There Is Treasure Everywhere: Reductive Plastid Evolution in Apicomplexa in Light of Their Close Relatives


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Historical Perspective
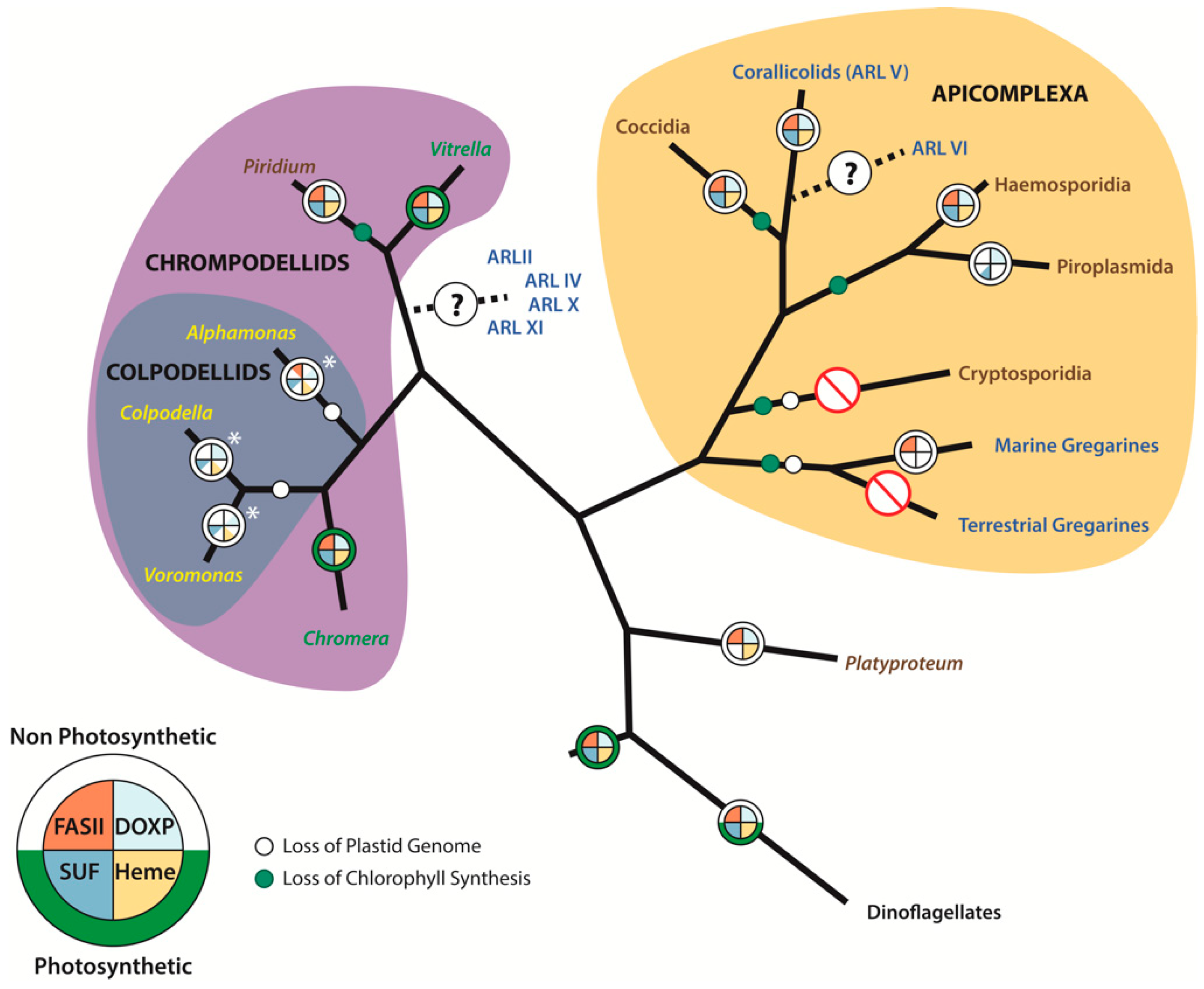
3. Apicomplexan Relatives and the Origins and Evolution of the Apicoplast
3.1. Chromerids
3.2. Colpodellids
3.3. Corallicolids
3.4. New Parasitic Lineages–Piridium and Platyproteum
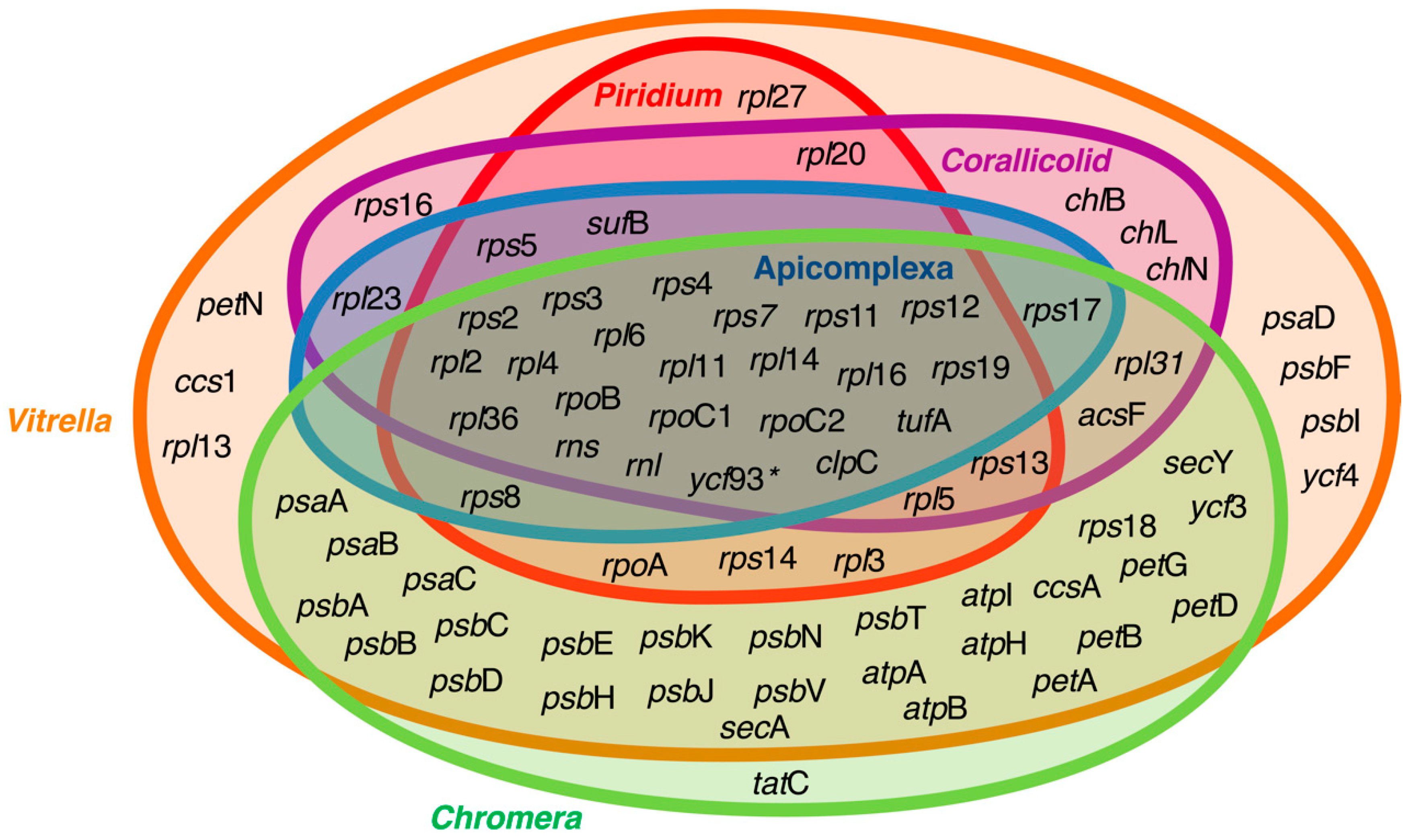
4. Plastid Genome Evolution within the Platyproteum-Chrompodellids-Apicomplexa Lineage
5. Apicoplast Metabolism
5.1. Heme Synthesis Pathway
5.2. Type II Fatty Acid Synthesis Pathway
5.3. MEP/DOXP Pathway
5.4. SUF Pathway
6. Hidden Treasure
7. What Gene Holds the Key to Apicoplast Retention?
8. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Adl, S.M.; Leander, B.S.; Simpson, A.G.B.; Archibald, J.M.; Anderson, O.R.; Bass, D.; Bowser, S.S.; Brugerolle, G.; Farmer, M.A.; Karpov, S.; et al. Diversity, nomenclature, and taxonomy of protists. Syst. Biol. 2007, 56, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Leander, B.; Clopton, R.; Keeling, P. Phylogeny of gregarines (Apicomplexa) as inferred from small-subunit rDNA and beta-tubulin. Int. J. Syst. Evol. Microbiol. 2003, 53, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Gentil, J.; Hempel, F.; Moog, D.; Zauner, S.; Maier, U.G. Review: Origin of complex algae by secondary endosymbiosis: A journey through time. Protoplasma 2017, 254, 1835–1843. [Google Scholar] [CrossRef] [PubMed]
- Archibald, J.M. Endosymbiosis and Eukaryotic Cell Evolution. Curr. Biol. 2015, 25, R911–R921. [Google Scholar] [CrossRef] [PubMed]
- Fussy, Z.; Obornik, M. Chromerids and their plastids. In Advances in Botanical Research; Hirakawa, Y., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 84, pp. 187–218. [Google Scholar]
- Waller, R.F.; Kořený, L. Chapter Four—Plastid complexity in dinoflagellates: A picture of gains, losses, replacements and revisions. In Advances in Botanical Research; Hirakawa, Y., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 84, pp. 105–143. [Google Scholar]
- Wetherbee, R.; Jackson, C.J.; Repetti, S.I.; Clementson, L.A.; Costa, J.F.; van de Meene, A.; Crawford, S.; Verbruggen, H. The golden paradox—A new heterokont lineage with chloroplasts surrounded by two membranes. J. Phycol. 2019, 55, 257–278. [Google Scholar] [CrossRef] [PubMed]
- Lemgruber, L.; Kudryashev, M.; Dekiwadia, C.; Riglar, D.T.; Baum, J.; Stahlberg, H.; Ralph, S.A.; Frischknecht, F. Cryo-electron tomography reveals four-membrane architecture of the Plasmodium apicoplast. Malaria J. 2013, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Köhler, S.; Delwiche, C.F.; Denny, P.W.; Tilney, L.G.; Webster, P.; Wilson, R.J.M.; Palmer, J.D.; Roos, D.S. A plastid of probable green algal origin in apicomplexan parasites. Science 1997, 275, 1485–1489. [Google Scholar] [CrossRef] [PubMed]
- Diniz, J.A.P.; Silva, E.O.; Lainson, R.; de Souza, W. The fine structure of Garnia gonadati and its association with the host cell. Parasitol. Res. 2000, 86, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Tomova, C.; Geerts, W.J.C.; Müller-Reichert, T.; Entzeroth, R.; Humbel, B.M. New comprehension of the apicoplast of Sarcocystis by transmission electron tomography. Biol. Cell 2006, 98, 535–545. [Google Scholar] [CrossRef]
