
Alcohol, Aldehydes, Adducts and Airways

Abstract
:1. Introduction
2. Role of Alcohol in Lung Disease
3. Role of Cigarette Smoking in Lung Disease
4. Other Environmental Oxidants
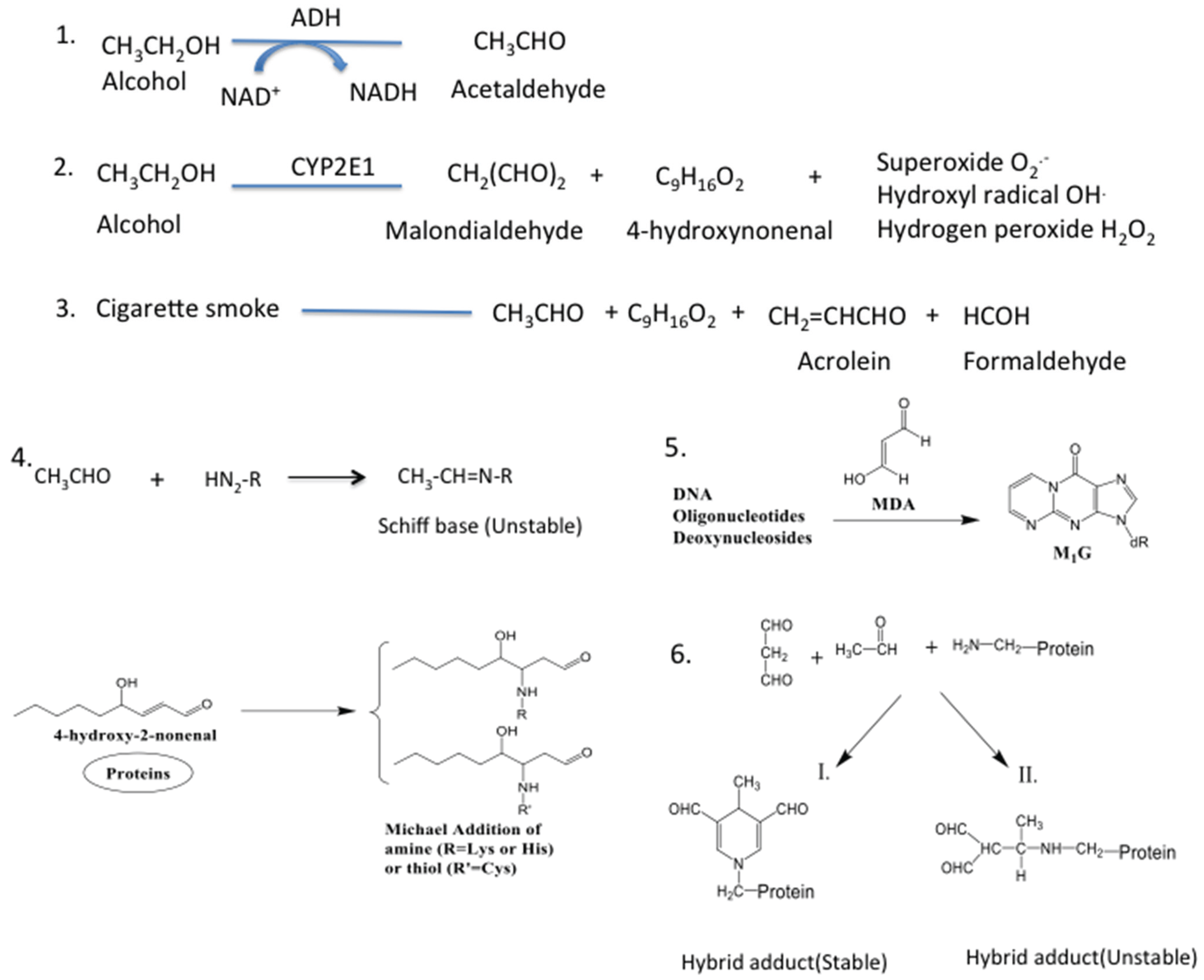
5. Source of Aldehydes in the Lung
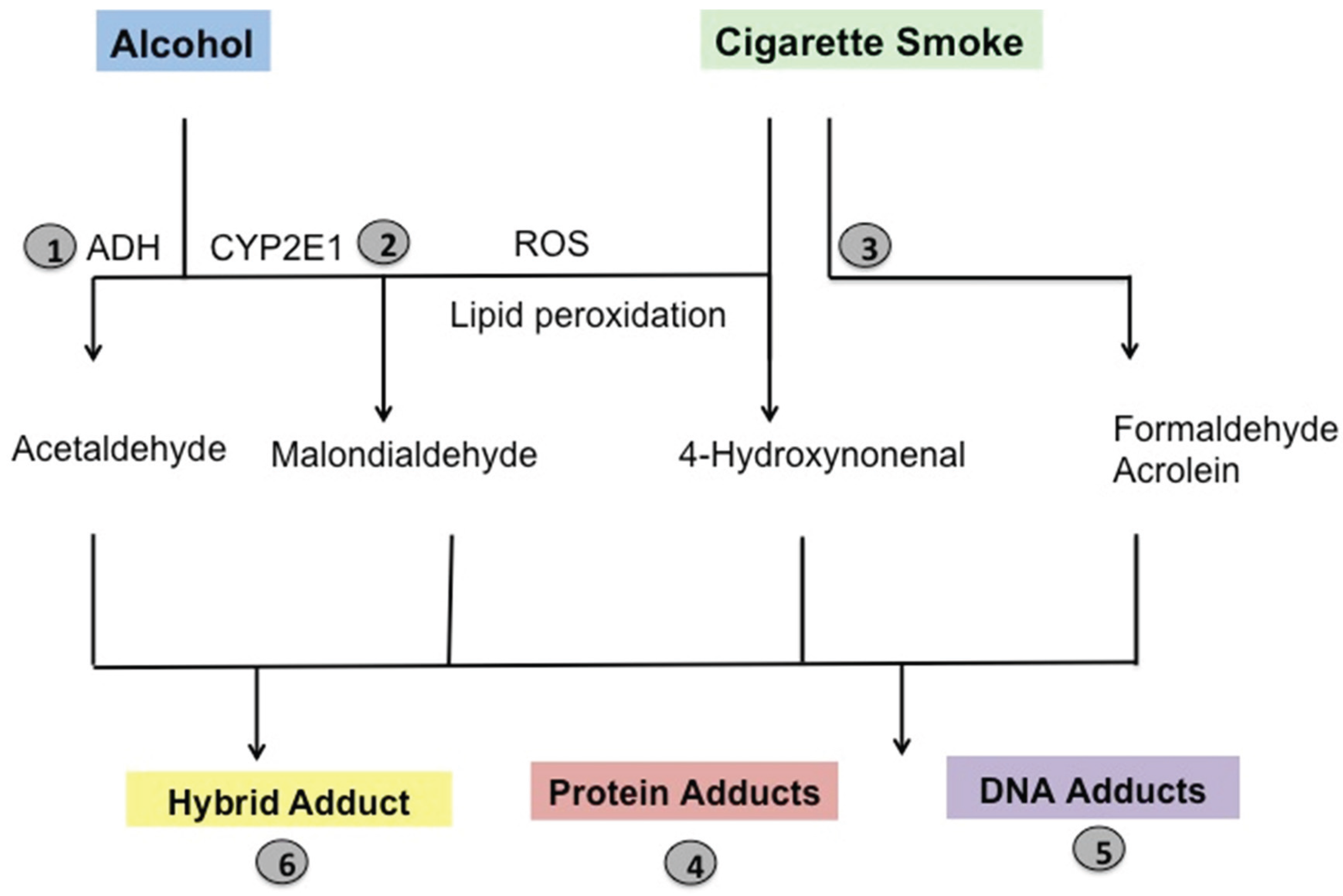
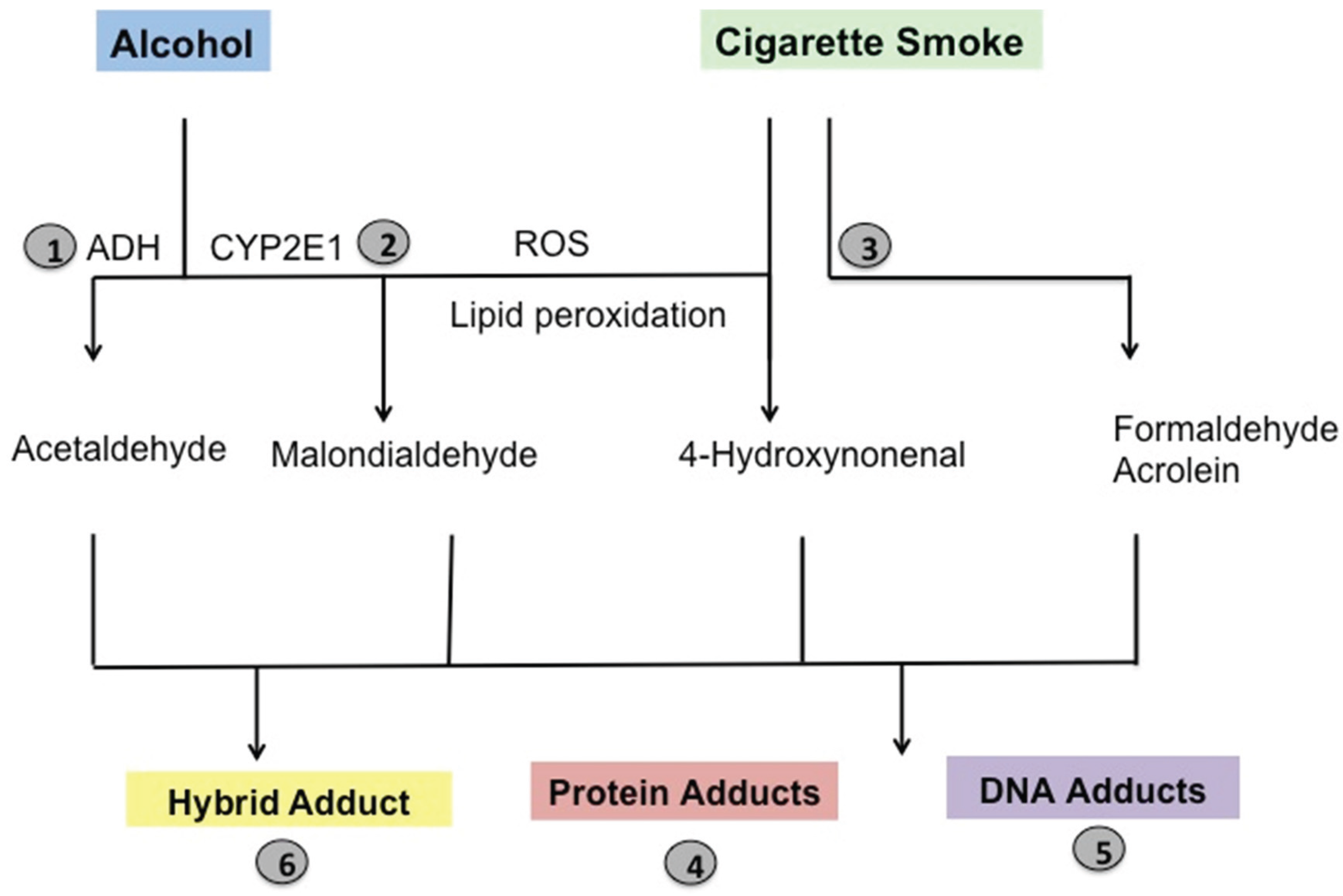
6. Pathological Implications of Lung Aldehydes

{kind=link}
{kind=link}
| Lung Aldehydes | Source | Lung Effect |
| Acetaldehyde | Oxidative stress [78,79] | |
| Malondialdehyde | Alcohol | COPD, asthma, emphysema [81,82] |
| 4-Hydroxynonenal | Cigarette smoke | Mild respiratory irritation [83] |
| Acrolein | Environmental toxicants | Release of pro-inflammatory cytokine [84,85] |
| Formaldehyde | Epigenetic and genetic effect [89] | |
| Lung Adducts | Aldehydes Involved | Lung Effect |
| Protein adduct | Damage protein structure and function [93,94] | |
| Acetaldehyde, | Slow cilia beating [95] | |
| Malondialdehyde, | Inhibition of anti-oxidative defense [96] | |
| 4-hydroxynonenal | Stimulation of fibrogenesis [97,98] | |
| and induction of immune response [99,100,101] | ||
| DNA adduct | Acetaldehyde, | Base pair mutation [102,103] |
| Malondialdehyde, | Carcinogenesis [104,105] | |
| Formaldehyde | Increased risk of mutation [102,106] | |
| Hybrid adduct | Acetaldehyde, Malondialdehyde | Induce pro-inflammatory chemokine [107] |
| Inhibit bronchial epithelial cell wound closure [108] | ||
| Increase influx of neutrophils [74] |
7. Lung Aldehydes and Protein Adduction
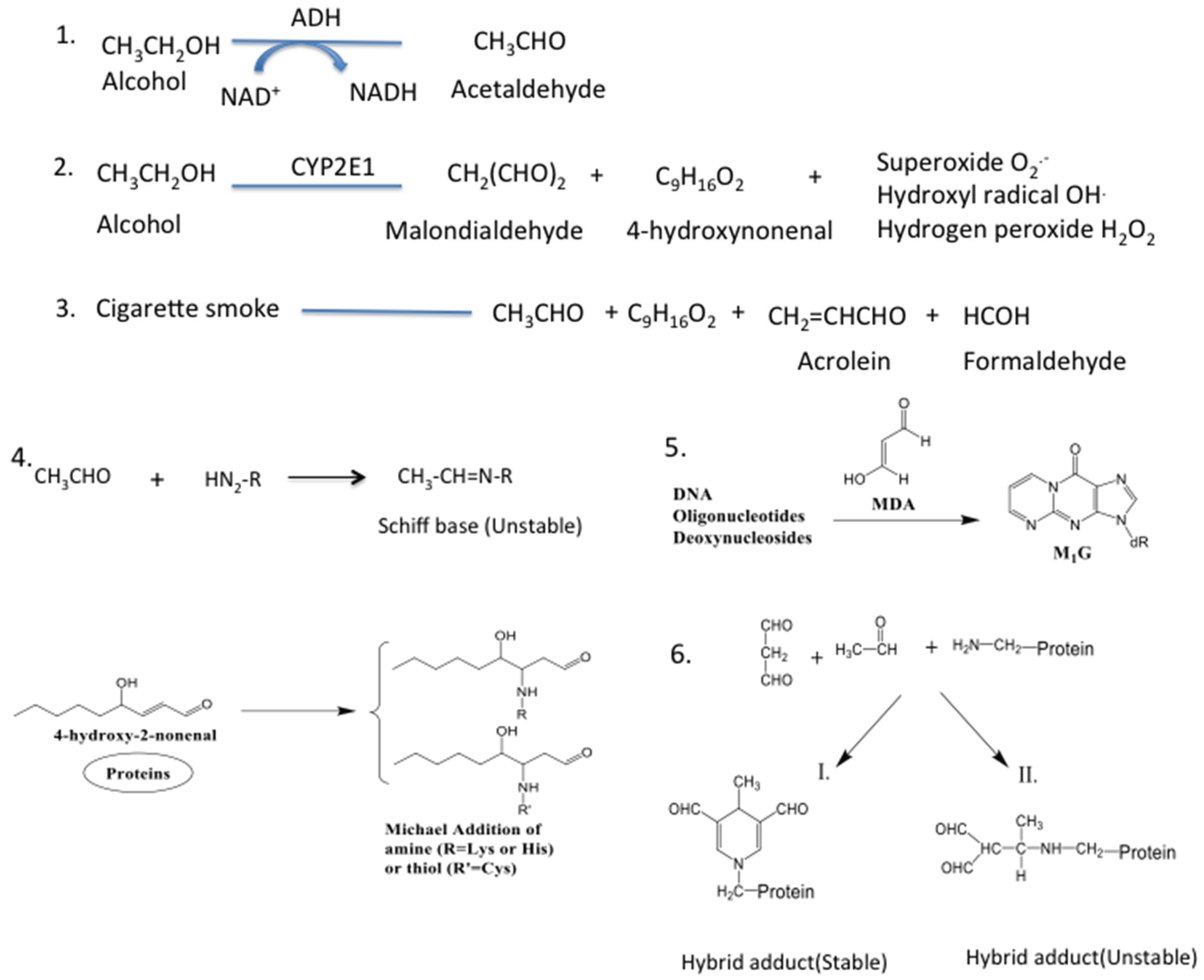
8. Pathological Implications of Protein Adducts
9. Lung Aldehydes and DNA Adduction
10. Pathological Implications of DNA Adducts
11. Lung Aldehydes and Hybrid Adducts
12. Pathological Implications of Hybrid Adduct
13. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Toews, G.B. Pulmonary defense mechanisms. Semin. Respir. Infect. 1993, 8, 160–167. [Google Scholar] [PubMed]
- Nicod, L. Lung defences: An overview. Eur. Respir. Rev. 2005, 14, 45–50. [Google Scholar] [CrossRef]
- Zoltaszek, R.; Kowalczyk, P.; Kowalczyk, M.C.; Hanausek, M.; Kilianska, Z.M.; Slaga, T.J.; Walaszek, Z. Dietary d-glucarate effects on the biomarkers of inflammation during early post-initiation stages of benzo[a]pyrene-induced lung tumorigenesis in A/J mice. Oncol. Lett. 2011, 2, 145–154. [Google Scholar] [PubMed]
- Hasani, A.; Pavia, D.; Spiteri, M.A.; Yeo, C.T.; Agnew, J.E.; Clarke, S.W.; Chung, K.F. Inhaled frusemide does not affect lung mucociliary clearance in healthy and asthmatic subjects. Eur. Respir. J. 1994, 7, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Zaas, A.K.; Schwartz, D.A. Innate immunity and the lung: Defense at the interface between host and environment. Trends Cardiovasc. Med. 2005, 15, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Bals, R. Epithelial antimicrobial peptides in host defense against infection. Respir. Res. 2000, 1, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Azad, N.; Rojanasakul, Y.; Vallyathan, V. Inflammation and lung cancer: Roles of reactive oxygen/nitrogen species. J. Toxicol. Environ. Health Part B 2008, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rosanna, D.P.; Salvatore, C. Reactive oxygen species, inflammation, and lung diseases. Curr. Pharm. Des. 2012, 18, 3889–3900. