
The Unconventional Role of ABHD17A in Increasing the S-Palmitoylation and Antiviral Activity of IFITM1 by Downregulating ABHD16A

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Transient Transfection
2.2. Plasmid Construction
2.3. Immunofluorescence (IF)
2.4. Bimolecular Fluorescence Complementation Assay (BiFC)
2.5. Yeast Two-Hybrid Assay (Y2H)
2.6. Western Blotting (WB)
2.7. Co-Immunoprecipitation (Co-IP)
2.8. GST-ABHD17A Expression and Purification
2.9. GST Pull-Down
2.10. Quantitative RT-PCR (qRT-PCR)
2.11. Luciferase Reporter Assay
2.12. Short Hairpin RNA (shRNA) Interference Experiment
2.13. CRISPR/Cas9-Mediated Knockout of ABHD16A
2.14. Acyl-PEGyl Exchange Gel Shift Assay (APEGS)
2.15. Virus Preparation and Infection
2.16. Enzyme-Linked Immunosorbent Assay (ELISA) of Secreted Hepatitis B e Antigen (HbeAg) and Hepatitis Surface Antigen (HbsAg)
2.17. Extracellular HBV DNA Extraction
2.18. Cell Viability Analysis
2.19. Image Processing and Analysis
2.20. Quantification and Statistical Analysis
3. Results
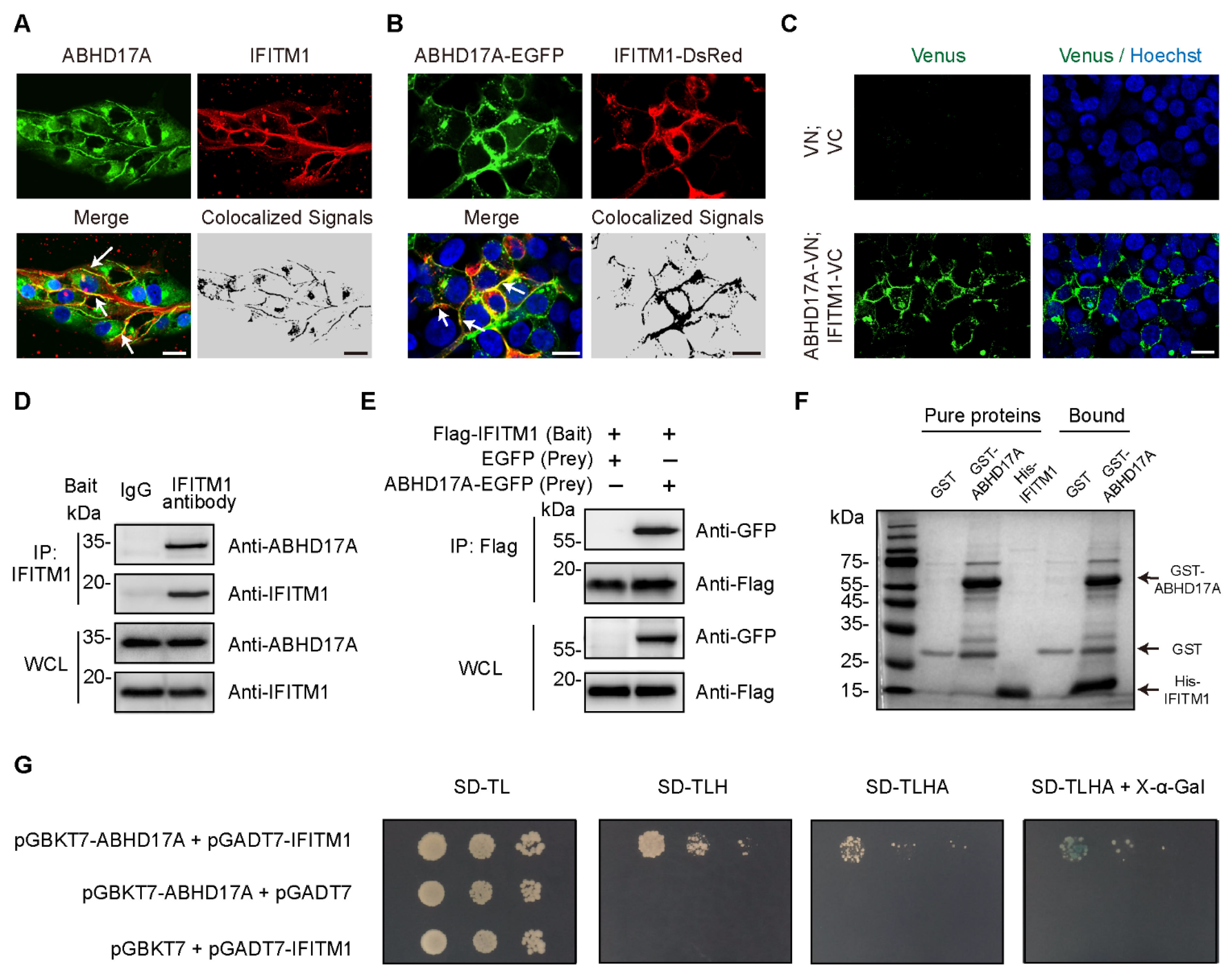
3.1. Identification of IFITM1 as ABHD17A-Interacting Protein
3.2. The Unconventional Role of ABHD17A in Increasing the S-Palmitoylation Modification of IFITM1
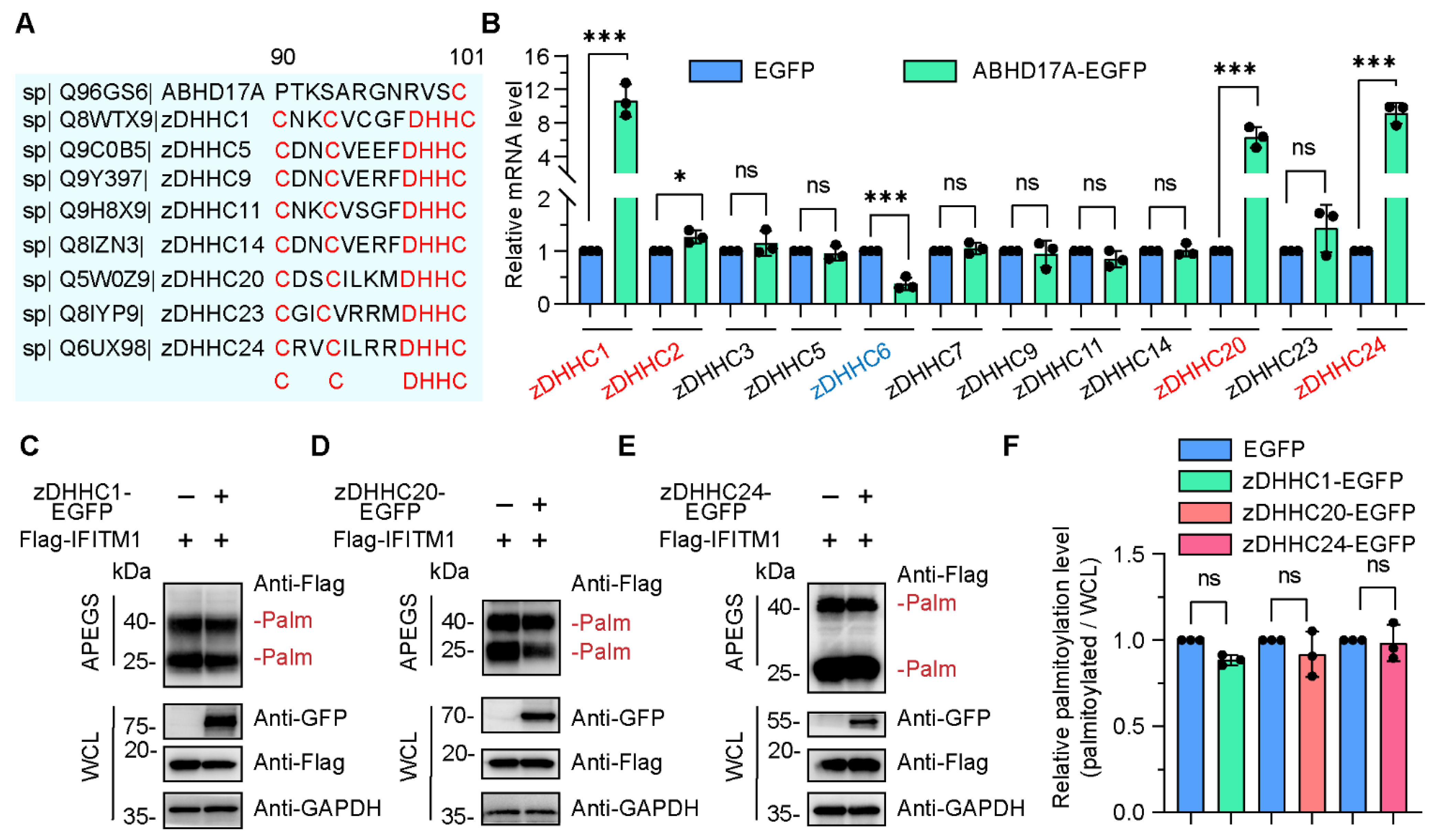
3.3. The zDHHC Family Members Are Dispensable for ABHD17A-Mediated S-Palmitoylation of IFITM1
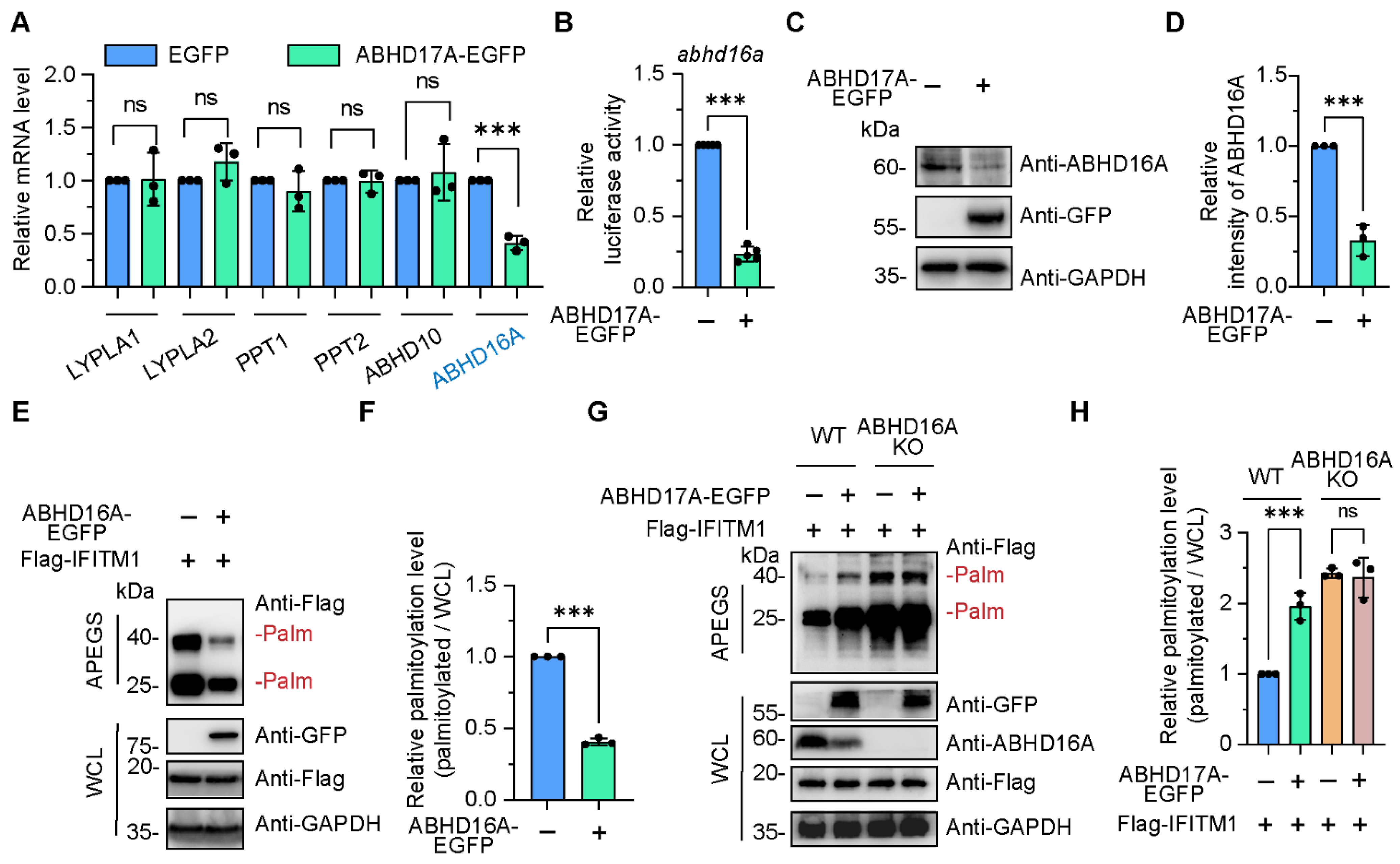
3.4. ABHD17A Promotes the S-Palmitoyl Modification Level of IFITM1 by Downregulating ABHD16A
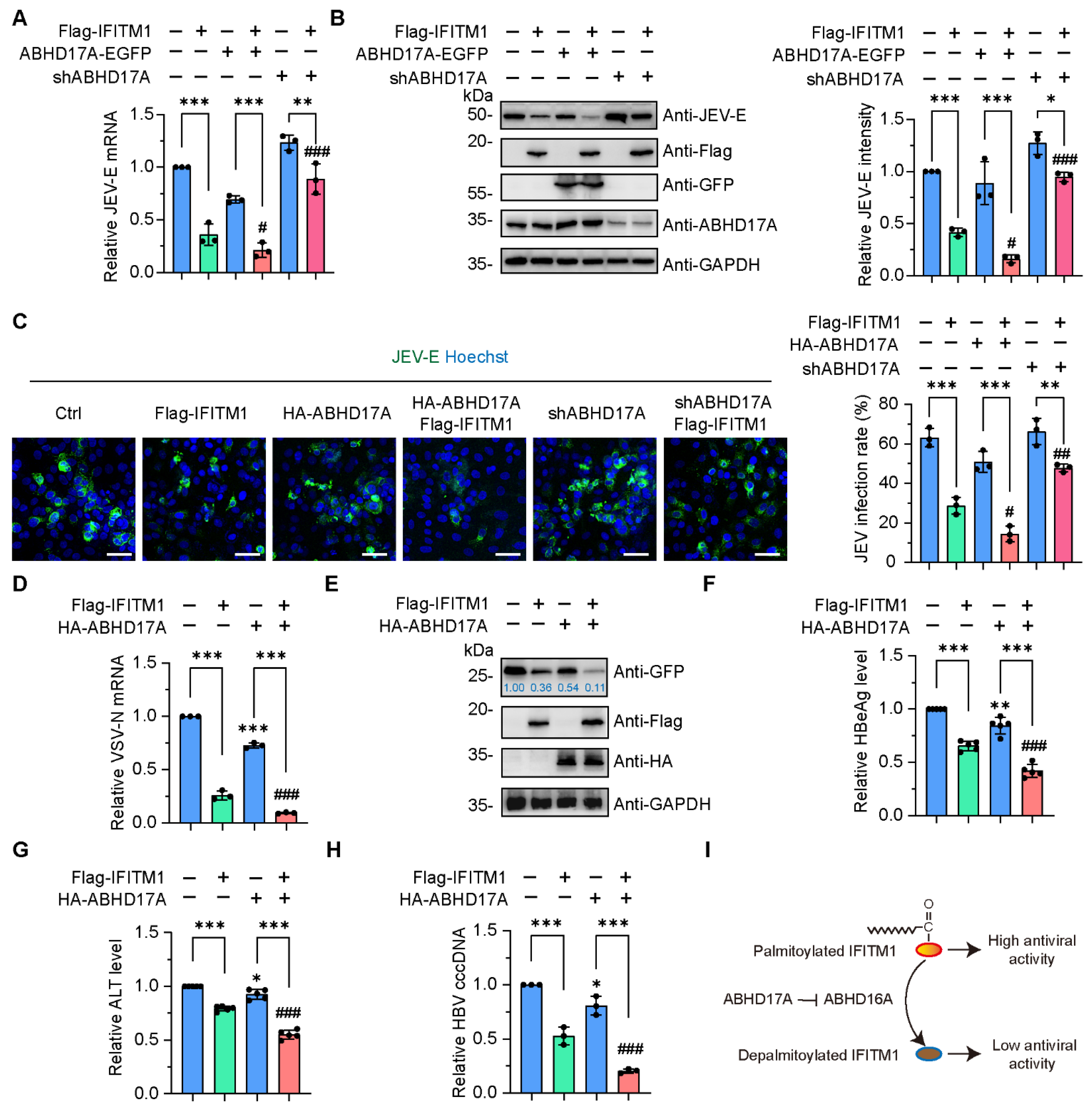
3.5. The Activity of IFITM1 to Restrict Virus Infection Is Positively Regulated by ABHD17A
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABHD | α/β-hydrolase domain |
| IFITMs | interferon-inducible transmembrane proteins |
| zDHHC | zinc finger Asp–His–His–Cys domain |
| PTM | post-translational modification |
| APEGS | acyl-PEGyl exchange gel shift |
| JEV | Japanese encephalitis virus |
| VSV | vesicular stomatitis virus |
| HBV | hepatitis B virus |
References
