The Antifungal Effect of Garlic Essential Oil on Phytophthora nicotianae and the Inhibitory Component Involved


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
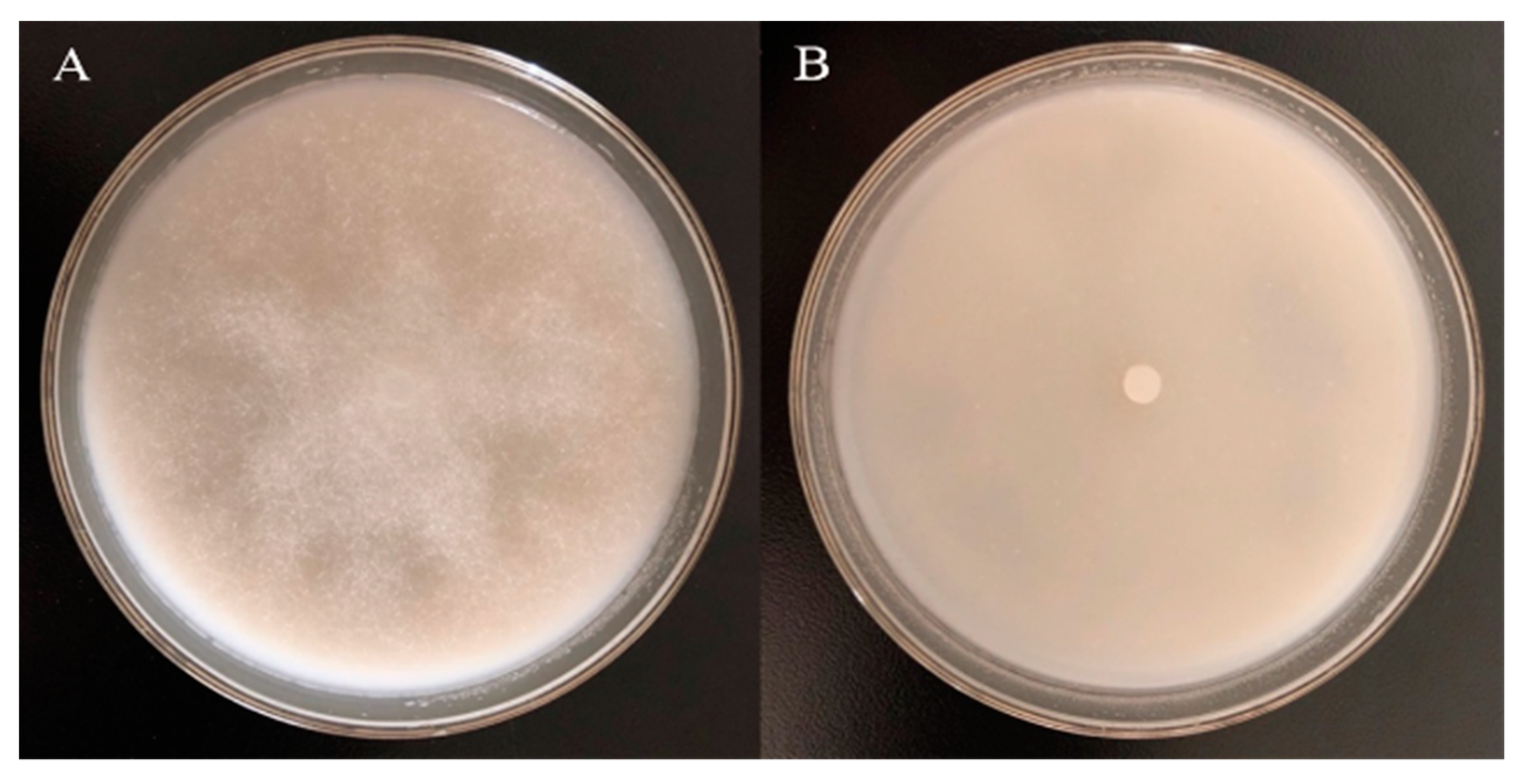
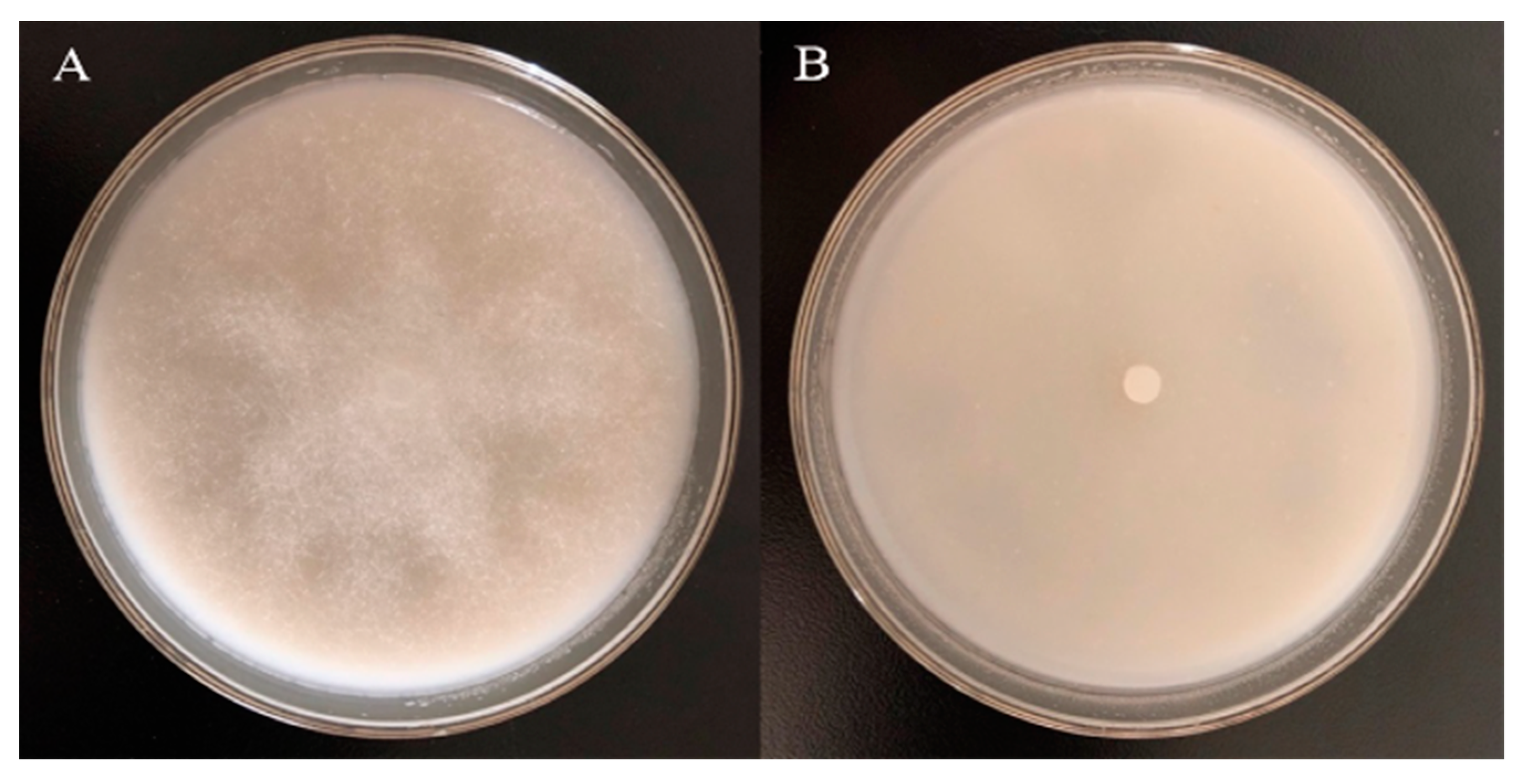
2.2. Inhibition Test of Garlic Essential Oil against Phytophthora Nicotianae
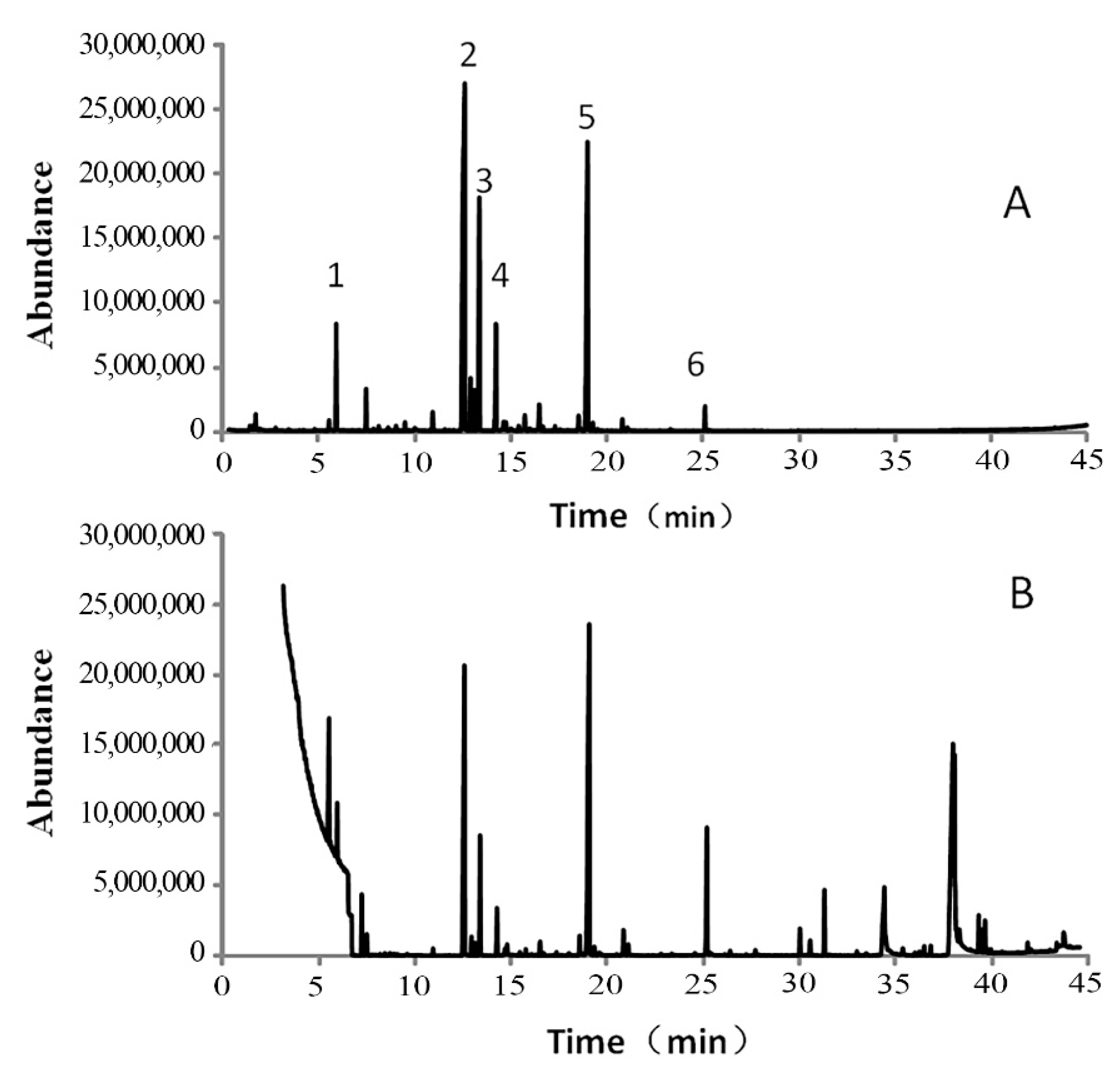
2.3. Gas Chromatography and Gas Chromatography–Mass Spectrometry Analysis
2.4. Headspace Solid Phase Microextraction Method for Sampling of Volatile Compounds
2.5. Inhibition Test of Diallyl Disulfide against Phytophthora Nicotianae
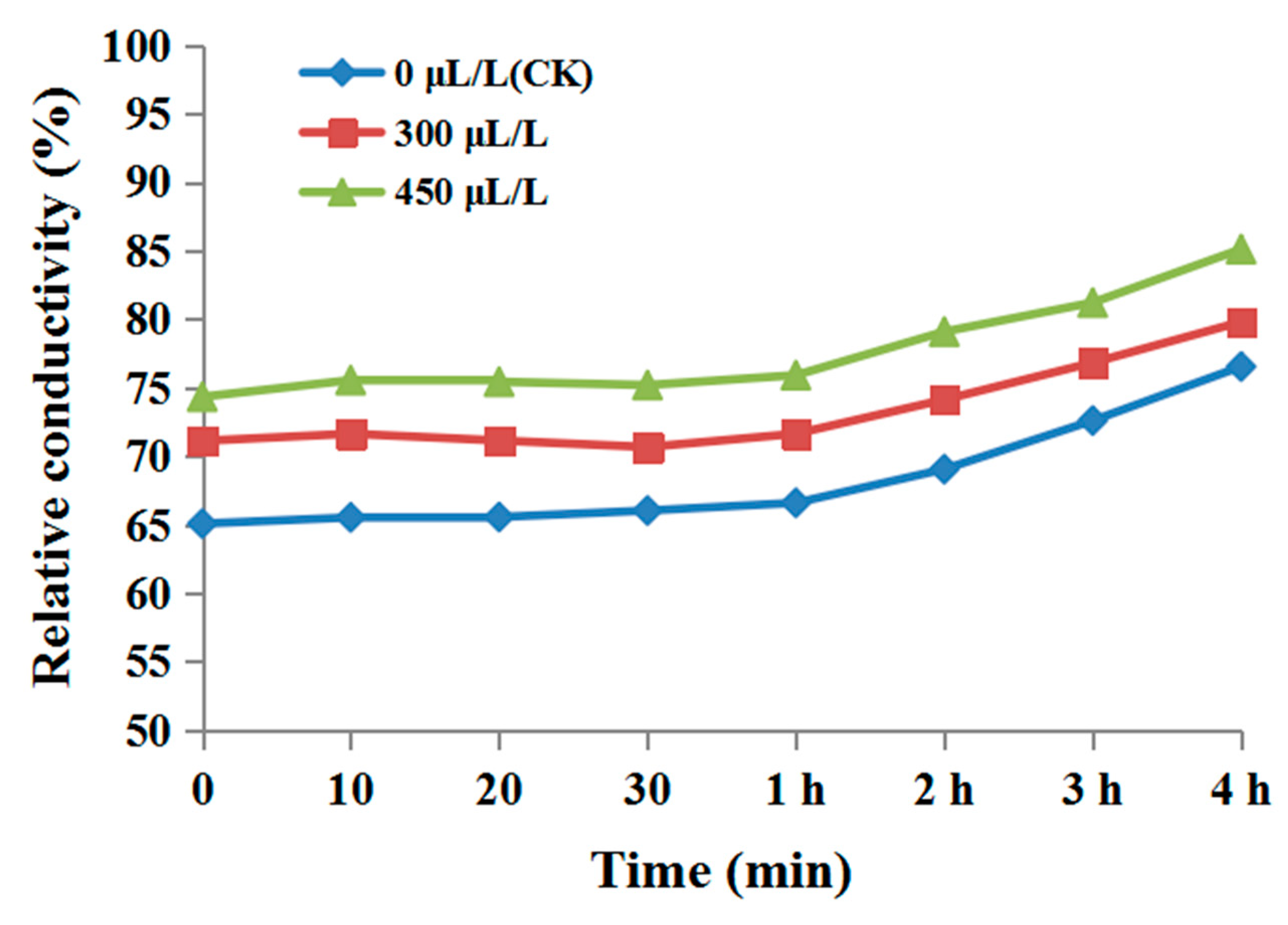
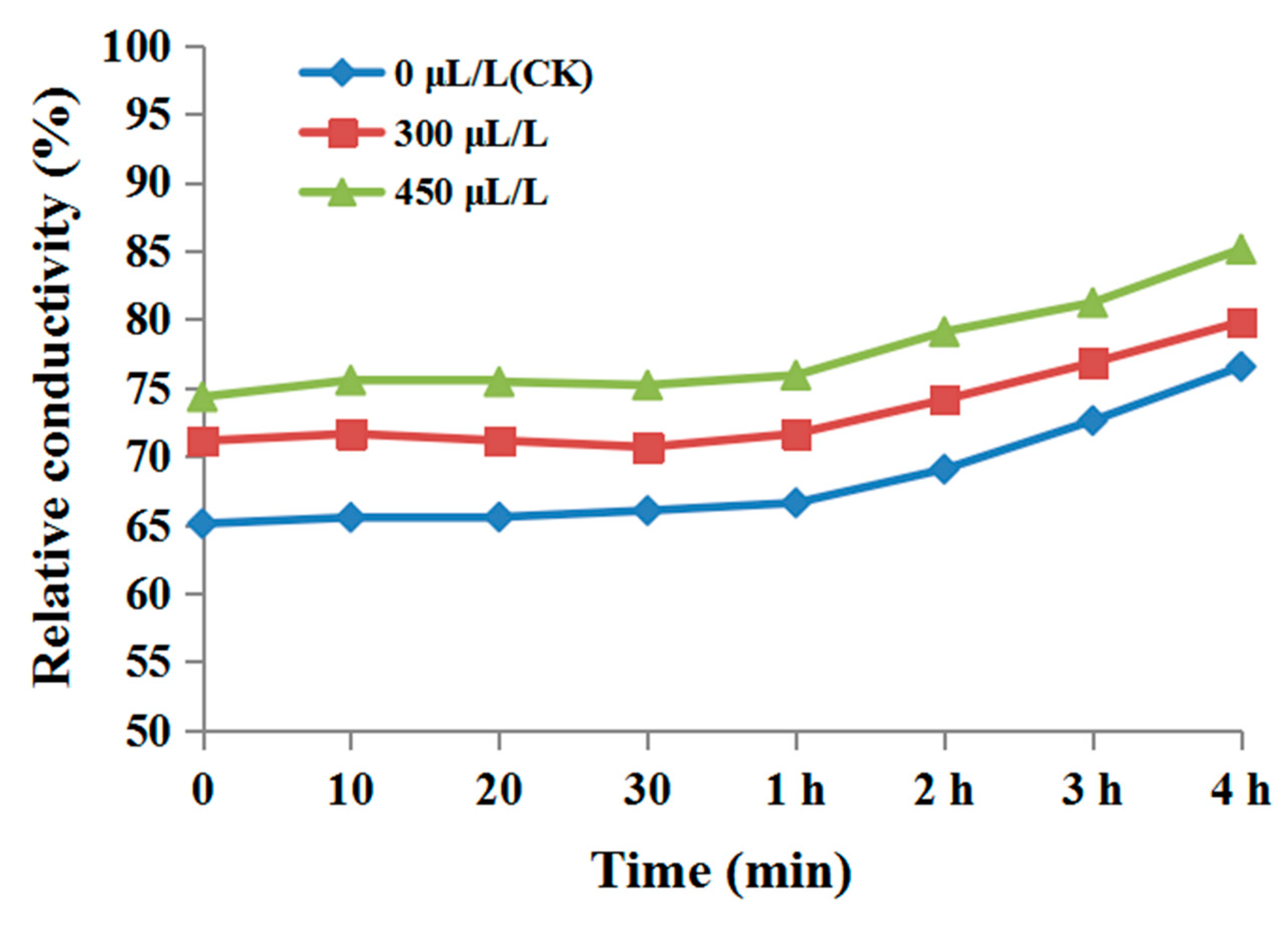
2.6. Cell Membrane Permeability
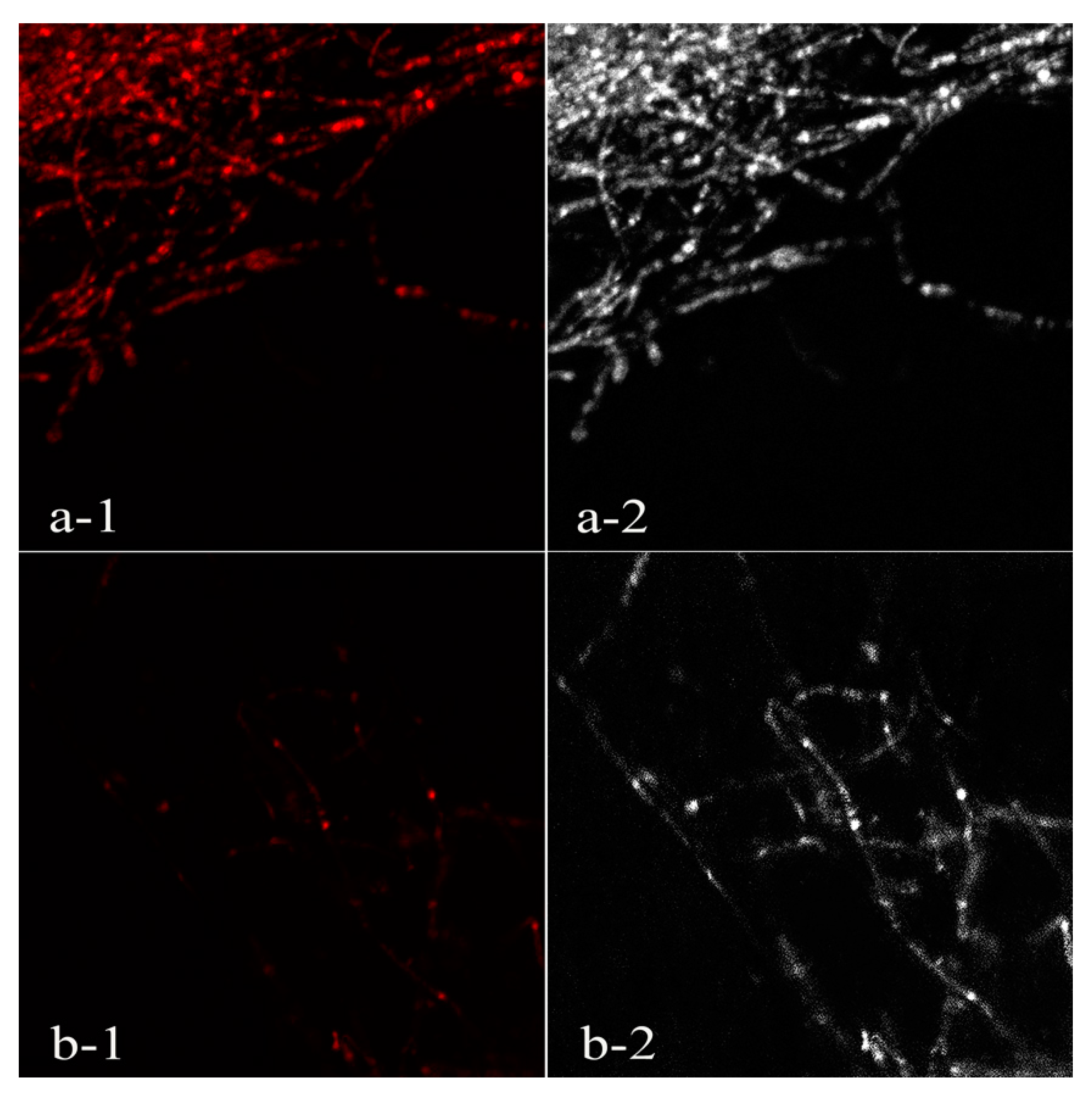
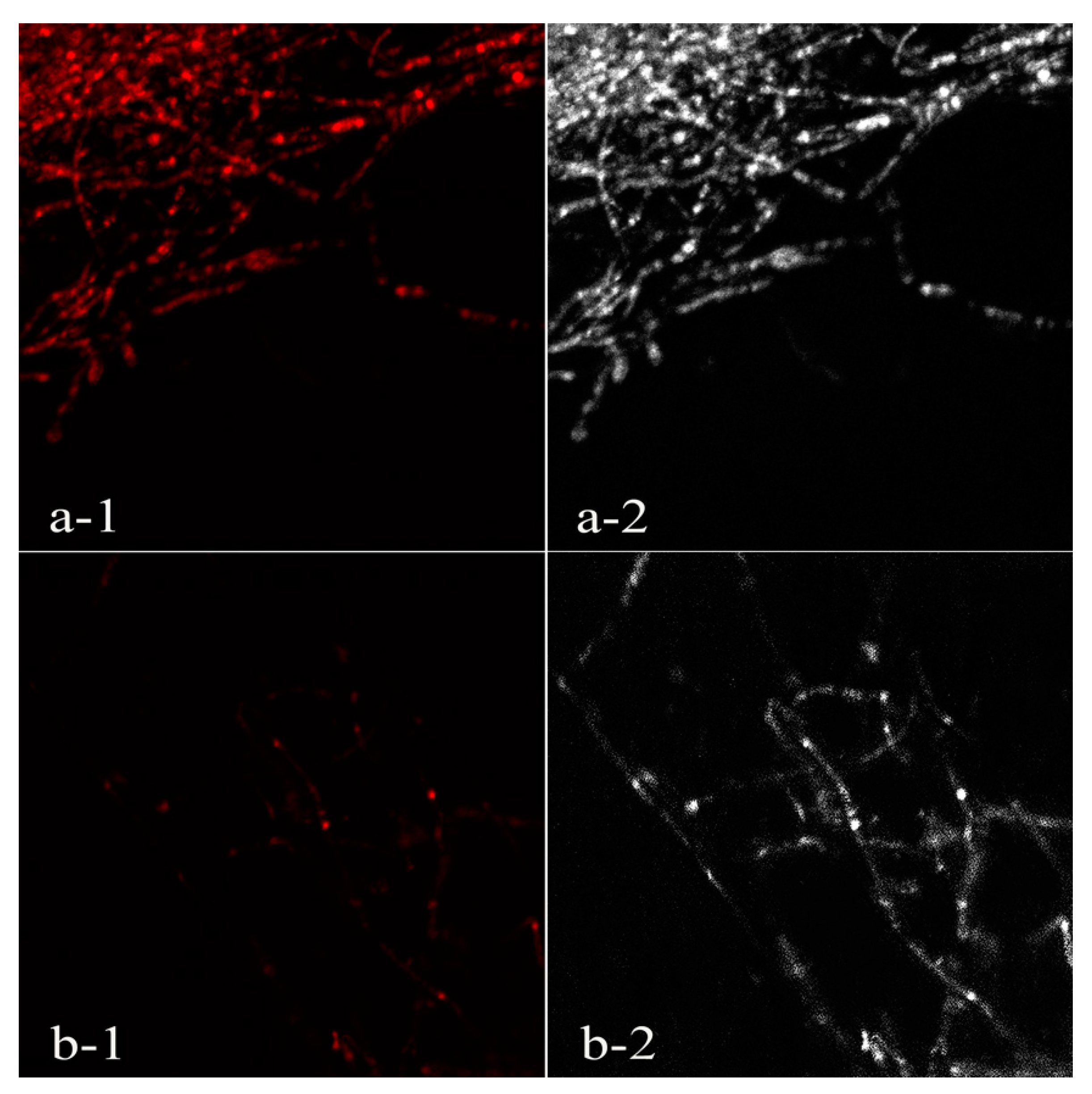
2.7. Propidium Iodide Fluorescence Assay of Phytophthora Nicotianae Mycelia
2.8. Pot Experiment of Garlic Essential Oil and diallyl disulfide for Disease Prevention
2.9. Dynamic Detection of Rhizosphere Pathogens by Real-Time Quantitative Polymerase Chain Reaction
2.10. Promoting Effect of Diallyl Disulfide
2.11. Statistical Analysis
3. Results and Discussion
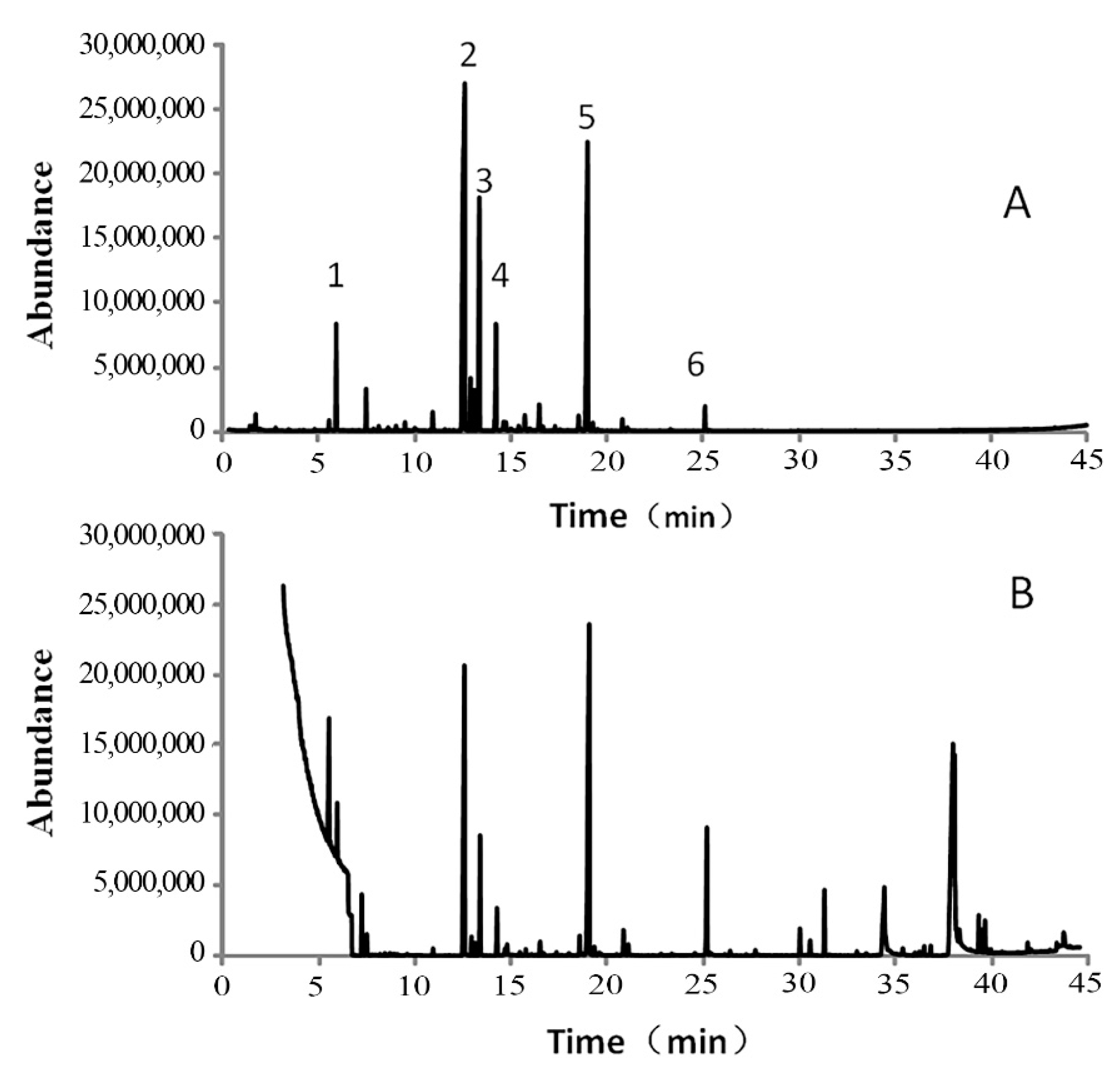
3.1. Main Chemical Components of Garlic Essential Oil
3.2. The Inhibitory Effect of Different Concentrations of Garlic Essential Oil and Diallyl Disulfide against Phytophthora Nicotianae
3.3. The Effect of Diallyl Disulfide on the Cell Membrane Permeability of Phytophthora Nicotianae
3.4. Propidium Iodide Fluorescence Assay of Phytophthora Nicotianae Mycelia
3.5. Prevention of Phytophthora Nicotianae by Garlic Essential Oil and Diallyl Disulfide in Pot Experiment
3.6. Dynamic Detection of Rhizosphere Pathogens by Real-Time Quantitative Polymerase Chain Reaction
3.7. Promoting Effect of Diallyl Disulfide