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.A.; Smith, R.A.; Safe, S.; Szabo, C.; Tjalkens, R.B.; Robertson, F.M. Toxicological and pathophysiological roles of reactive oxygen and nitrogen species. Toxicology 2010, 276, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I. Oxidative stress in pathogenesis of chronic obstructive pulmonary disease. Cell Biochem. Biophys. 2005, 43, 167–188. [Google Scholar] [CrossRef]
- Bouchery, E.E.; Harwood, H.J.; Sacks, J.J.; Simon, C.J.; Brewer, R.D. Economic costs of excessive alcohol consumption in the US, 2006. Am. J. Prev. Med. 2011, 41, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, K.; Roeber, J.; Kanny, D.; Tran, A.; Saiki, C.; Johnson, H.; Yeoman, K.; Safranek, T.; Creppage, K.; Lepp, A. Alcohol-attributable deaths and years of potential life lost—11 States, 2006–2010. Morb. Mortal. Wkly. Rep. 2014, 63, 213–216. [Google Scholar]
- Pearson, T.A. Alcohol and heart disease. Circulation 1996, 94, 3023–3025. [Google Scholar] [CrossRef] [PubMed]
- Moss, M.; Burnham, E.L. Alcohol abuse in the critically ill patient. Lancet 2007, 368, 2231–2242. [Google Scholar] [CrossRef]
- Allen, N.E.; Beral, V.; Casabonne, D.; Kan, S.W.; Reeves, G.K.; Brown, A.; Green, J. Moderate alcohol intake and cancer incidence in women. J. Natl. Cancer Inst. 2009, 101, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.A.S.; Cook, R.T.; Jerrells, T.R.; Kolls, J.K.; Nagy, L.E.; Szabo, G.; Wands, J.R.; Kovacs, E.J. Acute and chronic alcohol abuse modulate immunity. Alcohol. Clin. Exp. Res. 2006, 30, 1624–1631. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Yeligar, S.M.; Brown, L.A.S. Chronic-alcohol-abuse-induced oxidative stress in the development of acute respiratory distress syndrome. Sci. World J. 2012. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.W. Excretion of alcohol in urine and diuresis in healthy men in relation to their age, the dose administered and the time after drinking. Forensic Sci. Int. 1990, 45, 217–224. [Google Scholar] [CrossRef]
- Kalant, H. Absorption, diffusion, distribution, and elimination of ethanol: Effects on biological membranes. In The biology of alcoholism; Springer US: New York, NY, USA, 1971; pp. 1–62. [Google Scholar]
- Sisson, J.H. Alcohol and airways function in health and disease. Alcohol 2007, 41, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Hlastala, M.P.; Anderson, J.C. The impact of breathing pattern and lung size on the alcohol breath test. Annu. Biomed. Eng. 2007, 35, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, C.D.; Guidot, D.M. Alcoholic lung disease. Alcohol Res. Health 2008, 31, 66–75. [Google Scholar] [PubMed]