- Yánez, D.C.; Ross, S.; Crompton, T. The IFITM protein family in adaptive immunity. Immunology 2020, 159, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Herranz, M.; Taylor, J.; Sloan, R.D. IFITM proteins: Understanding their diverse roles in viral infection, cancer, and immunity. J. Biol. Chem. 2023, 299, 102741. [Google Scholar] [CrossRef] [PubMed]
- Buchrieser, J.; Degrelle, S.A.; Couderc, T.; Nevers, Q.; Disson, O.; Manet, C.; Donahue, D.A.; Porrot, F.; Hillion, K.H.; Perthame, E.; et al. IFITM proteins inhibit placental syncytiotrophoblast formation and promote fetal demise. Science 2019, 365, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Shen, L.; Chen, S.; Liu, M.; Wang, J.; Wen, X.; Liu, W.; Mao, L.; Ding, Y.; Yu, L.; et al. Swine RNF5 positively regulates the antiviral activity of IFITM1 by mediating the degradation of ABHD16A. J. Virol. 2025, 99, e0127724. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Li, X.; Xu, Z.; Shen, L.; Ding, Y.; Chen, S.; Mao, L.; Liu, W.; Xu, J. ABHD16A Negatively Regulates the Palmitoylation and Antiviral Function of IFITM Proteins. mBio 2022, 13, e0228922. [Google Scholar] [CrossRef]
- Xu, Z.; Li, X.; Xue, J.; Shen, L.; Zheng, W.; Yin, S.; Xu, J. S-palmitoylation of swine interferon-inducible transmembrane protein is essential for its anti-JEV activity. Virology 2020, 548, 82–92. [Google Scholar] [CrossRef]
- Narayana, S.K.; Helbig, K.J.; McCartney, E.M.; Eyre, N.S.; Bull, R.A.; Eltahla, A.; Lloyd, A.R.; Beard, M.R. The Interferon-induced Transmembrane Proteins, IFITM1, IFITM2, and IFITM3 Inhibit Hepatitis C Virus Entry. J. Biol. Chem. 2015, 290, 25946–25959. [Google Scholar] [CrossRef]
- Chen, H.W.; Zhang, Y.G.; Zhang, W.J.; Su, J.; Wu, H.; Fu, Z.F.; Cui, M. Palmitoylation of hIFITM1 inhibits JEV infection and contributes to BBB stabilization. Int. J. Biol. Macromol. 2024, 262, 129731. [Google Scholar] [CrossRef]
- Anderson, A.M.; Ragan, M.A. Palmitoylation: A protein S-acylation with implications for breast cancer. NPJ Breast Cancer 2016, 2, 16028. [Google Scholar] [CrossRef]