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Panabières, F.; Ali, G.S.; Allagui, M.B.; Dalio, R.J.D.; Gudmestad, N.C.; Kuhn, M.; Roy, S.G.; Schena, L.; Zampounis, A. Phytophthora nicotianae diseases worldwide: New knowledge of a long-recognised pathogen. Phytopatho. Mediterr. 2016, 55, 20–40. [Google Scholar] [CrossRef]
- Wang, H.C.; Chen, X.J.; Cai, L.T.; Cao, Y.; Lu, N.; Xia, H.Q.; Wang, M.S.; Shang, S.H. Race distribution and distribution of sensitivities to mefenoxam among isolates of Phytophthora parasitica var. nicotianae in Guizhou province of China. Crop Prot. 2013, 52, 136–140. [Google Scholar] [CrossRef]
- Taghavi, T.; Kim, C.; Rahemi, A. Role of Natural volatiles and essential oils in extending shelf life andcontrolling postharvest microorganisms of small fruits. Microorganisms 2018, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Mari, M.; Bautista-Baños, S.; Sivakumar, D. Decay control in the postharvest system: Role of microbial and plant volatile organic compounds. Postharvest Biol. Tec. 2016, 122, 70–81. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; Feo, V.D. Essential Oils and Antifungal Activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef]
- Prakash, B.; Kedia, A.; Mishra, P.K.; Dubey, N.K. Plant essential oils as food preservatives to control moulds, mycotoxin contamination and oxidative deterioration of agri-food commodities—Potentials and challenges. Food Ctrl. 2015, 47, 381–391. [Google Scholar] [CrossRef]
- Camiletti, B.X.; Asensio, C.M.; Pecci, M.D.L.P.G.; Lucini, E.I. Natural control of corn postharvest fungi Aspergillus flavus and Penicillium sp. using essential oils from plants grown in Argentina. J. Food Sci. 2014, 79, 2499–2506. [Google Scholar] [CrossRef]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Obagwu, J.; Korsten, L. Control of citrus green and blue molds with garlic extracts. Eur. J. Plant Pathol. 2003, 109, 221–225. [Google Scholar] [CrossRef]