- Boucher, M.J.; Yeh, E. Plastid–endomembrane connections in apicomplexan parasites. PLoS Pathog. 2019, 15, 1–9. [Google Scholar] [CrossRef]
- Waller, R.; McFadden, G. The Apicoplast: A review of the derived plastid of apicomplexan parasites. Curr. Issues Mol. Biol. 2005, 7, 57–80. [Google Scholar]
- Striepen, B.; Crawford, M.J.; Shaw, M.K.; Tilney, L.G.; Seeber, F.; Roos, D.S. The plastid of Toxoplasma gondii is divided by association with the centrosomes. J. Cell Biol. 2000, 151, 1423–1434. [Google Scholar] [CrossRef]
- Janouskovec, J.; Horak, A.; Obornik, M.; Lukes, J.; Keeling, P.J. A common red algal origin of the apicomplexan, dinoflagellate, and heterokont plastids. Proc. Natl. Acad. Sci. USA 2010, 107, 10949–10954. [Google Scholar] [CrossRef]
- Fussy, Z.; Obornik, M. Complex Endosymbioses I: From Primary to Complex Plastids, Multiple Independent Events. In Plastids; Marechal, E., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1829, pp. 17–35. [Google Scholar]
- Fichera, M.E.; Roos, D.S. A plastid organelle as a drug target in apicomplexan parasites. Nature 1997, 390, 407–409. [Google Scholar] [CrossRef]
- McFadden, G.; Reith, M.; Munholland, J.; LangUnnasch, N. Plastid in human parasites. Nature 1996, 381, 482. [Google Scholar] [CrossRef]
- Moore, R.B.; Obornik, M.; Janouskovec, J.; Chrudimsky, T.; Vancova, M.; Green, D.H.; Wright, S.W.; Davies, N.W.; Bolch, C.J.S.; Heimann, K.; et al. A photosynthetic alveolate closely related to apicomplexan parasites. Nature 2008, 451, 959–963. [Google Scholar] [CrossRef]
- Kwong, W.K.; del Campo, J.; Mathur, V.; Vermeij, M.J.A.; Keeling, P.J. A widespread coral-infecting apicomplexan with chlorophyll biosynthesis genes. Nature 2019, 568, 103–107. [Google Scholar] [CrossRef]
- Mathur, V.; del Campo, J.; Kolisko, M.; Keeling, P.J. Global diversity and distribution of close relatives of apicomplexan parasites. Environ. Microbiol. 2018, 20, 2824–2833. [Google Scholar] [CrossRef]
- Hepler, P.; Huff, C.; Sprinz, H. Fine structure of exoerythrocytic stages of Plasmodium fallax. J. Cell Biol. 1966, 30, 333–358. [Google Scholar] [CrossRef]
- Gardner, M.; Feagin, J.; Moore, D.; Spencer, D.; Gray, M.; Williamson, D.; Wilson, R. Organization and expression of small subunit ribosomal-RNA genes encoded by a 35-kilobase circular DNA in Plasmodium falciparum. Mol. Biochem. Parasitol. 1991, 48, 77–88. [Google Scholar] [CrossRef]
- Palmer, J.D. Green ancestry of malarial parasites? Curr. Biol. 1992, 2, 318–320. [Google Scholar] [CrossRef]
- Vaidya, A.B.; Akella, R.; Suplick, K. Sequences similar to genes for two mitochondrial proteins and portions of ribosomal RNA in tandemly arrayed 6-kilobase-pair DNA of a malarial parasite. Mol. Biol. Parasitol. 1989, 35, 97–107. [Google Scholar] [CrossRef]
- Blanchard, J.; Hicks, J. The non-photosynthetic plastid in malarial parasites and other apicomplexans is derived from outside the green plastid lineage. J. Eukaryot. Microbiol. 1999, 46, 367–375. [Google Scholar] [CrossRef]