- Mesquita, F.S.; Abrami, L.; Linder, M.E.; Bamji, S.X.; Dickinson, B.C.; van der Goot, F.G. Mechanisms and functions of protein S-acylation. Nat. Rev. Mol. Cell Biol. 2024, 25, 488–509. [Google Scholar] [CrossRef]
- Elliot Murphy, R.; Banerjee, A. In vitro reconstitution of substrate S-acylation by the zDHHC family of protein acyltransferases. Open Biol. 2022, 12, 210390. [Google Scholar] [CrossRef] [PubMed]
- McMichael, T.M.; Zhang, L.; Chemudupati, M.; Hach, J.C.; Kenney, A.D.; Hang, H.C.; Yount, J.S. The palmitoyltransferase ZDHHC20 enhances interferon-induced transmembrane protein 3 (IFITM3) palmitoylation and antiviral activity. J. Biol. Chem. 2017, 292, 21517–21526. [Google Scholar] [CrossRef]
- Won, S.J.; Cheung See Kit, M.; Martin, B.R. Protein depalmitoylases. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.; Liu, Y.; Song, Y.; Song, T.; Wang, T.; Ding, Z.; Liu, J.; Zhao, P. ZDHHC3-LYPLA1 regulates PRRSV-2 replication through reversible palmitoylation. Vet. Microbiol. 2025, 301, 110368. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhou, L.; Xu, Y.; Yang, M.; Xu, Y.; Komaniecki, G.P.; Kosciuk, T.; Chen, X.; Lu, X.; Zou, X.; et al. A STAT3 palmitoylation cycle promotes T(H)17 differentiation and colitis. Nature 2020, 586, 434–439. [Google Scholar] [CrossRef]
- Ni, H.; Wang, Y.; Yao, K.; Wang, L.; Huang, J.; Xiao, Y.; Chen, H.; Liu, B.; Yang, C.Y.; Zhao, J. Cyclical palmitoylation regulates TLR9 signalling and systemic autoimmunity in mice. Nat. Commun. 2024, 15, 1. [Google Scholar] [CrossRef]
- Remsberg, J.R.; Suciu, R.M.; Zambetti, N.A.; Hanigan, T.W.; Firestone, A.J.; Inguva, A.; Long, A.; Ngo, N.; Lum, K.M.; Henry, C.L.; et al. ABHD17 regulation of plasma membrane palmitoylation and N-Ras-dependent cancer growth. Nat. Chem. Biol. 2021, 17, 856–864. [Google Scholar] [CrossRef]
- Cao, Y.; Qiu, T.; Kathayat, R.S.; Azizi, S.A.; Thorne, A.K.; Ahn, D.; Fukata, Y.; Fukata, M.; Rice, P.A.; Dickinson, B.C. ABHD10 is an S-depalmitoylase affecting redox homeostasis through peroxiredoxin-5. Nat. Chem. Biol. 2019, 15, 1232–1240. [Google Scholar] [CrossRef]