- Song, W.G.; Li, B.J.; Shi, Y.X.; Liu, K.Q. Inhibitory effects of garlic extract on Botrytis cinerea. China Vegetables 2005, 21–22. [Google Scholar]
- Zhang, H.Q.; Lai, R.Q.; Chen, Z.M.; Zeng, W.L.; Zhong, X.J. Inhibitory effects of garlic extracts on Ralstonia solanacearum in laboratory. Tobacco Sci. Tech. 2009, 3, 62–64. [Google Scholar] [CrossRef]
- Lai, R.Q.; Jiang, L.C.; Chen, Z.M.; Zhou, D.X.; Zhou, D.J. Inhibitory effects of garlic extract on Phytophthora parasitica var. nicotianae in laboratory. Tobacco Sci. Tech. 2009, 9, 62–64. [Google Scholar] [CrossRef]
- Neufeld, J.D.; Knowles, R. Inhibition of nitrifiers and methanotrophs from an agricultural humisol by allylsulfide and its implications for environmental studies. Appl. Environ. Microb. 1999, 65, 2461–2465. [Google Scholar]
- Cheng, Z.H.; Tong, F.; Jin, R. Primary Study on the inhibitory effects on plant pathogen and the inhibitory ingredient of garlic straw aqueous extracts. Acta Bot. Boreal.-Occident. Sin. 2008, 28, 0324–0330. [Google Scholar] [CrossRef]
- Gong, B.; Bloszies, S.; Li, X.; Wei, M.; Yang, F.J.; Shi, Q.H.; Wang, X.F. Efficacy of garlic straw application against root-knot nematodes on tomato. Sci. Hortic. 2013, 161, 49–57. [Google Scholar] [CrossRef]
- Hayat, S.; Cheng, Z.H.; Ahmad, H.; Ali, M.; Chen, X.J.; Wang, M.Y. Garlic, from remedy to stimulant: Evaluation of antifungal potential reveals diversity in phytoalexin allicin content among garlic cultivars; allicin containing aqueous garlic extracts trigger antioxidants. Front. Plant Sci. 2016, 7, 15. [Google Scholar] [CrossRef]
- Avato, P.; Tursi, F.; Vitali, C.; Miccolis, V.; Candido, V. Allylsulfide constituents of garlic volatile oil as antimicrobial agents. Phytomedicine 2000, 7, 239–243. [Google Scholar] [CrossRef]
