Protein–Phospholipid Interaction Motifs: A Focus on Phosphatidic Acid


Abstract
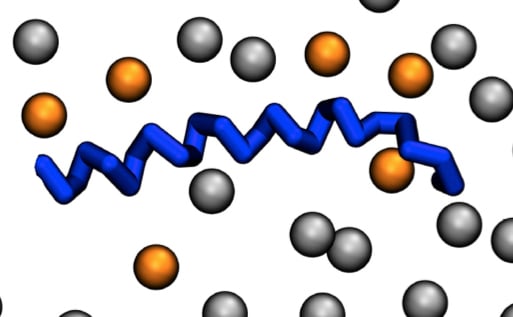
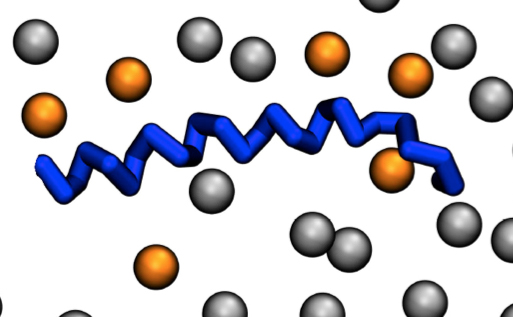{kind=link}
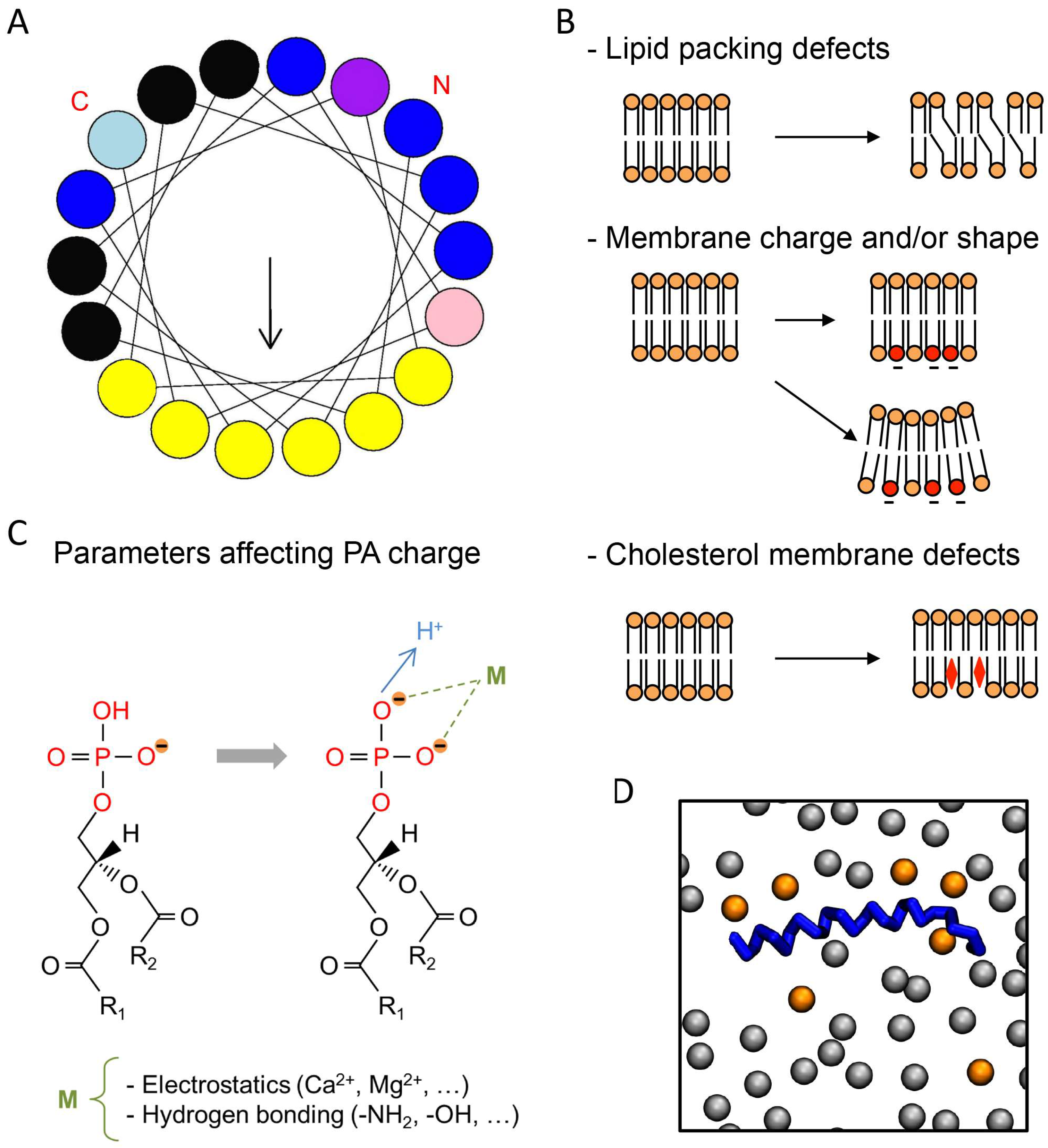{kind=link}
1. Introduction
2. Phosphatidic Acid-Binding Modules
3. Effect of Membrane Topology and Environment on Phosphatidic Acid Binding
4. Effect of Membrane Charge on Phosphatidic Acid Binding
5. Effect of Phosphatidic Acid Fatty Acyl Chain Composition
6. Effect of pH on Phosphatidic Acid Binding
7. Conclusions
Acknowledgments
Author Contributions
Conflict of Interests
References
- Arora, A.; Tamm, L.K. Biophysical approaches to membrane protein structure determination. Curr. Opin. Struct. Biol. 2001, 11, 540–547. [Google Scholar] [CrossRef]
- Basso, L.G.M.; Mendes, L.F.S.; Costa-Filho, A.J. The two sides of a lipid-protein story. Biophys. Rev. 2016, 8, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, S.; Shekhar, P.; Heinrich, F.; Daou, M.; Gericke, A.; Ross, A.H.; Lösche, M. Membrane association of the PTEN tumor suppressor: Molecular details of the protein–membrane complex from SPR binding studies and neutron reflection. PLoS ONE 2012, 7, e32591. [Google Scholar] [CrossRef] [PubMed]
- Raja, M.; Spelbrink, R.E.J.; de Kruijff, B.; Killian, J.A. Phosphatidic acid plays a special role in stabilizing and folding of the tetrameric potassium channel KcsA. FEBS Lett. 2007, 581, 5715–5722. [Google Scholar] [CrossRef] [PubMed]
- Ammar, M.R.; Kassas, N.; Bader, M.F.; Vitale, N. Phosphatidic acid in neuronal development: A node for membrane and cytoskeleton rearrangements. Biochimie 2014, 107, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Ammar, M.R.; Kassas, N.; Chasserot-Golaz, S.; Bader, M.F.; Vitale, N. Lipids in Regulated Exocytosis: What are They Doing? Front. Endocrinol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Stace, C.L.; Ktistakis, N.T. Phosphatidic acid- and phosphatidylserine-binding proteins. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2006, 1761, 913–926. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Lee, C.S.; Hwang, D.; Ryu, S.H. Understanding of the roles of phospholipase D and phosphatidic acid through their binding partners. Prog. Lipid Res. 2012, 51, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, H.; de los Santos, P.; Neiman, A.M. Positive and negative regulation of a SNARE protein by control of intracellular localization. Mol. Biol. Cell 2004, 15, 1802–1815. [Google Scholar] [CrossRef] [PubMed]
