Betulinic Acid Attenuates T-2-Toxin-Induced Testis Oxidative Damage Through Regulation of the JAK2/STAT3 Signaling Pathway in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
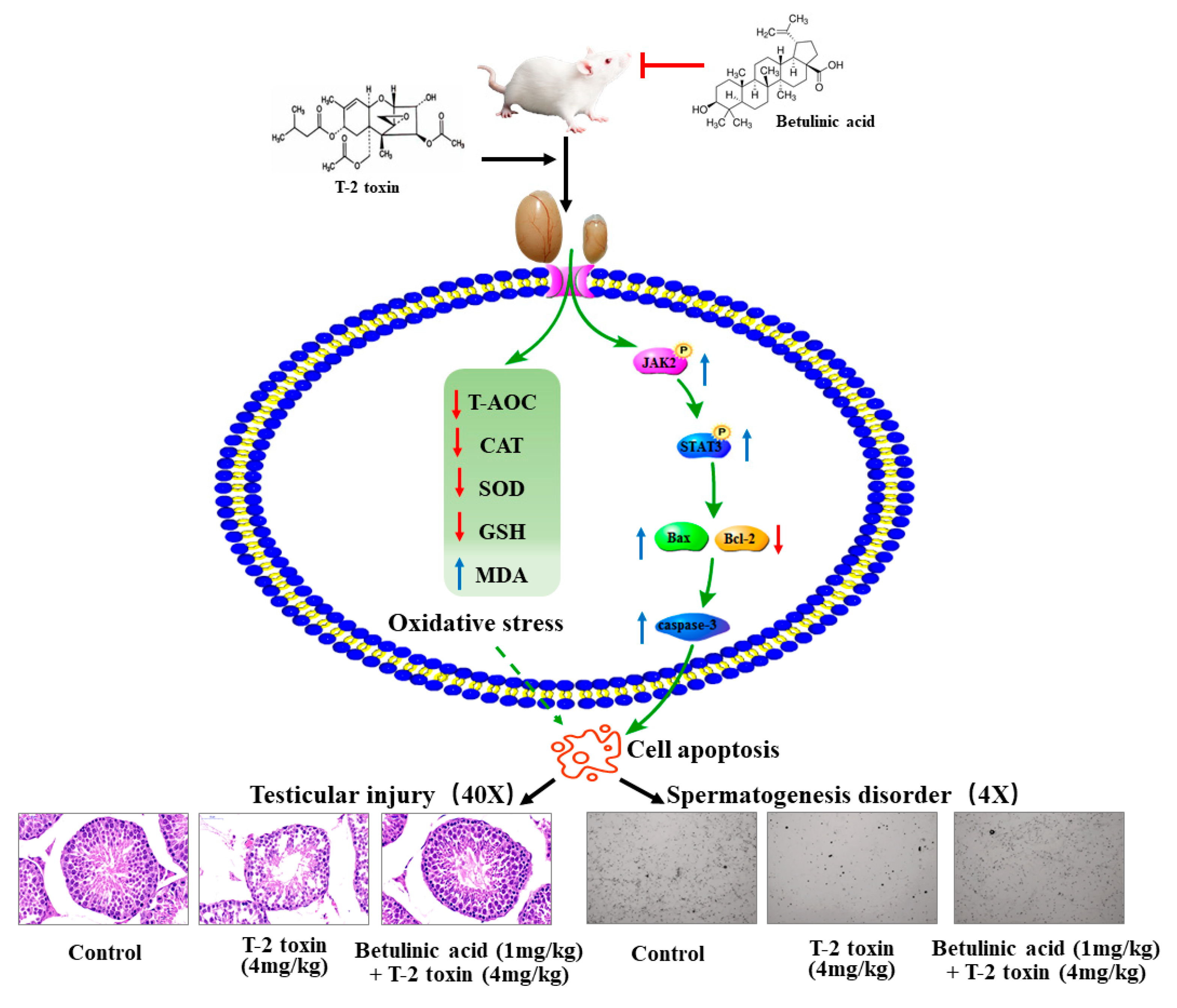
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Animals and Experimental Designs
2.3. Testosterone Enzyme-Linked Immunosorbent Assay
2.4. Analysis of Sperm Motility and Morphology
2.5. Histopathology of Testicular Tissue
2.6. Measurements of Antioxidative Capacity in Testes
2.7. Apoptosis Measured by Terminal Deoxynucleotidyl-Transferase-Mediated dUTP Nick End Labeling (TUNEL) Staining
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
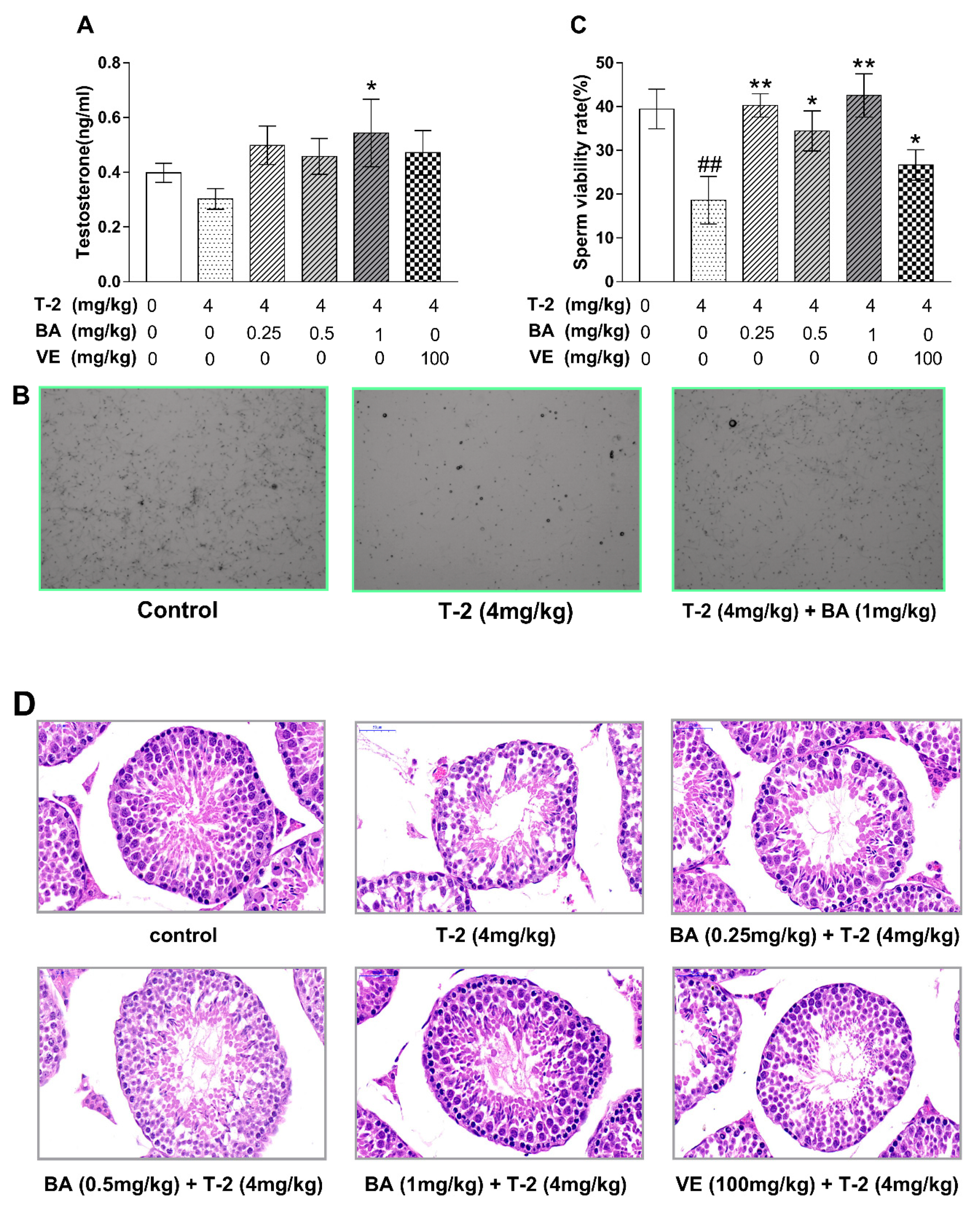
3.1. BA Repaired T-2-Toxin-Induced Testis Injury in Mice
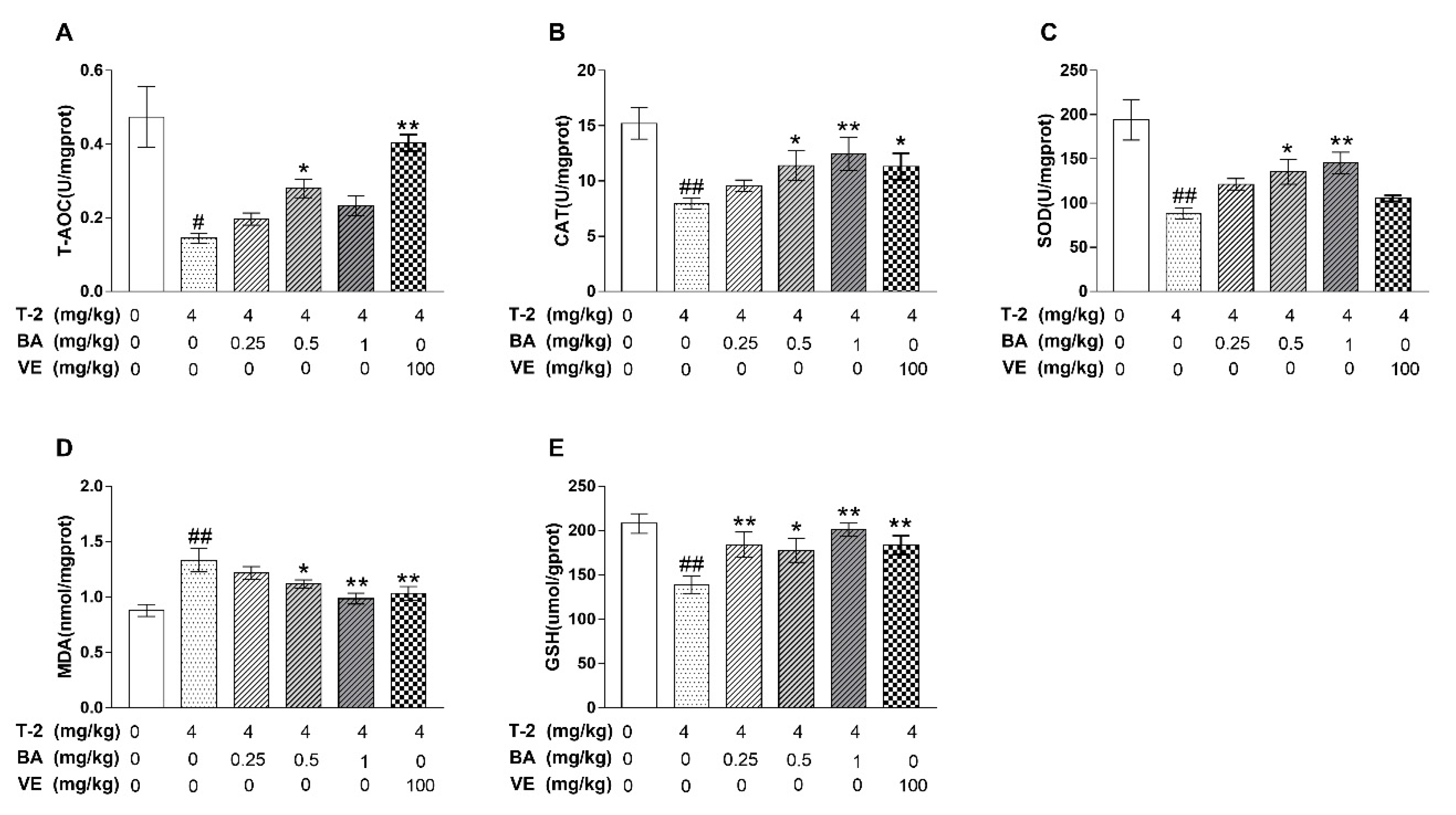
3.2. The Effect of BA on Testicular Antioxidant Capacity Induced by T-2 Toxin in Mice
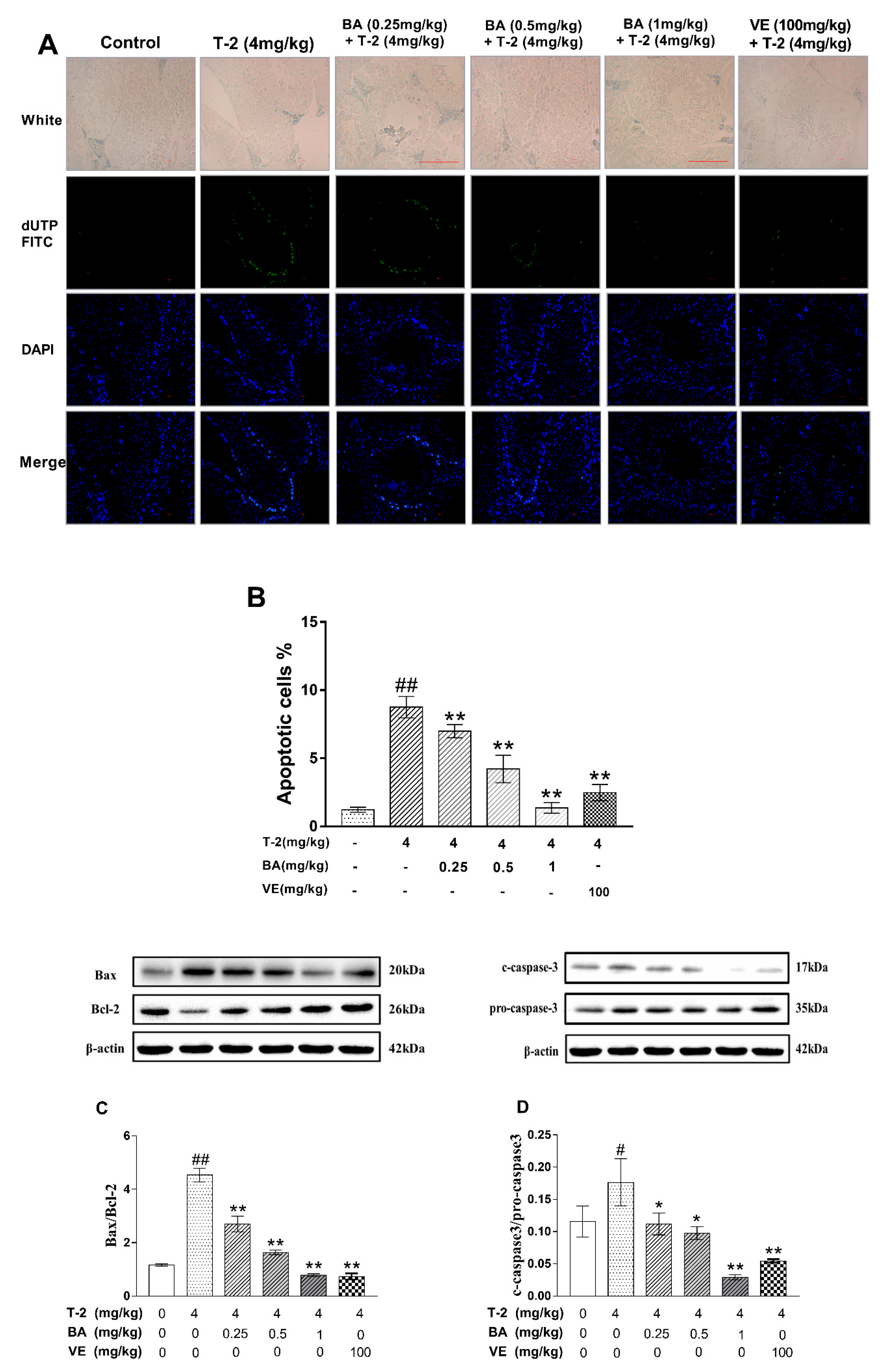
3.3. The Effect of BA on the Apoptosis of Testicular Cells and Apoptosis-Related Protein Expression Induced by T-2 Toxin in Mice
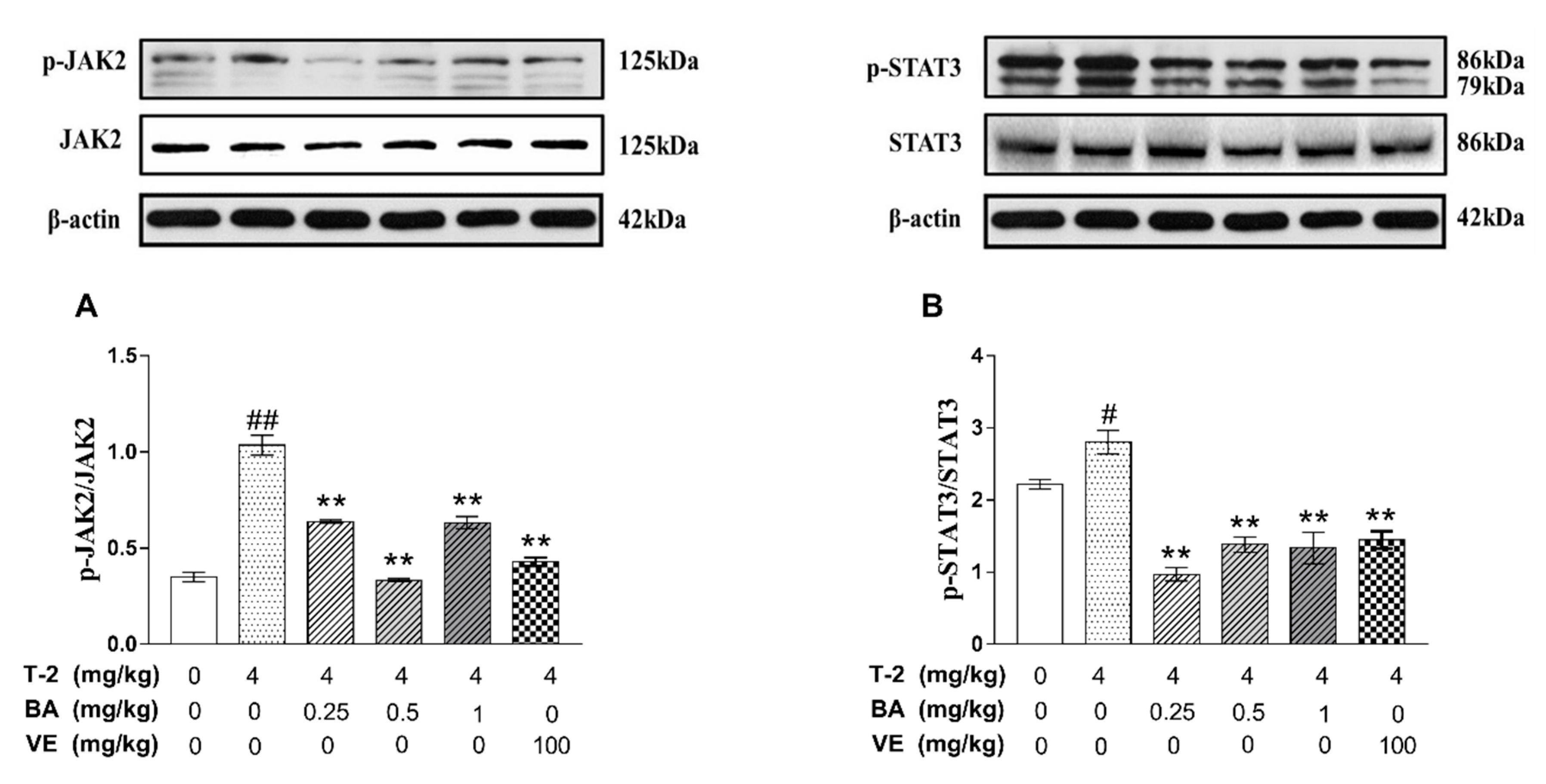
3.4. The Effects of BA on the Expression of Apoptosis-Related Proteins in the JAK2/STAT3 Signaling Pathway Induced by T-2 Toxin in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, Z.; Hu, X.; Zhang, Q.; Li, P. Determination for multiple mycotoxins in agricultural products using HPLC–MS/MS via a multiple antibody immunoaffinity column. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1021, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Li, S.; Bai, Y.; Prates, L.L.; Lei, Y.; Yu, P. Mycotoxin contamination of food and feed in China: Occurrence, detection techniques, toxicological effects and advances in mitigation technologies. Food Control. 2018, 91, 202–215. [Google Scholar] [CrossRef]