- Lehner, B.; Semple, J.I.; Brown, S.E.; Counsell, D.; Campbell, R.D.; Sanderson, C.M. Analysis of a high-throughput yeast two-hybrid system and its use to predict the function of intracellular proteins encoded within the human MHC class III region. Genomics 2004, 83, 153–167. [Google Scholar] [CrossRef]
- Xu, J.; Gu, W.; Ji, K.; Xu, Z.; Zhu, H.; Zheng, W. Sequence analysis and structure prediction of ABHD16A and the roles of the ABHD family members in human disease. Open Biol. 2018, 8. [Google Scholar] [CrossRef]
- Lin, D.T.; Conibear, E. ABHD17 proteins are novel protein depalmitoylases that regulate N-Ras palmitate turnover and subcellular localization. Elife 2015, 4, e11306. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, N.; Fukata, Y.; Sekiya, A.; Murakami, T.; Kobayashi, K.; Fukata, M. Identification of PSD-95 Depalmitoylating Enzymes. J. Neurosci. 2016, 36, 6431–6444. [Google Scholar] [CrossRef] [PubMed]
- McClafferty, H.; Runciman, H.; Shipston, M.J. Site-specific deacylation by ABHD17a controls BK channel splice variant activity. J. Biol. Chem. 2020, 295, 16487–16496. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Que, X.; Wang, S.; Zhou, Q.; Xing, X.; Chen, L.; Hou, C.; Ma, J.; An, P.; Peng, Y.; et al. ZDHHC5-mediated NLRP3 palmitoylation promotes NLRP3-NEK7 interaction and inflammasome activation. Mol. Cell 2023, 83, 4570–4585.e4577. [Google Scholar] [CrossRef]
- Dixon, C.L.; Martin, N.R.; Niphakis, M.J.; Cravatt, B.F.; Fairn, G.D. Attenuating ABHD17 Isoforms Augments the S-acylation and Function of NOD2 and a Subset of Crohn’s Disease-associated NOD2 Variants. Cell Mol. Gastroenterol. Hepatol. 2025, 19, 101491. [Google Scholar] [CrossRef]
- Trindade, B.C.; Chen, G.Y. NOD1 and NOD2 in inflammatory and infectious diseases. Immunol. Rev. 2020, 297, 139–161. [Google Scholar] [CrossRef]
- Miller, K.E.; Kim, Y.; Huh, W.K.; Park, H.O. Bimolecular Fluorescence Complementation (BiFC) Analysis: Advances and Recent Applications for Genome-Wide Interaction Studies. J. Mol. Biol. 2015, 427, 2039–2055. [Google Scholar] [CrossRef]
