Calcium-Binding Proteins with Disordered Structure and Their Role in Secretion, Storage, and Cellular Signaling

Abstract
1. Calcium Signaling in the Cell
2. Calcium-Binding Proteins
3. The Regulatory Potential of Intrinsically Disordered Proteins
4. Calcium-Binding IDPs
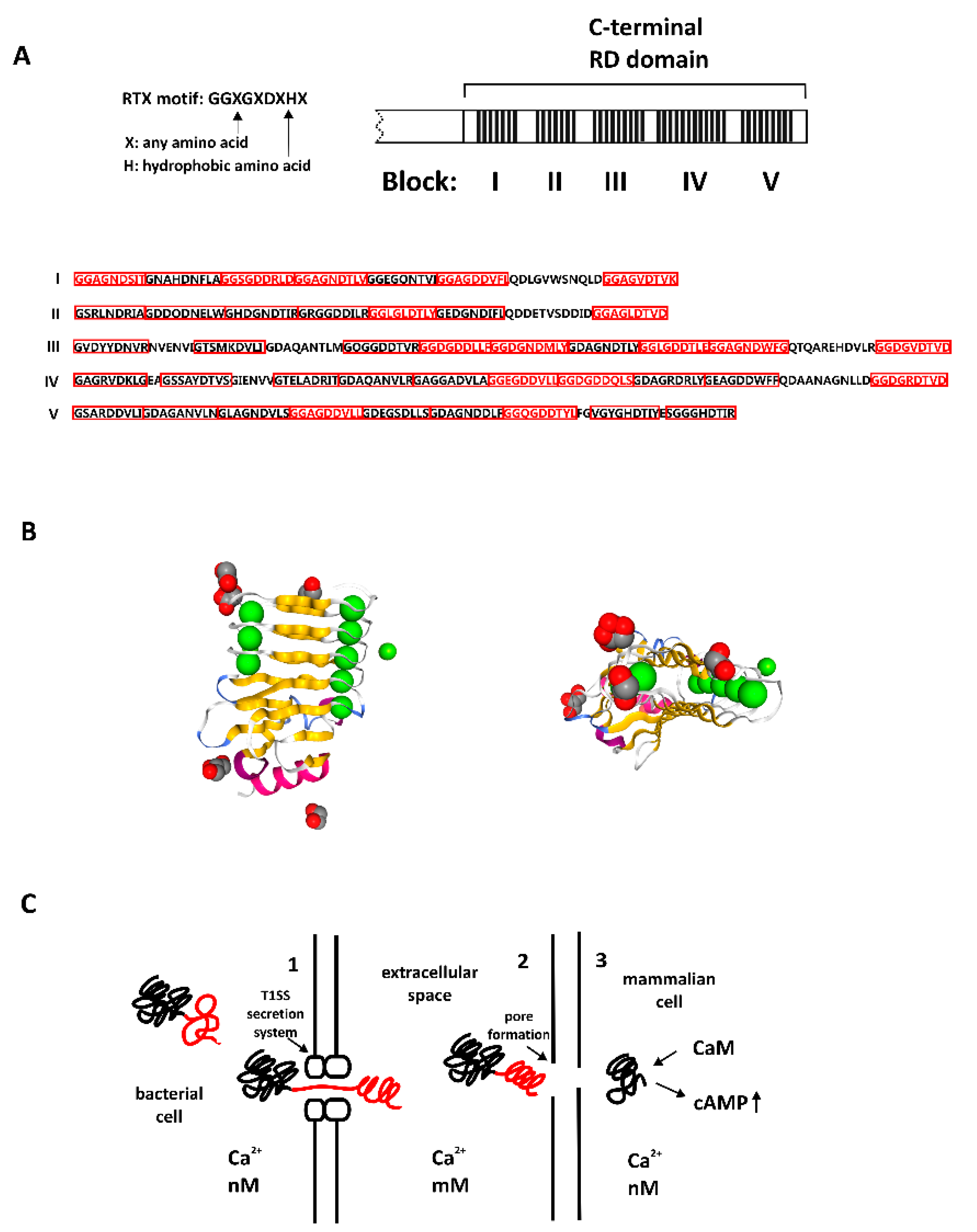
4.1. Bacterial Calcium-Binding Repeat-in-Toxin Motifs
4.2. Disordered Calcium-Binding IDPs in Biomineralization
4.2.1. Bone and Dentin Biomineralization
4.2.2. Fish Otolith Formation
4.2.3. The Role of Calcium in Biomineralization and Disorder
4.3. Calmodulin-Binding IDPs
4.4. Calcium Sequestering Proteins of the Reticulum
4.5. Other Calcium-Binding IDPs
5. Conclusions
- Ca2+ concentration in the cytoplasm is too low/calcium spike is too sharp and rapid (nevertheless such potential obstacles do not hinder calmodulin calcium-sensing).
