Emerging Insights into the Function of Kinesin-8 Proteins in Microtubule Length Regulation

Abstract
1. Microtubules and Dynamic Properties
1.1. Microtubule Structure
1.2. Dynamic Instability
2. Kinesin-8 Family of Motor Proteins
2.1. Localization
2.2. Cellular Roles
3. Biophysical Properties of Kinesin-8 Proteins
4. Emerging Insights into the Structure and Function of Kinesin-8 Motor Domain
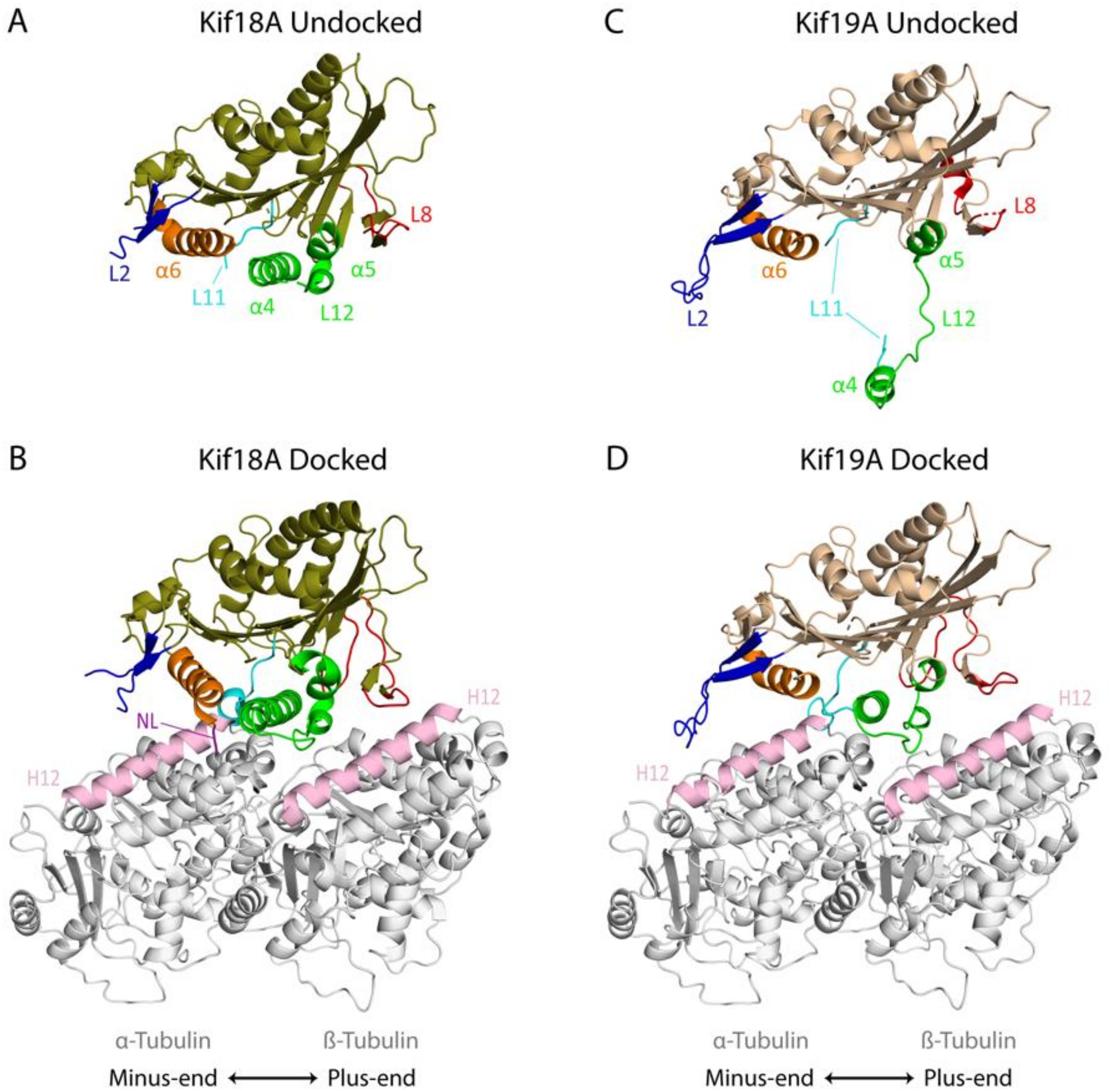
4.1. Insights from Structural Studies
4.1.1. Kif18A
4.1.2. Kif19A
4.2. Key Elements Associated with Microtubule Destabilization
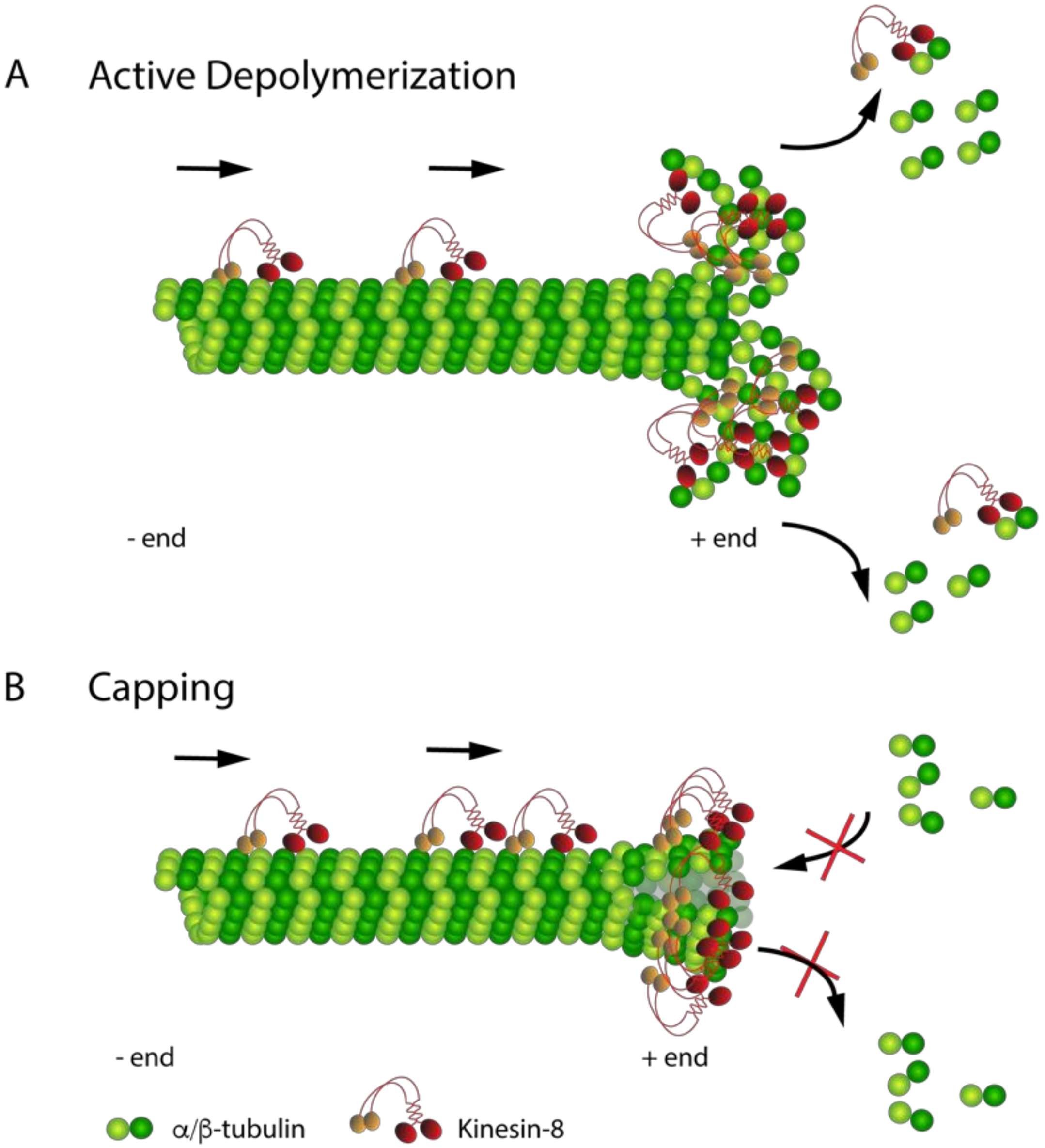
4.3. Emerging Microtubule Depolymerization Mechanisms of Kinesin-8 Proteins
5. Importance of C-Terminal tails in Modulating Kinesin-8 Function
5.1. Localization and Destabilization Activity
5.2. Crosslinking and Sliding
5.3. Regulation of Kinesin-8 proteins through Association with Other Regulators
6. Conclusions and Future Directions
Funding
Acknowledgments
Conflicts of Interest
References
- Desai, A.; Mitchison, T.J. Microtubule polymerization dynamics. Annu. Rev. Cell Dev. Biol. 1997, 13, 83–117. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, T.J. Localization of an exchangeable GTP binding site at the plus end of microtubules. Science 1993, 261, 1044–1047. [Google Scholar] [CrossRef] [PubMed]
- Weisenberg, R.C.; Deery, W.J.; Dickinson, P.J. Tubulin-nucleotide interactions during the polymerization and depolymerization of microtubules. Biochemistry 1976, 15, 4248–4254. [Google Scholar] [CrossRef] [PubMed]
- Carlier, M.F.; Pantaloni, D. Kinetic analysis of guanosine 5′-triphosphate hydrolysis associated with tubulin polymerization. Biochemistry 1981, 20, 1918–1924. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, T.J.; Kirschner, M.W. Dynamic instability of microtubule growth. Nature 1984, 312, 237–242. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, E.T.; Voter, W.A.; Erickson, H.P. GTP hydrolysis during microtubule assembly. Biochemistry 1987, 26, 4148–4156. [Google Scholar] [CrossRef] [PubMed]
- Drechsel, D.N.; Kirschner, M.W. The minimum GTP cap required to stabilize microtubules. Curr. Biol. 1994, 4, 1053–1061. [Google Scholar] [CrossRef]
- Seetapun, D.; Castle, B.T.; McIntyre, A.J.; Tran, P.T.; Odde, D.J. Estimating the microtubule GTP cap size in vivo. Curr. Biol. 2012, 22, 1681–1687. [Google Scholar] [CrossRef]
- Coombes, C.E.; Yamamoto, A.; Kenzie, M.R.; Odde, D.J.; Gardner, M.K. Evolving tip structures can explain age-dependent microtubule catastrophe. Curr. Biol. 2013, 23, 1342–1348. [Google Scholar] [CrossRef]
- Bowne-Anderson, H.; Hibbel, A.; Howard, J. Regulation of microtubule growth and catastrophe: Unifying theory and experiment. Trends Cell Biol. 2015, 25, 769–779. [Google Scholar] [CrossRef]
- Vemu, A.; Atherton, J.; Spector, J.O.; Moores, C.A.; Roll-Mecak, A. Tubulin isoform composition tunes microtubule dynamics. Mol. Biol. Cell 2017, 28, 3564–3572. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.R.; O’Toole, E.; Morgan, G.; Austin, J.; Ulyanov, E.; Ataullakhanov, F.; Gudimchuk, N. Microtubules grow by the addition of bent guanosine triphosphate tubulin to the tips of curved protofilaments. J. Cell Biol. 2018, 217, 2691–2708. [Google Scholar] [CrossRef] [PubMed]