- Zeniou-Meyer, M.; Zabari, N.; Ashery, U.; Chasserot-Golaz, S.; Haeberlé, A.M.; Demais, V.; Bailly, Y.; Gottfried, I.; Nakanishi, H.; Neiman, A.M.; et al. Phospholipase D1 production of phosphatidic acid at the plasma membrane promotes exocytosis of large dense-core granules at a late stage. J. Biol. Chem. 2007, 282, 21746–21757. [Google Scholar] [CrossRef] [PubMed]
- Kassas, N.; Tryoen-Tóth, P.; Corrotte, M.; Thahouly, T.; Bader, M.F.; Grant, N.J.; Vitale, N. Genetically encoded probes for phosphatidic acid. Methods Cell Biol. 2012, 108, 445–459. [Google Scholar] [PubMed]
- Loewen, C.J.; Gaspar, M.L.; Jesch, S.A.; Delon, C.; Ktistakis, N.T.; Henry, S.A.; Levine, T.P. Phospholipid metabolism regulated by a transcription factor sensing phosphatidic acid. Science 2004, 304, 1644–1647. [Google Scholar] [CrossRef] [PubMed]
- Baillie, G.S.; Huston, E.; Scotland, G.; Hodgkin, M.; Gall, I.; Peden, A.H.; MacKenzie, C.; Houslay, E.S.; Currie, R.; Pettitt, T.R.; et al. TAPAS-1, a novel microdomain within the unique N-terminal region of the PDE4A1 cAMP-specific phosphodiesterase that allows rapid, Ca2+-triggered membrane association with selectivity for interaction with phosphatidic acid. J. Biol. Chem. 2002, 277, 28298–282309. [Google Scholar] [CrossRef] [PubMed]
- Kassas, N.; Tanguy, E.; Thahouly, T.; Fouillen, L.; Heintz, D.; Chasserot-Golaz, S.; Bader, M.F.; Grant, N.J.; Vitale, N. Comparative Characterization of Phosphatidic Acid Sensors and Their Localization during Frustrated Phagocytosis. J. Biol. Chem. 2017, 292, 4266–4279. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Anderson, N.A.; Travesset, A.; Vaknin, D. Regulation of the electric charge in phosphatidic acid domains. J. Phys. Chem. B 2012, 116, 7213–7220. [Google Scholar] [CrossRef] [PubMed]
