The MMP-8 rs11225395 Promoter Polymorphism Increases Cancer Risk of Non-Asian Populations: Evidence from a Meta-Analysis

Abstract
1. Introduction
2. Materials and Methods
2.1. Data Sources and Identification
2.2. Study Eligibility Evaluation and Data Extraction
2.3. Statistics Analysis
3. Results
3.1. Characteristics of Included Studies
3.2. Main Analysis Results
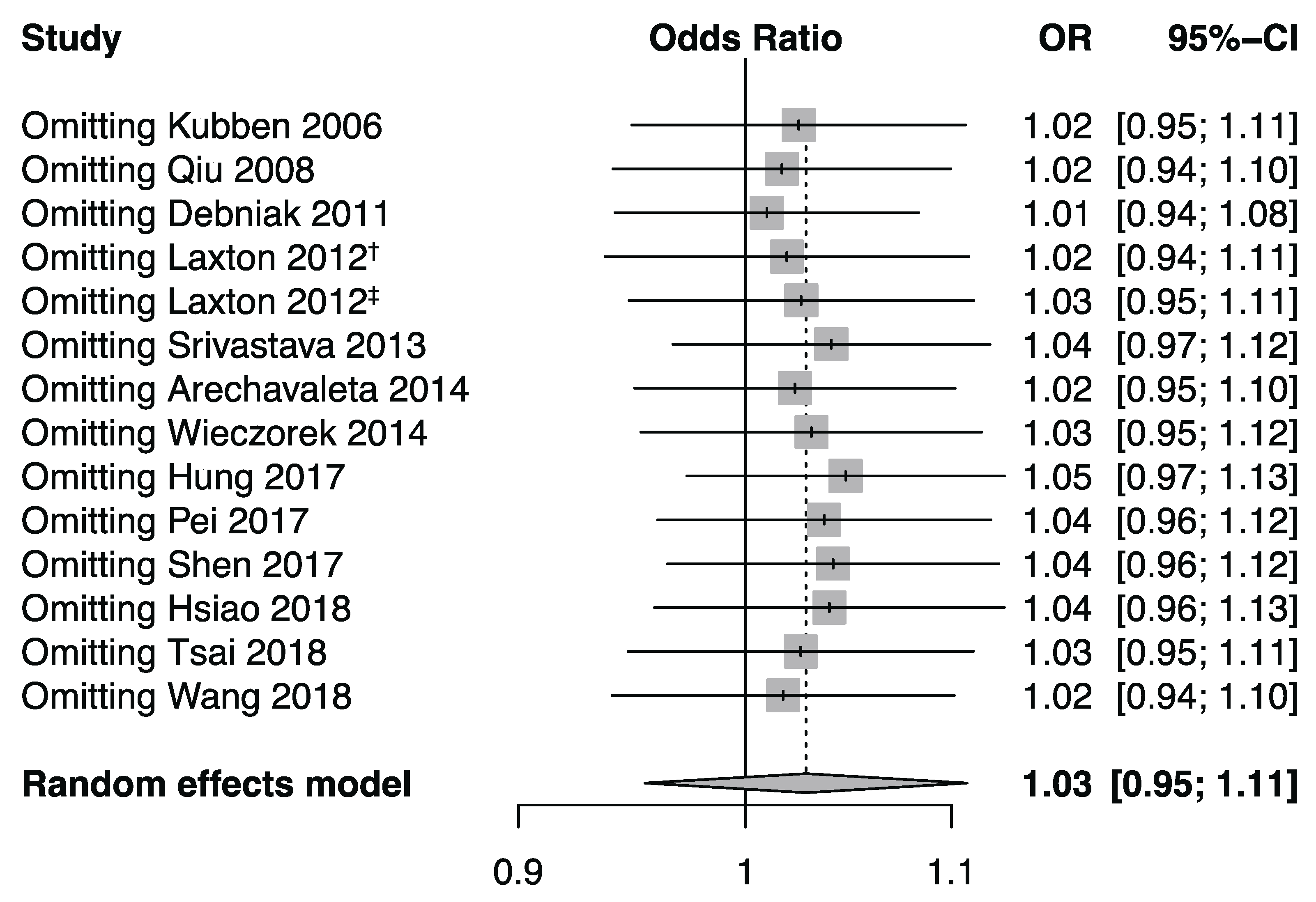
3.3. Publication Bias and Sensitivity Analysis
3.4. Between-Study Heterogeneity Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hao, X.; Xia, L.; Qu, R.; Yang, X.; Jiang, M.; Zhou, B. Association between miR-146a rs2910164 polymorphism and specific cancer susceptibility: An updated meta-analysis. Fam. Cancer 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.; Zhang, C.; Zhu, H.; Yang, X.; Xu, L.; Liu, J.; Lu, J.; Zhan, L.; Cheng, H.; Sun, X. Association between survivin-31G> C polymorphism and cancer risk: Meta-analysis of 29 studies. J. Cancer Res. Clin. Oncol. 2014, 140, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, Y.; Wang, G. Quantitative assessment of the influence of EPHX1 gene polymorphisms and cancer risk: A meta-analysis with 94,213 subjects. J. Exp. Clin. Cancer Res. 2014, 33, 82. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.-B.; Wang, D.-G.; Xuan, C.; Sun, G.-L.; Deng, K.-F. Genetic 135G/C polymorphism of RAD51 gene and risk of cancer: A meta-analysis of 28,956 cases and 28,372 controls. Fam. Cancer 2014, 13, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Astrom, P.; Juurikka, K.; Hadler-Olsen, E.S.; Svineng, G.; Cervigne, N.K.; Coletta, R.D.; Risteli, J.; Kauppila, J.H.; Skarp, S.; Kuttner, S.; et al. The interplay of matrix metalloproteinase-8, transforming growth factor-beta1 and vascular endothelial growth factor-C cooperatively contributes to the aggressiveness of oral tongue squamous cell carcinoma. Br J Cancer 2017, 117, 1007–1016. [Google Scholar] [CrossRef]
- Korpi, J.T.; Kervinen, V.; Maklin, H.; Vaananen, A.; Lahtinen, M.; Laara, E.; Ristimaki, A.; Thomas, G.; Ylipalosaari, M.; Astrom, P.; et al. Collagenase-2 (matrix metalloproteinase-8) plays a protective role in tongue cancer. Br J Cancer 2008, 98, 766–775. [Google Scholar] [CrossRef]