- Jong, G.; Hsiue, T.; Chen, C.; Chang, H.; Chen, C. Rapidly fatal outcome of bacteremic Klebsiella pneumoniae pneumonia in alcoholics. CHEST J. 1995, 107, 214–217. [Google Scholar] [CrossRef]
- Molina, P.E.; Happel, K.I.; Zhang, P.; Kolls, J.K.; Nelson, S. Focus on: Alcohol and the immune system. Alcohol Res. Health 2010, 33, 97–108. [Google Scholar] [PubMed]
- Sachs, C.W.; Christensen, R.H.; Pratt, P.C.; Lynn, W.S. Neutrophil elastase activity and superoxide production are diminished in neutrophils of alcoholics. Am. Rev. Respir. Dis. 1990, 141, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Guidot, D.M.; Modelska, K.; Lois, M.; Jain, L.; Moss, I.M.; Pittet, J.F.; Brown, L.A. Ethanol ingestion via glutathione depletion impairs alveolar epithelial barrier function in rats. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L127–L135. [Google Scholar] [PubMed]
- Guidot, D.; Moss, M.; Holguin, F.; Lois, M.; Brown, L. Ethanol ingestion impairs alveolar epithelial glutathione homeostasis and function, and predisposes to endotoxin-mediated acute lung injury. CHEST J. 1999, 116, 82S. [Google Scholar] [CrossRef]
- Birt, J.E.C.; Shuker, D.E.; Farmer, P.B. Stable acetaldehyde-protein adducts as biomarkers of alcohol exposure. Chem. Res. Toxicol. 1998, 11, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Current cigarette smoking among adults—United States, 2011. Morb. Mortal. Wkly. Rep. 2012, 61, 889–894. [Google Scholar]
- Blot, W.J.; McLaughlin, J.K.; Winn, D.M.; Austin, D.F.; Greenberg, R.S.; Preston-Martin, S.; Bernstein, L.; Schoenberg, J.B.; Stemhagen, A.; Fraumeni, J.F., Jr. Smoking and drinking in relation to oral and pharyngeal cancer. Cancer Res. 1988, 48, 3282–3287. [Google Scholar] [PubMed]
- Newcomb, P.A.; Carbone, P.P. The health consequences of smoking. Cancer. Med. Clin. North Am. 1992, 76, 305–331. [Google Scholar] [PubMed]
- Sasco, A.; Secretan, M.; Straif, K. Tobacco smoking and cancer: A brief review of recent epidemiological evidence. Lung Cancer 2004, 45, S3–S9. [Google Scholar] [CrossRef] [PubMed]
- Egleston, B.L.; Meireles, S.I.; Flieder, D.B.; Clapper, M.L. Population-based trends in lung cancer incidence in women. Semin. Oncol. 2009, 36, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Lopez, A.D.; Collishaw, N.E.; Piha, T. A descriptive model of the cigarette epidemic in developed countries. Tob. Control 1994, 3, 242–247. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (US); National Center for Chronic Disease Prevention and Health Promotion (US); Office on Smoking and Health (US). How Tobacco Smoke Causes Disease: The Biology and Behavioral Basis for Smoking-Attributable Disease; A Report of the Surgeon General: Atlanta, GA, USA, 2010.
