


Human Blood-Derived lncRNAs in Autism Spectrum Disorder

 , , and
, , and
Abstract

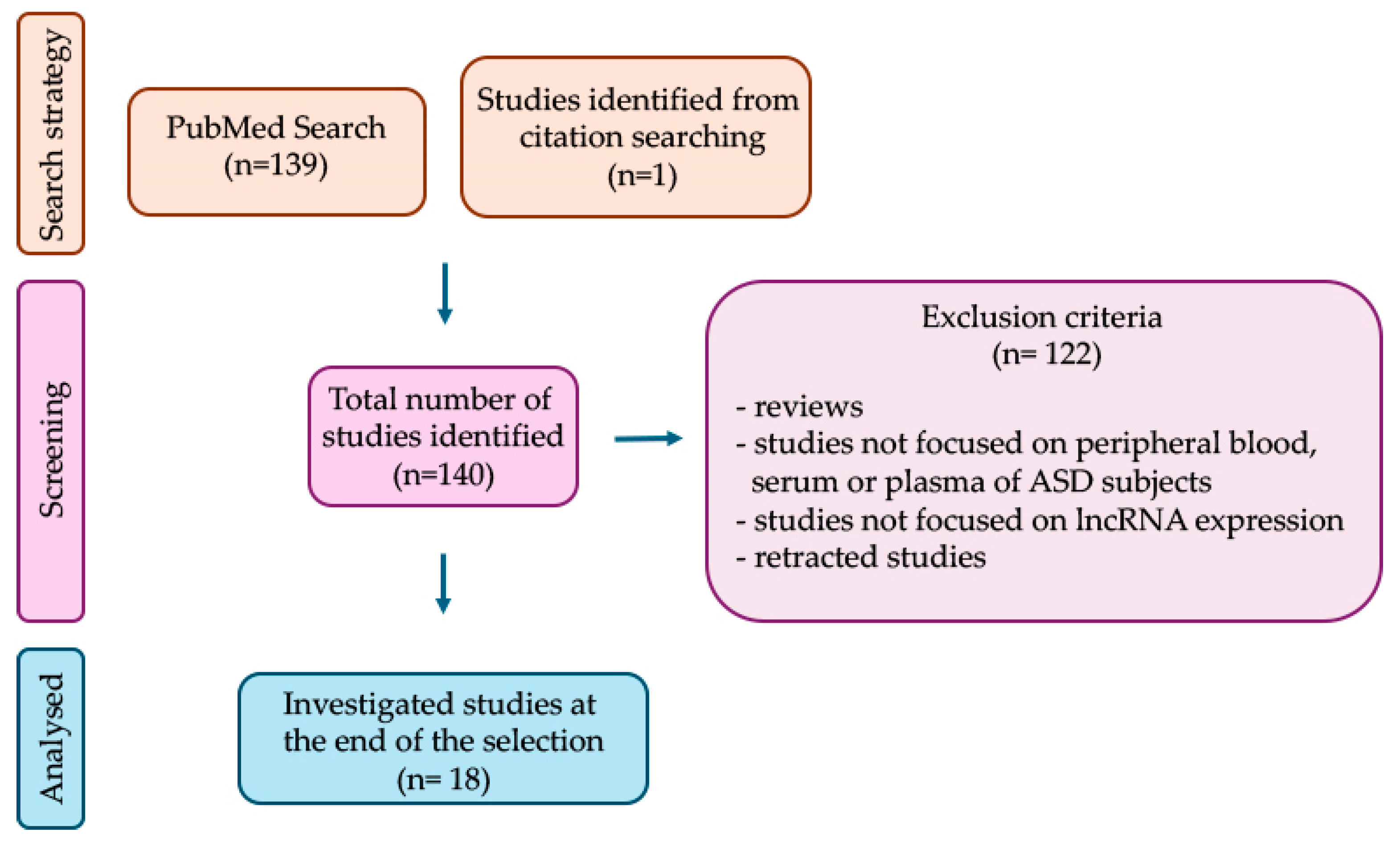
1. Introduction
2. Blood-Derived lncRNAs Dysregulated in Autism Spectrum Disorder (ASD)
2.1. LncRNAs Selected According to ASD-Related Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| lncRNAs | Subjects (Gender and Age) | Source | lncRNA Expression | ASD-Related Pathways Suggested by Authors | Molecular Mechanism in ASD | AUC | Sensitivity | Specificity | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| DISC2 | 30 ASD children (mean age: 6 ± 1.4 years); 41 healthy children (age-, gender-, and ethnicity-matched) | PB | ↑ | Neuronal differentiation | Unknown | AUC = 0.763 (p < 0.0001) | 83.33% | 73.17% | [53] |
| PRKAR2A-AS1 | 30 ASD children (mean age: 6 ± 1.4 years); 41 healthy children (age-, gender-, and ethnicity-matched) | PB | ↑ | Unknown | Unknown | AUC = 0.794 p < 0.0001 | 86.67% | 78.05% | [53] |
| LOC101928237 | 30 ASD children (mean age: 6 ± 1.4 years); 41 healthy children (age-, gender-, and ethnicity-matched) | PB | ↑ | Unknown | Unknown | AUC = 0.9 • (p < 0.0001) | 90% | 82.93% | [53] |
| LRRC2-AS1 | 30 ASD children (mean age: 6 ± 1.4 years); 41 healthy children (age-, gender-, and ethnicity-matched) | PB | ↓ | Unknown | Unknown | AUC = 0.929 • (p < 0.0001) | 86.67% | 100% | [53] |
| SNHG6 | 30 ASD children; 41 healthy children (age-, gender-, and ethnicity-matched; age unspecified) | PB | ↓ | Vitamin D receptor pathway | Interaction with miR-181c | AUC = 0.94 • (p = 0.002) | 60.00% | 73.17% | [57] |
| lincRNA-ROR | 30 ASD children (11 females and 19 males, mean age: 6 ± 1.4 years); 41 healthy children (11 females and 30 males, mean age: 6 ± 1.74) | PB | ↓ | Neuronal differentiation | Unknown | AUC = 0.85 (p < 0.0001) | 86.67% | 65.85% | [60] |