- Fast, N.; Kissinger, J.; Roos, D.; Keeling, P. Nuclear-encoded, plastid-targeted genes suggest a single common origin for apicomplexan and dinoflagellate plastids. Mol. Biol. Evol. 2001, 18, 418–426. [Google Scholar] [CrossRef]
- Funes, S.; Reyes-Prieto, A.; Perez-Martinez, X.; Gonzalez-Halphen, D. On the evolutionary origins of apicoplasts: Revisiting the rhodophyte vs. chlorophyte controversy. Microbes Infect. 2004, 6, 305–311. [Google Scholar] [CrossRef]
- Funes, S.; Davidson, E.; Reyes-Prieto, A.; Magallon, S.; Herion, P.; King, M.; Gonzalez-Halphen, D. A green algal apicoplast ancestor. Science 2002, 298, 2155. [Google Scholar] [CrossRef]
- Huang, Y.; He, L.; Hu, J.; He, P.; He, J.; Yu, L.; Malobi, N.; Zhou, Y.; Shen, B.; Zhao, J. Characterization and annotation of Babesia orientalis apicoplast genome. Parasite Vector 2015, 8, 543. [Google Scholar] [CrossRef]
- Cai, X.; Fuller, A.L.; McDougald, L.R.; Zhu, G. Apicoplast genome of the coccidian Eimeria tenella. Gene 2003, 321, 39–46. [Google Scholar] [CrossRef]
- Arisue, N.; Hashimoto, T.; Mitsui, H.; Palacpac, N.M.Q.; Kaneko, A.; Kawai, S.; Hasegawa, M.; Tanabe, K.; Horii, T. The Plasmodium apicoplast genome: Conserved structure and close relationship of P. ovale to rodent malaria parasites. Mol. Biol. Evol. 2012, 29, 2095–2099. [Google Scholar] [CrossRef]
- Waller, R.; Keeling, P.; Donald, R.; Striepen, B.; Handman, E.; Lang-Unnasch, N.; Cowman, A.; Besra, G.; Roos, D.; McFadden, G. Nuclear-encoded proteins target to the plastid in Toxoplasma gondii and Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 1998, 95, 12352–12357. [Google Scholar] [CrossRef]
- Ralph, S.A.; van Dooren, G.G.; Waller, R.F.; Crawford, M.J.; Fraunholz, M.J.; Foth, B.J.; Tonkin, C.J.; Roos, D.S.; McFadden, G.I. Metabolic maps and functions of the Plasmodium falciparum apicoplast. Nat. Rev. Microbiol. 2004, 2, 203–216. [Google Scholar] [CrossRef]
- Lim, L.; McFadden, G.I. The evolution, metabolism and functions of the apicoplast. Philos. Trans. R. Soc. B 2010, 365, 749–763. [Google Scholar] [CrossRef]
- Zhu, G.; Marchewka, M.J.; Keithly, J.S. Cryptosporidium parvum appears to lack a plastid genome. Microbiology 2000, 146, 315–321. [Google Scholar] [CrossRef]
- Toso, M.A.; Omoto, C.K. Gregarina niphandroides may lack both a plastid genome and organelle. J. Eukaryot. Microbiol. 2007, 54, 66–72. [Google Scholar] [CrossRef]
- Schrevel, J. Observations biologiques et ultrastructurales sur les selenidiidaeet leurs consequences sur la systematique des gregarinomorphes. J. Protozool. 1971, 18, 448–470. [Google Scholar] [CrossRef]
- Cumbo, V.R.; Baird, A.H.; Moore, R.B.; Negri, A.P.; Neilan, B.A.; Salih, A.; van Oppen, M.J.H.; Wang, Y.; Marquis, C.P. Chromera velia is endosymbiotic in larvae of the reef corals Acropora digitifera and A. tenuis. Protist 2013, 164, 237–244. [Google Scholar] [CrossRef]
- Foster, C.; Portman, N.; Chen, M.; Šlapeta, J. Increased growth and pigment content of Chromera velia in mixotrophic culture. FEMS Microbiol. Ecol. 2014, 88, 121–128. [Google Scholar] [CrossRef]
- Mohamed, A.R.; Cumbo, V.R.; Harii, S.; Shinzato, C.; Chan, C.X.; Ragan, M.A.; Satoh, N.; Ball, E.E.; Miller, D.J. Deciphering the nature of the coral—Chromera association. ISME J. 2018, 12, 776–790. [Google Scholar] [CrossRef]