- Decock, J.; Hendrickx, W.; Vanleeuw, U.; Van Belle, V.; Van Huffel, S.; Christiaens, M.-R.; Ye, S.; Paridaens, R. Plasma MMP1 and MMP8 expression in breast cancer: Protective role of MMP8 against lymph node metastasis. BMC Cancer 2008, 8, 77. [Google Scholar] [CrossRef]
- Gutierrez-Fernandez, A.; Fueyo, A.; Folgueras, A.R.; Garabaya, C.; Pennington, C.J.; Pilgrim, S.; Edwards, D.R.; Holliday, D.L.; Jones, J.L.; Span, P.N.; et al. Matrix metalloproteinase-8 functions as a metastasis suppressor through modulation of tumor cell adhesion and invasion. Cancer Res. 2008, 68, 2755–2763. [Google Scholar] [CrossRef]
- Stadlmann, S.; Pollheimer, J.; Moser, P.L.; Raggi, A.; Amberger, A.; Margreiter, R.; Offner, F.A.; Mikuz, G.; Dirnhofer, S.; Moch, H. Cytokine-regulated expression of collagenase-2 (MMP-8) is involved in the progression of ovarian cancer. Eur. J. Cancer (Oxford, England: 1990) 2003, 39, 2499–2505. [Google Scholar] [CrossRef]
- Lempinen, M.; Lyytinen, I.; Nordin, A.; Tervahartiala, T.; Makisalo, H.; Sorsa, T.; Isoniemi, H. Prognostic value of serum MMP-8, -9 and TIMP-1 in patients with hepatocellular carcinoma. Ann. Med. 2013, 45, 482–487. [Google Scholar] [CrossRef]
- Sirnio, P.; Tuomisto, A.; Tervahartiala, T.; Sorsa, T.; Klintrup, K.; Karhu, T.; Herzig, K.H.; Makela, J.; Karttunen, T.J.; Salo, T.; et al. High-serum MMP-8 levels are associated with decreased survival and systemic inflammation in colorectal cancer. Br. J. Cancer 2018, 119, 213–219. [Google Scholar] [CrossRef]
- Decock, J.; Long, J.-R.; Laxton, R.C.; Shu, X.-O.; Hodgkinson, C.; Hendrickx, W.; Pearce, E.G.; Gao, Y.-T.; Pereira, A.C.; Paridaens, R.; et al. Association of Matrix Metalloproteinase-8 Gene Variation with Breast Cancer Prognosis. Cancer Res. 2007, 67, 10214–10221. [Google Scholar] [CrossRef] [PubMed]
- Debniak, T.; Jakubowska, A.; Serrano-Fernandez, P.; Kurzawski, G.; Cybulski, C.; Chauhan, S.R.; Laxton, R.C.; Maleszka, R.; Lubinski, J.; Ye, S. Association of MMP8 gene variation with an increased risk of malignant melanoma. Melanoma Res. 2011, 21, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Z.; Du, X.M.; Jing, Q.M.; Li, X.X.; Gu, R.X.; Wang, J.; Han, Y.L. Impact of matrix metalloproteinase-8 gene variations on the risk of thoracic aortic dissection in a Chinese Han population. Mol Biol Rep. 2013, 40, 5953–5958. [Google Scholar] [CrossRef] [PubMed]