| LINC-PINT | 30 ASD children (11 females and 19 males, mean age: 6 ± 1.4 years); 41 healthy children (11 females and 30 males, mean age: 6 ± 1.74) | PB | ↓ | Neuronal differentiation | Unknown | AUC = 0.67 (p = 0.0138) | / | / | [60] |
| lincRNAp21 | 30 ASD children (11 females and 19 males, mean age: 6 ± 1.4 years); 41 healthy children (11 females and 30 males, mean age: 6 ± 1.74) | PB | ↓ | Neuronal differentiation | Unknown | AUC = 0.64 (p = 0.0394) | / | / | [60] |
| PCAT-29 | 30 ASD children (11 females and 19 males, mean age: 6 ± 1.4 years); 41 healthy children (11 females and 30 males, mean age: 6 ± 1.74) | PB | ↓ | Neuronal differentiation | Unknown | AUC = 0.74 (p = 0.0005) | / | / | [60] |
| PCAT-1 | 30 ASD children (11 females and 19 males, mean age: 6 ± 1.4 years); 41 healthy children (11 females and 30 males, mean age: 6 ± 1.74) | PB | ↓ | Neuronal differentiation | Unknown | AUC = 0.84 (p < 0.0001) | 80% | 70.73% | [60] |
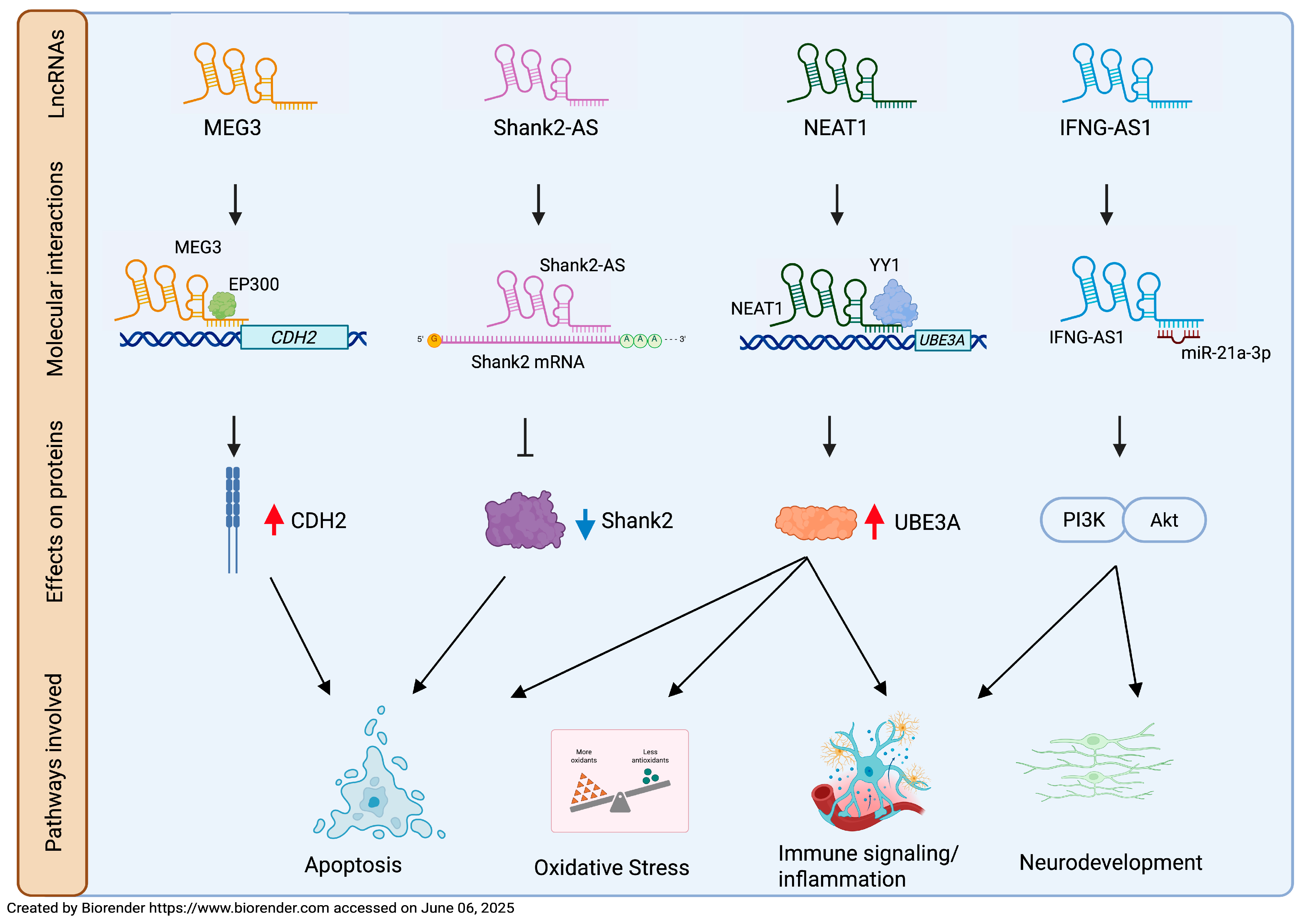
| MEG3 | 30 ASD children (mean age: 6.01 ± 1.39 years); 41 healthy children (age-, gender-, and ethnicity-matched) | PB | ↑ | Neuronal synaptic plasticity; apoptotic pathway | Increases CDH2 expression via EP300, repressing neuronal viability * | AUC = 0.792 (p < 0.0001) | 83.33% | 70.73% | [62] |
| Shank2-AS | 40 ASD children; 40 healthy children (age and gender unspecified) | Lymphocytes from PB | ↑ | Apoptotic pathway | Decreases Shank2 gene expression * | / | / | / | [69] |