- Voolstra, C.R.; Schwarz, J.A.; Schnetzer, J.; Sunagawa, S.; Desalvo, M.K.; Szmant, A.M.; Coffroth, M.A.; Medina, M. The host transcriptome remains unaltered during the establishment of coral–algal symbioses. Mol. Ecol. 2009, 18, 1823–1833. [Google Scholar] [CrossRef]
- Obornik, M.; Modry, D.; Lukes, M.; Cernotikova-Stribrna, E.; Cihlar, J.; Tesarova, M.; Kotabova, E.; Vancova, M.; Prasil, O.; Lukes, J. Morphology, Ultrastructure and life cycle of Vitrella brassicaformis n. sp., n. gen., a novel Chromerid from the Great Barrier Reef. Protist 2012, 163, 306–323. [Google Scholar] [CrossRef]
- Woo, Y.H.; Ansari, H.; Otto, T.D.; Klinger, C.M.; Kolisko, M.; Michalek, J.; Saxena, A.; Shanmugam, D.; Tayyrov, A.; Veluchamy, A.; et al. Chromerid genomes reveal the evolutionary path from photosynthetic algae to obligate intracellular parasites. eLife 2015, 4, e06974. [Google Scholar] [CrossRef]
- Sevcikova, T.; Horak, A.; Klimes, V.; Zbrankova, V.; Demir-Hilton, E.; Sudek, S.; Jenkins, J.; Schmutz, J.; Pribyl, P.; Fousek, J.; et al. Updating algal evolutionary relationships through plastid genome sequencing: Did alveolate plastids emerge through endosymbiosis of an ochrophyte? Sci. Rep. 2015, 5, 10134. [Google Scholar] [CrossRef]
- Janouskovec, J.; Horak, A.; Barott, K.L.; Rohwer, F.L.; Keeling, P.J. Global analysis of plastid diversity reveals apicomplexan-related lineages in coral reefs. Curr. Biol. 2012, 22, R518–R519. [Google Scholar] [CrossRef]
- Brugerolle, G.; Mignot, J.P. Observations sur le cycle l’ultrastructure et la position systématique de. Spiromonas perforans 1938, 15, 183–196. [Google Scholar]
- Cavalier-Smith, T.; Chao, E.E. Protalveolate phylogeny and systematics and the origins of Sporozoa and dinoflagellates (phylum Myzozoa nom. nov.). Eur. J. Protistol. 2004, 40, 185–212. [Google Scholar] [CrossRef]
- Kuvardina, O.; Leander, B.; Aleshin, V.; Myl’nikov, A.; Keeling, P.; Simdyanov, T. The phylogeny of colpodellids (alveolata) using small subunit rRNA gene sequences suggests they are the free-living sister group to apicomplexans. J. Eucariot. Microbiol. 2002, 49, 498–504. [Google Scholar] [CrossRef]
- Gile, G.H.; Slamovits, C.H. Transcriptomic analysis reveals evidence for a cryptic plastid in the colpodellid Voromonas pontica, a close relative of chromerids and apicomplexan parasites. PLoS ONE 2014, 9, e96258. [Google Scholar] [CrossRef]
- Janouskovec, J.; Tikhonenkov, D.V.; Burki, F.; Howe, A.T.; Kolisko, M.; Mylnikov, A.P.; Keeling, P.J. Factors mediating plastid dependency and the origins of parasitism in apicomplexans and their close relatives. Proc. Natl. Acad. Sci. USA 2015, 112, 10200–10207. [Google Scholar] [CrossRef]
- Janouskovec, J.; Horak, A.; Barott, K.L.; Rohwer, F.L.; Keeling, P.J. Environmental distribution of coral-associated relatives of apicomplexan parasites. ISME J. 2013, 7, 444–447. [Google Scholar] [CrossRef]
- Toller, W.; Rowan, R.; Knowlton, N. Genetic evidence for a protozoan (phylum Apicomplexa) associated with corals of the Montastraea annularis species complex. Coral Reefs 2002, 21, 143–146. [Google Scholar]