- Hecht, S.S. Cigarette smoking and lung cancer: Chemical mechanisms and approaches to prevention. Lancet Oncol. 2002, 3, 461–469. [Google Scholar] [CrossRef]
- Phillips, D.H. DNA adducts in human tissues: Biomarkers of exposure to carcinogens in tobacco smoke. Environ. Health Perspect. 1996, 104, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Van der Vaart, H.; Postma, D.S.; Timens, W.; ten Hacken, N.H. Acute effects of cigarette smoke on inflammation and oxidative stress: A review. Thorax 2004, 59, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Munnia, A.; Bonassi, S.; Verna, A.; Quaglia, R.; Pelucco, D.; Ceppi, M.; Neri, M.; Buratti, M.; Taioli, E.; Garte, S. Bronchial malondialdehyde DNA adducts, tobacco smoking, and lung cancer. Free Radic. Biol. Med. 2006, 41, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- RafiqKhan, M.; Sellappa, S. Serum malondialdehyde levels in lung cancer patients. J. Appl. Biol. Biotechnol. 2013, 1, 032–034. [Google Scholar]
- Behr, J.; Nowak, D. Tobacco smoke and respiratory disease. Eur. Respair. Mon. 2002, 21, 161–179. [Google Scholar]
- Hunninghake, G.W.; Crystal, R.G. Cigarette smoking and lung destruction: Accumulation of neutrophils in the lungs of cigarette smokers 1, 2. Am. Rev. Respir. Dis. 1983, 128, 833–838. [Google Scholar] [PubMed]
- Shapiro, S.D. The macrophage in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1999, 160, S29–S32. [Google Scholar] [CrossRef] [PubMed]
- Galvin, J.R.; Franks, T.J. Smoking-related lung disease. J. Thorac. Imaging 2009, 24, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Rahman, I. Current concepts on oxidative/carbonyl stress, inflammation and epigenetics in pathogenesis of chronic obstructive pulmonary disease. Toxicol. Appl. Pharmacol. 2011, 254, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, K.; Shibamoto, T. Determination of toxic carbonyl compounds in cigarette smoke. Environ. Toxicol. 2006, 21, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I. The role of oxidative stress in the pathogenesis of COPD. Treat. Respir. Med. 2005, 4, 175–200. [Google Scholar] [CrossRef] [PubMed]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, K.; Loridas, S. Pulmonary oxidative stress, inflammation and cancer: Respirable particulate matter, fibrous dusts and ozone as major causes of lung carcinogenesis through reactive oxygen species mechanisms. Int. J. Environ. Res. Public Health 2013, 10, 3886–3907. [Google Scholar] [CrossRef] [PubMed]
- Hiura, T.S.; Kaszubowski, M.P.; Li, N.; Nel, A.E. Chemicals in diesel exhaust particles generate reactive oxygen radicals and induce apoptosis in macrophages. J. Immunol. 1999, 163, 5582–5591. [Google Scholar] [PubMed]
- Frampton, M.W.; Pryor, W.A.; Cueto, R.; Cox, C.; Morrow, P.E.; Utell, M.J. Ozone exposure increases aldehydes in epithelial lining fluid in human lung. Am. J. Respir. Crit. Care Med. 1999, 159, 1134–1137. [Google Scholar] [CrossRef] [PubMed]
- Gardi, C.; Valacchi, G. Cigarette smoke and ozone effect on murine inflammatory responses. Ann. N. Y. Acad. Sci. 2012, 1259, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Sunil, V.R.; Patel-Vayas, K.; Shen, J.; Laskin, J.D.; Laskin, D.L. Classical and alternative macrophage activation in the lung following ozone-induced oxidative stress. Toxicol. Appl. Pharmacol. 2012, 263, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Lodovici, M.; Bigagli, E. Oxidative stress and air pollution exposure. J. Toxicol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Kaphalia, L.; Calhoun, W.J. Alcoholic lung injury: Metabolic, biochemical and immunological aspects. Toxicol. Lett. 2013, 222, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Matsuse, H.; Fukushima, C.; Shimoda, T.; Asai, S.; Kohno, S. Effects of Acetaldehyde on human airway constriction and inflammation. In Novartis Foundation Symposium; John Wiley: New York, NY, USA, 2007; Volume 285, p. 97. [Google Scholar]
- Aytacoglu, B.N.; Calikoglu, M.; Tamer, L.; Coskun, B.; Sucu, N.; Kose, N.; Aktas, S.; Dikmengil, M. Alcohol-induced lung damage and increased oxidative stress. Respiration 2006, 73, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Hukkanen, J.; Pelkonen, O.; Hakkola, J.; Raunio, H. Expression and regulation of xenobiotic-metabolizing cytochrome P450 (CYP) enzymes in human lung. Crit. Rev. Toxicol. 2002, 32, 391–411. [Google Scholar] [CrossRef] [PubMed]
- Kaphalia, L.; Boroumand, N.; Hyunsu, J.; Kaphalia, B.S.; Calhoun, W.J. Ethanol metabolism, oxidative stress, and endoplasmic reticulum stress responses in the lungs of hepatic alcohol dehydrogenase deficient deer mice after chronic ethanol feeding. Toxicol. Appl. Pharmacol. 2014, 277, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Vasiliou, V.; Zakhari, S.; Seitz, H.K.; Hoek, J.B. Biological Basis of Alcohol-Induced Cancer; Springer: New York, NY, USA, 2015. [Google Scholar]
- Zheng, R.; Dragomir, A.; Mishin, V.; Richardson, J.R.; Heck, D.E.; Laskin, D.L.; Laskin, J.D. Differential metabolism of 4-hydroxynonenal in liver, lung and brain of mice and rats. Toxicol. Appl. Pharmacol. 2014, 279, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Hoberman, H.D.; George, R.C.S. Reaction of tobacco smoke aldehydes with human hemoglobin. J. Biochem. Toxicol. 1988, 3, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Piadé, J.; Wajrock, S.; Jaccard, G.; Janeke, G. Formation of mainstream cigarette smoke constituents prioritized by the World Health Organization—Yield patterns observed in market surveys, clustering and inverse correlations. Food Chem. Toxicol. 2013, 55, 329–347. [Google Scholar] [CrossRef] [PubMed]
- Chepiga, T.; Morton, M.; Murphy, P.; Avalos, J.; Bombick, B.; Doolittle, D.; Borgerding, M.; Swauger, J. A comparison of the mainstream smoke chemistry and mutagenicity of a representative sample of the US cigarette market with two Kentucky reference cigarettes (K1R4F and K1R5F). Food Chem. Toxicol. 2000, 38, 949–962. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Allil Compounds, Aldehydes, Epoxides and Peroxides; International Agency for Research on Cancer: Lyon, France, 1985; Volume 36. [Google Scholar]
- Tuma, J.; Casey, C.A. Dangerous byproducts of alcohol breakdown-focus on adducts. Alcohol Res. Health 2003, 27, 285–290. [Google Scholar] [PubMed]
