Distribution of Glycerophospholipids in the Adult Human Lens

{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Lenses
2.3. Matrix-Assisted Laser Desorption Ionisation Mass Spectrometry
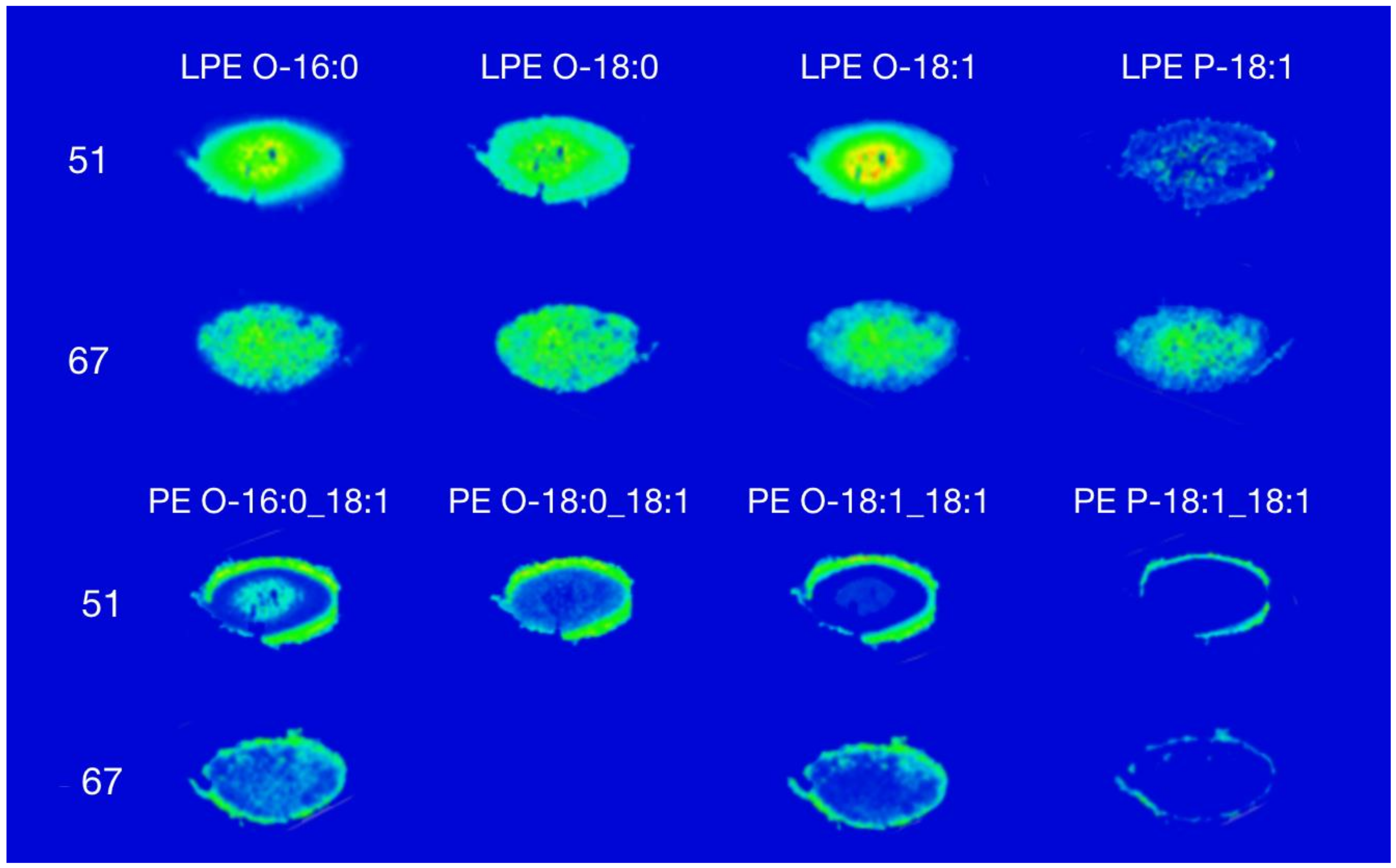
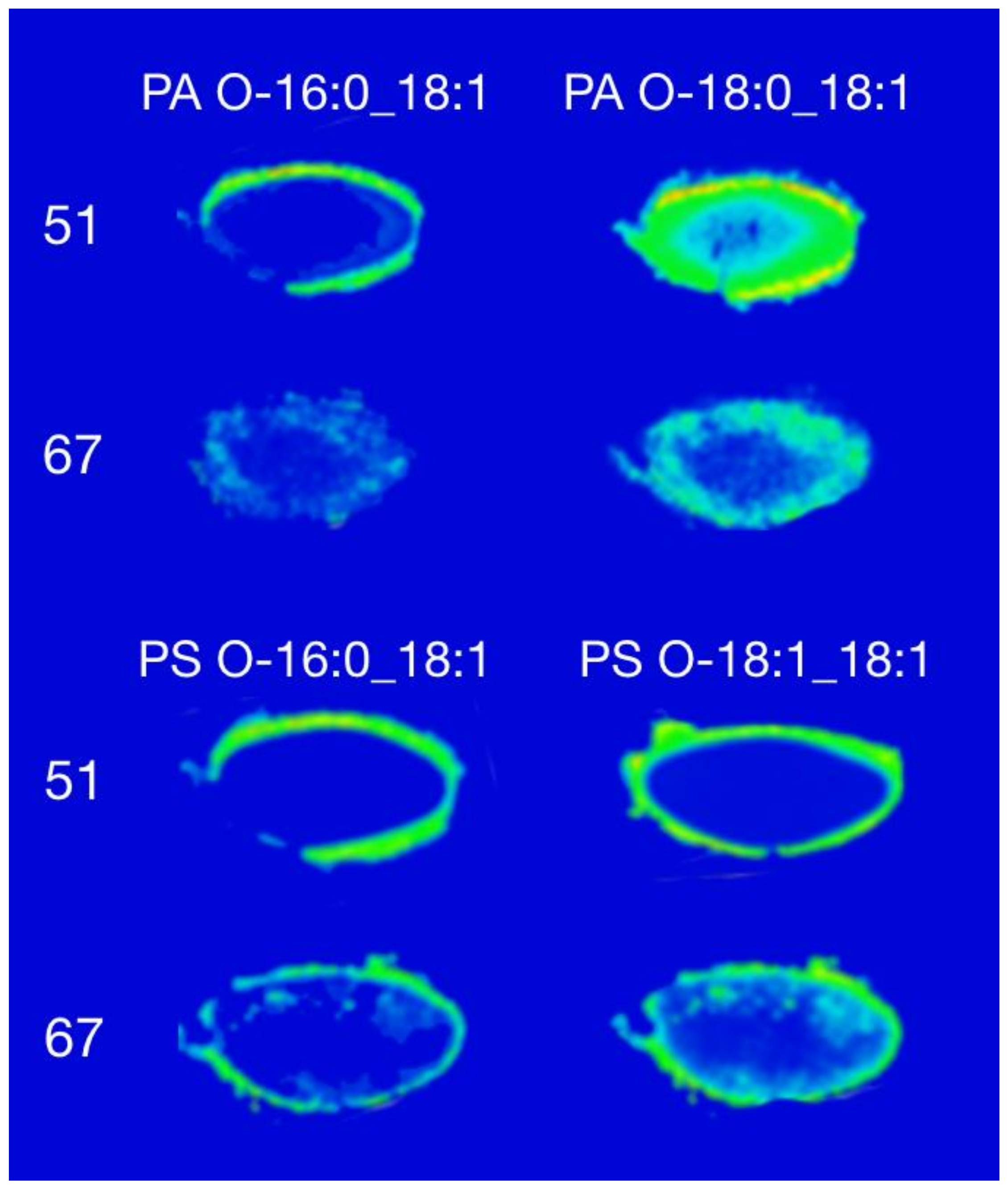
3. Results and Discussion
3.1. Phosphatidylethanolamine
3.2. Phosphatidic Acid and Phosphatidylserine
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Harding, J. Biochemistry of the Eye; Taylor & Francis: Abingdon, UK, 1998. [Google Scholar]
- Heys, K.R.; Crams, S.L.; Truscott, R.J.W. Massive increase in the stiffness of the human lens nucleus with age: the basis for presbyopia? Mol. Vis. 2004, 10, 956–963. [Google Scholar] [PubMed]
- Hughes, J.R.; Levchenko, V.A.; Blanksby, S.J.; Mitchell, T.W.; Williams, A.; Truscott, R.J.W. No turnover in lens lipids for the entire human lifespan. eLife 2015, 4, e06003. [Google Scholar] [CrossRef] [PubMed]
- Wride, M.A. Lens fibre cell differentiation and organelle loss: Many paths lead to clarity. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1219–1233. [Google Scholar] [CrossRef] [PubMed]
- Borchman, D.; Yappert, M.C. Lipids and the ocular lens. J. Lipid Res. 2010, 51, 2473–2488. [Google Scholar] [CrossRef] [PubMed]
- Berman, E.R. Biochemistry of the Eye; Plenum Press: New York, NY, USA, 1991. [Google Scholar]
- Deeley, J.M.; Mitchell, T.W.; Wei, X.; Korth, J.; Nealon, J.R.; Blanksby, S.J.; Truscott, R.J.W. Human lens lipids differ markedly from those of commonly used experimental animals. Biochim. Biophys. Acta (BBA) Mol. Biol. Lipids 2008, 1781, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Seng, J.A.; Ellis, S.R.; Hughes, J.R.; Maccarone, A.T.; Truscott, R.J.W.; Blanksby, S.J.; Mitchell, T.W. Characterisation of sphingolipids in the human lens by thin layer chromatography–desorption electrospray ionisation mass spectrometry. Biochim. Biophys. Acta (BBA) Mol. Biol. Lipids 2014, 1841, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Estrada, R.; Puppato, A.; Borchman, D.; Yappert, M.C. Reevaluation of the phospholipid composition in membranes of adult human lenses by 31P NMR and MALDI MS. Biochim. Biophys. Acta (BBA) Biomembr. 2010, 1798, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Deeley, J.M.; Thomas, M.C.; Truscott, R.J.W.; Mitchell, T.W.; Blanksby, S.J. Identification of abundant alkyl ether glycerophospholipids in the human lens by tandem mass spectrometry techniques. Anal. Chem. 2009, 81, 1920–1930. [Google Scholar] [CrossRef] [PubMed]