- Mandelkow, E.M.; Mandelkow, E.; Milligan, R.A. Microtubule dynamics and microtubule caps: A time-resolved cryo-electron microscopy study. J. Cell Biol. 1991, 114, 977–991. [Google Scholar] [CrossRef] [PubMed]
- Chretien, D.; Fuller, S.D.; Karsenti, E. Structure of growing microtubule ends: Two-dimensional sheets close into tubes at variable rates. J. Cell Biol. 1995, 129, 1311–1328. [Google Scholar] [CrossRef] [PubMed]
- Brouhard, G.J. Dynamic instability 30 years later: Complexities in microtubule growth and catastrophe. Mol. Biol. Cell 2015, 26, 1207–1210. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.A.; O’Brien, E.T.; Pryer, N.K.; Sobeiro, M.F.; Voter, W.A.; Erickson, H.P.; Salmon, E.D. Dynamic instability of individual microtubules analysed by video light microscopy: Rate constants and transition frequencies. J. Cell Biol. 1988, 107, 1437–1448. [Google Scholar] [CrossRef] [PubMed]
- Alushin, G.M.; Lander, G.C.; Kellogg, E.H.; Zhang, R.; Baker, D.; Nogales, E. High-resolution microtubule structures reveal the structural transitions in alphabeta-tubulin upon GTP hydrolysis. Cell 2014, 157, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.K.; Zanic, M.; Gell, C.; Bormuth, V.; Howard, J. Depolymerizing kinesins Kip3 and MCAK shape cellular microtubule architecture by differential control of catastrophe. Cell 2011, 147, 1092–1103. [Google Scholar] [CrossRef] [PubMed]
- Vemu, A.; Szczesna, E.; Zehr, E.A.; Spector, J.O.; Grigorieff, N.; Deaconescu, A.M.; Roll-Mecak, A. Severing enzymes amplify microtubule arrays through lattice GTP-tubulin incorporation. Science 2018, 361. [Google Scholar] [CrossRef] [PubMed]
- Belmont, L.D.; Hyman, A.A.; Sawin, K.E.; Mitchison, T.J. Real-time visualization of cell cycle-dependent changes in microtubule dynamics in cytoplasmic extracts. Cell 1990, 62, 579–589. [Google Scholar] [CrossRef]
- Verde, F.; Dogterom, M.; Stelzer, E.; Karsenti, E.; Leibler, S. Control of microtubule dynamics and length by cyclin A- and cyclin B-dependent kinases in Xenopus egg extracts. J. Cell Biol. 1992, 118, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Rusan, N.M.; Fagerstrom, C.J.; Yvon, A.M.; Wadsworth, P. Cell cycle-dependent changes in microtubule dynamics in living cells expressing green fluorescent protein-alpha tubulin. Mol. Biol. Cell 2001, 12, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Rieder, C.L.; Davison, E.A.; Jensen, L.C.; Cassimeris, L.; Salmon, E.D. Oscillatory movements of monooriented chromosomes and their position relative to the spindle pole result from the ejection properties of the aster and half-spindle. J. Cell Biol. 1986, 103, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Saxton, W.M.; Stemple, D.L.; Leslie, R.J.; Salmon, E.D.; Zavortink, M.; McIntosh, J.R. Tubulin dynamics in cultured mammalian cells. J. Cell Biol. 1984, 99, 2175–2186. [Google Scholar] [CrossRef] [PubMed]
- Gorbsky, G.J.; Borisy, G.G. Microtubules of the kinetochore fiber turn over in metaphase but not in anaphase. J. Cell Biol. 1989, 109, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt, J. Spindle assembly: Asters part their separate ways. Nat. Cell Biol. 2005, 7, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Wuhr, M.; Dumont, S.; Groen, A.C.; Needleman, D.J.; Mitchison, T.J. How does a millimeter-sized cell find its center? Cell Cycle 2009, 8, 1115–1121. [Google Scholar] [CrossRef]