- Ma, C.Z.; Li, S.D.; Miao, Z.Q.; Guo, R.J.; Yang, S.J.; Gu, Z.R. Efficacy evaluation of applying organic sulfide fumigants to plant pathogens-polluted soil in vegetable greenhouse. Acta. Agr. Shanghai 2009, 25, 41–46. [Google Scholar] [CrossRef]
- Zhang, C.; Gao, J.; Han, T.; Tian, X.; Wang, F. Integrated control of tobacco black shank by combined use of riboflavin and Bacillus subtilis strain Tpb55. BioControl 2017, 62, 835–845. [Google Scholar] [CrossRef]
- Han, X.; Zhao, J.; Cao, J.; Zhang, C. Essential oil of Chrysanthemum indicum L.: Potential biocontrol agent against plant pathogen Phytophthora nicotianae. Environ. Sci. Pollu. Res. 2019, 26, 7013–7023. [Google Scholar] [CrossRef]
- Paul, S.; Dubey, R.C.; Maheswari, D.K.; Kang, S.C. Trachyspermum ammi L. fruit essential oil influencing on membrane permeability and surface characteristics in inhibiting food-borne pathogens. Food Ctrl. 2011, 22, 725–731. [Google Scholar] [CrossRef]
- Fan, S.; Chang, J.; Zong, Y.; Hu, G.; Jia, J. GC-MS analysis of the composition of the essential oil from Dendranthema indicum Var. Aromaticum using three extraction methods and two columns. Molecules 2018, 23, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Ge, C.; Liu, S.; Chen, C.; Zhou, M. Effect of phenylpyrrole fungicide fludioxonil on morphological and physiological characteristics of Sclerotinia sclerotiorum. Pestic. Biochem. Phys. 2013, 106, 61–67. [Google Scholar] [CrossRef]
- Guntiya, N.; Bussaban, B.; Faiyue, B.; Uthaibutra, J.; Saengnil, K. Application of gaseous chlorine dioxide for control of fungal fruit rotdisease of harvested ‘Daw’ longan. Sci. Hortic-Amsterdam 2016, 213, 164–172. [Google Scholar] [CrossRef]
