Structural Lesions of Proteins Connected to Lipid Membrane Damages Caused by Radical Stress: Assessment by Biomimetic Systems and Raman Spectroscopy

 ,
, 

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Protein Models and Their Analysis
2.2. Radiolytic Production of Transients
2.3. Biomimetic Models
3. Results and Discussion
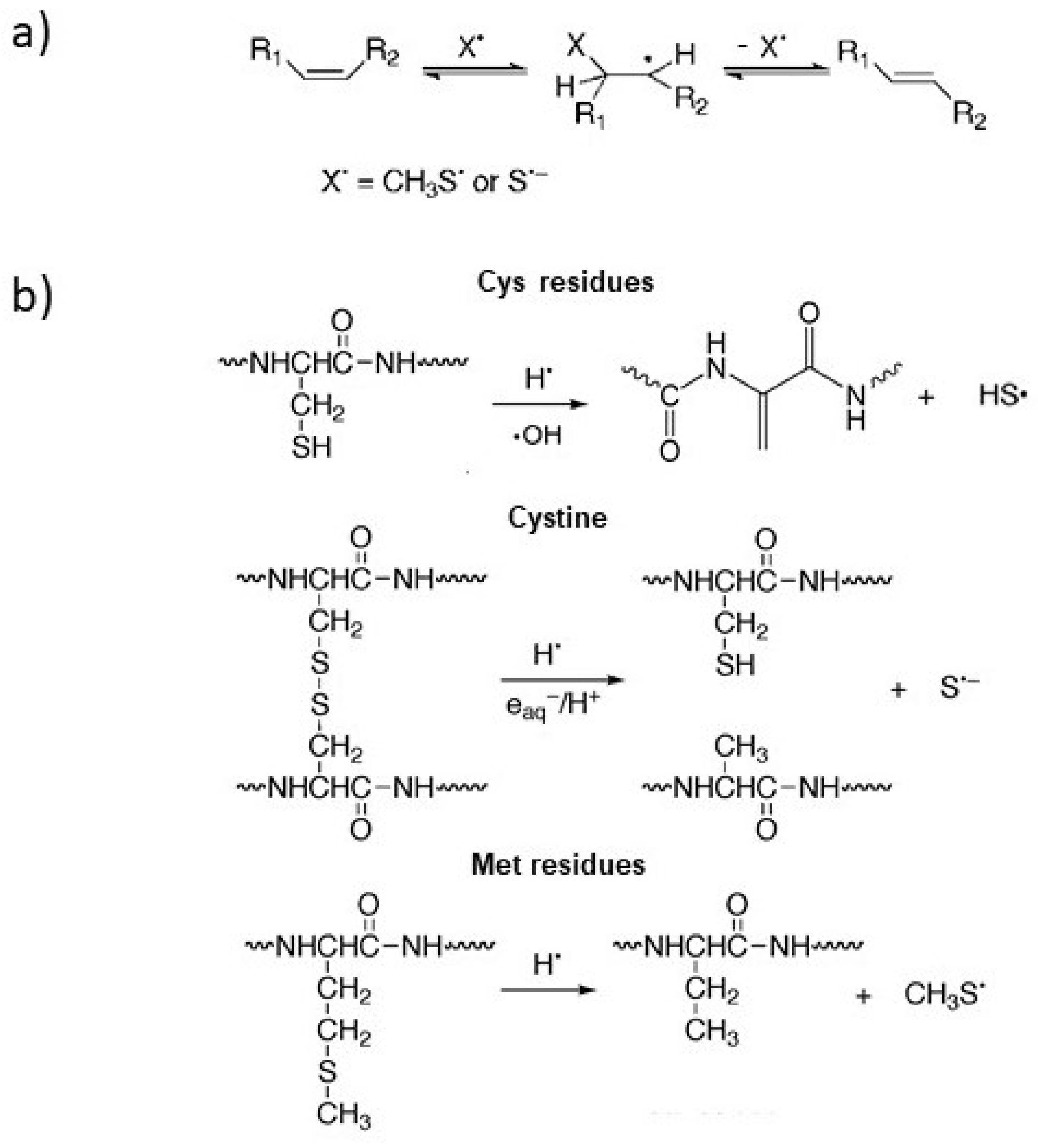
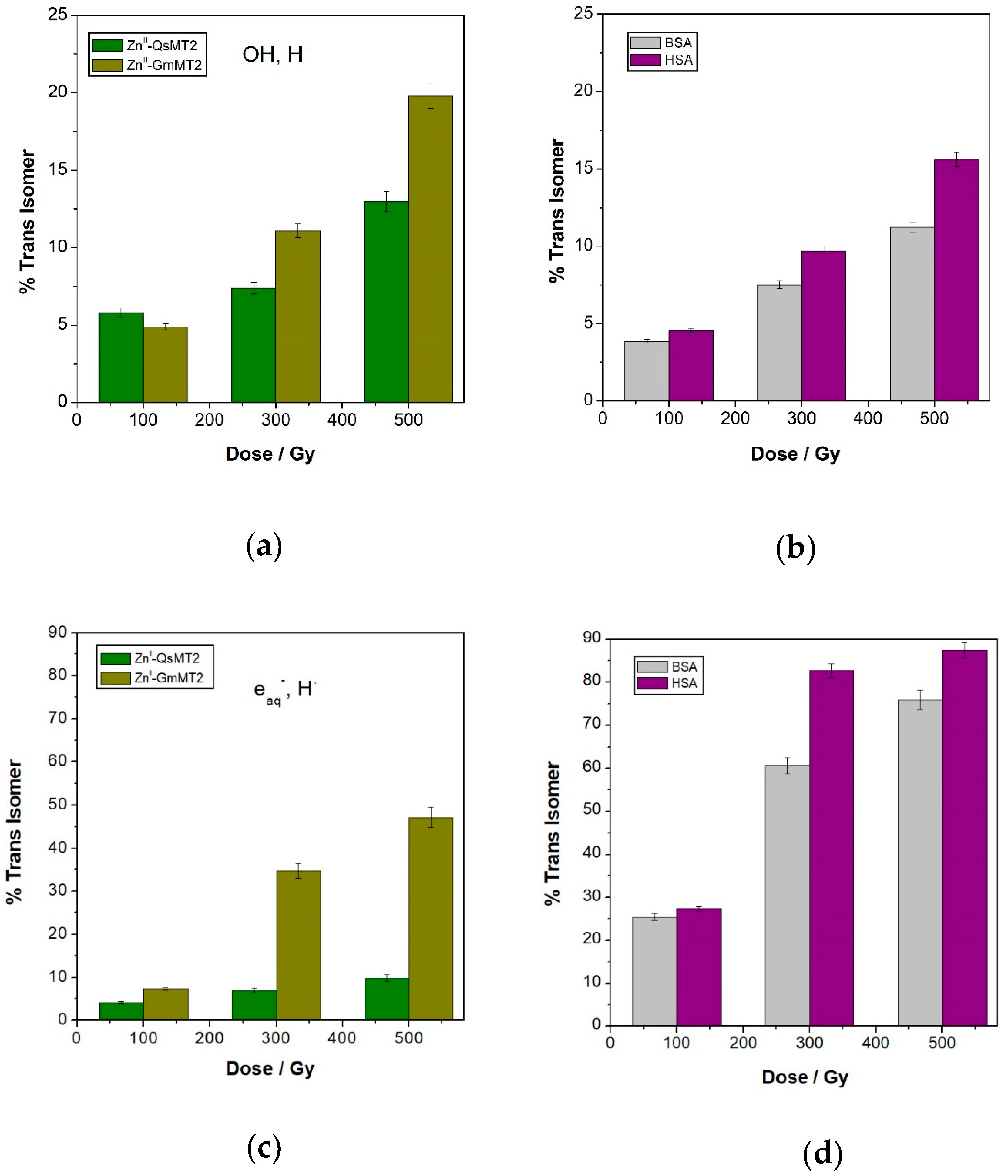
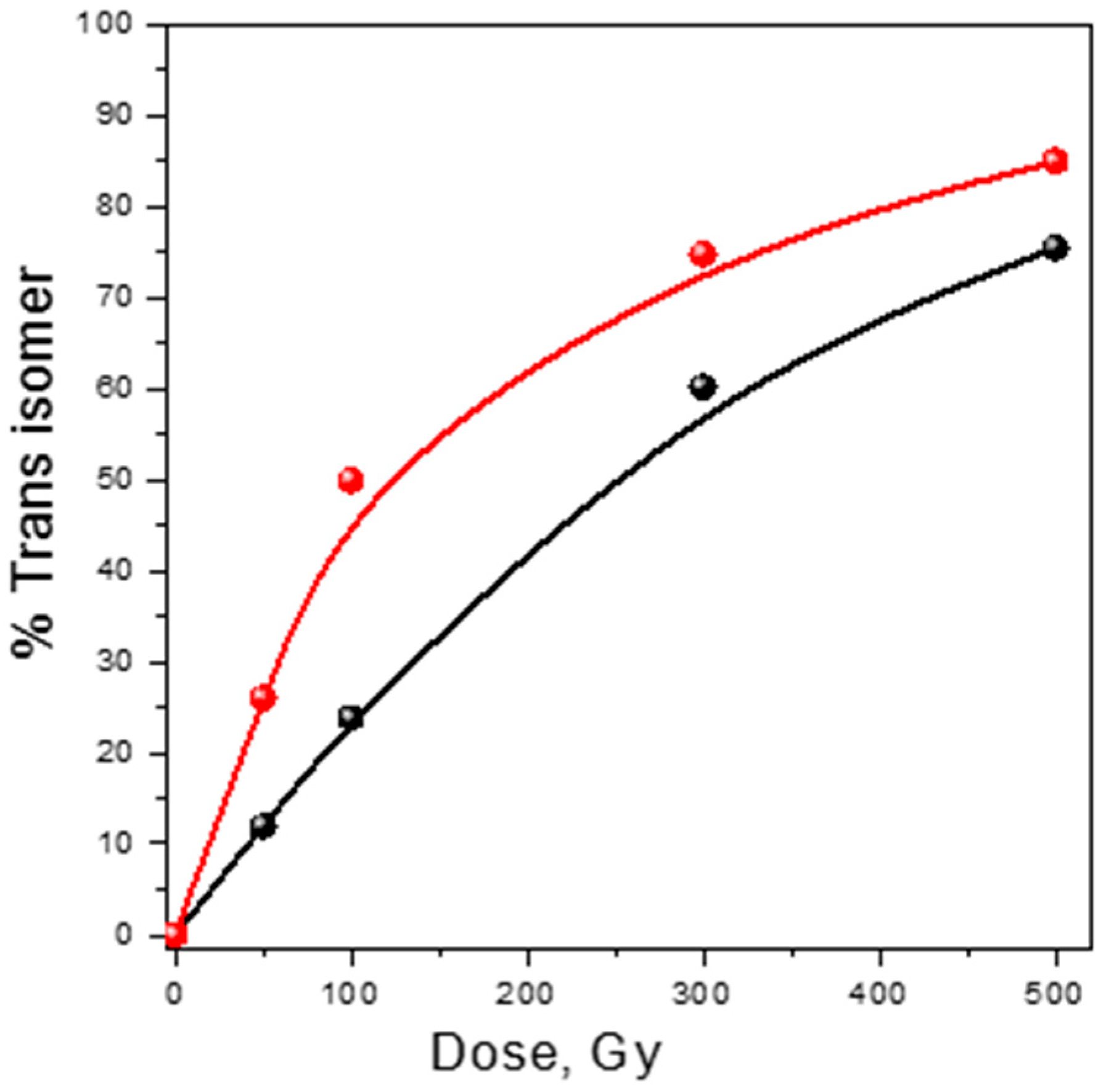
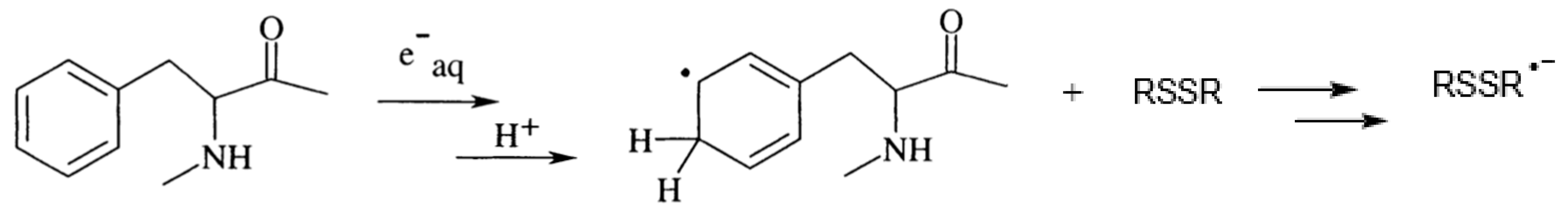
3.1. Damage Transfer from Protein to Lipid
3.2. Protein Analysis by Raman Spectroscopy
3.2.1. Protein Secondary Structure Change
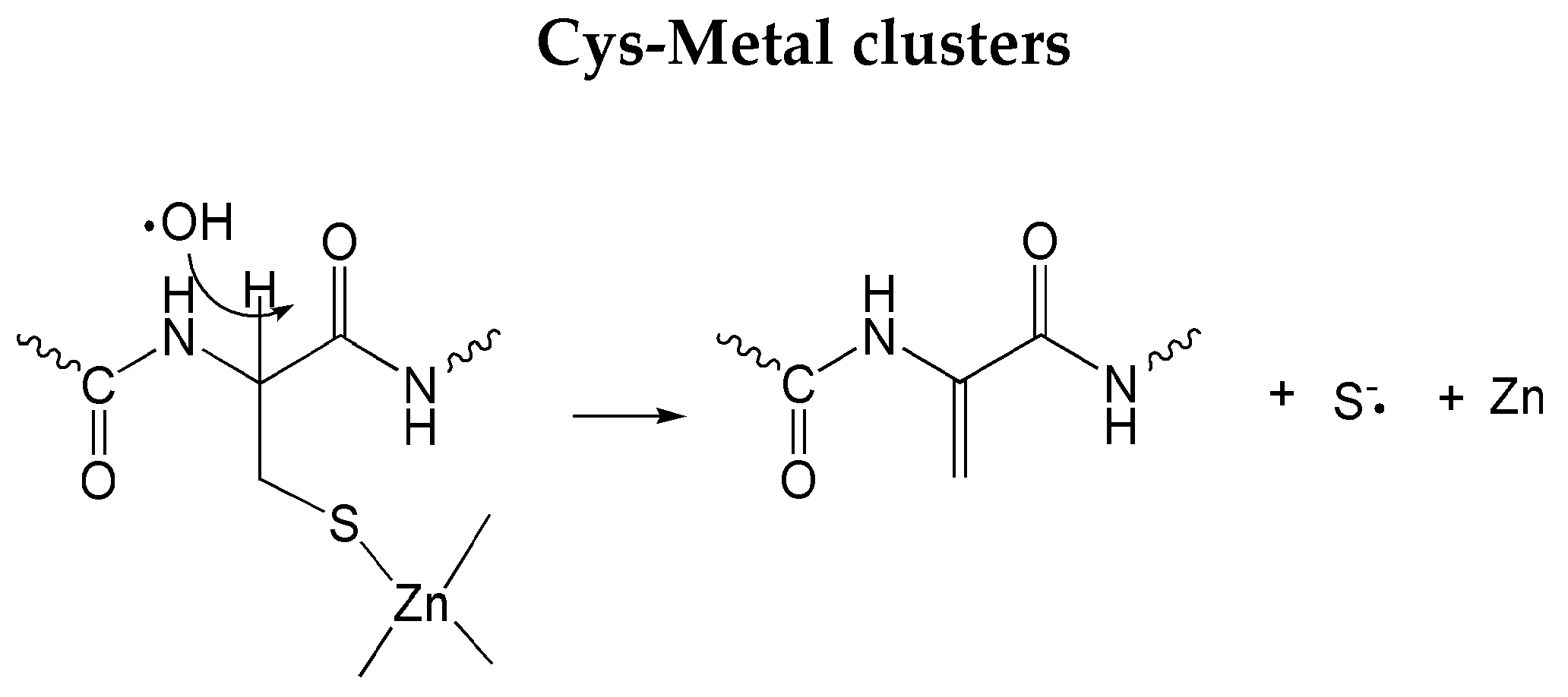
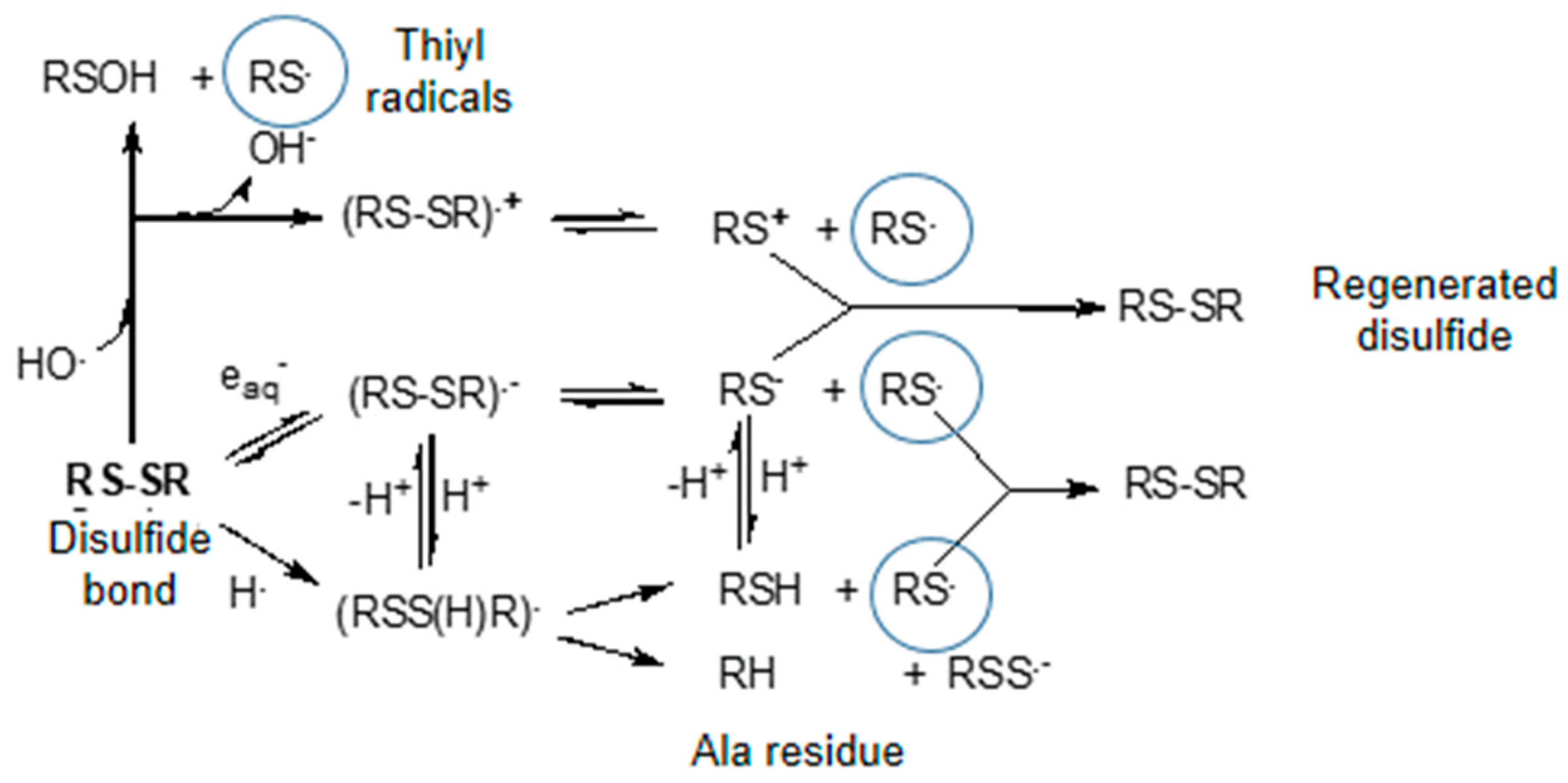
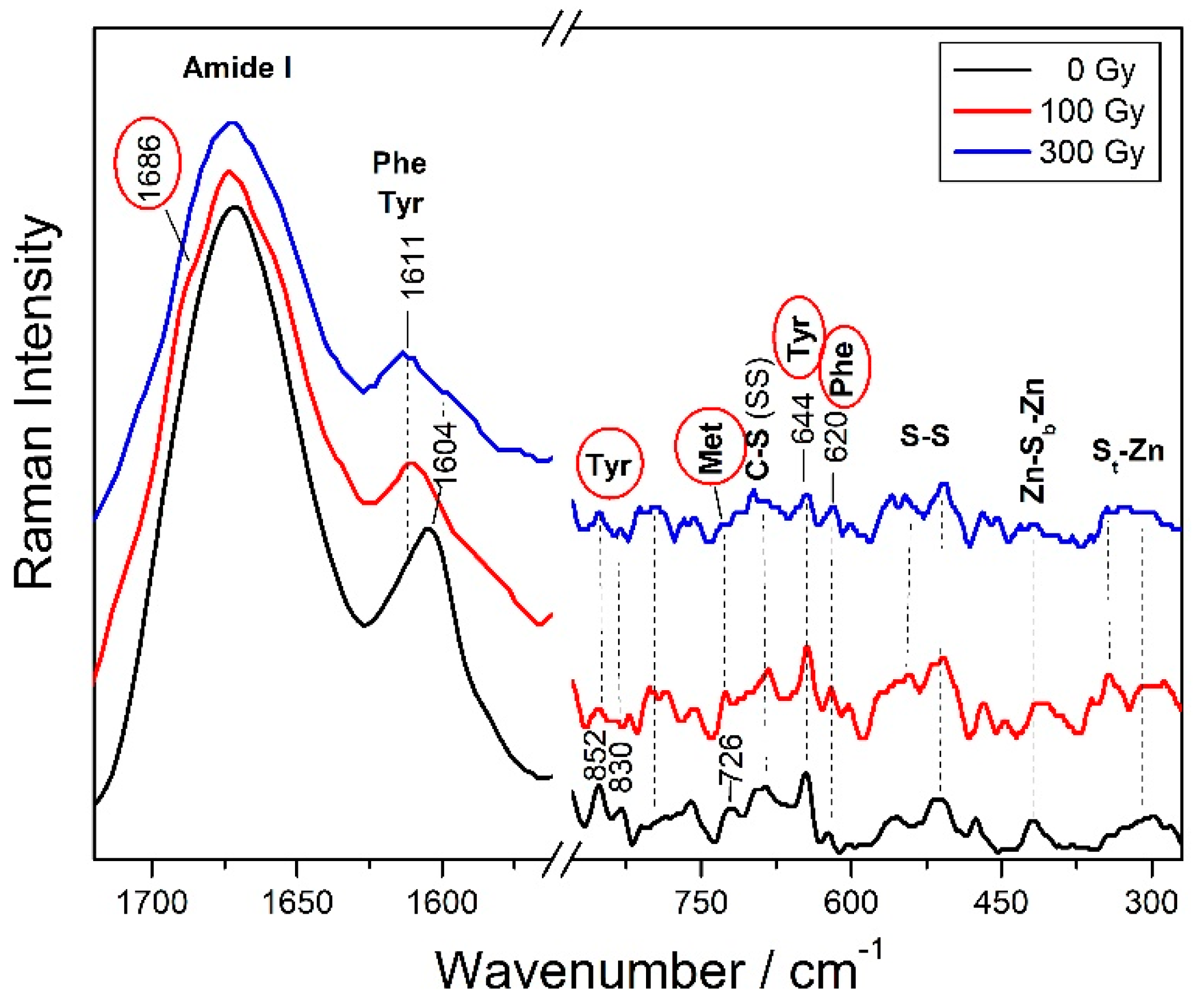
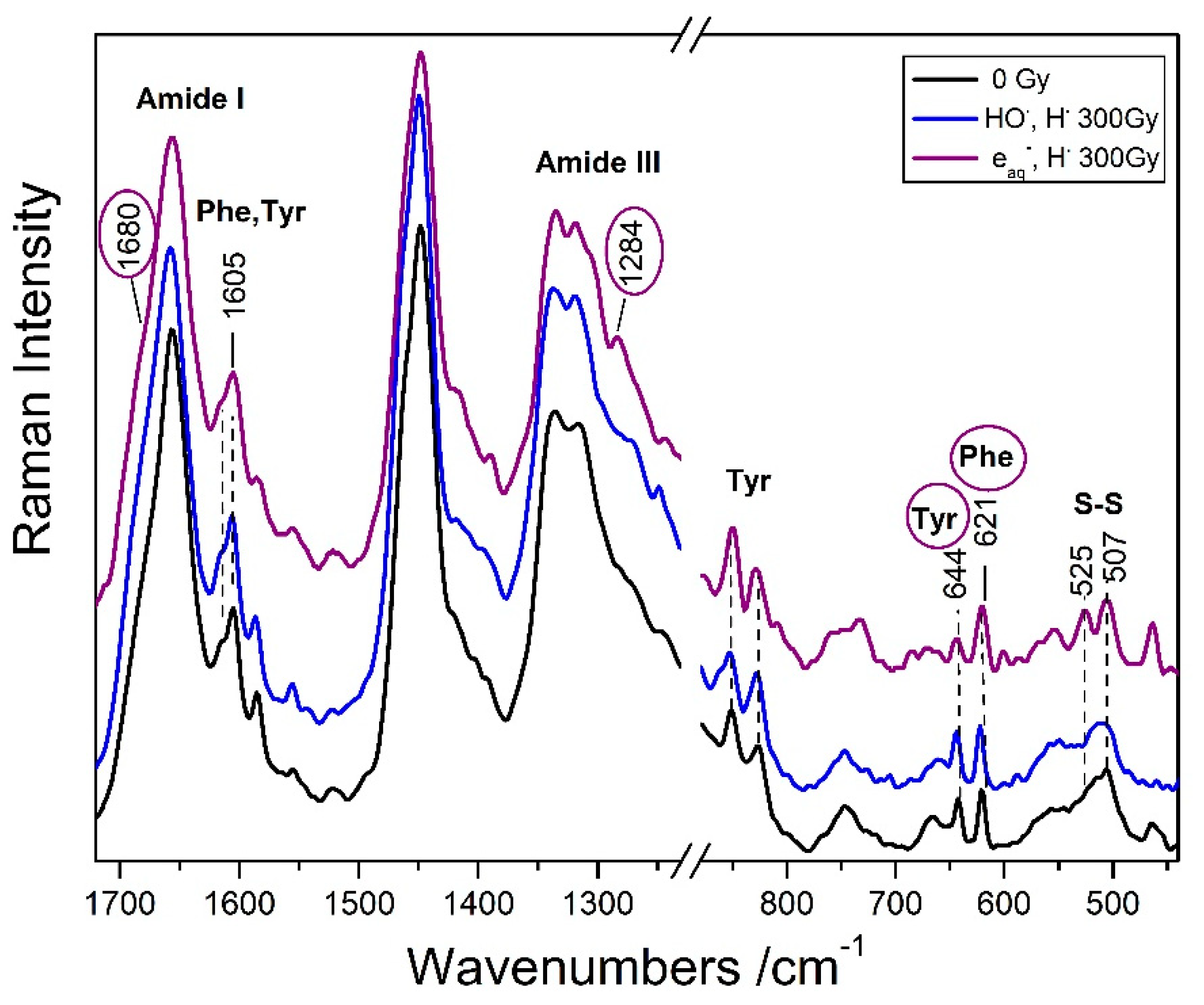
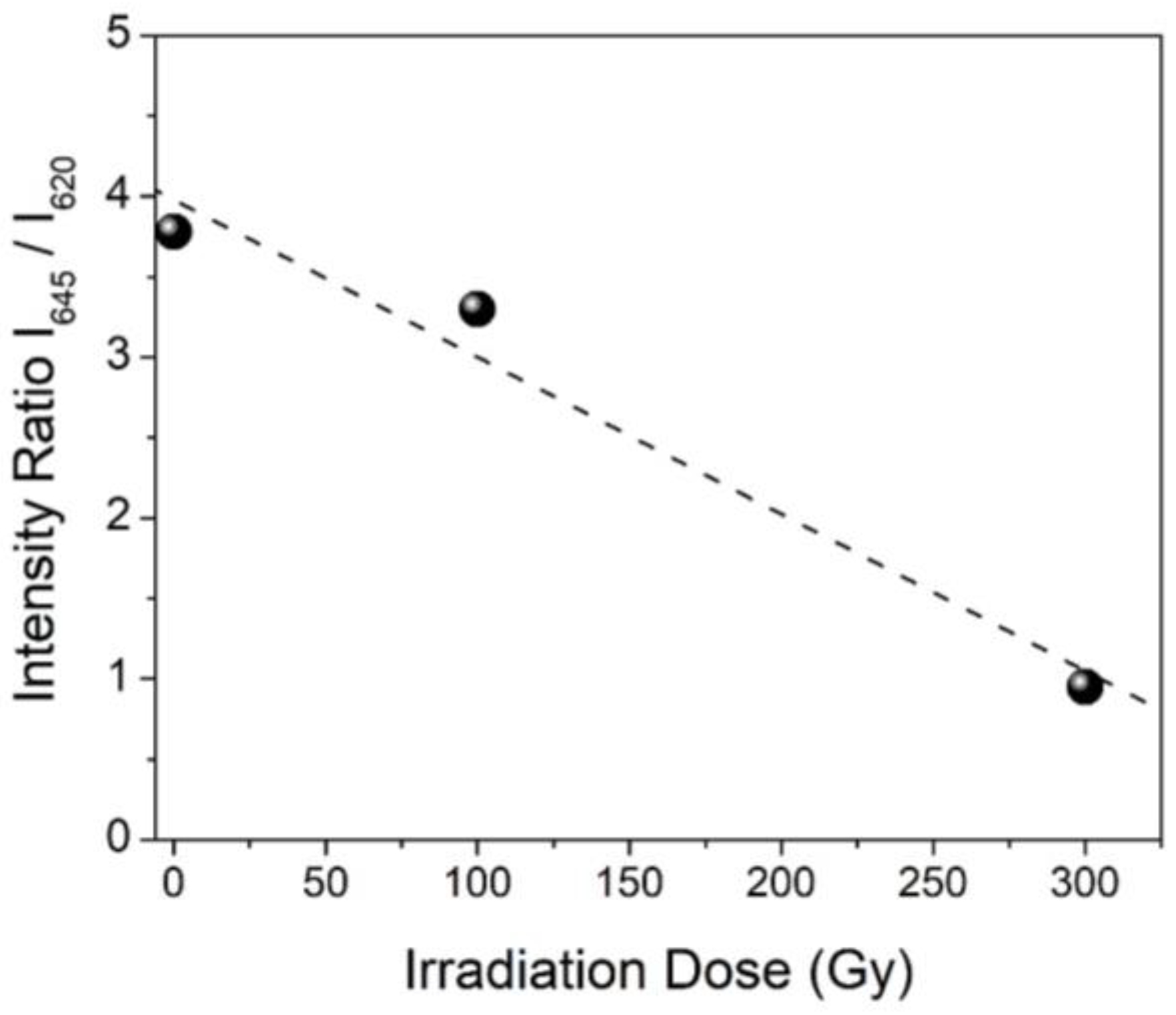
3.2.2. Specific Amino Acid Residues Damage
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heipieper, H.J.; Meinhardt, F.; Segyra, A. The cis-trans isomerase of unsaturated fatty acids in Pseudomonas and Vibrio: Biochemistry, molecular biology and physiological function of a unique stress adaptive mechanism. FEMS Microbiol. Lett. 2003, 229, 1–7. [Google Scholar] [CrossRef]
- Wolff, R.L.; Entressangles, B. Steady-state fluorescence polarization study of structurally defined phospholipids from liver mitochondria of rats fed elaidic acid. Biochem. Biophys. Acta 1994, 1211, 198–206. [Google Scholar] [CrossRef]
- Murakami, Y.; Tsuyama, M.; Kobayashi, Y.; Kodama, H.; Iba, K. Trienoic fatty acids and plant tolerance of high temperature. Science 2000, 287, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake, W.M.N.; Chen, Z.-Y. Trans, ω-3, and ω-6 fatty acids in Canadian human milk. Lipids 1996, 31, S279–S282. [Google Scholar] [CrossRef]
- Wardman, P. Thiyl radicals in biology: Their role as a ‘molecular switch’ central to cellular oxidative stress. In S-Centered Radicals; Alfassi, Z.B., Ed.; Wiley: Chichester, Great Britain, UK, 1999; pp. 289–309. [Google Scholar]