- Olmsted, J.B.; Stemple, D.L.; Saxton, W.M.; Neighbors, B.W.; McIntosh, J.R. Cell cycle-dependent changes in the dynamics of MAP 2 and MAP 4 in cultured cells. J. Cell Biol. 1989, 109, 211–223. [Google Scholar] [CrossRef]
- Walczak, C.E.; Heald, R. Mechanisms of mitotic spindle assembly and function. Int. Rev. Cytol. 2008, 265, 111–158. [Google Scholar] [CrossRef]
- Wickstead, B.; Gull, K. A “holistic” kinesin phylogeny reveals new kinesin families and predicts protein functions. Mol. Biol. Cell 2006, 17, 1734–1743. [Google Scholar] [CrossRef]
- Varga, V.; Helenius, J.; Tanaka, K.; Hyman, A.A.; Tanaka, T.U.; Howard, J. Yeast kinesin-8 depolymerizes microtubules in a length-dependent manner. Nat. Cell Biol. 2006, 8, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Mayr, M.I.; Hummer, S.; Bormann, J.; Gruner, T.; Adio, S.; Woehlke, G.; Mayer, T.U. The human kinesin Kif18A is a motile microtubule depolymerase essential for chromosome congression. Curr. Biol. 2007, 17, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.L., Jr.; Carvalho, P.; Roof, D.M.; Pellman, D. Plus end-specific depolymerase activity of Kip3, a kinesin-8 protein, explains its role in positioning the yeast mitotic spindle. Nat. Cell Biol. 2006, 8, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Stout, J.R.; Yount, A.L.; Powers, J.A.; Leblanc, C.; Ems-McClung, S.C.; Walczak, C.E. Kif18B interacts with EB1 and controls astral microtubule length during mitosis. Mol. Biol. Cell 2011, 22, 3070–3080. [Google Scholar] [CrossRef] [PubMed]
- Tanenbaum, M.E.; Macurek, L.; van der Vaart, B.; Galli, M.; Akhmanova, A.; Medema, R.H. A complex of Kif18b and MCAK promotes microtubule depolymerization and is negatively regulated by Aurora kinases. Curr. Biol. 2011, 21, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Savoian, M.S.; Gatt, M.K.; Riparbelli, M.G.; Callaini, G.; Glover, D.M. Drosophila Klp67A is required for proper chromosome congression and segregation during meiosis I. J. Cell Sci. 2004, 117, 3669–3677. [Google Scholar] [CrossRef]
- Niwa, S.; Nakajima, K.; Miki, H.; Minato, Y.; Wang, D.; Hirokawa, N. KIF19A is a microtubule-depolymerizing kinesin for ciliary length control. Dev. Cell 2012, 23, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Stumpff, J.; von Dassow, G.; Wagenbach, M.; Asbury, C.; Wordeman, L. The kinesin-8 motor Kif18A suppresses kinetochore movements to control mitotic chromosome alignment. Dev. Cell 2008, 14, 252–262. [Google Scholar] [CrossRef]
- Fukuda, Y.; Luchniak, A.; Murphy, E.R.; Gupta, M.L., Jr. Spatial control of microtubule length and lifetime by opposing stabilizing and destabilizing functions of Kinesin-8. Curr. Biol. 2014, 24, 1826–1835. [Google Scholar] [CrossRef]
- Savoian, M.S.; Glover, D.M. Drosophila Klp67A binds prophase kinetochores to subsequently regulate congression and spindle length. J. Cell Sci. 2010, 123, 767–776. [Google Scholar] [CrossRef]
- Cottingham, F.R.; Hoyt, M.A. Mitotic spindle positioning in Saccharomyces cerevisiae is accomplished by antagonistically acting microtubule motor proteins. J. Cell Biol. 1997, 138, 1041–1053. [Google Scholar] [CrossRef] [PubMed]