- Beeghly-Fadiel, A.C.; Long, J.; Gao, Y.; Zheng, Y.; Cai, Q.; Gu, K.; Chen, Z.; Lu, W.; Zheng, W.; Shu, X.O. Abstract 905: Associations with breast cancer survival replicated for three SNPs: A report from the Shanghai Breast Cancer Survival Study and Shanghai Breast Cancer Study. Cancer Res. 2010, 70, 905. [Google Scholar] [CrossRef]
- Arechavaleta-Velasco, F.; Cuevas-Antonio, R.; Dominguez-Lopez, P.; Estrada-Moscoso, I.; Imani-Razavi, F.S.; Zeferino-Toquero, M.; Diaz-Cueto, L. Matrix metalloproteinase-8 promoter gene polymorphisms in Mexican women with ovarian cancer. Med Oncol. (NorthwoodLond. Engl.) 2014, 31, 132. [Google Scholar] [CrossRef]
- DerSimonian, R.; Laird, N. Meta-analysis in clinical trials. Control. Clin. Trials 1986, 7, 177–188. [Google Scholar] [CrossRef]
- Mantel, N.; Haenszel, W. Statistical aspects of the analysis of data from retrospective studies of disease. J. Natl. Cancer Inst. 1959, 22, 719–748. [Google Scholar]
- Galbraith, R. A note on graphical presentation of estimated odds ratios from several clinical trials. Stat. Med. 1988, 7, 889–894. [Google Scholar] [CrossRef]
- Egger, M.; Smith, G.D.; Schneider, M.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef]
- Begg, C.B.; Mazumdar, M. Operating characteristics of a rank correlation test for publication bias. Biometrics 1994, 1088–1101. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. BMJ 2009, 339. [Google Scholar] [CrossRef] [PubMed]
- Laxton, R.C. A study of matrix metalloproteinases in cancer and atherosclerosis; Queen Mary University of London: London, UK, 2012. [Google Scholar]
- Hashim, N.A.N.; Ramzi, N.H.; Velapasamy, S.; Alex, L.; Chahil, J.K.; Lye, S.H.; Munretnam, K.; Haron, M.R.; Ler, L.W. Identification of genetic and non-genetic risk factors for nasopharyngeal carcinoma in a Southeast Asian population. Asian Pac. J. Cancer Prev. 2012, 13, 6005–6010. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, C.-L.; Liu, L.-C.; Shih, T.-C.; Chuang, C.-L.; Chen, G.-L.; Wang, H.-C.; Pan, S.-Y.; Shen, T.-C.; Tsai, C.-W.; Chang, W.-S.; et al. The Association of Matrix Metalloproteinase-8 Promoter Genotypes in Breast Cancer. Anticancer Res. 2018, 38, 2181–2185. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.-W.; Tsai, C.-W.; Wu, C.-N.; Shih, L.-C.; Chen, Y.-Y.; Liu, Y.-F.; Hung, H.-S.; Shen, M.-Y.; Chang, W.-S.; Bau, D.-T. The Contribution of Matrix Metalloproteinase-8 Promoter Polymorphism to Oral Cancer Susceptibility. In Vivo 2017, 31, 585–590. [Google Scholar] [CrossRef]