- Rodemer, C.; Thai, T.-P.; Brugger, B.; Kaercher, T.; Werner, H.; Nave, K.-A.; Wieland, F.; Gorgas, K.; Just, W.W. Inactivation of ether lipid biosynthesis causes male infertility, defects in eye development and optic nerve hypoplasia in mice. Hum. Mol. Genet. 2003, 12, 1881–1895. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Deeley, J.; Blanksby, S.; Leisch, F.; Ellis, S.; Truscott, R.; Mitchell, T. Instability of the cellular lipidome with age. Age 2012, 34, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Grami, V.; Marrero, Y.; Tang, D.; Yappert, M.C.; Rasi, V.; Borchman, D. Human lens phospholipid changes with age and cataract. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1682–1689. [Google Scholar] [CrossRef] [PubMed]
- Ellis, S.R.; Wu, C.; Deeley, J.M.; Zhu, X.; Truscott, R.J.W.; Panhuis, M.; Cooks, R.G.; Mitchell, T.W.; Blanksby, S.J. Imaging of human lens lipids by desorption electrospray ionisation mass spectrometry. J. Am. Soc. Mass Spectrom. 2010, 21, 2095–2104. [Google Scholar] [CrossRef] [PubMed]
- Deeley, J.M.; Hankin, J.A.; Friedrich, M.G.; Murphy, R.C.; Truscott, R.J.W.; Mitchell, T.W.; Blanksby, S.J. Sphingolipid distribution changes with age in the human lens. J. Lipid Res. 2010, 51, 2753–2760. [Google Scholar] [CrossRef] [PubMed]
- Borchman, D.; Byrdwell, W.C.; Yappert, M.C. Regional and age-dependent differences in the phospholipid composition of human lens membranes. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3938–3942. [Google Scholar]
- Byrdwell, W.C. Dual parallel mass spectrometers for analysis of sphingolipid, glycerophospholipid and plasmalogen molecular species. Rapid Commun. Mass Spectrom. 1998, 12, 256–272. [Google Scholar] [CrossRef]
- Liebisch, G.; Vizcaíno, J.A.; Köfeler, H.; Trötzmüller, M.; Griffiths, W.J.; Schmitz, G.; Spener, F.; Wakelam, M.J.O. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res. 2013, 54, 1523–1530. [Google Scholar] [CrossRef] [PubMed]