- Sies, H. Glutathione and its role in cellular functions. Free-Radic. Biol. Med. 1999, 27, 916–921. [Google Scholar] [CrossRef]
- Mozziconacci, O.; Williams, T.D.; Kerwin, B.A.; Schoneich, C. Intramolecular hydrogen transfer between protein cysteine thiyl radicals and αC−H bonds in insulin: Control of selectivity by secondary structure. J. Phys. Chem. B 2008, 112, 15921–15932. [Google Scholar] [CrossRef]
- Douki, T.; Odin, F.; Caillat, S.; Favier, A.; Cadet, J. Predominance of the 1,N2-propano′-deoxyguanosine adduct among 4-hydroxy -2-nonenal-induced DNA lesions. Free Radic. Biol. Med. 2004, 37, 62–70. [Google Scholar] [CrossRef]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Lykakis, I.N.; Ferreri, C.; Chatgilialoglu, C. The sulfhydryl radical (HS(.)/S(.-)): A contender for the isomerization of double bonds in membrane lipids. Angew. Chem. Int. Ed. 2007, 46, 1914–1916. [Google Scholar] [CrossRef]
- Chatgilialoglu, C.; Ferreri, C.; Torreggiani, A.; Renzone, G.; Salzano, A.M.; Scaloni, A. Radiation-induced reductive modifications of sulfur-containing amino acids within peptides and proteins. J. Proteom. 2011, 74, 2264–2273. [Google Scholar] [CrossRef]
- Chatgilialoglu, C.; Ferreri, C.; Melchiorre, M.; Sansone, A.; Torreggiani, A. Lipid Geometrical isomerism: From Chemistry to Biology and Diagnostics. Chem. Rev. 2014, 114, 255–284. [Google Scholar] [CrossRef] [PubMed]
- Torreggiani, A.; Tamba, M.; Manco, I.; Faraone-Mennella, M.R.; Ferreri, C.; Chatgilialoglu, C. Radiation damage of lysozyme in a biomimetic model: Some insights by Raman spectroscopy. J. Mol. Struct. 2005, 744–747, 767–773. [Google Scholar] [CrossRef]
- Torreggiani, A.; Tamba, M.; Manco, I.; Faraone-Mennella, M.R.; Ferreri, C.; Chatgilialoglu, C. Investigation of radical-based damage of RNase A in aqueous solution and lipid vesicles. Biopolymers 2006, 81, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, C.; Manco, I.; Faraone-Mennella, M.R.; Torreggiani, A.; Tamba, M.; Manara, S.; Chatgilialoglu, C. The reaction of hydrogen atoms with methionine residues: A model of reductive radical stress-causing tandem protein-lipid damage. ChemBioChem 2006, 7, 1738–1744. [Google Scholar] [CrossRef] [PubMed]