- Yang, S.; Yong, W.; Li, R.H.; Xing, Z.; Jun, T.F. Antifungal activity and action mode of Cuminic acid from the aeeds of Cuminum cyminum L. against Fusarium oxysporum f. sp. Niveum (FON) causing fusarium wilt on watermelon. Molecules 2017, 22, 2053. [Google Scholar] [CrossRef]
- Jing, C.L.; Gou, J.Y.; Han, X.B.; Wu, Q.; Zhang, C.S. In vitro and in vivo activities of eugenol against tobacco black shank caused by Phytophthora nicotianae. Pestic. Biochem. Phys. 2017, 142, 148–154. [Google Scholar] [CrossRef]
- Han, T.; You, C.; Zhang, L.; Feng, C.; Zhang, C.; Wang, J.; Kong, F. Biocontrol potential of antagonist Bacillus subtilis Tpb55 against tobacco black shank. BioControl 2016, 61, 195–205. [Google Scholar] [CrossRef]
- Liu, X. Consruction of Compound Microorganism against Phytophthora parasitica var. nicotianae. Master’s Thesis, Chinese Academy of Agricultural Sciences, Qingdao, China, 2012. [Google Scholar]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of garlic and its bioactive components. J. Nutr. 2001, 131, 955S–962S. [Google Scholar] [CrossRef]
- Block, E. Chemistry of garlic and onions. Sci. Am. 1985, 252, 114–122. [Google Scholar] [CrossRef]
- Mnayer, D.; Fabiano-Tixier, A.S.; Petitcolas, E.; Hamieh, T.; Nehme, N.; Ferrant, C.; Fernandez, X.; Chemat, F. Chemical composition, antibacterial and antioxidant activities of six essentials oils from the Alliaceae family. Molecules 2014, 19, 20034–20053. [Google Scholar] [CrossRef] [PubMed]
- Oyedemi, S.; Okoh, A.; Mabinya, L.V.; Pirochenva, G.; Afolayan, A. The proposed mechanism of bactericidal action of eugenol, ∝-terpineol and g-terpinene against Listeria monocytogenes, Streptococcus pyogenes, Proteus vulgaris and Escherichia coli. Afr. J. Biotechnol. 2009, 8, 1280–1286. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, S.G.; Liang, W.; Mei, J.; Di, Y.; Lan, H.H.; Yang, Y.; Wang, W.W.; Luo, Y.Y.; Wang, H.Z. Antibacterial activity and mode of action of Mentha arvensis ethanol extract against multidrug-resistant Acinetobacter baumannii. Trop. J. Pharm. Res. 2015, 14, 2099–2106. [Google Scholar] [CrossRef]