- Corradi, M.; Rubinstein, I.; Andreoli, R.; Manini, P.; Caglieri, A.; Poli, D.; Alinovi, R.; Mutti, A. Aldehydes in exhaled breath condensate of patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2003, 167, 1380–1386. [Google Scholar] [CrossRef] [PubMed]
- Salaspuro, V.; Salaspuro, M. Synergistic effect of alcohol drinking and smoking on in vivo acetaldehyde concentration in saliva. Int. J. Cancer 2004, 111, 480–483. [Google Scholar] [CrossRef] [PubMed]
- Pryor, W.A.; Bermúdez, E.; Cueto, R.; Squadrito, G.L. Detection of aldehydes in bronchoalveolar lavage of rats exposed to ozone. Toxicol. Sci. 1996, 34, 148–156. [Google Scholar] [CrossRef]
- Hecht, S.S. Smoking and lung cancer—A new role for an old toxicant? Proc. Natl. Acad. Sci. USA 2006, 103, 15725–15726. [Google Scholar] [CrossRef] [PubMed]
- Lodovici, M.; Bigagli, E. Biomarkers of induced active and passive smoking damage. Int. J. Environ. Res. Public Health 2009, 6, 874–888. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.H. Smoking-related DNA and protein adducts in human tissues. Carcinogenesis 2002, 23, 1979–2004. [Google Scholar] [CrossRef] [PubMed]
- Araneda, O.; García, C.; Lagos, N.; Quiroga, G.; Cajigal, J.; Salazar, M.; Behn, C. Lung oxidative stress as related to exercise and altitude. Lipid peroxidation evidence in exhaled breath condensate: A possible predictor of acute mountain sickness. Eur. J. Appl. Physiol. 2005, 95, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Mercken, E.M.; Hageman, G.J.; Schols, A.M.; Akkermans, M.A.; Bast, A.; Wouters, E.F. Rehabilitation decreases exercise-induced oxidative stress in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2005, 172, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.A.; Kharbanda, K.K.; McCaskill, M.L.; Tuma, D.J.; Yanov, D.; DeVasure, J.; Sisson, J.H. Malondialdehyde-acetaldehyde-adducted protein inhalation causes lung injury. Alcohol 2012, 46, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Sherman, D.; Watson, R.R. Ethanol and the Liver: Mechanisms and Management; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Negre-Salvayre, A.; Coatrieux, C.; Ingueneau, C.; Salvayre, R. Advanced lipid peroxidation end products in oxidative damage to proteins. Potential role in diseases and therapeutic prospects for the inhibitors. Br. J. Pharmacol. 2008, 153, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Niedernhofer, L.J.; Daniels, J.S.; Rouzer, C.A.; Greene, R.E.; Marnett, L.J. Malondialdehyde, a product of lipid peroxidation, is mutagenic in human cells. J. Biol. Chem. 2003, 278, 31426–31433. [Google Scholar] [CrossRef] [PubMed]
- Negre-Salvayre, A.; Auge, N.; Ayala, V.; Basaga, H.; Boada, J.; Brenke, R.; Chapple, S.; Cohen, G.; Feher, J.; Grune, T. Pathological aspects of lipid peroxidation. Free Radic. Res. 2010, 44, 1125–1171. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Park, Y.S. Role of lipid peroxidation-derived α, β-unsaturated aldehydes in vascular dysfunction. Oxid. Med. Cell. Longev. 2013. [Google Scholar] [CrossRef] [PubMed]
- Al-Rubaye, F.G. Correlation of serum Malonialdehyde Acetylaldehyde Adduct to serum Malondialdehyde as oxidative stress markers in Acute Coronary Syndrome patients. Indian J. Appl. Res. 2014, 4, 44–47. [Google Scholar] [CrossRef]
- Parola, M.; Bellomo, G.; Robino, G.; Barrera, G.; Dianzani, M.U. 4-Hydroxynonenal as a biological signal: Molecular basis and pathophysiological implications. Antioxid. Redox Signal. 1999, 1, 255–284. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, T.; Kohno, S.; Takao, A.; Fujiwara, C.; Matsuse, H.; Sakai, H.; Watanabe, T.; Hara, K.; Asai, S. Investigation of the mechanism of alcohol-induced bronchial asthma. J. Allergy Clin. Immunol. 1996, 97, 74–84. [Google Scholar] [CrossRef]
- McConnell, E. Emergency and Continuous Exposure Guidance Levels for Selected Submarine Contaminants: Volume, 1; The National Academies Press: Washington, DC, USA, 2007; pp. 195–208. [Google Scholar]
- Moretto, N.; Facchinetti, F.; Southworth, T.; Civelli, M.; Singh, D.; Patacchini, R. α, β-Unsaturated aldehydes contained in cigarette smoke elicit IL-8 release in pulmonary cells through mitogen-activated protein kinases. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L839–L848. [Google Scholar] [CrossRef] [PubMed]
- Facchinetti, F.; Amadei, F.; Geppetti, P.; Tarantini, F.; di Serio, C.; Dragotto, A.; Gigli, P.M.; Catinella, S.; Civelli, M.; Patacchini, R. α, β-Unsaturated aldehydes in cigarette smoke release inflammatory mediators from human macrophages. Am. J. Respir. Cell Mol. Biol. 2007, 37, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, E.I.; Ruben, J.; Koot, C.W.; Hristova, M.; van der Vliet, A. Regulation of constitutive neutrophil apoptosis by the α, β-unsaturated aldehydes acrolein and 4-hydroxynonenal. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 289, L1019–L1028. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, H.S.; Shaver, C.; Case, L.M.; Dietsch, M.; Wesselkamper, S.C.; Hardie, W.D.; Korfhagen, T.R.; Corradi, M.; Nadel, J.A.; Borchers, M.T. Acrolein-activated matrix metalloproteinase 9 contributes to persistent mucin production. Am. J. Respir. Cell Mol. Biol. 2008, 38, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Van der Toorn, M.; Smit-de Vries, M.P.; Slebos, D.J.; de Bruin, H.G.; Abello, N.; van Oosterhout, A.J.; Bischoff, R.; Kauffman, H.F. Cigarette smoke irreversibly modifies glutathione in airway epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L1156–L1162. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.D.; Lim, R.W. Epigenetic effects of ethanol on the liver and gastrointestinal system. Alcohol. Res. 2013, 35, 47–55. [Google Scholar] [PubMed]
- Shukla, S.D.; Aroor, A.R. Epigenetic effects of ethanol on liver and gastrointestinal injury. World J. Gastroenterol. 2006, 12, 5265–5271. [Google Scholar] [PubMed]
- Manzo-Avalos, S.; Saavedra-Molina, A. Cellular and mitochondrial effects of alcohol consumption. Int. J. Environ. Res. Public Health 2010, 7, 4281–4304. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Fang, L.; Li, H.; Tang, M.S.; Jin, C. Cigarette smoke component acrolein modulates chromatin assembly by inhibiting histone acetylation. J. Biol. Chem. 2013, 288, 21678–21687. [Google Scholar] [CrossRef] [PubMed]