- Wang, H.J.; Xue, H.L.; Zhao, J.; Yang, B.I.; Lu-Mei, P.U.; Mao, X.R.; Wang, Y.; University, G.A. Advance on researches of production, toxicity and detoxification of T-2 toxin. J. Food Saf. Qual. 2014, 5, 2392–2398. [Google Scholar]
- Fatima, Z.; Guo, P.; Huang, D.; Lu, Q.; Wu, Q.; Dai, M.; Cheng, G.; Peng, D.; Tao, Y.; Ayub, M.; et al. The critical role of p16/Rb pathway in the inhibition of GH3 cell cycle induced by T-2 toxin. Toxicology 2018, 400–401, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Harkema, J.R.; Islam, Z.; Cuff, C.F.; Pestka, J.J. T-2 toxin impairs murine immune response to respiratory reovirus and exacerbates viral bronchiolitis. Toxicol. Appl. Pharmacol. 2006, 217, 76–85. [Google Scholar] [CrossRef]
- Yan, D.; Pei, F.; Song, Y. Serum levels of M-CSF, RANKL and OPG in rats fed with Kashin-Beck disease-affected diet. J. Orthop. Surg. Res. 2014, 9, 78. [Google Scholar] [CrossRef]
- Xu, J.; Pan, S.; Gan, F.; Hao, S.; Liu, D.; Xu, H.; Huang, K. Selenium deficiency aggravates T-2 toxin-induced injury of primary neonatal rat cardiomyocytes through ER stress. Chem. Biol. Interact. 2018, 285, 96–105. [Google Scholar] [CrossRef]
- Cristina, C.; Fabiola, P.; Spicer, L.J.; Francesca, C. Fusarium mycotoxins: Effects on reproductive function in domestic animals--A review. Theriogenology 2013, 80, 557–564. [Google Scholar]