- Arasu, M.V.; Viayaraghavan, P.; Ilavenil, S.; Al-Dhabi, N.A.; Choi, K.C. Essential oil of four medicinal plants and protective properties in plum fruits against the spoilage bacteria and fungi. Ind. Crop Prod. 2019, 133, 54–62. [Google Scholar] [CrossRef]
- Silva, F.; Ferreira, S.; Duarte, A.; Mendonça, D.I.; Domingues, F.C. Antifungal activity of Coriandrum sativum essential oil, its mode of action against Candida species and potential synergism with amphotericin B. Phytomedicine 2011, 19, 42–47. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z.H.; Meng, H.W.; Tang, X.W. The Garlic Allelochemical Diallyl Disulfide Affects Tomato Root Growth by Influencing Cell Division, Phytohormone Balance and Expansin Gene Expression. Front. Plant Sci. 2016, 7, 1134–1199. [Google Scholar] [CrossRef]
- Ren, K.L.; Hayat, S.; Qi, X.F.; Liu, T.; Cheng, Z.H. The garlic allelochemical DADS influences cucumber root growth involved in regulating hormone levels and modulating cell cycling. Plant Physiol. 2018, 230, 51–60. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | RT a, min | Relative Amount (%) | Identification Method d | |
|---|---|---|---|---|
| HS-SPME | SD b | |||
| Disulfide, dimethyl | 3.43 | 0.05 | Tr c | MS |
| Diacetone alcohol | 5.53 | 0.25 | 7 | MS, RI |
| Diallyl sulfide (DAS) | 5.9 | 5.69 | 1.78 | MS, RI |
| Disulfide, methyl 2-propenyl | 7.48 | 1.39 | tr | MS, RI |
| Disulfide, methyl (1E) -1-propenyl | 8.13 | 0.15 | 0.08 | MS, RI |
| 1,2-Dithiole | 8.64 | 0.08 | 0.05 | MS, RI |
| Dimethyl trisulfide | 9.04 | 0.08 | tr | MS, RI |
| α-Limonene | 10.92 | 0.73 | 0.19 | MS, RI |
| Diallyl disulfide (DADS) | 12.62 | 60.12 | 19.09 | MS, RI |