- Mee, L.K.; Adelstein, S.J.; Steinhart, C.M.; Lichtin, N.N. Reactions of aqueous L-methionine, L-phenylalanine, L-methionyl-L-phenylalanine, L-phenylalanyl-L-methionine and their mixtures with H atoms during steady radiolysis at pH 6.5. Radiat. Res. 1977, 71, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Davies, M.J. Generation and propagation of radical reactions on proteins. Biochim. Biophys. Acta 2001, 1504, 196–219. [Google Scholar] [CrossRef]
- Garrison, W.M. Reaction mechanisms in the radiolysis of peptides, polypeptides, and proteins. Chem. Rev. 1987, 87, 381–398. [Google Scholar] [CrossRef]
- Zhang, X.; Min, X.; Li, C.; Benjamin, I.J.; Qian, B.; Zhang, X.; Ding, Z.; Gao, X.; Yao, Y.; Ma, Y.; et al. Involvement of reductive stress in the cardiomyopathy in transgenic mice with cardiac-specific overexpression of heat shock Protein 27. Hypertension 2010, 55, 1412–1417. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta–Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Wadley, A.J.; Holliday, A.; Morgan, R.G.; Heesom, K.J.; Aldred, S.; Peters, D.M.; Bueno, A.A.; Coles, S.J. Preliminary evidence of reductive stress in human cytotoxic T-cells following exercise. J. Appl. Physiol. 2018, 125, 586–595. [Google Scholar] [CrossRef]
- Majorek, K.A.; Porebski, P.J.; Dayal, A.; Zimmerman, M.D.; Jablonska, K.; Stewart, A.J.; Chruszcz, M.; Minor, W. Structural and immunologic characterization of bovine, horse, and rabbit serum albumins. Mol. Immunol. 2012, 52, 174–182. [Google Scholar] [CrossRef]
- Vašák, M.; Meloni, G. Mammalian Metallothionein-3: New Functional and Structural Insights. Int. J. Mol. Sci. 2017, 18, 1117. [Google Scholar]
- Capdevila, M.; Bofill, R.; Palacios, Ò.; Atrian, S. State-of-the-art of metallothioneins at the beginning of the 21st century. Coord. Chem. Rev. 2012, 256, 46–62. [Google Scholar] [CrossRef]