- Wu, J.; Tu, D.; Yuan, L.Y.; Yuan, H.; Wen, L.X. T-2 toxin exposure induces apoptosis in rat ovarian granulosa cells through oxidative stress. Environ. Toxicol. Pharmacol. 2013, 36, 493–500. [Google Scholar] [CrossRef]
- Jing, W.; Li, J.; Hui, Y.; Shuang-Qing, P. T-2 toxin induces apoptosis in ovarian granulosa cells of rats through reactive oxygen species-mediated mitochondrial pathway. Toxicol. Lett. 2011, 202, 168–177. [Google Scholar]
- Jing, W.; Di, T.; Li-Yun, Y.; Jin-E, Y.; Yanan, T. T-2 toxin regulates steroid hormone secretion of rat ovarian granulosa cells through cAMP-PKA pathway. Toxicol. Lett. 2015, 232, 573–579. [Google Scholar]
- Yuan, Z.; Matias, F.B.; Yi, J.E.; Jing, W. T-2 toxin-induced cytotoxicity and damage on TM3 Leydig cells. Comp. Biochem. Physiol. C Toxicol. Pharm. 2015, 181–182, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Manjari, C.; Jayaraj, R.; Bhaskar, A.S.B.; Rao, P.V. Lakshmana. Oxidative stress induction by T-2 toxin causes DNA damage and triggers apoptosis via caspase pathway in human cervical cancer cells. Toxicology 2009, 262, 153–161. [Google Scholar]
- Rao, F.Z.; Tian, H.; Li, W.Q.; Hung, H.L.; Sun, F. Potential role of punicalagin against oxidative stress induced testicular damage. Asian J. Androl. 2016, 18, 627–632. [Google Scholar] [CrossRef] [PubMed]
- KaracaoäŸLu, E.; SelmanoäŸLu, G. T-2 toxin induces cytotoxicity and disrupts tight junction barrier in SerW3 cells. Env. Toxicol. Pharm. 2017, 56, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Adeleke, G.E.; Adaramoye, O.A. Betulinic acid protects against N-nitrosodimethylamine-induced redox imbalance in testes of rats. Redox Rep. Commun. Free Radic. Res. 2017, 22, 1. [Google Scholar]