- Patten, R. Notes on a new protozoon, Piridium sociabile n.gen., n.sp., from the foot of Buccinum undatum. Parasitology 1936, 28, 502–516. [Google Scholar] [CrossRef]
- Adl, S.M.; Simpson, A.G.B.; Lane, C.E.; Lukes, J.; Bass, D.; Bowser, S.S.; Brown, M.W.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The revised classification of eukaryotes. J. Eukaryot. Mikrobiol. 2012, 59, 429–493. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Gregarine site-heterogeneous 18S rDNA trees, revision of gregarine higher classification, and the evolutionary diversification of Sporozoa. Eur. J. Protistol. 2014, 50, 472–495. [Google Scholar] [CrossRef]
- Mathur, V.; Kolisko, M.; Hehenberger, E.; Irwin, N.A.; Leander, B.S.; Kristmundsson, Á.; Freeman, M.A.; Keeling, P.J. Multiple independent origins of apicomplexan-like parasites. Biorxiv 2019. [Google Scholar] [CrossRef]
- Barbrook, A.C.; Voolstra, C.R.; Howe, C.J. The chloroplast genome of a Symbiodinium sp. clade c3 isolate. Protist 2014, 165, 1–13. [Google Scholar] [CrossRef]
- Smith, D.R.; Keeling, P.J. Mitochondrial and plastid genome architecture: Reoccurring themes, but significant differences at the extremes. Proc. Natl. Acad. Sci. USA 2015, 112, 10177–10184. [Google Scholar] [CrossRef]
- Garg, A.; Stein, A.; Zhao, W.; Dwivedi, A.; Frutos, R.; Cornillot, E.; Ben Mamoun, C. Sequence and annotation of the apicoplast genome of the human pathogen Babesia microti. PLoS ONE 2014, 9, e107939. [Google Scholar] [CrossRef]
- Sato, S. The apicomplexan plastid and its evolution. Cell Mol. Life Sci. 2011, 68, 1285–1296. [Google Scholar] [CrossRef]
- de Vries, J.; Archibald, J.M. Plastid genomes. Curr. Biol. 2018, 28, R336–R337. [Google Scholar] [CrossRef]
- Salomaki, E.D.; Nickles, K.R.; Lane, C.E. The ghost plastid of Choreocolax polysiphoniae. J. Phycol. 2015, 51, 217–221. [Google Scholar] [CrossRef]
- Salomaki, E.D.; Lane, C.E. Molecular phylogenetics supports a clade of red algal parasites retaining native plastids: Taxonomy and terminology revised. J. Phycol. 2019, 55, 279–288. [Google Scholar] [CrossRef]
- Evans, J.R.; Amour, N.S.; Verbruggen, H.; Salomaki, E.D.; Vis, M.L. Chloroplast and mitochondrial genomes of Balbiania investiens (Balbianiales, Nemaliophycidae). Phycologia 2019, 58, 310–318. [Google Scholar] [CrossRef]
- McFadden, G.I. The apicoplast. Protoplasma 2011, 248, 641–650. [Google Scholar] [CrossRef]
- Glaser, S.; van Dooren, G.G.; Agrawal, S.; Brooks, C.F.; McFadden, G.I.; Striepen, B.; Higgins, M.K. Tic22 is an essential chaperone required for protein import into the apicoplast. J. Biol. Chem. 2012, 287, 39505–39512. [Google Scholar] [CrossRef]
- Soll, J.; Schleiff, E. Protein import into chloroplasts. Nat. Rev. Mol. Cell Biol. 2004, 5, 198–208. [Google Scholar] [CrossRef]
- Sommer, M.S.; Gould, S.B.; Lehmann, P.; Gruber, A.; Przyborski, J.M.; Maier, U.-G. Der1-mediated preprotein import into the periplastid compartment of chromalveolates? Mol. Biol. Evol. 2007, 24, 918–928. [Google Scholar] [CrossRef]
- Hempel, F.; Bullmann, L.; Lau, J.; Zauner, S.; Maier, U.G. ERAD-derived preprotein transport across the second outermost plastid membrane of diatoms. Mol. Biol. Evol. 2009, 26, 1781–1790. [Google Scholar] [CrossRef]