- DeZwaan, T.M.; Ellingson, E.; Pellman, D.; Roof, D.M. Kinesin-related KIP3 of Saccharomyces cerevisiae is required for a distinct step in nuclear migration. J. Cell Biol. 1997, 138, 1023–1040. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.K.; Heller, K.K.; Frisen, L.; Wallack, D.L.; Loayza, D.; Gammie, A.E.; Rose, M.D. The kinesin-related proteins, Kip2p and Kip3p, function differently in nuclear migration in yeast. Mol. Biol. Cell 1998, 9, 2051–2068. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.T.; Marsh, L.; Doye, V.; Inoue, S.; Chang, F. A mechanism for nuclear positioning in fission yeast based on microtubule pushing. J. Cell Biol. 2001, 153, 397–411. [Google Scholar] [CrossRef] [PubMed]
- McHugh, T.; Gluszek, A.A.; Welburn, J.P.I. Microtubule end tethering of a processive kinesin-8 motor Kif18b is required for spindle positioning. J. Cell Biol. 2018, 217, 2403–2416. [Google Scholar] [CrossRef] [PubMed]
- Straight, A.F.; Sedat, J.W.; Murray, A.W. Time-lapse microscopy reveals unique roles for kinesins during anaphase in budding yeast. J. Cell Biol. 1998, 143, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Arellano-Santoyo, H.; Portran, D.; Gaillard, J.; Vantard, M.; Thery, M.; Pellman, D. Microtubule-sliding activity of a kinesin-8 promotes spindle assembly and spindle-length control. Nat. Cell Biol. 2013, 15, 948–957. [Google Scholar] [CrossRef]
- Gandhi, R.; Bonaccorsi, S.; Wentworth, D.; Doxsey, S.; Gatti, M.; Pereira, A. The Drosophila kinesin-like protein KLP67A is essential for mitotic and male meiotic spindle assembly. Mol. Biol. Cell 2004, 15, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.N.; Ems-McClung, S.C.; Stout, J.R.; LeBlanc, C.; Shaw, S.L.; Gardner, M.K.; Walczak, C.E. Kif18A uses a microtubule binding site in the tail for plus-end localization and spindle length regulation. Curr. Biol. 2011, 21, 1500–1506. [Google Scholar] [CrossRef] [PubMed]
- West, R.R.; Malmstrom, T.; Troxell, C.L.; McIntosh, J.R. Two related kinesins, klp5+ and klp6+, foster microtubule disassembly and are required for meiosis in fission yeast. Mol. Biol. Cell 2001, 12, 3919–3932. [Google Scholar] [CrossRef] [PubMed]
- West, R.R.; Malmstrom, T.; McIntosh, J.R. Kinesins klp5+ and klp6+ are required for normal chromosome movement in mitosis. J. Cell Sci. 2002, 115, 931–940. [Google Scholar] [PubMed]
- Gergely, Z.R.; Crapo, A.; Hough, L.E.; McIntosh, J.R.; Betterton, M.D. Kinesin-8 effects on mitotic microtubule dynamics contribute to spindle function in fission yeast. Mol. Biol. Cell 2016, 27, 3490–3514. [Google Scholar] [CrossRef] [PubMed]
- Walczak, C.E.; Zong, H.; Jain, S.; Stout, J.R. Spatial regulation of astral microtubule dynamics by Kif18B in PtK cells. Mol. Biol. Cell 2016, 27, 3021–3030. [Google Scholar] [CrossRef] [PubMed]
- Rizk, R.S.; Discipio, K.A.; Proudfoot, K.G.; Gupta, M.L., Jr. The kinesin-8 Kip3 scales anaphase spindle length by suppression of midzone microtubule polymerization. J. Cell Biol. 2014, 204, 965–975. [Google Scholar] [CrossRef]
- Liu, X.S.; Zhao, X.D.; Wang, X.; Yao, Y.X.; Zhang, L.L.; Shu, R.Z.; Ren, W.H.; Huang, Y.; Huang, L.; Gu, M.M.; et al. Germinal cell aplasia in Kif18a mutant male mice due to impaired chromosome congression and dysregulated BubR1 and CENP-E. Genes Cancer 2010, 1, 26–39. [Google Scholar] [CrossRef]