- Ríos, J.L.; Máñez, S. New Pharmacological Opportunities for Betulinic Acid. Planta Med. 2018, 84, 8–19. [Google Scholar] [CrossRef]
- Agnieszka, S.C.; Krzysztof, P.; Jadwiga, D.; Martyna, K.S. Betulin and betulinic acid attenuate ethanol-induced liver stellate cell activation by inhibiting reactive oxygen species (ROS), cytokine (TNF-α, TGF-β) production and by influencing intracellular signaling. Toxicology 2011, 280, 152–163. [Google Scholar]
- Wang, M.; Zhu, P.; Jiang, C.; Ma, L.; Zhang, Z.; Zenga, X. Preliminary characterization, antioxidant activity in vitro and hepatoprotective effect on acute alcohol-induced liver injury in mice of polysaccharides from the peduncles of Hovenia dulcis. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2012, 50, 2964–2970. [Google Scholar] [CrossRef]
- Zhan-Wei, Z.; Shun-Zong, S.; Yan-Ling, W.; Li-Hua, L.; Ying, W.; Ji-Xing, N. Betulinic acid prevention of d-galactosamine/lipopolysaccharide liver toxicity is triggered by activation of Bcl-2 and antioxidant mechanisms. J. Pharm. Pharmacol. 2011, 63, 572–578. [Google Scholar]
- Yi, J.; Zhu, R.; Wu, J.; Wu, J.; Tan, Z. Ameliorative effect of betulinic acid on oxidative damage and apoptosis in the splenocytes of dexamethasone treated mice. Int. Immunopharmacol. 2015, 27, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Zhu, R.; Wu, J.; Wu, J.; Xia, W.; Zhu, L.; Jiang, W.; Xiang, S.; Tan, Z. In vivo protective effect of betulinic acid on dexamethasone induced thymocyte apoptosis by reducing oxidative stress. Pharmacol. Rep. Pr. 2016, 68, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Xia, W.; Wu, J.; Yuan, L.; Wu, J.; Tu, D.; Fang, J.; Tan, Z. Betulinic