- Percher, A.; Ramakrishnan, S.; Thinon, E.; Yuan, X.; Yount, J.S.; Hang, H.C. Mass-tag labeling reveals site-specific and endogenous levels of protein S-fatty acylation. Proc. Natl. Acad. Sci. USA 2016, 113, 4302–4307. [Google Scholar] [CrossRef]
- Li, X.; Shen, L.; Xu, Z.; Liu, W.; Li, A.; Xu, J. Protein Palmitoylation Modification During Viral Infection and Detection Methods of Palmitoylated Proteins. Front. Cell Infect. Microbiol. 2022, 12, 821596. [Google Scholar] [CrossRef]
- Lemonidis, K.; Werno, M.W.; Greaves, J.; Diez-Ardanuy, C.; Sanchez-Perez, M.C.; Salaun, C.; Thomson, D.M.; Chamberlain, L.H. The zDHHC family of S-acyltransferases. Biochem. Soc. Trans. 2015, 43, 217–221. [Google Scholar] [CrossRef]
- Tabaczar, S.; Czogalla, A.; Podkalicka, J.; Biernatowska, A.; Sikorski, A.F. Protein palmitoylation: Palmitoyltransferases and their specificity. Exp. Biol. Med. 2017, 242, 1150–1157. [Google Scholar] [CrossRef]
- Fan, Y.; Gao, Y.; Nie, L.; Hou, T.; Dan, W.; Wang, Z.; Liu, T.; Wei, Y.; Wang, Y.; Liu, B.; et al. Targeting LYPLAL1-mediated cGAS depalmitoylation enhances the response to anti-tumor immunotherapy. Mol. Cell 2023, 83, 3520–3532.e3527. [Google Scholar] [CrossRef]
- Zeng, Q.; Liu, J.; Li, Z.; Zhang, Y.; Zu, S.; Ding, X.; Zhang, H. Japanese encephalitis virus NS4B inhibits interferon beta production by targeting TLR3 and TRIF. Vet. Microbiol. 2023, 284, 109849. [Google Scholar] [CrossRef]
- Wen, X.; Liu, M.; Fan, Y.; Xu, J.; Wang, Z.; Mao, L.; Gu, W.; Shi, X.; Xu, J. Depalmitoylase ABHD16A negatively regulates the anti-hepatitis B virus activity of IFITM1. Microbiol. Spectr. 2025, 13, e0309524. [Google Scholar] [CrossRef]
- Chen, W.; Wen, Z.; Zhang, J.; Li, C.; Huang, K.; Bu, Z. Establishing a safe, rapid, convenient and low-cost antiviral assay of interferon bioactivity based on recombinant VSV expressing GFP. J. Virol. Methods 2018, 252, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Fu, Y.; Lin, C.; Shen, Y.; Yu, Q.; Yao, X.; Huang, Q.; Liu, C.; Zeng, Y.; Chen, T.; et al. Oncostatin M Induces IFITM1 Expression to Inhibit Hepatitis B Virus Replication Via JAK-STAT Signaling. Cell Mol. Gastroenterol. Hepatol. 