- Aravindakumar, C.T.; Ceulemans, J.; De Ley, M. Nitric oxide induces Zn2+ release from metallothionein by destroying zinc-sulphur clusters without concomitant formation of S-nitrosothiol. Biochem. J. 1999, 344, 253–258. [Google Scholar] [PubMed]
- Hartmann, H.J.; Weser, U. Copper-release from yeast Cu(I)-metallothionein by nitric oxide (NO). Biometals 2000, 13, 153–156. [Google Scholar] [CrossRef]
- Capdevila, M.; Domènech, J.; Pagani, A.; Tío, L.; Villarreal, L.; Atrian, S. Zn- and Cd-metallothionein recombinant species from the most diverse phyla may contain sulfide (S2−) ligands. Angew. Chem. Int. Ed. Engl. 2005, 44, 4618–4622. [Google Scholar] [CrossRef]
- Maret, W.; Vallee, B.L. Thiolate ligands in metallothionein confer redox activity on zinc clusters. Proc. Natl. Acad. Sci. USA 1998, 95, 3478–3482. [Google Scholar] [CrossRef]
- Domenech, J.; Tinti, A.; Capdevila, M.; Atrian, S.; Torreggiani, A. Structural study of the zinc and cadmium complexes of a type 2 plant (Quercus suber) metallothionein: Insights by vibrational spectroscopy. Biopolymers 2007, 86, 240–248. [Google Scholar] [CrossRef]
- Tomas, M.; Tinti, A.; Bofill, R.; Capdevila, M.; Atrian, S.; Torreggiani, A. Comparative Raman study of four plant metallothionein isoforms: Insights into their Zn(II) clusters and protein conformations. J. Inorg. Biochem. 2016, 156, 55–63. [Google Scholar] [CrossRef]
- Capdevila, M.; Cols, N.; Romero-Isart, N.; Gonzàlez-Duarte, R.; Atrian, S.; Gonzàlez-Duarte, P. Recombinant synthesis of mouse Zn3-ß and Zn4-α metallothionein 1 domains and characterization of their cadmium(II) binding capacity. Cell. Mol. Life Sci. 1997, 53, 681–688. [Google Scholar] [CrossRef]
- Cols, N.; Romero-Isart, N.; Capdevila, M.; Oliva, B.; Gonzàlez-Duarte, P.; Gonzàlez-Duarte, R.; Atrian, S. Binding of excess cadmium(II) to Cd7-metallothionein from recombinant mouse Zn7-metallothionein 1. UV-VIS absorption and circular dichroism studies and theoretical location approach by surface accessibility analysis. J. Inorg. Biochem. 1997, 68, 157–166. [Google Scholar] [CrossRef]
- Tu, A.T. Spectroscopy of Biological Systems; Clark, R.J.H., Hester, R.E., Eds.; Wiley: Chichester, UK, 1986; Volume 13, pp. 47–112. [Google Scholar]