acid prevents alcohol-induced liver damage by improving the antioxidant system in mice. J. Vet. Sci. 2014, 15, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Andreas, J.; Aleksander, G.; Herman, T.; Thorsten, D.; Zsolt, K.; Gert, D.; Csilla, K. European Association of Urology guidelines on Male Infertility: The 2012 update. Eur. Urol. 2012, 62, 324–332. [Google Scholar]
- Haider, S.G. Cell Biology of Leydig Cells in the Testis. Int. Rev. Cytol. A Surv. Cell Biol. 2004, 233, 181–241. [Google Scholar]
- Azad, F.; Nejati, V.; Shalizar-Jalali, A.; Najafi, G.; Rahmani, F. Antioxidant and anti-apoptotic effects of royal jelly against nicotine-induced testicular injury in mice. Env. Toxicol. 2019, 34, 708–718. [Google Scholar] [CrossRef]
- Mäkelä, J.A.; Toppari, J. Spermatogenesis. In Endocrinology of the Testis and Male Reproduction; Simoni, M., Huhtaniemi, I., Eds.; Springer: Cham, Switzerland, 2017; pp. 417–455. [Google Scholar]
- Ren, Y.P.; Shao, W.H.; Zuo, L.J.; Zhao, W.; Qin, H.Z.; Hua, Y.J.; Lu, D.J.; Mi, C.; Zeng, S.E.; Zu, L. Mechanism of cadmium poisoning on testicular injury in mice. Oncol. Lett. 2019, 18, 1035–1042. [Google Scholar] [CrossRef]
- Gandhi, J.; Hernandez, R.J.; Chen, A.; Smith, N.L.; Sheynkin, Y.R.; Joshi, G.; Khan, S.A. Impaired hypothalamic-pituitary-testicular axis activity, spermatogenesis, and sperm function promote infertility in males with lead poisoning. Zygote 2017, 25, 103–110. [Google Scholar] [CrossRef]