2024, 17, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, E.; Adolfs, Y.; Fukata, M.; Pasterkamp, R.J.; Kapitein, L.C.; Hoogenraad, C.C. Dynamic Palmitoylation Targets MAP6 to the Axon to Promote Microtubule Stabilization during Neuronal Polarization. Neuron 2017, 94, 809–825.e807. [Google Scholar] [CrossRef] [PubMed]
- Mesev, E.V.; LeDesma, R.A.; Ploss, A. Decoding type I and III interferon signalling during viral infection. Nat. Microbiol. 2019, 4, 914–924. [Google Scholar] [CrossRef]
- He, W.R.; Yuan, J.; Ma, Y.H.; Zhao, C.Y.; Yang, Z.Y.; Zhang, Y.; Han, S.; Wan, B.; Zhang, G.P. Modulation of Host Antiviral Innate Immunity by African Swine Fever Virus: A Review. Animals 2022, 12, 2935. [Google Scholar] [CrossRef]
- Chow, K.T.; Gale, M., Jr. SnapShot: Interferon Signaling. Cell 2015, 163, 1808–1808.e1. [Google Scholar] [CrossRef]
- Das, T.; Yang, X.; Lee, H.; Garst, E.H.; Valencia, E.; Chandran, K.; Im, W.; Hang, H.C. S-Palmitoylation and Sterol Interactions Mediate Antiviral Specificity of IFITMs. ACS Chem. Biol. 2022, 17, 2109–2120. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Voeltz, G.K. An ER phospholipid hydrolase drives ER-associated mitochondrial constriction for fission and fusion. Elife 2022, 11, e84279. [Google Scholar] [CrossRef] [PubMed]
- Holme, S.; Sapia, J.; Davey, M.; Vanni, S.; Conibear, E. An S-acylated N-terminus and a conserved loop regulate the activity of the ABHD17 deacylase. J. Cell Biol. 2025, 224, e202405042. [Google Scholar] [CrossRef]
- Ko, P.J.; Dixon, S.J. Protein palmitoylation and cancer. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef]
- Yang, L.; Hou, Y.; Du, Y.E.; Li, Q.; Zhou, F.; Li, Y.; Zeng, H.; Jin, T.; Wan, X.; Guan, S.; et al. Mirtronic miR-4646-5p promotes gastric cancer metastasis by regulating ABHD16A and metabolite lysophosphatidylserines. Cell Death Differ. 2021, 28, 2708–2727. [Google Scholar] [CrossRef] [PubMed]
