander N. Orekhov, and Igor A. Sobenin. 2019. "Creation of Cybrid Cultures Containing mtDNA Mutations m.12315G>A and m.1555G>A, Associated with Atherosclerosis" Biomolecules 9, no. 9: 499. https://doi.org/10.3390/biom9090499
APA StyleSazonova, M. A., Sinyov, V. V., Ryzhkova, A. I., Sazonova, M. D., Khasanova, Z. B., Shkurat, T. P., Karagodin, V. P., Orekhov, A. N., & Sobenin, I. A. (2019). Creation of Cybrid Cultures Containing mtDNA Mutations m.12315G>A and m.1555G>A, Associated with Atherosclerosis. Biomolecules, 9(9), 499. https://doi.org/10.3390/biom9090499