- Spork, S.; Hiss, J.A.; Mandel, K.; Sommer, M.; Kooij, T.W.A.; Chu, T.; Schneider, G.; Maier, U.G.; Przyborski, J.M. An unusual ERAD-like complex is targeted to the apicoplast of Plasmodium falciparum. Eukaryot. Cell 2009, 8, 1134–1145. [Google Scholar] [CrossRef]
- Nagaraj, V.A.; Arumugam, R.; Prasad, D.; Rangarajan, P.N.; Padmanaban, G. Protoporphyrinogen IX oxidase from Plasmodium falciparum is anaerobic and is localized to the mitochondrion. Mol. Biochem. Parasitol. 2010, 174, 44–52. [Google Scholar] [CrossRef]
- Nagaraj, V.A.; Prasad, D.; Arumugam, R.; Rangarajan, P.N.; Padmanaban, G. Characterization of coproporphyrinogen III oxidase in Plasmodium falciparum cytosol. Parasitol. Int. 2010, 59, 121–127. [Google Scholar] [CrossRef]
- Sato, S.; Clough, B.; Coates, L.; Wilson, R.J.M. Enzymes for heme biosynthesis are found in both the mitochondrion and plastid of the malaria parasite Plasmodium falciparum. Protist 2004, 155, 117–125. [Google Scholar] [CrossRef]
- Koreny, L.; Sobotka, R.; Janouskovec, J.; Keeling, P.J.; Obornik, M. Tetrapyrrole synthesis of photosynthetic chromerids is likely homologous to the unusual pathway of apicomplexan parasites. Plant Cell. 2011, 23, 3454–3462. [Google Scholar] [CrossRef][Green Version]
- Nagaraj, V.A.; Sundaram, B.; Varadarajan, N.M.; Subramani, P.A.; Kalappa, D.M.; Ghosh, S.K.; Padmanaban, G. Malaria parasite-synthesized heme is essential in the mosquito and liver stages and complements host heme in the blood stages of infection. PLoS Pathogens 2013, 9, 1–13. [Google Scholar] [CrossRef]
- Botte, C.Y.; Yamaryo-Botte, Y. Complex Endosymbioses II: The nonphotosynthetic plastid of apicomplexa parasites (the apicoplast) and its integrated metabolism. In Plastids; Marechal, E., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1829, pp. 37–54. [Google Scholar]
- Gornicki, P. Apicoplast fatty acid biosynthesis as a target for medical intervention in Apicomplexan parasites. Int. J. Parasitol. 2003, 33, 885–896. [Google Scholar] [CrossRef]
- Goodman, C.D.; McFadden, G.I. Fatty acid biosynthesis as a drug target in apicomplexan parasites. Curr. Drug Targets 2007, 8, 15–30. [Google Scholar] [CrossRef]
- Gardner, M.J.; Bishop, R.; Shah, T.; de Villiers, E.P.; Carlton, J.M.; Hall, N.; Ren, Q.; Paulsen, I.T.; Pain, A.; Berriman, M.; et al. Genome sequence of Theileria parva, a bovine pathogen that transforms lymphocytes. Science 2005, 309, 134–137. [Google Scholar] [CrossRef]
- Vaughan, A.M.; O’Neill, M.T.; Tarun, A.S.; Camargo, N.; Phuong, T.M.; Aly, A.S.I.; Cowman, A.F.; Kappe, S.H.I. Type II fatty acid synthesis is essential only for malaria parasite late liver stage development. Cell. Microbiol. 2009, 11, 506–520. [Google Scholar] [CrossRef]
- van Schaijk, B.C.L.; Kumar, T.R.S.; Vos, M.W.; Richman, A.; van Gemert, G.-J.; Li, T.; Eappen, A.G.; Williamson, K.C.; Morahan, B.J.; Fishbaugher, M.; et al. Type II fatty acid biosynthesis is essential for Plasmodium falciparum sporozoite development in the midgut of Anopheles mosquitoes. Eukaryot. Cell 2014, 13, 550–559. [Google Scholar] [CrossRef]