- Licata, L.; Briganti, L.; Peluso, D.; Perfetto, L.; Iannuccelli, M.; Galeota, E.; Sacco, F.; Palma, A.; Nardozza, A.P.; Santonico, E.; et al. MINT, the molecular interaction database: 2012 update. Nucleic Acids Res. 2012, 40, D857–D861. [Google Scholar] [CrossRef]
- Yu, H.; Tardivo, L.; Tam, S.; Weiner, E.; Gebreab, F.; Fan, C.; Svrzikapa, N.; Hirozane-Kishikawa, T.; Rietman, E.; Yang, X.; et al. Next-generation sequencing to generate interactome datasets. Nat. Methods 2011, 8, 478–480. [Google Scholar] [CrossRef] [PubMed]
- Trojer, P.; Zhang, J.; Yonezawa, M.; Schmidt, A.; Zheng, H.; Jenuwein, T.; Reinberg, D. Dynamic Histone H1 Isotype 4 Methylation and Demethylation by Histone Lysine Methyltransferase G9a/KMT1C and the Jumonji Domain-containing JMJD2/KDM4 Proteins. J. Biol. Chem. 2009, 284, 8395–8405. [Google Scholar] [CrossRef] [PubMed]
- Zaman, M.M.; Nomura, T.; Takagi, T.; Okamura, T.; Jin, W.; Shinagawa, T.; Tanaka, Y.; Ishii, S. Ubiquitination-deubiquitination by the TRIM27-USP7 complex regulates tumor necrosis factor alpha-induced apoptosis. Mol. Cell Biol. 2013, 33, 4971–4984. [Google Scholar] [CrossRef]
- Sobocińska, J.; Roszczenko-Jasińska, P.; Ciesielska, A.; Kwiatkowska, K. Protein Palmitoylation and Its Role in Bacterial and Viral Infections. Front. Immunol. 2017, 8, 2003. [Google Scholar] [CrossRef]
- Das, T.; Yount, J.S.; Hang, H.C. Protein S-palmitoylation in immunity. Open Biol. 2021, 11, 200411. [Google Scholar] [CrossRef]
- Mukai, K.; Konno, H.; Akiba, T.; Uemura, T.; Waguri, S.; Kobayashi, T.; Barber, G.N.; Arai, H.; Taguchi, T. Activation of STING requires palmitoylation at the Golgi. Nat. Commun. 2016, 7, 11932. [Google Scholar] [CrossRef] [PubMed]
- An, C.; Li, Z.; Chen, Y.; Huang, S.; Yang, F.; Hu, Y.; Xu, T.; Zhang, C.; Ge, S. The cGAS-STING pathway in cardiovascular diseases: From basic research to clinical perspectives. Cell Biosci. 2024, 14, 58. [Google Scholar] [CrossRef] [PubMed]
- Claudinon, J.; Gonnord, P.; Beslard, E.; Marchetti, M.; Mitchell, K.; Boularan, C.; Johannes, L.; Eid, P.; Lamaze, C. Palmitoylation of interferon-alpha (IFN-alpha) receptor subunit IFNAR1 is required for the activation of Stat1 and Stat2 by IFN-alpha. J. Biol. Chem. 2009, 284, 24328–24340. [Google Scholar] [CrossRef]