- No secondary structure formed upon calcium binding in the cytoplasm can be as effective as the EF-hand motif (energetically unfavorable?).
- After Ca2+ binding and folding, how would the message be propagated?
- The Ca2+ dependent IDP sensor would have to bind too many protein targets; a universal calcium sensor (calmodulin) binding to many IDPs works better.
- There are presumably unknown calcium-binding IDPs.
- There are more examples of transient disorder; they may be difficult to spot and cannot be easily predicted from the sequence.
Funding
Conflicts of Interest
References
- Zhou, Y.; Xue, S.; Yang, J.J. Calciomics: Integrative studies of Ca2+-binding proteins and their interactomes in biological systems. Metallomics 2013, 5, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Hepler, P.K. The cytoskeleton and its regulation by calcium and protons. Plant Physiol. 2016, 170, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Balaji, R.; Bielmeier, C.; Harz, H.; Bates, J.; Stadler, C.; Hildebrand, A.; Classen, A.K. Calcium spikes, waves and oscillations in a large, patterned epithelial tissue. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Yanez, M.; Gil-Longo, J.; Campos-Toimil, M. Calcium binding proteins. Adv. Exp. Med. Biol. 2012, 740, 461–482. [Google Scholar] [PubMed]
- Kirberger, M.; Wang, X.; Deng, H.; Yang, W.; Chen, G.; Yang, J.J. Statistical analysis of structural characteristics of protein Ca2+-binding sites. J. Biol. Inorg. Chem. 2008, 13, 1169–1181. [Google Scholar] [CrossRef] [PubMed]
- Dunker, A.K.; Silman, I.; Uversky, V.N.; Sussman, J.L. Function and structure of inherently disordered proteins. Curr. Opin. Struct. Biol. 2008, 18, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Tompa, P. Intrinsically unstructured proteins. Trends Biochem. Sci. 2002, 27, 527–533. [Google Scholar] [CrossRef]
- Wright, P.E.; Dyson, H.J. Intrinsically disordered proteins in cellular signalling and regulation. Nat. Rev. Mol. Cell Biol. 2015, 16, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.J.; Sodhi, J.S.; McGuffin, L.J.; Buxton, B.F.; Jones, D.T. Prediction and functional analysis of native disorder in proteins from the three kingdoms of life. J. Mol. Biol. 2004, 337, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Dunker, A.K.; Oldfield, C.J.; Meng, J.; Romero, P.; Yang, J.Y.; Chen, J.W.; Vacic, V.; Obradovic, Z.; Uversky, V.N. The unfoldomics decade: An update on intrinsically disordered proteins. BMC Genom. 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Querfurth, C.; Diernfellner, A.C.; Gin, E.; Malzahn, E.; Hofer, T.; Brunner, M. Circadian conformational change of the neurospora clock protein frequency triggered by clustered hyperphosphorylation of a basic domain. Mol. Cell 2011, 43, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Mittag, T.; Marsh, J.; Grishaev, A.; Orlicky, S.; Lin, H.; Sicheri, F.; Tyers, M.; Forman-Kay, J.D. Structure/function implications in a dynamic complex of the intrinsically disordered Sic1 with the Cdc4 subunit of an SCF ubiquitin ligase. Structure 2010, 18, 494–506. [Google Scholar] [CrossRef] [PubMed]
- Lyons, N.A.; Fonslow, B.R.; Diedrich, J.K.; Yates, J.R., 3rd; Morgan, D.O. Sequential primed kinases create a damage-responsive phosphodegron on Eco1. Nat. Struct. Mol. Biol. 2013, 20, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Sotomayor-Perez, A.C.; Ladant, D.; Chenal, A. Disorder-to-order transition in the CyaA toxin RTX domain: Implications for toxin secretion. Toxins 2014, 7, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Linhartova, I.; Bumba, L.; Masin, J.; Basler, M.; Osicka, R.; Kamanova, J.; Prochazkova, K.; Adkins, I.; Hejnova-Holubova, J.; Sadilkova, L.; et al. RTX proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef] [PubMed]
- Sotomayor-Perez, A.C.; Ladant, D.; Chenal, A. Calcium-induced folding of intrinsically disordered repeat-in-toxin (RTX) motifs via changes of protein charges and oligomerization states. J. Biol. Chem. 2011, 286, 16997–17004. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.S.; Hildebrand, P.W. NGL viewer: A web application for molecular visualization. Nucleic Acids Res. 2015, 43, W576–W579. [Google Scholar] [CrossRef] [PubMed]