| 1-Allyl-2-isopropyldisulfane | 12.92 | 1.43 | 0.43 | MS, RI |
| (E)-1-Allyl-2-(prop-1-en-1-yl) disulfane | 13.1 | 1.07 | 0.32 | MS, RI |
| Disulfide, dipropyl | 13.38 | 9.13 | 2.95 | MS, RI |
| Trisulfide, methyl 2-propenyl | 14.25 | 2.24 | 0.86 | MS, RI |
| 4-Methyl-1,2,3-trithiolane | 14.77 | 0.17 | 0.22 | MS, RI |
| 3-Vinyl-3,6-dihydro-1,2-dithiine | 15.75 | 0.35 | 0.15 | MS, RI |
| 2-Vinyl-4H-1,3-dithiine | 16.5 | 0.66 | 0.36 | MS, RI |
| Carvone | 17.31 | 0.12 | 0.11 | MS, RI |
| Anethol | 18.53 | 0.42 | 0.57 | MS, RI |
| Trisulfide, di-2-propenyl (DATS) | 19.02 | 14.18 | 17.98 | MS, RI |
| 1-Allyl-3-propyltrisulfane | 19.29 | 0.14 | 0.17 | MS, RI |
| 5-Methyl-1,2,3,4-tetrathiane | 20.83 | 0.14 | 0.33 | MS, RI |
| Tetrasulfide, di-2-propenyl | 25.12 | 0.45 | 3.85 | MS, RI |
| 1-Allyl-3-(2-(allylthio) propyl) trisulfane | 31.23 | tr | 1.31 | MS, RI |
| n-Hexadecanoic acid | 34.37 | tr | 4.26 | MS, RI |
| 1-Allyl-3-(2-(allyldisulfanyl) propyl) trisulfane | 36.41 | tr | 0.28 | MS, RI |
| 9,12-Octadecadienoic acid (Z,Z)- | 37.92 | tr | 16.72 | MS, RI |
| 9-Octadecenoic acid, (E)- | 38.01 | tr | 7.74 | MS, RI |
| Octadecanoic acid | 38.2 | tr | 0.42 | MS, RI |
| Isopropyl linoleate | 39.28 | tr | 0.64 | MS, RI |
| β-Monolinolein | 43.7 | tr | 0.61 | MS, RI |
| Total identified (%) | 99.03 | 88.47 | ||
| Treatment | Root Irrigation Effect | Fumigation Effect | ||
|---|---|---|---|---|
| Disease Index | Control Effect (%) | Disease Index | Control Effect (%) | |
| GEO 1000 a | 66.67 ± 1.10 b | 19.40 ± 0.53 d | 48.15 ± 1.33 b | 41.80 ± 0.265 c |
| GEO 500 b | 44.44 ± 2.42 d | 46.35 ± 1.70 b | 41.98 ± 1.39 c | 49.27 ± 0.51 b |
| DADS 1000 c | 51.85 ± 2.02 c | 37.36 ± 1.00 c | 46.91 ± 1.85 bc | 43.33 ± 0.93 c |
| DADS 500 d | 41.48 ± 1.21 d | 49.87 ± 0.31 a | 33.33 ± 2.42 d | 59.80 ± 2.01 a |