- Kubben, F.J.j.M.; Sier, C.F.M.; Meijer, M.J.W.; van den Berg, M.; van der Reijden, J.J.; Griffioen, G.; van de Velde, C.J.H.; Lamers, C.B.H.W.; Verspaget, H.W. Clinical impact of MMP and TIMP gene polymorphisms in gastric cancer. Br. J. Cancer 2006, 95, 744–751. [Google Scholar] [CrossRef]
- Pei, J.S.; Chang, W.S.; Hsu, P.C.; Hung, Y.W.; Cheng, S.P.; Tsai, C.W.; Bau, D.T.; Gong, C.L. The Contribution of MMP-8 Promoter Genotypes to Childhood Leukemia. In Vivo 2017, 31, 1059–1064. [Google Scholar] [CrossRef][Green Version]
- Qiu, W.; Zhou, G.; Zhai, Y.; Zhang, X.; Xie, W.; Zhang, H.; Yang, H.; Zhi, L.; Yuan, X.; Zhang, X.; et al. No association of MMP-7, MMP-8, and MMP-21 polymorphisms with the risk of hepatocellular carcinoma in a Chinese population. Cancer Epidemiol. Biomark. Prev. A Publ. Am. Assoc. Cancer Res. Cosponsored By Am. Soc. Prev. Oncol. 2008, 17, 2514–2518. [Google Scholar] [CrossRef]
- Shen, T.-C.; Hsia, T.-C.; Chao, C.-Y.; Chen, W.-C.; Chen, C.-Y.; Chen, W.-C.; Lin, Y.-T.; Hsiao, C.-L.; Chang, W.-S.; Tsai, C.-W.; et al. The Contribution of MMP-8 Promoter Polymorphisms in Lung Cancer. Anticancer Res. 2017, 37, 3563–3567. [Google Scholar] [CrossRef]
- Srivastava, P.; Kapoor, R.; Mittal, R.D. Association of single nucleotide polymorphisms in promoter of matrix metalloproteinase-2, 8 genes with bladder cancer risk in Northern India. Urol. Oncol. 2013, 31, 247–254. [Google Scholar] [CrossRef]
- Tsai, T.-H.; Wang, Y.-M.; Chang, W.-S.; Tsai, C.-W.; Wu, H.-C.; Hsu, H.-M.; Wang, Y.-C.; Li, H.-T.; Gong, C.-L.; Bau, D.-T.; et al. Association of Matrix Metalloproteinase-8 Genotypes with the Risk of Bladder Cancer. Anticancer Res. 2018, 38, 5159–5164. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhou, Y.; Li, G.; Wen, X.; Kou, Y.; Yu, J.; He, H.; Zhao, Q.; Xue, F.; Wang, J. MMP8 and MMP9 gene polymorphisms were associated with breast cancer risk in a Chinese Han population. Sci. Rep. 2018, 8, 13422. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, E.; Reszka, E.; Wasowicz, W.; Grzegorczyk, A.; Konecki, T.; Sosnowski, M.; Jablonowski, Z. MMP7 and MMP8 genetic polymorphisms in bladder cancer patients. Cent. Eur. J. Urol. 2014, 66, 405–410. [Google Scholar] [CrossRef] [PubMed]
- 1000 Genomes Project Consortium. A global reference for human genetic variation. Nature 2015, 526, 68. [Google Scholar] [CrossRef] [PubMed]