- Mazumdar, J.; Striepen, B. Make it or take it: Fatty acid metabolism of Apicomplexan parasites. Eukaryot. Cell 2007, 6, 1727–1735. [Google Scholar] [CrossRef]
- Zhu, G.; Marchewka, M.J.; Woods, K.M.; Upton, S.J.; Keithly, J.S. Molecular analysis of a Type I fatty acid synthase in Cryptosporidium parvum. Mol. Biochem. Parasitol. 2000, 105, 253–260. [Google Scholar] [CrossRef]
- Abrahamsen, M.S.; Templeton, T.J.; Enomoto, S.; Abrahante, J.E.; Zhu, G.; Lancto, C.A.; Deng, M.; Liu, C.; Widmer, G.; Tzipori, S.; et al. Complete genome sequence of the apicomplexan, Cryptosporidium parvum. Science 2004, 304, 441. [Google Scholar] [CrossRef]
- Caballero, M.C.; Pedroni, M.J.; Palmer, G.H.; Suarez, C.E.; Davitt, C.; Lau, A.O.T. Characterization of acyl carrier protein and LytB in Babesia bovis apicoplast. Mol. Biochem. Parasit. 2012, 181, 125–133. [Google Scholar] [CrossRef]
- Cassera, M.; Gozzo, F.; D’Alexandri, F.; Merino, E.; del Portillo, H.; Peres, V.; Almeida, I.; Eberlin, M.; Wunderlich, G.; Wiesner, J.; et al. The methylerythritol phosphate pathway is functionally active in all intraerythrocytic stages of Plasmodium falciparum. J. Biol. Chem. 2004, 279, 51749–51759. [Google Scholar] [CrossRef]
- Yeh, E.; DeRisi, J.L. Chemical rescue of malaria parasites lacking an apicoplast defines organelle function in blood-stage Plasmodium falciparum. PLoS Biol. 2011, 9, e1001138. [Google Scholar] [CrossRef]
- Suazo, K.F.; Schaber, C.; Palsuledesai, C.C.; Odom John, A.R.; Distefano, M.D. Global proteomic analysis of prenylated proteins in Plasmodium falciparum using an alkyne-modified isoprenoid analogue. Sci. Rep. 2016, 6, 38615. [Google Scholar] [CrossRef]
- Gisselberg, J.E.; Zhang, L.; Elias, J.E.; Yeh, E. The prenylated proteome of Plasmodium falciparum reveals pathogen-specific prenylation activity and drug mechanism-of-action. Mol. Cell. Proteom. 2017, 16, S54. [Google Scholar] [CrossRef]
- Johnson, D.; Dean, D.; Smith, A.; Johnson, M. Structure, function, and formation of biological iron-sulfur clusters. Annu. Rev. Biochem. 2005, 74, 247–281. [Google Scholar] [CrossRef]
- Gisselberg, J.E.; Dellibovi-Ragheb, T.A.; Matthews, K.A.; Bosch, G.; Prigge, S.T. The suf iron-sulfur cluster synthesis pathway is required for apicoplast maintenance in malaria parasites. PLoS Pathogens 2013, 9, e1003655. [Google Scholar] [CrossRef]
- Strassert, J.F.H.; Karnkowska, A.; Hehenberger, E.; del Campo, J.; Kolisko, M.; Okamoto, N.; Burki, F.; Janouskovec, J.; Poirier, C.; Leonard, G.; et al. Single cell genomics of uncultured marine alveolates shows paraphyly of basal dinoflagellates. ISME J. 2018, 12, 304–308. [Google Scholar] [CrossRef]
- Gawryluk, R.M.R.; del Campo, J.; Okamoto, N.; Strassert, J.F.H.; Lukeš, J.; Richards, T.A.; Worden, A.Z.; Santoro, A.E.; Keeling, P.J. Morphological identification and single-cell genomics of marine diplonemids. Curr. Biol. 2016, 26, 3053–3059. [Google Scholar] [CrossRef]
- Kolisko, M.; Boscaro, V.; Burki, F.; Lynn, D.H.; Keeling, P.J. Single-cell transcriptomics for microbial eukaryotes. Curr. Biol. 2014, 24, R1081–R1082. [Google Scholar] [CrossRef]