- Vass, E.; Hollosi, M.; Besson, F.; Buchet, R. Vibrational spectroscopic detection of beta and gamma-turns in synthetic and natural peptides and proteins. Chem. Rev. 2003, 103, 1917–1954. [Google Scholar] [CrossRef]
- Torreggiani, A.; Tinti, A. Raman spectroscopy a promising technique for investigations of metallothioneins. Metallomics 2010, 2, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Jurasekova, Z.; Tinti, A.; Torreggiani, A. Use of Raman spectroscopy for the identification of radical-mediated damages in human serum albumin. Anal. Bioanal. Chem. 2011, 400, 2921–2931. [Google Scholar] [CrossRef]
- Kramer, A.C.; Torreggiani, A.; Davies, M.J. Effect of Oxidation and Protein Unfolding on Cross-Linking of beta-Lactoglobulin and alpha-Lactalbumin. J. Agric. Food Chem. 2017, 65, 10258–10269. [Google Scholar] [CrossRef]
- Buxton, G.V.; Greenstock, C.L.; Helman, W.P.; Ross, A.B. Critical-review of rate constants for reactions of hydrated electrons, hydrogen atoms and hydroxyl radicals (•OH/•O−) in aqueous-solution. J. Phys. Chem. Ref. Data 1988, 17, 513–886. [Google Scholar] [CrossRef] [Green Version]
- Spinks, J.W.T.; Woods, R.J. An Introduction to Radiation Chemistry, 3rd ed.; Wiley & Sons: New York, NY, USA, 1990; pp. 243–313. [Google Scholar]
- Winterbourn, C.C. Toxicity of iron and hydrogen peroxide: The Fenton reaction. Toxicol. Lett. 1995, 82/83, 969–974. [Google Scholar] [CrossRef]
- Steinberg, D.; Witztum, J. Is the oxidative modification hypothesis relevant to human atherosclerosis? Do the antioxidant trials conducted to date refute the hypothesis? Circulation 2002, 105, 2107–2111. [Google Scholar] [CrossRef] [Green Version]
- Kramer, J.F.K.; Fellner, V.; Dugan, M.E.R.; Sauer, F.D.; Mossoba, M.M.; Yurawecz, M.P. Evaluating acid and base catalysts in the methylation of milk and rumen fatty acids with special emphasis on conjugated dienes and total trans fatty acids. Lipids 1997, 32, 1219–1228. [Google Scholar] [CrossRef]