| NEAT1 | 30 ASD children (mean age: 6.01 ± 1.4 years); 41 healthy children (mean age: 6 ± 1.4 years, gender-matched) | PB | ↑ | Apoptotic pathway | Interaction with miR-497/BDNF pathway; recruits YY1 to regulate UBE3A expression * | AUC = 0.759 (p < 0.0001) | 70% | 75.61% | [71] |
| TUG1 | 30 ASD children (mean age: 6.01 ± 1.4 years); 41 healthy children (mean age: 6 ± 1.4 years, gender-matched) | PB | ↑ | Apoptotic pathway | Sponge of miR-9 | AUC = 0.733 (p = 0.0001) | 76.67% | 65.85% | [71] |
| 16 ASD children (12 males and 4 females; mean age: 6 ± 1.8 years) 16 healthy controls (11 males and 5 females; mean age: 7.6 ± 2.1 years) | PB mononuclear cells (PBMCs) | ↑ | Protection against oxidative stress | Regulation of TLDC1 expression | / | / | / | [82] | |
| PVT1 | 60 ASD children (mean age: 7.07 ± 2.56 years); 58 healthy children (mean age: 7.76 ± 2.67 years) | serum | ↓ | Neuro- protection | Unknown | AUC = 0.848 (95% CI = 0.78–0.92) | 85.0% | 79.3% | [90] |
| LINC01231 | 30 ASD children (11 females mean age: 6 ± 1.73 years and 19 males, mean age: 6 ± 1.33 years); 41 healthy children (11 females, mean age: 5.63 ± 1.28 years and 30 males, mean age: 6.2 ± 1.88 years) | PB | ↑ | Ca2+ signaling | Unknown | AUC = 0.75 ± 0.06 (p = 0.0003) | 0.77 | 0.76 | [98] |
| THRIL | 10 ASD children; 41 healthy children (age and gender unspecified) | PB | ↓ | Immune signaling/ inflammation | Regulation of TNF-α expression | / | / | / | [114] |
| IFNG-AS1 | 50 ASD children (15 females and 35 males, mean age: 6 ± 1.4 years); 50 healthy controls (14 females and 36 males, mean age: 6 ± 1.74 years) | PB | ↓ | Immune signaling/ inflammation neurogenesis | Regulation of IFNG; regulation of miR-21a-3p/PI3K (p110α)/ AKT axis * | / | / | / | [122] |