- Grimsrud, P.A.; Xie, H.; Griffin, T.J.; Bernlohr, D.A. Oxidative stress and covalent modification of protein with bioactive aldehydes. J. Biol. Chem. 2008, 283, 21837–21841. [Google Scholar] [CrossRef] [PubMed]
- Doorn, J.A.; Petersen, D.R. Covalent adduction of nucleophilic amino acids by 4-hydroxynonenal and 4-oxononenal. Chem. Biol. Interact. 2003, 143, 93–100. [Google Scholar] [CrossRef]
- Sisson, J.H.; Tuma, D.J.; Rennard, S.I. Acetaldehyde-mediated cilia dysfunction in bovine bronchial epithelial cells. Am. J. Physiol. 1991, 260, L29–L36. [Google Scholar] [PubMed]
- Sultana, R.; Raju, B.S.S.; Sharma, V.; Reddanna, P.; Babu, P.P. Formation of acetaldehyde adducts of glutathione S-transferase A3 in the liver of rats administered alcohol chronically. Alcohol 2005, 35, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Casini, A.; Cunningham, M.; Rojkind, M.; Lieber, C.S. Acetaldehyde increases procollagen type I and fibronectin gene transcription in cultured rat fat-storing cells through a protein synthesis-dependent mechanism. Hepatology 1991, 13, 758–765. [Google Scholar] [CrossRef]
- Brenner, D.A.; Chojkier, M. Acetaldehyde increases collagen gene transcription in cultured human fibroblasts. J. Biol. Chem. 1987, 262, 17690–17695. [Google Scholar] [PubMed]
- Niemelä, O.; Klajner, F.; Orrego, H.; Vidins, E.; Blendis, L.; Israel, Y. Antibodies against acetaldehyde-modified protein epitopes in human alcoholics. Hepatology 1987, 7, 1210–1214. [Google Scholar] [CrossRef] [PubMed]
- Niemelä, O. Distribution of ethanol-induced protein adducts in vivo: Relationship to tissue injury 1, 2. Free Radic. Biol. Med. 2001, 31, 1533–1538. [Google Scholar] [CrossRef]
- Xu, D.; Thiele, G.M.; Beckenhauer, J.L.; Klassen, L.W.; Sorrell, M.F.; Tuma, D.J. Detection of circulating antibodies to malondialdehyde-acetaldehyde adducts in ethanol-fed rats. Gastroenterology 1998, 115, 686–692. [Google Scholar] [CrossRef]
- VanderVeen, L.A.; Hashim, M.F.; Shyr, Y.; Marnett, L.J. Induction of frameshift and base pair substitution mutations by the major DNA adduct of the endogenous carcinogen malondialdehyde. Proc. Natl. Acad. Sci. USA 2003, 100, 14247–14252. [Google Scholar] [CrossRef] [PubMed]
- Marnett, L.J. Oxyradicals and DNA damage. Carcinogenesis 2000, 21, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Riedel, F.; Goessler, U.; Hörmann, K. Alcohol-related diseases of the mouth and throat. Best Pract. Res. Clin. Gastroenterol. 2003, 17, 543–555. [Google Scholar] [CrossRef]
- Theruvathu, J.A.; Jaruga, P.; Nath, R.G.; Dizdaroglu, M.; Brooks, P.J. Polyamines stimulate the formation of mutagenic 1,N2-propanodeoxyguanosine adducts from acetaldehyde. Nucleic Acids Res. 2005, 33, 3513–3520. [Google Scholar] [CrossRef] [PubMed]
- Peluso, M.; Srivatanakul, P.; Munnia, A.; Jedpiyawongse, A.; Ceppi, M.; Sangrajrang, S.; Piro, S.; Boffetta, P. Malondialdehyde-deoxyguanosine adducts among workers of a Thai industrial estate and nearby residents. Environ. Health Perspect. 2010, 118, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.A.; Kharbanda, K.K.; Tuma, D.J.; Sisson, J.H. Malondialdehyde-acetaldehyde-adducted bovine serum albumin activates protein kinase C and stimulates interleukin-8 release in bovine bronchial epithelial cells. Alcohol 2001, 25, 159–166. [Google Scholar] [CrossRef]
- Wyatt, T.A.; Kharbanda, K.K.; Tuma, D.J.; Sisson, J.H.; Spurzem, J.R. Malondialdehyde-acetaldehyde adducts decrease bronchial epithelial wound repair. Alcohol 2005, 36, 31–40. [Google Scholar] [CrossRef] [PubMed]
- McCaskill, M.L.; Kharbanda, K.K.; Tuma, D.J.; Reynolds, J.D.; DeVasure, J.M.; Sisson, J.H.; Wyatt, T.A. Hybrid malondialdehyde and acetaldehyde protein adducts form in the lungs of mice exposed to alcohol and cigarette smoke. Alcohol. Clin. Exp. Res. 2011, 35, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Lopachin, R.M.; Decaprio, A.P. Protein adduct formation as a molecular mechanism in neurotoxicity. Toxicol. Sci. 2005, 86, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Fenaille, F.; Guy, P.A.; Tabet, J. Study of protein modification by 4-hydroxy-2-nonenal and other short chain aldehydes analyzed by electrospray ionization tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2003, 14, 215–226. [Google Scholar] [CrossRef]
- Cai, J.; Bhatnagar, A.; Pierce, W.M., Jr. Protein modification by acrolein: Formation and stability of cysteine adducts. Chem. Res. Toxicol. 2009, 22, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Donohue, T.M.; Tuma, D.J.; Sorrell, M.F. Acetaldehyde adducts with proteins: Binding of [14 C] acetaldehyde to serum albumin. Arch. Biochem. Biophys. 1983, 220, 239–246. [Google Scholar] [CrossRef]
- Tuma, D.J.; Newman, M.R.; Donohue, T.M.; Sorrell, M.F. Covalent binding of acetaldehyde to proteins: Participation of lysine residues. Alcohol. Clin. Exp. Res. 1987, 11, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Tuma, D.J.; Jennett, R.B.; Sorrell, M.F. The interaction of acetaldehyde with tubulina. Ann. N. Y. Acad. Sci. 1987, 492, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Tuma, D.J.; Hoffman, T.; Sorrell, M.F. The chemistry of acetaldehyde-protein adducts. Alcohol Alcohol. Suppl. 1991, 1, 271–276. [Google Scholar] [PubMed]
- Requena, J.R.; Fu, M.X.; Ahmed, M.U.; Jenkins, A.J.; Lyons, T.J.; Thorpe, S.R. Lipoxidation products as biomarkers of oxidative damage to proteins during lipid peroxidation reactions. Nephrol. Dial. Transplant. 1996, 11, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, M.F.; Tuma, D.J. The functional implications of acetaldehyde binding to cell constituentsa. Ann. N. Y. Acad. Sci. 1987, 492, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Eiserich, J.P.; van der Vliet, A.; Handelman, G.J.; Halliwell, B.; Cross, C.E. Dietary antioxidants and cigarette smoke-induced biomolecular damage: A complex interaction. Am. J. Clin. Nutr. 1995, 62, 1490S–1500S. [Google Scholar] [PubMed]
- Uchida, K.; Stadtman, E.R. Modification of histidine residues in proteins by reaction with 4-hydroxynonenal. Proc. Natl. Acad. Sci. USA 1992, 89, 4544–4548. [Google Scholar] [CrossRef] [PubMed]