- Ferreri, C.; Samadi, A.; Sassatelli, F.; Landi, L.; Chatgilialoglu, C. Regioselective cis-trans isomerization of arachidonic double bonds by thiyl radicals: The influence of phospholipid supramolecular organization. J. Am. Chem. Soc. 2004, 126, 1063–1072. [Google Scholar] [CrossRef]
- Talamo, F.; D’Ambrosio, C.; Arena, S.; Del Vecchio, P.; Ledda, L.; Zehender, G.; Ferrara, L.; Scaloni, A. Proteins from bovine tissues and biological fluids: Defining a reference electrophoresis map for liver, kidney, muscle, plasma and red blood cells. Proteomics 2003, 3, 440–460. [Google Scholar] [CrossRef] [PubMed]
- Melchiorre, M.; Ferreri, C.; Tinti, A.; Chatgilialoglu, C.; Torreggiania, A. A Promising Raman Spectroscopy Technique for the Investigation of trans and cis Cholesteryl Ester Isomers in Biological Samples. Appl. Spectrosc. 2015, 69, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Mihaljević, B.; Tartaro, I.; Ferreri, C.; Chatgilialoglu, C. Linoleic acid peroxidation vs. isomerization: A biomimetic model of free radical reactivity in the presence of thiols. Org. Biomol. Chem. 2011, 9, 3541–3548. [Google Scholar]
- Chatgilialoglu, C.; Ferreri, C.; Ballestri, M.; Mulazzani, Q.G.; Landi, L. Cis-trans isomerization of monounsaturated fatty acid residues in phospholipids by thiyl radicals. J. Am. Chem. Soc. 2000, 122, 4593–4601. [Google Scholar] [CrossRef]
- Torreggiani, A.; Domènech, J.; Orihuela, R.; Ferreri, C.; Atrian, S.; Capdevila, M.; Chatgilialoglu, C. Zinc and cadmium complexes of a plant metallothionein under radical stress: Desulfurisation reactions associated with the formation of trans-lipids in model membranes. Chem. Eur. J. 2009, 15, 6015–6024. [Google Scholar] [CrossRef]
- Torreggiani, A.; Chatgilialoglu, C.; Ferreri, C.; Melchiorre, M.; Atrian, S.; Capdevila, M. Non-enzymatic modifications in metallothioneins connected to lipid membrane damages: Structural and biomimetic studies under reductive radical stress. J. Proteom. 2013, 92, 204–2015. [Google Scholar] [CrossRef]
- Salzano, M.; Renzone, G.; Scaloni, A.; Torreggiani, A.; Ferreri, C.; Chatgilialoglu, C. Human serum albumin modifications associated with reductive radical stress. Mol. BioSyst. 2011, 7, 889–898. [Google Scholar] [CrossRef]