| ADINR | 30 ASD children (11 females and 19 males, mean age: 6 ± 1.39 years); 41 healthy children (11 females and 30 males, age- and ethnicity- matched) | PB | ↑ | Immune signaling/ inflammation: NF-κB pathway | Unknown | AUC = 0.735 | / | / | [125] |
| ANRIL | 30 ASD children (11 females and 19 males, mean age: 6 ± 1.39 years); 41 healthy children (11 females and 30 males, age- and ethnicity- matched) | PB | ↑ | Immune signaling/ inflammation: NF-κB pathway | Unknown | AUC = 0.857 | / | / | [125] |
| NKILA | 30 ASD children (11 females and 19 males, mean age: 6 ± 1.39 years); 41 healthy children (11 females and 30 males, age- and ethnicity- matched) | PB | ↑ | Immune signaling/ inflammation: NF-κB pathway | Unknown | AUC = 0.757 | / | / | [125] |
| CCAT1 | 30 ASD children (11 females and 19 males mean age: 6.01 ± 1.39 years); 41 healthy controls (11 females and 30 males (age-, sex-, and ethnicity-matched) | PB | ↓ | Immune signaling/ inflammation | Unknown | AUC = 0.663 (p = 0.016) | 54.33% | 82.93% | [128] |
| CCAT2 | 30 ASD children (11 females and 19 males mean age: 6.01 ± 1.39 years); 41 healthy controls (11 females and 30 males (age-, sex-, and ethnicity-matched) | PB | ↑ | Immune signaling/ inflammation | Unknown | AUC = 0.779 (p < 0.0001) | 86.67% | 73.17% | [128] |
2.2. LncRNAs Selected by High-Throughput Analysis
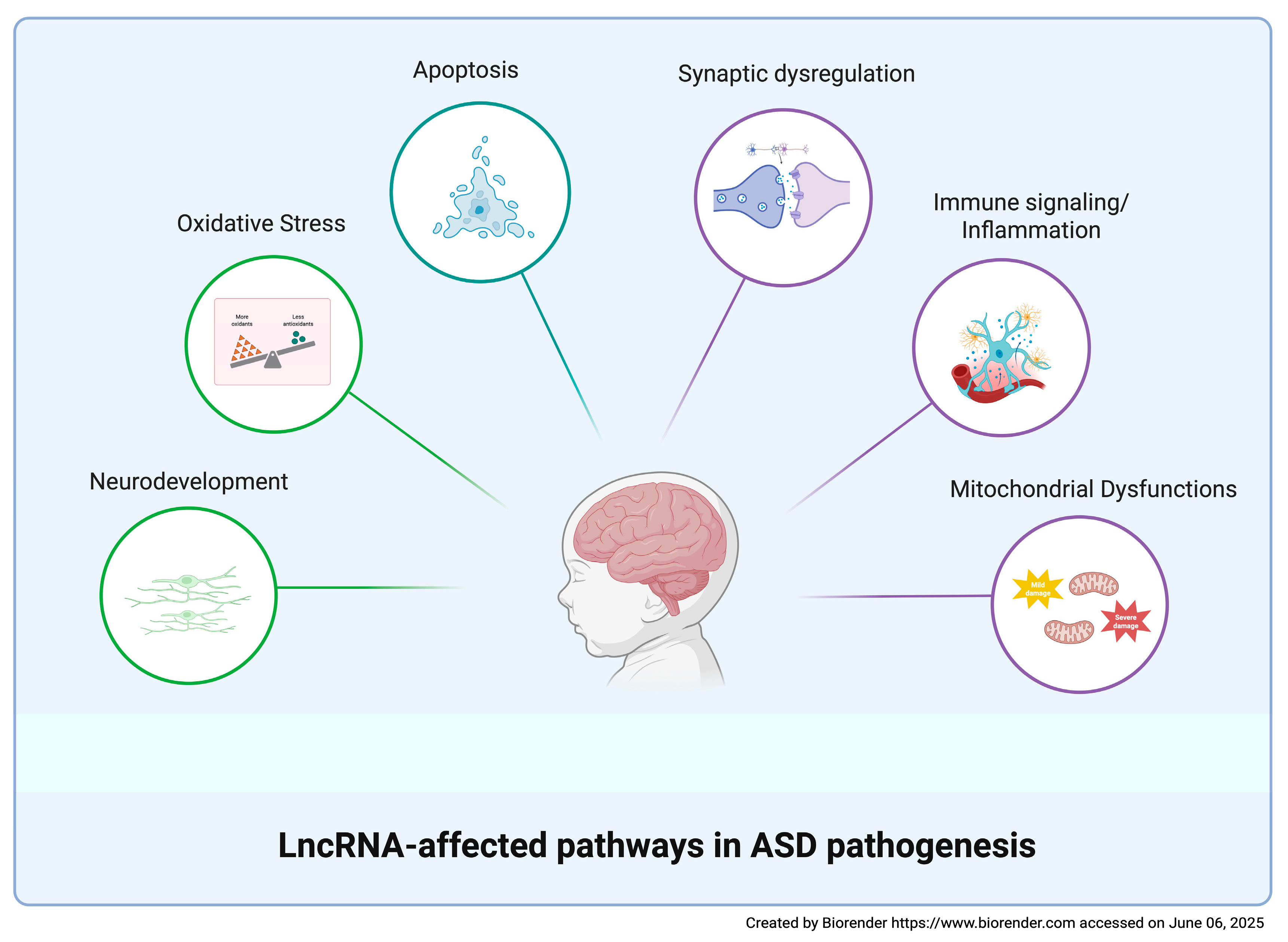
3. Discussion
4. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rogge, N.; Janssen, J. The Economic Costs of Autism Spectrum Disorder: A Literature Review. J. Autism Dev. Disord. 2019, 49, 2873–2900. [Google Scholar] [CrossRef] [PubMed]
- Hirota, T.; King, B.H. Autism Spectrum Disorder: A Review. JAMA 2023, 329, 157–168. [Google Scholar] [CrossRef]