- Potocký, M.; Pleskot, R.; Pejchar, P.; Vitale, N.; Kost, B.; Zárský, V. Live-cell imaging of phosphatidic acid dynamics in pollen tubes visualized by Spo20p-derived biosensor. New Phytol. 2014, 203, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Putta, P.; Rankenberg, J.; Korver, R.A.; van Wijk, R.; Munnik, T.; Testerink, C.; Kooijman, E.E. Phosphatidic acid binding proteins display differential binding as a function of membrane curvature stress and chemical properties. Biochim. Biophys. Acta 2016, 1858, 2709–2716. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, T.J. Hydration properties of lamellar and non-lamellar phases of phosphatidylcholine and phosphatidylethanolamine. Chem. Phys. Lipids 1996, 81, 117–131. [Google Scholar] [CrossRef]
- Horchani, H.; de Saint-Jean, M.; Barelli, H.; Antonny, B. Interaction of the Spo20 membrane-sensor motif with phosphatidic acid and other anionic lipids, and influence of the membrane environment. PLoS ONE 2014, 9, e113484. [Google Scholar] [CrossRef] [PubMed]
- Boroda, S.; Takkellapati, S.; Lawrence, R.T.; Entwisle, S.W.; Pearson, J.M.; Granade, M.E.; Mullins, G.R.; Eaton, J.M.; Villén, J.; Harris, T.E. The phosphatidic acid-binding, polybasic domain is responsible for the differences in the phosphoregulation of lipins 1 and 3. J. Biol. Chem. 2017, 292, 20481–20493. [Google Scholar] [CrossRef] [PubMed]
- Kooijman, E.E.; Carter, K.M.; van Laar, E.G.; Chupin, V.; Burger, K.N.; de Kruijff, B. What makes the bioactive lipids phosphatidic acid and lysophosphatidic acid so special? Biochemistry 2005, 44, 17007–17015. [Google Scholar] [CrossRef] [PubMed]
- Loew, S.; Kooijman, E.E.; May, S. Increased pH-sensitivity of protein binding to lipid membranes through the electrostatic-hydrogen bond switch. Chem. Phys. Lipids 2013, 169, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Faraudo, J.; Travesset, A. Phosphatidic acid domains in membranes: Effect of divalent counterions. Biophys. J. 2007, 92, 2806–2818. [Google Scholar] [CrossRef] [PubMed]
- Andreyev, A.Y.; Fahy, E.; Guan, Z.; Kelly, S.; Li, X.; McDonald, J.G.; Milne, S.; Myers, D.; Park, H.; Ryan, A.; et al. Subcellular organelle lipidomics in TLR-4-activated macrophages. J. Lipid Res. 2010, 51, 2785–2797. [Google Scholar] [CrossRef] [PubMed]
- Hammond, G.R.; Balla, T. Polyphosphoinositide binding domains: Key to inositol lipid biology. Biochim. Biophys. Acta 2015, 1851, 746–758. [Google Scholar] [CrossRef] [PubMed]