- Greco, T.; Zangrillo, A.; Biondi-Zoccai, G.; Landoni, G. Meta-analysis: Pitfalls and hints. Heart Lung Vessel 2013, 5, 219–225. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Year | Country | Region | Cancer Type | Matching Criteria | Genotyping Methods | Case-Control | HWE (p) | NOS |
|---|---|---|---|---|---|---|---|---|---|
| Kubben | 2006 | Dutch | Europe | gastric cancer | NA | PCR-RFLP | 79-169 | 0.75 | 6 |
| Qiu | 2008 | China | Asia | hepatocellular carcinoma | NA | PCR-RFLP | 417-480 | 0.22 | 2 |
| Debniak | 2011 | UK | Europe | melanoma | Age, gender | TaqMan | 296-290 | 0.75 | 6 |
| Hashim | 2012 | Malaysia | Asia | nasopharyngeal carcinoma | Age, gender | Microarray | 48-48 | NA | 6 |
| Laxton | 2012 | UK | Europe | breast cancer | Age | TaqMan | 2172-2268 | 0.43 | 6 |
| Laxton | 2012 | Poland | Europe | breast cancer | Age, gender | TaqMan | 297-303 | NA | 5 |
| Srivastava | 2013 | India | Asia | bladder cancer | Age, gender | PCR-RFLP | 200-200 | 0.48 | 8 |
| Arechavaleta | 2014 | Mexico | Latin America | ovarian cancer | NA | PCR-RFLP | 35-37 | 0.01 | 5 |
| Wieczorek | 2014 | Poland | Europe | bladder cancer | NA | TaqMan | 241-199 | 0.70 | 5 |
| Hung | 2017 | China | Asia | oral cancer | Age, gender | PCR-RFLP | 788-956 | <0.01 | 7 |
| Pei | 2017 | China | Asia | leukemia | Age | PCR-RFLP | 266-266 | 0.14 | 7 |
| Shen | 2017 | China | Asia | lung cancer | Age, gender | PCR-RFLP | 358-716 | <0.01 | 8 |
| Hsiao | 2018 | China | Asia | breast cancer | Age, gender | PCR-RFLP | 1232-1232 | <0.01 | 6 |
| Tsai | 2018 | China | Asia | bladder cancer | Age, gender | PCR-RFLP | 375-375 | 0.08 | 8 |
| Wang | 2018 | China | Asia | breast cancer | NA | Mass spectrum | 571-578 | NA | 7 |
| Author | Year | Country | Cancer | Control | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CC | CT | TT | CC+CT | CT+TT | C | T | MAF | CC | CT | TT | CC+CT | CT+TT | C | T | MAF | ||||
| Kubben | 2006 | Dutch | 19 | 46 | 14 | 65 | 60 | 84 | 74 | 0.47 | 55 | 81 | 33 | 136 | 114 | 191 | 147 | 0.43 | |
| Qiu | 2008 | China | 140 | 196 | 81 | 336 | 277 | 476 | 358 | 0.43 | 184 | 216 | 80 | 400 | 296 | 584 | 376 | 0.39 | |
| Debniak | 2011 | UK | 86 | 152 | 58 | 238 | 210 | 324 | 268 | 0.45 | 113 | 134 | 43 | 247 | 177 | 360 | 220 | 0.38 | |