| CK e | 82.72 ± 1.91 a | - | 82.72 ± 1.91 a | - |
| Treatment | Copies of Root Irrigation Effect | Copies of Fumigation Effect |
|---|---|---|
| GEO 1000 1 | 558,251.04 ± 9385.42 b | 15,531.74 ± 617.53 d |
| GEO 500 2 | 38,276.60 ± 2752.51 d | 1413.87 ± 98.33 d |
| DADS 1000 3 | 238,512.00 ± 22,176.61 cd | 4623.86 ± 246.49 d |
| DADS 500 4 | 62,955.76 ± 80.94 d | 752.19 ± 2.50 d |
| CK 5 | 9,234,399.25 ± 324,740.51 a | 9,234,399.25 ± 324,740.51 a |
| Treatment | Root Length (mm) |
|---|---|
| CK 1 | 19.50 ± 0.764 d |
| DADS 1000 2 | 27.33 ± 0.601 c |
| DADS 500 3 | 27.83 ± 0.603 c |
| DADS 200 4 | 41.50 ± 0.866 b |
| DADS 100 5 | 44.83 ± 0.928 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wei, K.; Han, X.; Zhao, D.; Zheng, Y.; Chao, J.; Gou, J.; Kong, F.; Zhang, C.-S. The Antifungal Effect of Garlic Essential Oil on Phytophthora nicotianae and the Inhibitory Component Involved. Biomolecules 2019, 9, 632. https://doi.org/10.3390/biom9100632
Wang Y, Wei K, Han X, Zhao D, Zheng Y, Chao J, Gou J, Kong F, Zhang C-S. The Antifungal Effect of Garlic Essential Oil on Phytophthora nicotianae and the Inhibitory Component Involved. Biomolecules. 2019; 9(10):632. https://doi.org/10.3390/biom9100632
Chicago/Turabian StyleWang, Yaochen, Keke Wei, Xiaobin Han, Donglin Zhao, Yanfen Zheng, Jianmin Chao, Jianyu Gou, Fanyu Kong, and Cheng-Sheng Zhang. 2019. "The Antifungal Effect of Garlic Essential Oil on Phytophthora nicotianae and the Inhibitory Component Involved" Biomolecules 9, no. 10: 632. https://doi.org/10.3390/biom9100632
APA StyleWang, Y., Wei, K., Han, X., Zhao, D., Zheng, Y., Chao, J., Gou, J., Kong, F., & Zhang, C.-S. (2019). The Antifungal Effect of Garlic Essential Oil on Phytophthora nicotianae and the Inhibitory Component Involved. Biomolecules, 9(10), 632. https://doi.org/10.3390/biom9100632