- Tomas, M.; Jiménez-Martí, E.; Bofill, R.; Tinti, A.; Di Foggia, M.; Chatgilialoglu, C.; Torreggiani, A. Comparative analysis of the soybean metallothionein system under radical stress: Protein modification connected to lipid membrane damages. Metallomics 2018, 10, 1792–1804. [Google Scholar] [CrossRef]
- Tuma, R. Raman spectroscopy of proteins: From peptides to large assemblies. J. Raman Spectrosc. 2005, 36, 307–319. [Google Scholar] [CrossRef]
- Torreggiani, A.; Domenech, J.; Tinti, A. Structural modifications in metal complexes of a plant metallothionein caused by reductive radical stress: A Raman study. J. Raman Spectrosc. 2009, 40, 1687–1693. [Google Scholar] [CrossRef]
- Krimm, S.; Bandekar, J. Vibrational analysis of peptides, polypeptides, and proteins. V. Normal vibrations of beta-turns. Biopolymers 1980, 19, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Torreggiani, A. Structural lesion to ribonuclease A caused by reductive stress: Assessment by Raman spectroscopy. Spectroscopy 2008, 22, 279–286. [Google Scholar] [CrossRef]
- Mee, L.K.; Adelstein, S.J.; Stein, G. Inactivation of Ribonuclease by the Primary Aqueous Radicals. Rad. Res. 1972, 52, 588–602. [Google Scholar] [CrossRef]
- Davies, M.J.; Fu, S.; Wang, H.; Dean, R.T. Stable markers of oxidant damage to proteins and their application in the study of human disease. Free Radic. Biol. Med. 1999, 27, 1151–1163. [Google Scholar] [CrossRef]
- Liddle, W.K.; Tu, A.T. Evaluation of phenylalanine and tyrosine Raman lines for the amino acid ratio. Appl. Spectrosc. 1981, 35, 444–446. [Google Scholar] [CrossRef]
- Ploder, M.; Neurauter, G.; Spittler, A.; Schroecksnadel, K.; Roth, E.; Fuchs, D. Serum phenylalanine in patients post trauma and with sepsis correlate to neopterin concentrations. Amino Acids 2008, 35, 303–307. [Google Scholar] [CrossRef]