- Czechanski, A.; Kim, H.; Byers, C.; Greenstein, I.; Stumpff, J.; Reinholdt, L.G. Kif18a is specifically required for mitotic progression during germ line development. Dev. Biol. 2015, 402, 253–262. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, C.; Chen, H.; Li, L.; Guo, L.; Jiang, W.; Lu, S.H. Kif18A is involved in human breast carcinogenesis. Carcinogenesis 2010, 31, 1676–1684. [Google Scholar] [CrossRef]
- Schiewek, J.; Schumacher, U.; Lange, T.; Joosse, S.A.; Wikman, H.; Pantel, K.; Mikhaylova, M.; Kneussel, M.; Linder, S.; Schmalfeldt, B.; et al. Clinical relevance of cytoskeleton associated proteins for ovarian cancer. J. Cancer Res. Clin. Oncol. 2018. [Google Scholar] [CrossRef]
- Liao, W.; Huang, G.; Liao, Y.; Yang, J.; Chen, Q.; Xiao, S.; Jin, J.; He, S.; Wang, C. High KIF18A expression correlates with unfavorable prognosis in primary hepatocellular carcinoma. Oncotarget 2014, 5, 10271–10279. [Google Scholar] [CrossRef]
- Luo, W.; Liao, M.; Liao, Y.; Chen, X.; Huang, C.; Fan, J.; Liao, W. The role of kinesin KIF18A in the invasion and metastasis of hepatocellular carcinoma. World J. Surg. Oncol. 2018, 16, 36. [Google Scholar] [CrossRef]
- Zhu, H.; Xu, W.; Zhang, H.; Liu, J.; Xu, H.; Lu, S.; Dang, S.; Kuang, Y.; Jin, X.; Wang, Z. Targeted deletion of Kif18a protects from colitis-associated colorectal (CAC) tumors in mice through impairing Akt phosphorylation. Biochem. Biophs. Res. Commun. 2013, 438, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, M.; Nishida, N.; Iwatsuki, M.; Ishimaru, S.; Mimori, K.; Tanaka, F.; Nakagawa, T.; Sato, T.; Sugihara, K.; Hoon, D.S.; et al. Kinesin 18A expression: Clinical relevance to colorectal cancer progression. Int. J. Cancer 2011, 129, 2543–2552. [Google Scholar] [CrossRef]
- Kasahara, M.; Nagahara, M.; Nakagawa, T.; Ishikawa, T.; Sato, T.; Uetake, H.; Sugihara, K. Clinicopathological relevance of kinesin family member 18A expression in invasive breast cancer. Oncol. Lett. 2016, 12, 1909–1914. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, S.; Sun, R.; Lu, M.; Wu, Y.; Li, Y. Expression of KIF18A in gastric cancer and its association with prognosis. Chin. J. Gastrointest. Surg. 2016, 19, 585–589. [Google Scholar]
- Wu, Y.; Wang, A.; Zhu, B.; Huang, J.; Lu, E.; Xu, H.; Xia, W.; Dong, G.; Jiang, F.; Xu, L. KIF18B promotes tumor progression through activating the Wnt/β-catenin pathway in cervical cancer. Onco Targets Ther. 2018, 11, 1707–1720. [Google Scholar] [CrossRef] [PubMed]
- Itzel, T.; Scholz, P.; Maass, T.; Krupp, M.; Marquardt, J.U.; Strand, S.; Becker, D.; Staib, F.; Binder, H.; Roessler, S.; et al. Translating bioinformatics in oncology: Guilt-by-profiling analysis and identification of KIF18B and CDCA3 as novel driver genes in carcinogenesis. Bioinformatics 2015, 31, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Qiu, W.; Gupta, M.L., Jr.; Pereira-Leal, J.B.; Reck-Peterson, S.L.; Pellman, D. Mechanisms underlying the dual-mode regulation of microtubule dynamics by Kip3/kinesin-8. Mol. Cell 2011, 43, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Mayr, M.I.; Storch, M.; Howard, J.; Mayer, T.U. A non-motor microtubule binding site is essential for the high processivity and mitotic function of kinesin-8 Kif18A. PLoS ONE 2011, 6, e27471. [Google Scholar] [CrossRef]