- Elliott, K.; FitzSimons, D.W. The Human Lens—In Relation to Cataract; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Chen, Y.-L.; Montedonico, A.E.; Kauffman, S.; Dunlap, J.R.; Menn, F.-M.; Reynolds, T.B. Phosphatidylserine synthase and phosphatidylserine decarboxylase are essential for cell wall integrity and virulence in Candida albicans. Mol. Microbiol. 2010, 75, 1112–1132. [Google Scholar] [CrossRef] [PubMed]
- Cotlier, E.; Obara, Y.; Toftness, B. Cholesterol and phospholipids in protein fractions of human lens and senile cataract. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1978, 530, 267–278. [Google Scholar] [CrossRef]
- Oborina, E.M.; Yappert, M.C. Effect of sphingomyelin versus dipalmitoylphosphatidylcholine on the extent of lipid oxidation. Chem. Phys. Lipids 2003, 123, 223–232. [Google Scholar] [CrossRef]
- Grey, A.C.; Schey, K.L. Age-related changes in the spatial distribution of human lens α-crystallin products by MALDI imaging mass spectrometry. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4319–4329. [Google Scholar] [CrossRef] [PubMed]
- Grey, A.C.; Chaurand, P.; Caprioli, R.M.; Schey, K.L. MALDI imaging mass spectrometry of integral membrane proteins from ocular lens and retinal tissue. J. Proteome Res. 2009, 8, 3278–3283. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seng, J.A.; Nealon, J.R.; Blanksby, S.J.; Mitchell, T.W. Distribution of Glycerophospholipids in the Adult Human Lens. Biomolecules 2018, 8, 156. https://doi.org/10.3390/biom8040156
Seng JA, Nealon JR, Blanksby SJ, Mitchell TW. Distribution of Glycerophospholipids in the Adult Human Lens. Biomolecules. 2018; 8(4):156. https://doi.org/10.3390/biom8040156
Chicago/Turabian StyleSeng, Jo Ann, Jessica R. Nealon, Stephen J. Blanksby, and Todd W. Mitchell. 2018. "Distribution of Glycerophospholipids in the Adult Human Lens" Biomolecules 8, no. 4: 156. https://doi.org/10.3390/biom8040156
APA StyleSeng, J. A., Nealon, J. R., Blanksby, S. J., & Mitchell, T. W. (2018). Distribution of Glycerophospholipids in the Adult Human Lens. Biomolecules, 8(4), 156. https://doi.org/10.3390/biom8040156