- Ferreri, C.; Angelini, F.; Chatgilialoglu, C.; Dellonte, S.; Moschese, V.; Rossi, P.; Chini, L. Trans fatty acids and atopic eczema/dermatitis syndrome: The relationship with a free radical cis-trans isomerization of membrane lipids. Lipids 2005, 40, 661–667. [Google Scholar] [CrossRef]
- Balazy, M.; Chemtob, S. Trans-arachidonic acids: New mediators of nitro-oxidative stress. Pharmacol. Ther. 2008, 119, 275–290. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Total AA | Cys | Met | Arom. AA |
|---|---|---|---|---|
| ZnII-QsMT2 | 81 | 14 | 3 | 2 Phe, 1 Hys |
| ZnII-GmMT2 | 77 | 14 | 4 | 2 Tyr, 1 Phe |
| BSA | 583 | 35 | 4 | 2 Trp, 20 Tyr, 27 Phe, 17 His |
| HSA | 585 | 35 | 6 | 1 Trp, 18 Tyr, 31 Phe, 14 His |
| Reactive Species | Dose (Gy) | Beta-Strands (%) | Beta-Turns (%) | Random (%) |
|---|---|---|---|---|
| - | 0 | 45 | 43 | 12 |
| HO• and H• | 100 | 25 | 55 | 19 |
| HO• and H• | 300 | 20 | 53 | 28 |
| eaq− and H• | 100 | 10 | 51 | 39 |
| Reactive Species | Alpha-Helix (%) | Beta-Sheets (%) | Beta-Turns (%) | Random (%) |
|---|---|---|---|---|
| - | 54 | 22 | 14 | 10 |
| HO• and H• | 43 | 32 | 12 | 13 |
| eaq− and H• | 46 | 23 | 22 | 9 |
| H• | 38 | 31 | 18 | 13 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torreggiani, A.; Tinti, A.; Jurasekova, Z.; Capdevila, M.; Saracino, M.; Foggia, M.D. Structural Lesions of Proteins Connected to Lipid Membrane Damages Caused by Radical Stress: Assessment by Biomimetic Systems and Raman Spectroscopy. Biomolecules 2019, 9, 794. https://doi.org/10.3390/biom9120794
Torreggiani A, Tinti A, Jurasekova Z, Capdevila M, Saracino M, Foggia MD. Structural Lesions of Proteins Connected to Lipid Membrane Damages Caused by Radical Stress: Assessment by Biomimetic Systems and Raman Spectroscopy. Biomolecules. 2019; 9(12):794. https://doi.org/10.3390/biom9120794
Chicago/Turabian StyleTorreggiani, Armida, Anna Tinti, Zuzana Jurasekova, Mercè Capdevila, Michela Saracino, and Michele Di Foggia. 2019. "Structural Lesions of Proteins Connected to Lipid Membrane Damages Caused by Radical Stress: Assessment by Biomimetic Systems and Raman Spectroscopy" Biomolecules 9, no. 12: 794. https://doi.org/10.3390/biom9120794