- Novitskiy, G.; Ravi, R.; Potter, J.J.; Rennie-Tankersley, L.; Wang, L.; Mezey, E. Effects of acetaldehyde and TNFα on the inhibitory kappa B-α protein and nuclear factor kappa B activation in hepatic stellate cells. Alcohol Alcohol. 2005, 40, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Kirichenko, A.; Li, L.; Morandi, M.T.; Holian, A. 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murine lung cells after acute ozone exposure. Toxicol. Appl. Pharmacol. 1996, 141, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.B.; Worrall, S.; Emery, P.W.; Preedy, V.R. Protein adduct species in muscle and liver of rats following acute ethanol administration. Alcohol Alcohol. 2005, 40, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G.; Pizzimenti, S.; Ciamporcero, E.S.; Daga, M.; Ullio, C.; Arcaro, A.; Cetrangolo, G.P.; Ferretti, C.; Dianzani, C.; Lepore, A. Role of 4-hydroxynonenal-protein adducts in human diseases. Antioxid. Redox Signal. 2014, 22, 1681–1702. [Google Scholar] [CrossRef] [PubMed]
- Shamoto-Nagai, M.; Maruyama, W.; Hashizume, Y.; Yoshida, M.; Osawa, T.; Riederer, P.; Naoi, M. In parkinsonian substantia nigra, α-synuclein is modified by acrolein, a lipid-peroxidation product, and accumulates in the dopamine neurons with inhibition of proteasome activity. J. Neural Transm. 2007, 114, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Kanematsu, M.; Sakai, K.; Matsuda, T.; Hattori, N.; Mizuno, Y.; Suzuki, D.; Miyata, T.; Noguchi, N.; Niki, E.; et al. Protein-bound acrolein: Potential markers for oxidative stress. Proc. Natl. Acad. Sci. USA 1998, 95, 4882–4887. [Google Scholar] [CrossRef] [PubMed]
- Moretto, N.; Volpi, G.; Pastore, F.; Facchinetti, F. Acrolein effects in pulmonary cells: Relevance to chronic obstructive pulmonary disease. Ann. N. Y. Acad. Sci. 2012, 1259, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Chavez, J.; Chung, W.; Miranda, C.L.; Singhal, M.; Stevens, J.F.; Maier, C.S. Site-specific protein adducts of 4-hydroxy-2(E)-nonenal in human THP-1 monocytic cells: Protein carbonylation is diminished by ascorbic acid. Chem. Res. Toxicol. 2009, 23, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Niemelä, O.; Parkkila, S.; Pasanen, M.; Limuro, Y.; Bradford, B.; Thurman, R.G. Early alcoholic liver injury: Formation of protein adducts with acetaldehyde and lipid peroxidation products, and expression of CYP2E1 and CYP3A. Alcohol. Clin. Exp. Res. 1998, 22, 2118–2124. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Nanji, A.A.; Siakotos, A.N.; Lin, R.C. Acetaldehyde-modified and 4-hydroxynonenal-modified proteins in the livers of rats with alcoholic liver disease. Hepatology 1997, 26, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, P.A.; Caramori, G.; Casolari, P.; Papi, A.A.; Edwards, M.; Shamji, B.; Triantaphyllopoulos, K.; Hussain, F.; Pinart, M.; Khan, Y. Oxidative stress-induced antibodies to carbonyl-modified protein correlate with severity of chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2011, 184, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Sampey, B.P.; Stewart, B.J.; Petersen, D.R. Ethanol-induced modulation of hepatocellular extracellular signal-regulated kinase-1/2 activity via 4-hydroxynonenal. J. Biol. Chem. 2007, 282, 1925–1937. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.M.; Mullally, J.E.; Fitzpatrick, F.A. Reactive lipid species from cyclooxygenase-2 inactivate tumor suppressor LKB1/STK11: Cyclopentenone prostaglandins and 4-hydroxy-2-nonenal covalently modify and inhibit the AMP-kinase kinase that modulates cellular energy homeostasis and protein translation. J. Biol. Chem. 2006, 281, 2598–2604. [Google Scholar] [PubMed]
- Parola, M.; Robino, G.; Marra, F.; Pinzani, M.; Bellomo, G.; Leonarduzzi, G.; Chiarugi, P.; Camandola, S.; Poli, G.; Waeg, G.; et al. HNE interacts directly with JNK isoforms in human hepatic stellate cells. J. Clin. Investig. 1998, 102, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Schutt, F.; Bergmann, M.; Holz, F.G.; Kopitz, J. Proteins modified by malondialdehyde, 4-hydroxynonenal, or advanced glycation end products in lipofuscin of human retinal pigment epithelium. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3663–3668. [Google Scholar] [CrossRef]
- Wang, M.; McIntee, E.J.; Cheng, G.; Shi, Y.; Villalta, P.W.; Hecht, S.S. Identification of DNA adducts of acetaldehyde. Chem. Res. Toxicol. 2000, 13, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Vaca, C.E.; Fang, J.; Schweda, E.K. Studies of the reaction of acetaldehyde with deoxynucleosides. Chem. Biol. Interact. 1995, 98, 51–67. [Google Scholar] [CrossRef]
- Wang, M.; Yu, N.; Chen, L.; Villalta, P.W.; Hochalter, J.B.; Hecht, S.S. Identification of an acetaldehyde adduct in human liver DNA and quantitation as N 2-ethyldeoxyguanosine. Chem. Res. Toxicol. 2006, 19, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Brooks, P.J.; Theruvathu, J.A. DNA adducts from acetaldehyde: Implications for alcohol-related carcinogenesis. Alcohol 2005, 35, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Bono, R.; Romanazzi, V.; Munnia, A.; Piro, S.; Allione, A.; Ricceri, F.; Guarrera, S.; Pignata, C.; Matullo, G.; Wang, P. Malondialdehyde-deoxyguanosine adduct formation in workers of pathology wards: The role of air formaldehyde exposure. Chem. Res. Toxicol. 2010, 23, 1342–1348. [Google Scholar] [CrossRef] [PubMed]
- Blair, I.A. DNA adducts with lipid peroxidation products. J. Biol. Chem. 2008, 283, 15545–15549. [Google Scholar] [CrossRef] [PubMed]
- Linhart, K.; Glassen, K.; Peccerella, T.; Waldherr, R.; Linhart, H.; Bartsch, H.; Seitz, H.K. The generation of carcinogenic etheno-DNA adducts in the liver of patients with nonalcoholic fatty liver disease. Hepatobil. Surg. Nutr. 2015, 4, 117–123. [Google Scholar]
- Hang, B. Formation and repair of tobacco carcinogen-derived bulky DNA adducts. J. Nucleic Acids 2010. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.K.; Nokubo, M.; Reddy, G.R.; Yeola, S.N.; Morrow, J.D.; Blair, I.A.; Marnett, L.J. Detection of endogenous malondialdehyde-deoxyguanosine adducts in human liver. Science 1994, 265, 1580–1582. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Dhingra, K.; Hittelman, W.N.; Liehr, J.G.; de Andrade, M.; Li, D. Lipid peroxidation-induced putative malondialdehyde-DNA adducts in human breast tissues. Cancer Epidemiol. Biomark. Prev. 1996, 5, 705–710. [Google Scholar]