- Stumpff, J.; Du, Y.; English, C.A.; Maliga, Z.; Wagenbach, M.; Asbury, C.L.; Wordeman, L.; Ohi, R. A tethering mechanism controls the processivity and kinetochore-microtubule plus-end enrichment of the kinesin-8 Kif18A. Mol. Cell 2011, 43, 764–775. [Google Scholar] [CrossRef]
- Mockel, M.M.; Heim, A.; Tischer, T.; Mayer, T.U. Xenopus laevis Kif18A is a highly processive kinesin required for meiotic spindle integrity. Biol. Open 2017, 6, 463–470. [Google Scholar] [CrossRef]
- Shin, Y.; Du, Y.; Collier, S.E.; Ohi, M.D.; Lang, M.J.; Ohi, R. Biased Brownian motion as a mechanism to facilitate nanometer-scale exploration of the microtubule plus end by a kinesin-8. Proc. Natl. Acad. Sci. USA 2015, 112, 3826–3835. [Google Scholar] [CrossRef] [PubMed]
- Varga, V.; Leduc, C.; Bormuth, V.; Diez, S.; Howard, J. Kinesin-8 motors act cooperatively to mediate length-dependent microtubule depolymerization. Cell 2009, 138, 1174–1183. [Google Scholar] [CrossRef] [PubMed]
- Grissom, P.M.; Fiedler, T.; Grishchuk, E.L.; Nicastro, D.; West, R.R.; McIntosh, J.R. Kinesin-8 from fission yeast: A heterodimeric, plus-end-directed motor that can couple microtubule depolymerization to cargo movement. Mol. Biol. Cell 2009, 20, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Erent, M.; Drummond, D.R.; Cross, R.A. S. pombe kinesins-8 promote both nucleation and catastrophe of microtubules. PLoS ONE 2012, 7, e30738. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.J.; Dalby, B.; Stewart, R.J.; Doxsey, S.J.; Goldstein, L.S. Mitochondrial association of a plus end-directed microtubule motor expressed during mitosis in Drosophila. J. Cell Biol. 1997, 136, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Arellano-Santoyo, H.; Geyer, E.A.; Stokasimov, E.; Chen, G.Y.; Su, X.; Hancock, W.; Rice, L.M.; Pellman, D. A tubulin binding switch underlies Kip3/Kinesin-8 depolymerase activity. Dev. Cell 2017, 42, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Locke, J.; Joseph, A.P.; Pena, A.; Mockel, M.M.; Mayer, T.U.; Topf, M.; Moores, C.A. Structural basis of human kinesin-8 function and inhibition. Proc. Natl. Acad. Sci. USA 2017, 114, 9539–9548. [Google Scholar] [CrossRef]
- Du, Y.; English, C.A.; Ohi, R. The kinesin-8 Kif18A dampens microtubule plus-end dynamics. Curr. Biol. 2010, 20, 374–380. [Google Scholar] [CrossRef]
- Wang, D.; Nitta, R.; Morikawa, M.; Yajima, H.; Inoue, S.; Shigematsu, H.; Kikkawa, M.; Hirokawa, N. Motility and microtubule depolymerization mechanisms of the Kinesin-8 motor, KIF19A. eLife 2016, 5. [Google Scholar] [CrossRef]
- Unsworth, A.; Masuda, H.; Dhut, S.; Toda, T. Fission yeast kinesin-8 Klp5 and Klp6 are interdependent for mitotic nuclear retention and required for proper microtubule dynamics. Mol. Biol. Cell 2008, 19, 5104–5115. [Google Scholar] [CrossRef]
- Stumpff, J.; Wagenbach, M.; Franck, A.; Asbury, C.L.; Wordeman, L. Kif18A and chromokinesins confine centromere movements via microtubule growth suppression and spatial control of kinetochore tension. Dev. Cell 2012, 22, 1017–1029. [Google Scholar] [CrossRef]
- Peters, C.; Brejc, K.; Belmont, L.; Bodey, A.J.; Lee, Y.; Yu, M.; Guo, J.; Sakowicz, R.; Hartman, J.; Moores, C.A. Insight into the molecular mechanism of the multitasking kinesin-8 motor. EMBO J. 2010, 29, 3437–3447. [Google Scholar] [CrossRef] [PubMed]