- Picelli, S.; Faridani, O.R.; Björklund, Å.K.; Winberg, G.; Sagasser, S.; Sandberg, R. Full-length RNA-seq from single cells using Smart-seq2. Nat. Protocols 2014, 9, 171. [Google Scholar] [CrossRef]
- Leger, M.M.; Kolisko, M.; Kamikawa, R.; Stairs, C.W.; Kume, K.; Cepicka, I.; Silberman, J.D.; Andersson, J.O.; Xu, F.; Yabuki, A.; et al. Organelles that illuminate the origins of Trichomonas hydrogenosomes and Giardia mitosomes. Nat. Rvol. Evol. 2017, 1, 0092. [Google Scholar] [CrossRef]
- Vacek, V.; Novak, L.V.F.; Treitli, S.C.; Taborsky, P.; Cepicka, I.; Kolisko, M.; Keeling, P.J.; Hampl, V. Fe-S cluster assembly in oxymonads and related protists. Mol. Biol. Evol. 2018, 35, 2712–2718. [Google Scholar] [CrossRef]
- McFadden, G.I.; Yeh, E. The apicoplast: Now you see it, now you don’t. Int. J. Parasitol. 2017, 47, 137–144. [Google Scholar] [CrossRef]
- Tovar, J.; León-Avila, G.; Sánchez, L.B.; Sutak, R.; Tachezy, J.; van der Giezen, M.; Hernández, M.; Müller, M.; Lucocq, J.M. Mitochondrial remnant organelles of Giardia function in iron-sulphur protein maturation. Nature 2003, 426, 172–176. [Google Scholar] [CrossRef]
- Lill, R.; Dutkiewicz, R.; Freibert, S.A.; Heidenreich, T.; Mascarenhas, J.; Netz, D.J.; Paul, V.D.; Pierik, A.J.; Richter, N.; Stümpfig, M.; et al. The role of mitochondria and the CIA machinery in the maturation of cytosolic and nuclear iron–sulfur proteins. Eur. J. Cell Biol. 2015, 94, 280–291. [Google Scholar] [CrossRef]
- Karnkowska, A.; Vacek, V.; Zubáčová, Z.; Treitli, S.C.; Petrželková, R.; Eme, L.; Novák, L.; Žárský, V.; Barlow, L.D.; Herman, E.K.; et al. A eukaryote without a mitochondrial organelle. Curr. Biol. 2016, 26, 1274–1284. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Liu, J.; Liu, A.; He, X.; Xu, J.; Li, Z.; Zhao, S.; Li, Y.; Yin, H.; et al. Comparative analysis of apicoplast genomes of Babesia infective to small ruminants in China. Parasites Vectors 2019, 12, 312. [Google Scholar] [CrossRef]
- Barbrook, A.C.; Howe, C.J.; Purton, S. Why are plastid genomes retained in non-photosynthetic organisms? Trends Plant Sci. 2006, 11, 101–108. [Google Scholar] [CrossRef]
- Goodman, C.D.; McFadden, G.I. Ycf93 (Orf105), a small Apicoplast-encoded membrane protein in the relict plastid of the malaria parasite Plasmodium falciparum that is conserved in apicomplexa. PLoS ONE 2014, 9, e91178. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salomaki, E.D.; Kolisko, M. There Is Treasure Everywhere: Reductive Plastid Evolution in Apicomplexa in Light of Their Close Relatives. Biomolecules 2019, 9, 378. https://doi.org/10.3390/biom9080378
Salomaki ED, Kolisko M. There Is Treasure Everywhere: Reductive Plastid Evolution in Apicomplexa in Light of Their Close Relatives. Biomolecules. 2019; 9(8):378. https://doi.org/10.3390/biom9080378
Chicago/Turabian StyleSalomaki, Eric D., and Martin Kolisko. 2019. "There Is Treasure Everywhere: Reductive Plastid Evolution in Apicomplexa in Light of Their Close Relatives" Biomolecules 9, no. 8: 378. https://doi.org/10.3390/biom9080378
APA StyleSalomaki, E. D., & Kolisko, M. (2019). There Is Treasure Everywhere: Reductive Plastid Evolution in Apicomplexa in Light of Their Close Relatives. Biomolecules, 9(8), 378. https://doi.org/10.3390/biom9080378