- Kalmar, L.; Homola, D.; Varga, G.; Tompa, P. Structural disorder in proteins brings order to crystal growth in biomineralization. Bone 2012, 51, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Boskey, A.L.; Villarreal-Ramirez, E. Intrinsically disordered proteins and biomineralization. Matrix Biol. 2016, 52–54, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Golub, E.E. Biomineralization and matrix vesicles in biology and pathology. Semin. Immunopathol. 2011, 33, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Veis, A.; Dorvee, J.R. Biomineralization mechanisms: A new paradigm for crystal nucleation in organic matrices. Calcif. Tissue Int. 2013, 93, 307–315. [Google Scholar] [CrossRef] [PubMed]
- George, A.; Veis, A. Phosphorylated proteins and control over apatite nucleation, crystal growth, and inhibition. Chem. Rev. 2008, 108, 4670–4693. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Baba, O.; Butler, W.T. Post-translational modifications of sibling proteins and their roles in osteogenesis and dentinogenesis. Crit. Rev. Oral Biol. Med. 2004, 15, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Sollner, C.; Burghammer, M.; Busch-Nentwich, E.; Berger, J.; Schwarz, H.; Riekel, C.; Nicolson, T. Control of crystal size and lattice formation by starmaker in otolith biomineralization. Science 2003, 302, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Kaplon, T.M.; Rymarczyk, G.; Nocula-Lugowska, M.; Jakob, M.; Kochman, M.; Lisowski, M.; Szewczuk, Z.; Ozyhar, A. Starmaker exhibits properties of an intrinsically disordered protein. Biomacromolecules 2008, 9, 2118–2125. [Google Scholar] [CrossRef] [PubMed]
- Poznar, M.; Holubowicz, R.; Wojtas, M.; Gapinski, J.; Banachowicz, E.; Patkowski, A.; Ozyhar, A.; Dobryszycki, P. Structural properties of the intrinsically disordered, multiple calcium ion-binding otolith matrix macromolecule-64 (OMM-64). Biochim. Biophys. Acta 2017, 1865, 1358–1371. [Google Scholar] [CrossRef] [PubMed]
- Wojtas, M.; Holubowicz, R.; Poznar, M.; Maciejewska, M.; Ozyhar, A.; Dobryszycki, P. Calcium ion binding properties and the effect of phosphorylation on the intrinsically disordered starmaker protein. Biochemistry 2015, 54, 6525–6534. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.; Carver, J.A.; Ecroyd, H.; Thorn, D.C. Invited review: Caseins and the casein micelle: Their biological functions, structures, and behavior in foods. J. Dairy Sci. 2013, 96, 6127–6146. [Google Scholar] [CrossRef] [PubMed]
- Tercinier, L.; Ye, A.; Anema, S.G.; Singh, A.; Singh, H. Interactions of casein micelles with calcium phosphate particles. J. Agric. Food Chem. 2014, 62, 5983–5992. [Google Scholar] [CrossRef] [PubMed]
- Maximciuc, A.A.; Putkey, J.A.; Shamoo, Y.; Mackenzie, K.R. Complex of calmodulin with a ryanodine receptor target reveals a novel, flexible binding mode. Structure 2006, 14, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Salarian, M.; Chen, Y.; Veenstra, R.; Louis, C.F.; Yang, J.J. Gap junction regulation by calmodulin. FEBS Lett. 2014, 588, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Permyakov, S.E.; Permyakov, E.A.; Uversky, V.N. Intrinsically disordered caldesmon binds calmodulin via the “buttons on a string” mechanism. PeerJ 2015, 3, e1265. [Google Scholar] [CrossRef] [PubMed]
- Kleerekoper, Q.K.; Putkey, J.A. PEP-19, an intrinsically disordered regulator of calmodulin signaling. J. Biol. Chem. 2009, 284, 7455–7464. [Google Scholar] [CrossRef] [PubMed]
- Radivojac, P.; Vucetic, S.; O’Connor, T.R.; Uversky, V.N.; Obradovic, Z.; Dunker, A.K. Calmodulin signaling: Analysis and prediction of a disorder-dependent molecular recognition. Proteins 2006, 63, 398–410. [Google Scholar] [CrossRef] [PubMed]
- Creamer, T.P. Transient disorder: Calcineurin as an example. Intrinsically Disord. Proteins 2013, 1, e26412. [Google Scholar] [CrossRef] [PubMed]