- Sablin, E.P.; Kull, F.J.; Cooke, R.; Vale, R.D.; Fletterick, R.J. Crystal structure of the motor domain of the kinesin-related motor ncd. Nature 1996, 380, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Kull, F.J.; Sablin, E.P.; Lau, R.; Fletterick, R.J.; Vale, R.D. Crystal structure of the kinesin motor domain reveals a structural similarity to myosin. Nature 1996, 380, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Woehlke, G.; Ruby, A.K.; Hart, C.L.; Ly, B.; Hom-Booher, N.; Vale, R.D. Microtubule interaction site of the kinesin motor. Cell 1997, 90, 207–216. [Google Scholar] [CrossRef]
- Shipley, K.; Hekmat-Nejad, M.; Turner, J.; Moores, C.; Anderson, R.; Milligan, R.; Sakowicz, R.; Fletterick, R. Structure of a kinesin microtubule depolymerization machine. EMBO J. 2004, 23, 1422–1432. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Saijo, S.; Shimizu, N.; Jiang, X.; Hirokawa, N. Mechanism of Catalytic Microtubule Depolymerization via KIF2-Tubulin Transitional Conformation. Cell Rep. 2017, 20, 2626–2638. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Wang, W.; Jiang, Q.; Wang, C.; Knossow, M.; Gigant, B. The structure of apo-kinesin bound to tubulin links the nucleotide cycle to movement. Nat. Commun. 2014, 5, 5364. [Google Scholar] [CrossRef]
- Shang, Z.; Zhou, K.; Xu, C.; Csencsits, R.; Cochran, J.C.; Sindelar, C.V. High-resolution structures of kinesin on microtubules provide a basis for nucleotide-gated force-generation. eLife 2014, 3, e04686. [Google Scholar] [CrossRef]
- Atherton, J.; Farabella, I.; Yu, I.M.; Rosenfeld, S.S.; Houdusse, A.; Topf, M.; Moores, C.A. Conserved mechanisms of microtubule-stimulated ADP release, ATP binding, and force generation in transport kinesins. eLife 2014, 3, e03680. [Google Scholar] [CrossRef]
- Goulet, A.; Behnke-Parks, W.M.; Sindelar, C.V.; Major, J.; Rosenfeld, S.S.; Moores, C.A. The structural basis of force generation by the mitotic motor kinesin-5. J. Biol. Chem. 2012, 287, 44654–44666. [Google Scholar] [CrossRef] [PubMed]
- Sindelar, C.V.; Downing, K.H. An atomic-level mechanism for activation of the kinesin molecular motors. Proc. Natl. Acad. Sci. USA 2010, 107, 4111–4116. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Nitta, R.; Okada, Y.; Hirokawa, N. A common mechanism for microtubule destabilizers-M type kinesins stabilize curling of the protofilament using the class-specific neck and loops. Cell 2004, 116, 591–602. [Google Scholar] [CrossRef]
- Tan, D.; Rice, W.J.; Sosa, H. Structure of the kinesin13-microtubule ring complex. Structure 2008, 16, 1732–1739. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Fonseca, C.; Stumpff, J. A unique kinesin-8 surface loop provides specificity for chromosome alignment. Mol. Biol. Cell 2014, 25, 3319–3329. [Google Scholar] [CrossRef] [PubMed]
- Moores, C.A.; Cooper, J.; Wagenbach, M.; Ovechkina, Y.; Wordeman, L.; Milligan, R.A. The role of the kinesin-13 neck in microtubule depolymerization. Cell Cycle 2006, 5, 1812–1815. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, L.S. Molecular motors: From one motor many tails to one motor many tales. Trends Cell Biol. 2001, 11, 477–482. [Google Scholar] [CrossRef]
- Verhey, K.J.; Hammond, J.W. Traffic control: Regulation of kinesin motors. Nat. Rev. Mol. Cell Biol. 2009, 10, 765–777. [Google Scholar] [CrossRef]