- Zhang, Y.; Chen, S.Y.; Hsu, T.; Santella, R.M. Immunohistochemical detection of malondialdehyde-DNA adducts in human oral mucosa cells. Carcinogenesis 2002, 23, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Munnia, A.; Amasio, M.E.; Peluso, M. Exocyclic malondialdehyde and aromatic DNA adducts in larynx tissues. Free Radic. Biol. Med. 2004, 37, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, M.; Hottor, T.K.; deVasure, J.M.; Wyatt, T.A.; McCaskill, M.L. Protective role of CYP2E1 inhibitor diallyl disulfide (DADS) on alcohol-induced malondialdehyde-deoxyguanosine (M1dG) adduct formation. Alcohol. Clin. Exp. Res. 2014, 38, 1550–1558. [Google Scholar] [CrossRef] [PubMed]
- Linhart, K.; Bartsch, H.; Seitz, H.K. The role of reactive oxygen species (ROS) and cytochrome P-450 2E1 in the generation of carcinogenic etheno-DNA adducts. Redox Biol. 2014, 3, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.K.; Stickel, F. Molecular mechanisms of alcohol-mediated carcinogenesis. Nat. Rev. Cancer 2007, 7, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Drobes, D.J. Concurrent alcohol and tobacco dependence: Mechanisms and treatment. Alcohol Res. Health 2002, 26, 136–142. [Google Scholar]
- Miller, N.S.; Gold, M.S. Comorbid cigarette and alcohol addiction: Epidemiology and treatment. J. Addict. Dis. 1998, 17, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Ström, K. Alcohol, smoking and lung disease. Addict. Biol. 1999, 4, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Niemela, O. Aldehyde-protein adducts in the liver as a result of ethanol-induced oxidative stress. Front. Biosci. 1999, 4, D506–D513. [Google Scholar] [CrossRef] [PubMed]
- Tuma, D.J.; Kearley, M.L.; Thiele, G.M.; Worrall, S.; Haver, A.; Klassen, L.W.; Sorrell, M.F. Elucidation of reaction scheme describing malondialdehyde-acetaldehyde-protein adduct formation. Chem. Res. Toxicol. 2001, 14, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Tuma, D.J. Role of malondialdehyde-acetaldehyde adducts in liver injury 1, 2. Free Radic. Biol. Med. 2002, 32, 303–308. [Google Scholar] [CrossRef]
- Worrall, S.; de Jersey, J.; Wilce, P.A. Comparison of the formation of proteins modified by direct and indirect ethanol metabolites in the liver and blood of rats fed the Lieber-DeCarli liquid diet. Alcohol Alcohol. 2000, 35, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.P.; Simet, S.M.; DeVasure, J.M.; Boten, J.A.; Sweeter, J.M.; Kharbanda, K.K.; Sisson, J.H.; Wyatt, T.A. Malondialdehyde-acetaldehyde (MAA) adducted proteins bind to scavenger receptor A in airway epithelial cells. Alcohol 2014, 48, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Nkadi, P.O.; Merritt, T.A.; Pillers, D.M. An overview of pulmonary surfactant in the neonate: Genetics, metabolism, and the role of surfactant in health and disease. Mol. Genet. Metab. 2009, 97, 95–101. [Google Scholar] [PubMed]
- Duryee, M.J.; Freeman, T.L.; Willis, M.S.; Hunter, C.D.; Hamilton, B.C., 3rd; Suzuki, H.; Tuma, D.J.; Klassen, L.W.; Thiele, G.M. Scavenger receptors on sinusoidal liver endothelial cells are involved in the uptake of aldehyde-modified proteins. Mol. Pharmacol. 2005, 68, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- De Winther, M.P.; van Dijk, K.W.; Havekes, L.M.; Hofker, M.H. Macrophage scavenger receptor class A: A multifunctional receptor in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Hill, G.E.; Miller, J.A.; Baxter, B.T.; Klassen, L.W.; Duryee, M.J.; Tuma, D.J.; Thiele, G.M. Association of malondialdehyde-acetaldehyde (MAA) adducted proteins with atherosclerotic-induced vascular inflammatory injury. Atherosclerosis 1998, 141, 107–116. [Google Scholar] [CrossRef]
- Rolla, R.; Vay, D.; Mottaran, E.; Parodi, M.; Traverso, N.; Aricó, S.; Sartori, M.; Bellomo, G.; Klassen, L.W.; Thiele, G.M. Detection of circulating antibodies against malondialdehyde-acetaldehyde adducts in patients with alcohol-induced liver disease. Hepatology 2000, 31, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Duryee, M.J.; Klassen, L.W.; Freeman, T.L.; Willis, M.S.; Tuma, D.J.; Thiele, G.M. Lipopolysaccharide is a cofactor for malondialdehyde-acetaldehyde adduc mediated cytokine/chemokine release by rat sinusoidal liver endothelial and kupffer cells. Alcohol. Clin. Exp. Res. 2004, 28, 1931–1938. [Google Scholar] [CrossRef] [PubMed]
- Thiele, G.M.; Duryee, M.J.; Anderson, D.R.; Klassen, L.W.; Mohring, S.M.; Young, K.A.; Benissan-Messan, D.; Sayles, H.; Dusad, A.; Hunter, C.D. Malondialdehyde-acetaldehyde adducts and anti-malondialdehyde-acetaldehyde antibodies in rheumatoid arthritis. Arthritis Rheumatol. 2015, 67, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Willis, M.S.; Klassen, L.W.; Tuma, D.J.; Sorrell, M.F.; Thiele, G.M. Adduction of soluble proteins with malondialdehyde-acetaldehyde (MAA) induces antibody production and enhances T-cell proliferation. Alcohol. Clin. Exp. Res. 2002, 26, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.S.; Xiong, W.; Dale, M.; Yu, F.; Duryee, M.J.; Anderson, D.R.; Thiele, G.M.; Baxter, B.T. Antibodies against malondialdehyde-acetaldehyde adducts can help identify patients with abdominal aortic aneurysm. J. Vasc. Surg. 2015. [Google Scholar] [CrossRef] [PubMed]
- Kaur, I.; Katyal, A. Modification of mouse A2M B (620–792) and A2M N (168–230) by malondialdehyde and acetaldehyde attenuates the proteinase and TGF-β1 binding ability of A2MB. FEBS Lett. 2011, 585, 829–833. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sapkota, M.; Wyatt, T.A. Alcohol, Aldehydes, Adducts and Airways. Biomolecules 2015, 5, 2987-3008. https://doi.org/10.3390/biom5042987
Sapkota M, Wyatt TA. Alcohol, Aldehydes, Adducts and Airways. Biomolecules. 2015; 5(4):2987-3008. https://doi.org/10.3390/biom5042987
Chicago/Turabian StyleSapkota, Muna, and Todd A. Wyatt. 2015. "Alcohol, Aldehydes, Adducts and Airways" Biomolecules 5, no. 4: 2987-3008. https://doi.org/10.3390/biom5042987
APA StyleSapkota, M., & Wyatt, T. A. (2015). Alcohol, Aldehydes, Adducts and Airways. Biomolecules, 5(4), 2987-3008. https://doi.org/10.3390/biom5042987