- Barneda, D.; Planas-Iglesias, J.; Gaspar, M.L.; Mohammadyani, D.; Prasannan, S.; Dormann, D.; Han, G.S.; Jesch, S.A.; Carman, G.M.; Kagan, V.; et al. The brown adipocyte protein CIDEA promotes lipid droplet fusion via a phosphatidic acid-binding amphipathic helix. Elife 2015, 26, e07485. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Itoh, K.; Yamada, T.; Cerveny, K.L.; Suzuki, T.L.; Macdonald, P.; Frohman, M.A.; Ramachandran, R.; Iijima, M.; Sesaki, H. Coincident Phosphatidic Acid Interaction Restrains Drp1 in Mitochondrial Division. Mol. Cell 2016, 63, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Iijima, M.; Sesaki, H. An unstructured loop that is critical for interactions of the stalk domain of Drp1 with saturated phosphatidic acid. Small GTPases 2017, 23, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Young, B.P.; Shin, J.J.; Orij, R.; Chao, J.T.; Li, S.C.; Guan, X.L.; Khong, A.; Jan, E.; Wenk, M.R.; Prinz, W.A.; et al. Phosphatidic acid is a pH biosensor that links membrane biogenesis to metabolism. Science 2010, 329, 1085–1088. [Google Scholar] [CrossRef] [PubMed]
- Capelluto, D.G.; Zhao, X.; Lucas, A.; Lemkul, J.A.; Xiao, S.; Fu, X.; Sun, F.; Bevan, D.R.; Finkielstein, C.V. Biophysical and molecular-dynamics studies of phosphatidic acid binding by the Dvl-2 DEP domain. Biophys J. 2014, 106, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Parry, M.J.; Alakoskela, J.M.; Khandelia, H.; Kumar, S.A.; Jaattela, M.; Mahalka, A.K.; Kinnunen, P.K. High-affinity small molecule-phospholipid complex formation: Binding of siramesine to phosphatidic acid. J. Am. Chem. Soc. 2008, 130, 12953–12960. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanguy, E.; Kassas, N.; Vitale, N. Protein–Phospholipid Interaction Motifs: A Focus on Phosphatidic Acid. Biomolecules 2018, 8, 20. https://doi.org/10.3390/biom8020020
Tanguy E, Kassas N, Vitale N. Protein–Phospholipid Interaction Motifs: A Focus on Phosphatidic Acid. Biomolecules. 2018; 8(2):20. https://doi.org/10.3390/biom8020020
Chicago/Turabian StyleTanguy, Emeline, Nawal Kassas, and Nicolas Vitale. 2018. "Protein–Phospholipid Interaction Motifs: A Focus on Phosphatidic Acid" Biomolecules 8, no. 2: 20. https://doi.org/10.3390/biom8020020
APA StyleTanguy, E., Kassas, N., & Vitale, N. (2018). Protein–Phospholipid Interaction Motifs: A Focus on Phosphatidic Acid. Biomolecules, 8(2), 20. https://doi.org/10.3390/biom8020020