| Hashim | 2012 | Malaysia | NA | NA | 1 | 47 | NA | NA | NA | NA | NA | NA | 8 | 40 | NA | NA | NA | NA | |
| Laxton | 2012 | UK | 628 | 1105 | 439 | 1733 | 1544 | 2361 | 1983 | 0.46 | 735 | 1096 | 437 | 1831 | 1533 | 2566 | 1970 | 0.43 | |
| Laxton | 2012 | Poland | NA | NA | NA | NA | NA | 332 | 262 | 0.44 | NA | NA | NA | NA | NA | 348 | 258 | 0.43 | |
| Srivastava | 2013 | India | 99 | 90 | 11 | 189 | 101 | 288 | 112 | 0.28 | 92 | 84 | 24 | 176 | 108 | 268 | 132 | 0.33 | |
| Arechavaleta | 2014 | Mexico | 6 | 16 | 13 | 22 | 29 | 28 | 42 | 0.60 | 6 | 26 | 5 | 32 | 31 | 38 | 36 | 0.49 | |
| Wieczorek | 2014 | Poland | 72 | 125 | 44 | 197 | 169 | 269 | 213 | 0.44 | 60 | 101 | 38 | 161 | 139 | 221 | 177 | 0.44 | |
| Hung | 2017 | China | 414 | 284 | 90 | 698 | 374 | 1112 | 464 | 0.29 | 466 | 364 | 126 | 830 | 490 | 1296 | 616 | 0.32 | |
| Pei | 2017 | China | 139 | 98 | 29 | 237 | 127 | 376 | 156 | 0.29 | 129 | 105 | 32 | 234 | 137 | 363 | 169 | 0.31 | |
| Shen | 2017 | China | 188 | 130 | 40 | 318 | 170 | 506 | 210 | 0.29 | 351 | 273 | 92 | 624 | 365 | 975 | 457 | 0.32 | |
| Hsiao | 2018 | China | 648 | 466 | 118 | 1114 | 584 | 1762 | 702 | 0.28 | 633 | 468 | 131 | 1101 | 599 | 1734 | 730 | 0.30 | |
| Tsai | 2018 | China | 186 | 152 | 37 | 338 | 189 | 524 | 226 | 0.30 | 197 | 140 | 38 | 337 | 178 | 534 | 216 | 0.29 | |
| Wang | 2018 | China | NA | NA | NA | NA | NA | 724 | 418 | 0.37 | NA | NA | NA | NA | NA | 769 | 387 | 0.33 | |
| Comparison | Allele Model (T vs. C) | Homozygote Model (TT vs. CC) | Heterozygote Model (CT vs. CC) | Dominant Model (CT+TT vs. CC) | Recessive Model (TT vs. CC+CT) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (Number of Study) | OR (95% CI) | p | ph | OR (95% CI) | p | ph | OR (95% CI) | p | ph | OR (95% CI) | p | ph | OR (95% CI) | p | ph | |
| Overall (15) | 1.03(0.95,1.11) | 0.46 | 0.02 | 1.01(0.85,1.20) | 0.90 | 0.03 | 1.06(0.98,1.14) | 0.14 | 0.13 | 1.04(0.93,1.16) | 0.49 | 0.03 | 0.97(0.83,1.13) | 0.70 | 0.05 | |
| Region | ||||||||||||||||
| Asia (9) | 0.97(0.89,1.07) | 0.57 | 0.07 | 0.89(0.77,1.03) | 0.12 | 0.18 | 0.97(0.88,1.07) | 0.55 | 0.57 | 0.95(0.87,1.04) | 0.26 | 0.34 | 0.89(0.77,1.02) | 0.09 | 0.14 | |
| Others (6) | 1.11(1.04,1.19) | <0.01 | 0.43 | 1.22(1.05,1.41) | 0.01 | 0.39 | 1.21(1.07,1.36) | <0.01 | 0.43 | 1.21(1.08,1.35) | <0.01 | 0.45 | 1.09(0.96,1.24) | 0.19 | 0.17 | |
| Cancer type | ||||||||||||||||