- Yang, J.Y.; Zhang, Y.F.; Li, Y.X.; Meng, X.P.; Bao, J.F. l-arginine protects against oxidative damage induced by T-2 toxin in mouse Leydig cells. J. Biochem. Mol. Toxicol. 2018, 32, e22209. [Google Scholar] [CrossRef]
- Ammar, O.; Houas, Z.; Mehdi, M. The association between iron, calcium, and oxidative stress in seminal plasma and sperm quality. Env. Sci. Pollut. Res. Int. 2019, 26, 14097–14105. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, X.; Zhang, J.; Ji, Q.; Huang, W.; Zhang, X.; Li, Y. Spermatogenesis disorder caused by T-2 toxin is associated with germ cell apoptosis mediated by oxidative stress. Env. Pollut. 2019, 251, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Bhadauria, A.S.; Singh, A.K.; Saha, S. Betulinic acid as apoptosis activator: Molecular mechanisms, mathematical modeling and chemical modifications. Life Sci. 2018, 209, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.Q.; Zhang, T.; Zhegn, J.; Ding, W.M.; Wang, Y.; Li, Y.; Zhu, L.; Murray, M.; Zhou, F.F. Betulinic acid derivatives can protect human Muller cells from glutamate-induced oxidative stress. Exp. Cell Res. 2019, 383. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Liu, H.; Cheng, Y.T.; Yan, L.X.; Krittanawong, C.; Li, S.H.; Qian, W.; Su, W.; Chen, X.Z.; Hou, X.J.; et al. 2,3,5,4 ‘-Tetrahydroxystilbene-2-O-beta-d-glucoside eliminates ischemia/reperfusion injury-induced H9c2 cardiomyocytes apoptosis involving in Bcl-2, Bax, caspsae-3, and Akt activation. J. Cell. Biochem. 2019, 120, 10972–10977. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhou, Y.; Yuan, Z.H.; Yi, J.E.; Chen, J.S.; Wang, N.D.; Tian, Y.A. Autophagy and Apoptosis Interact to Modulate T-2 Toxin-Induced Toxicity in Liver Cells. Toxins 2019, 11, 45. [Google Scholar] [CrossRef]