- Bal, N.C.; Jena, N.; Chakravarty, H.; Kumar, A.; Chi, M.; Balaraju, T.; Rawale, S.V.; Rawale, J.S.; Sharon, A.; Periasamy, M. The C-terminal calcium-sensitive disordered motifs regulate isoform-specific polymerization characteristics of calsequestrin. Biopolymers 2015, 103, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Park, I.Y.; Kim, E.; Youn, B.; Fields, K.; Dunker, A.K.; Kang, C. Comparing skeletal and cardiac calsequestrin structures and their calcium binding: A proposed mechanism for coupled calcium binding and protein polymerization. J. Biol. Chem. 2004, 279, 18026–18033. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Corbett, E.F.; Mesaeli, N.; Nakamura, K.; Opas, M. Calreticulin: One protein, one gene, many functions. Biochem. J. 1999, 344, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Shivarov, V.; Ivanova, M.; Tiu, R.V. Mutated calreticulin retains structurally disordered C terminus that cannot bind Ca2+: Some mechanistic and therapeutic implications. Blood Cancer J. 2014, 4, e185. [Google Scholar] [CrossRef] [PubMed]
- Wijeyesakere, S.J.; Gafni, A.A.; Raghavan, M. Calreticulin is a thermostable protein with distinct structural responses to different divalent cation environments. J. Biol. Chem. 2011, 286, 8771–8785. [Google Scholar] [CrossRef] [PubMed]
- Hanin, M.; Brini, F.; Ebel, C.; Toda, Y.; Takeda, S.; Masmoudi, K. Plant dehydrins and stress tolerance: Versatile proteins for complex mechanisms. Plant Signal. Behav. 2011, 6, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Heyen, B.J.; Alsheikh, M.K.; Smith, E.A.; Torvik, C.F.; Seals, D.F.; Randall, S.K. The calcium-binding activity of a vacuole-associated, dehydrin-like protein is regulated by phosphorylation. Plant Physiol. 2002, 130, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N. Unreported intrinsic disorder in proteins: Building connections to the literature on IDPs. Intrinsically Disord. Proteins 2014, 2, e970499. [Google Scholar] [CrossRef] [PubMed]
- Balcerak, A.; Rowinski, S.; Szafron, L.M.; Grzybowska, E.A. The calcium binding properties and structure prediction of the Hax-1 protein. Acta Biochim. Pol. 2017, 64, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Hirasaka, K.; Mills, E.M.; Haruna, M.; Bando, A.; Ikeda, C.; Abe, T.; Kohno, S.; Nowinski, S.M.; Lago, C.U.; Akagi, K.; et al. UCP3 is associated with Hax-1 in mitochondria in the presence of calcium ion. Biochem. Biophys. Res. Commun. 2016, 472, 108–113. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Protein Symbol | Protein Name | Organism | Function | Swiss Prot ID |
|---|---|---|---|---|
| OPN (SSP1) | Osteopontin, secreted phosphoprotein 1 | Homo sapiens | Matrix mineralization, immunity | P10451 |
| DMP1 | Dentin matrix protein 1 | Homo sapiens | Osteoblast differentiation, nucleation of hydroxyapatite | Q13316 |
| MEPE | Matrix extracellular phosphoglycoprotein | Homo sapiens | Phosphate excretion, mineralization | Q9NQ76 |
| BSP | Bone sialoprotein | Homo sapiens | Matrix mineralization, hydroxyapatite binding, cell attachment | P21815 |
| DSPP | Dentin sialophosphoprotein | Homo sapiens | Dentinogenesis | Q9NZW4 |
| AMELX | Amylogenin | Homo sapiens | Enamel formation; extracellular scaffold, crystal growth | Q99217 |
| Stm | Starmaker | Danio rerio (zebrafish) | Otolith formation | A2VD23 |
| OMM-64 | Otolith matrix macromolecule-64 | Oncorhynchus mykiss (rainbow trout) | Otolith formation | B1Q2L9 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzybowska, E.A. Calcium-Binding Proteins with Disordered Structure and Their Role in Secretion, Storage, and Cellular Signaling. Biomolecules 2018, 8, 42. https://doi.org/10.3390/biom8020042
Grzybowska EA. Calcium-Binding Proteins with Disordered Structure and Their Role in Secretion, Storage, and Cellular Signaling. Biomolecules. 2018; 8(2):42. https://doi.org/10.3390/biom8020042
Chicago/Turabian StyleGrzybowska, Ewa A. 2018. "Calcium-Binding Proteins with Disordered Structure and Their Role in Secretion, Storage, and Cellular Signaling" Biomolecules 8, no. 2: 42. https://doi.org/10.3390/biom8020042
APA StyleGrzybowska, E. A. (2018). Calcium-Binding Proteins with Disordered Structure and Their Role in Secretion, Storage, and Cellular Signaling. Biomolecules, 8(2), 42. https://doi.org/10.3390/biom8020042