- Ren, W.; Jhala, U.S.; Du, K. Proteomic analysis of protein palmitoylation in adipocytes. Adipocyte 2013, 2, 17–28. [Google Scholar] [CrossRef]
- Kordyukova, L.V.; Serebryakova, M.V.; Baratova, L.A.; Veit, M. Site-specific attachment of palmitate or stearate to cytoplasmic versus transmembrane cysteines is a common feature of viral spike proteins. Virology 2010, 398, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Xia, Q.; Liu, K.; Wu, Z.; Li, C.; Xiao, C.; Dong, N.; Hameed, M.; Anwar, M.N.; Li, Z.; et al. Palmitoylation at Residue C221 of Japanese Encephalitis Virus NS2A Protein Contributes to Viral Replication Efficiency and Virulence. J. Virol. 2023, 97, e0038223. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, X.; Chen, S.; Liu, M.; Fan, Y.; Wen, X.; Wang, J.; Li, X.; Liu, H.; Mao, L.; Yu, L.; et al. The Unconventional Role of ABHD17A in Increasing the S-Palmitoylation and Antiviral Activity of IFITM1 by Downregulating ABHD16A. Biomolecules 2025, 15, 992. https://doi.org/10.3390/biom15070992
Shi X, Chen S, Liu M, Fan Y, Wen X, Wang J, Li X, Liu H, Mao L, Yu L, et al. The Unconventional Role of ABHD17A in Increasing the S-Palmitoylation and Antiviral Activity of IFITM1 by Downregulating ABHD16A. Biomolecules. 2025; 15(7):992. https://doi.org/10.3390/biom15070992
Chicago/Turabian StyleShi, Xuemeng, Shuaiwu Chen, Mingyang Liu, Yali Fan, Xin Wen, Jingyi Wang, Xiaoling Li, Huimin Liu, Lin Mao, Li Yu, and et al. 2025. "The Unconventional Role of ABHD17A in Increasing the S-Palmitoylation and Antiviral Activity of IFITM1 by Downregulating ABHD16A" Biomolecules 15, no. 7: 992. https://doi.org/10.3390/biom15070992
APA StyleShi, X., Chen, S., Liu, M., Fan, Y., Wen, X., Wang, J., Li, X., Liu, H., Mao, L., Yu, L., Hu, Y., & Xu, J. (2025). The Unconventional Role of ABHD17A in Increasing the S-Palmitoylation and Antiviral Activity of IFITM1 by Downregulating ABHD16A. Biomolecules, 15(7), 992. https://doi.org/10.3390/biom15070992