- Lu, Z.; Lu, C.; Li, C.; Jiao, Y.; Li, Y.; Zhang, G. Dracorhodin perchlorate induces apoptosis and G2/M cell cycle arrest in human esophageal squamous cell carcinoma through inhibition of the JAK2/STAT3 and AKT/FOXO3a pathways. Mol. Med. Rep. 2019, 20, 2091–2100. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, P.; Guo, X.; Liu, T.; Luo, X.; Zhu, Y.T. Inhibition of JAK2/STAT3 signaling pathway protects mice from the DDP-induced acute kidney injury in lung cancer. Inflamm. Res. 2019, 68, 751–760. [Google Scholar] [CrossRef]
- Wu, L.; Li, J.; Liu, T.; Li, S.; Feng, J.; Yu, Q.; Zhang, J.; Chen, J.; Zhou, Y.; Ji, J.; et al. Quercetin shows anti-tumor effect in hepatocellular carcinoma LM3 cells by abrogating JAK2/STAT3 signaling pathway. Cancer Med. 2019. [Google Scholar] [CrossRef]
- He, P.; Zhang, D.; Li, H.; Yang, X.; Li, D.; Zhai, Y.; Ma, L.; Feng, G. Hepatitis B virus X protein modulates apoptosis in human renal proximal tubular epithelial cells by activating the JAK2/STAT3 signaling pathway. Int. J. Mol. Med. 2013, 31, 1017–1029. [Google Scholar] [CrossRef]
- Xu, T.; Pang, Q.; Wang, Y.; Yan, X. Betulinic acid induces apoptosis by regulating PI3K/Akt signaling and mitochondrial pathways in human cervical cancer cells. Int. J. Mol. Med. 2017, 40, 1669–1678. [Google Scholar] [CrossRef]
- Cai, Y.; Zheng, Y.; Gu, J.; Wang, S.; Wang, N.; Yang, B.; Zhang, F.; Wang, D.; Fu, W.; Wang, Z. Betulinic acid chemosensitizes breast cancer by triggering ER stress-mediated apoptosis by directly targeting GRP78. Cell Death Dis. 2018, 9, 636. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Gao, Y.Q.; Dai, W.B.; Hu, Y.; Wu, Y.F.; Mei, Q.X. Helicteric Acid, Oleanic Acid, and Betulinic Acid, Three Triterpenes from Helicteres angustifolia L., Inhibit Proliferation and Induce Apoptosis in HT-29 Colorectal Cancer Cells via Suppressing NF-κB and STAT3 Signaling. Evid. Based Complement. Altern. Med. 2017, 2017, 1–8. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Yang, C.; Liu, J.; Chen, J.; Huang, C.; Wang, J.; Liang, Z.; Wen, L.; Yi, J.-e.; Yuan, Z. Betulinic Acid Attenuates T-2-Toxin-Induced Testis Oxidative Damage Through Regulation of the JAK2/STAT3 Signaling Pathway in Mice. Biomolecules 2019, 9, 787. https://doi.org/10.3390/biom9120787
Wu J, Yang C, Liu J, Chen J, Huang C, Wang J, Liang Z, Wen L, Yi J-e, Yuan Z. Betulinic Acid Attenuates T-2-Toxin-Induced Testis Oxidative Damage Through Regulation of the JAK2/STAT3 Signaling Pathway in Mice. Biomolecules. 2019; 9(12):787. https://doi.org/10.3390/biom9120787
Chicago/Turabian StyleWu, Jing, Chenglin Yang, Juan Liu, Jiaxin Chen, Chao Huang, Ji Wang, Zengenni Liang, Lixin Wen, Jin-e Yi, and Zhihang Yuan. 2019. "Betulinic Acid Attenuates T-2-Toxin-Induced Testis Oxidative Damage Through Regulation of the JAK2/STAT3 Signaling Pathway in Mice" Biomolecules 9, no. 12: 787. https://doi.org/10.3390/biom9120787
APA StyleWu, J., Yang, C., Liu, J., Chen, J., Huang, C., Wang, J., Liang, Z., Wen, L., Yi, J.-e., & Yuan, Z. (2019). Betulinic Acid Attenuates T-2-Toxin-Induced Testis Oxidative Damage Through Regulation of the JAK2/STAT3 Signaling Pathway in Mice. Biomolecules, 9(12), 787. https://doi.org/10.3390/biom9120787