- Gatt, M.K.; Savoian, M.S.; Riparbelli, M.G.; Massarelli, C.; Callaini, G.; Glover, D.M. Klp67A destabilises pre-anaphase microtubules but subsequently is required to stabilise the central spindle. J. Cell Sci. 2005, 118, 2671–2682. [Google Scholar] [CrossRef]
- Wang, H.; Brust-Mascher, I.; Cheerambathur, D.; Scholey, J.M. Coupling between microtubule sliding, plus-end growth and spindle length revealed by kinesin-8 depletion. Cytoskeleton 2010, 67, 715–728. [Google Scholar] [CrossRef]
- Cavazza, T.; Vernos, I. The RanGTP Pathway: From Nucleo-Cytoplasmic Transport to Spindle Assembly and Beyond. Front. Cell Dev. Biol. 2015, 3, 82. [Google Scholar] [CrossRef] [PubMed]
- Dephoure, N.; Zhou, C.; Villen, J.; Beausoleil, S.A.; Bakalarski, C.E.; Elledge, S.J.; Gygi, S.P. A quantitative atlas of mitotic phosphorylation. Proc. Natl. Acad. Sci. USA 2008, 105, 10762–10767. [Google Scholar] [CrossRef] [PubMed]
- Dave, S.; Anderson, S.J.; Sinha Roy, P.; Nsamba, E.T.; Bunning, A.R.; Fukuda, Y.; Gupta, M.L., Jr. Discrete regions of the kinesin-8 Kip3 tail differentially mediate astral microtubule stability and spindle disassembly. Mol. Biol. Cell 2018, 29, 1866–1877. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Organism | Family Member(s) |
|---|---|
| Aspergillus nidulans | KipB |
| Caenorhabditis elegans | KLP-13 |
| Drosophila melanogaster | KLP67A, Kif19A |
| Homo sapiens | Kif18A, Kif18B, Kif19 |
| Mus musculus | Kif18A, Kif18B, Kif19A |
| Saccharomyces cerevisiae | Kip3 |
| Schizosaccharomyces pombe | Klp5/Klp6 (heterodimer) |
| Xenopus laevis | Kif18A, Kif18B |
| Kip3 | Klp5/6 | Kif18A | hsKif18B | mKif19A | Klp67A | |
|---|---|---|---|---|---|---|
| Velocity (µm min−1) | 3 [31] 0.71 ± 0.23 [33] 3.2 ± 0.3 [72] | 2.3 ± 0.2 [73] Klp5: 0.4 ± 0.22 * Klp6: 5.22 ± 1.08 * [74] | 11.9 ± 2.3 [68] 18.6 ± 5.4 [70] | 3.12 ± 0.18 [71] 20.9 ± 0.4 [45] | 1.3 ± 0.18 [37] | 3 ± 1.2 [75] |
| Run Length (µm) | 12.4 ± 2.3 [31] 11 ± 2 [72] | 7.2 ± 5.9 [73] | 9.4 ± 5.7 [68] 10.1 ± 4.6 [70] | 0.74 ± 0.22 [71] >7 [45] | N.D. | N.D. |
| End Dwell Time (s) | 36 ± 4 [72] 38.2 ± 6 [76] | Klp6: 42 ± 24* [74] | ~55 [69] | 1.42 ± 0.57 [71] 22.8 [45] | N.D. | N.D. |
| Depolymerization Rate (µm min−1) | ≤2 [31] 0.06 [33] 2.5–4 [72] | N.S. [73,74] | 0.052 ± 0.026 ** [77] N.S. [78] 0.21 ± 0.08–1.25 ± 0.14 [32] | N.S. [45] | 1.07 ± 0.23 [37] | N.D. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shrestha, S.; Hazelbaker, M.; Yount, A.L.; Walczak, C.E. Emerging Insights into the Function of Kinesin-8 Proteins in Microtubule Length Regulation. Biomolecules 2019, 9, 1. https://doi.org/10.3390/biom9010001
Shrestha S, Hazelbaker M, Yount AL, Walczak CE. Emerging Insights into the Function of Kinesin-8 Proteins in Microtubule Length Regulation. Biomolecules. 2019; 9(1):1. https://doi.org/10.3390/biom9010001
Chicago/Turabian StyleShrestha, Sanjay, Mark Hazelbaker, Amber L. Yount, and Claire E. Walczak. 2019. "Emerging Insights into the Function of Kinesin-8 Proteins in Microtubule Length Regulation" Biomolecules 9, no. 1: 1. https://doi.org/10.3390/biom9010001
APA StyleShrestha, S., Hazelbaker, M., Yount, A. L., & Walczak, C. E. (2019). Emerging Insights into the Function of Kinesin-8 Proteins in Microtubule Length Regulation. Biomolecules, 9(1), 1. https://doi.org/10.3390/biom9010001