| Bladder (3) | 0.97(0.83,1.12) | 0.67 | 0.29 | 0.85(0.61,1.17) | 0.32 | 0.14 | 1.08(0.87,1.33) | 0.49 | 0.84 | 1.02(0.84,1.25) | 0.81 | 0.58 | 0.83(0.61,1.12) | 0.22 | 0.15 | |
| Breast (4) | 1.06(1.00,1.13) | 0.07 | 0.20 | 1.04(0.78,1.37) | 0.80 | 0.08 | 1.08(0.89,1.30) | 0.43 | 0.08 | 1.07(0.86,1.31) | 0.55 | 0.04 | 1.02(0.89,1.16) | 0.79 | 0.25 | |
| Digestive System (3) | 1.03(0.83,1.28) | 0.78 | 0.05 | 0.99(0.79,1.24) | 0.95 | 0.11 | 1.10(0.80,1.49) | 0.57 | 0.07 | 1.09(0.79,1.51) | 0.59 | 0.04 | 0.97(0.79,1.20) | 0.80 | 0.30 | |
| Others (5) | 1.07(0.83,1.39) | 0.60 | 0.02 | 1.14(0.71,1.86) | 0.58 | 0.05 | 1.01(0.74,1.37) | 0.96 | 0.09 | 1.03(0.75,1.44) | 0.84 | 0.04 | 1.08(0.65,1.79) | 0.78 | 0.02 | |
| Sample Size | ||||||||||||||||
| ≤500 (5) | 0.98(0.82,1.16) | 0.80 | 0.20 | 0.87(0.60,1.27) | 0.48 | 0.11 | 1.08(0.83,1.41) | 0.56 | 0.46 | 1.02(0.79,1.31) | 0.88 | 0.51 | 0.82(0.41,1.64) | 0.57 | <0.01 | |
| >500 (10) | 1.03(0.95,1.12) | 0.41 | 0.01 | 1.03(0.87,1.23) | 0.71 | 0.04 | 1.05(0.93,1.18) | 0.43 | 0.06 | 1.04(0.92,1.19) | 0.52 | <0.01 | 1.01(0.91,1.11) | 0.86 | 0.44 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Chen, Y.; Hua, W.; Sun, X.; Chen, Y.; Liu, Y.; Fan, J.; Zhao, Y.; Zhao, L.; Xu, X.; et al. The MMP-8 rs11225395 Promoter Polymorphism Increases Cancer Risk of Non-Asian Populations: Evidence from a Meta-Analysis. Biomolecules 2019, 9, 570. https://doi.org/10.3390/biom9100570
Feng J, Chen Y, Hua W, Sun X, Chen Y, Liu Y, Fan J, Zhao Y, Zhao L, Xu X, et al. The MMP-8 rs11225395 Promoter Polymorphism Increases Cancer Risk of Non-Asian Populations: Evidence from a Meta-Analysis. Biomolecules. 2019; 9(10):570. https://doi.org/10.3390/biom9100570
Chicago/Turabian StyleFeng, Jiarong, Yudi Chen, Wenxi Hua, Xiaohan Sun, Yanjie Chen, Yu Liu, Jiaying Fan, Yuening Zhao, Lixiang Zhao, Xiaojing Xu, and et al. 2019. "The MMP-8 rs11225395 Promoter Polymorphism Increases Cancer Risk of Non-Asian Populations: Evidence from a Meta-Analysis" Biomolecules 9, no. 10: 570. https://doi.org/10.3390/biom9100570
APA StyleFeng, J., Chen, Y., Hua, W., Sun, X., Chen, Y., Liu, Y., Fan, J., Zhao, Y., Zhao, L., Xu, X., & Yang, X. (2019). The MMP-8 rs11225395 Promoter Polymorphism Increases Cancer Risk of Non-Asian Populations: Evidence from a Meta-Analysis. Biomolecules, 9(10), 570. https://doi.org/10.3390/biom9100570