- Lugo Marín, J.; Alviani Rodríguez-Franco, M.; Mahtani Chugani, V.; Magán Maganto, M.; Díez Villoria, E.; Canal Bedia, R. Prevalence of Schizophrenia Spectrum Disorders in Average-IQ Adults with Autism Spectrum Disorders: A Meta-analysis. J. Autism Dev. Disord. 2018, 48, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Capal, J.K.; Jeste, S.S. Autism and Epilepsy. Pediatr. Clin. N. Am. 2024, 71, 241–252. [Google Scholar] [CrossRef]
- Santarone, M.E.; Zambrano, S.; Zanotta, N.; Mani, E.; Minghetti, S.; Pozzi, M.; Villa, L.; Molteni, M.; Zucca, C. EEG Features in Autism Spectrum Disorder: A Retrospective Analysis in a Cohort of Preschool Children. Brain Sci. 2023, 13, 345. [Google Scholar] [CrossRef] [PubMed]
- Pizzolorusso, F.; Paparella, M.T.; Pizzolorusso, I.; Masino, F.; Guglielmi, G. Magnetic Resonance Imaging in Autism Spectrum Disorders: Clinical and neuroradiological phenotypes. Acta Biomed. 2023, 94, e2023027. [Google Scholar]
- Maenner, M.J.; Warren, Z.; Williams, A.R.; Amoakohene, E.; Bakian, A.V.; Bilder, D.A.; Durkin, M.S.; Fitzgerald, R.T.; Furnier, S.M.; Hughes, M.M.; et al. Prevalence and Characteristics of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2020. MMWR Surveill. Summ. 2023, 72, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.R.; Gonda, X.; Tarazi, F.I. Autism Spectrum Disorder: Classification, diagnosis and therapy. Pharmacol. Ther. 2018, 190, 91–104. [Google Scholar] [CrossRef]
- van ‘t Hof, M.; Tisseur, C.; van Berckelear-Onnes, I.; van Nieuwenhuyzen, A.; Daniels, A.M.; Deen, M.; Hoek, H.W.; Ester, W.A. Age at autism spectrum disorder diagnosis: A systematic review and meta-analysis from 2012 to 2019. Autism 2021, 25, 862–873. [Google Scholar] [CrossRef]
- Chlebowski, C.; Green, J.A.; Barton, M.L.; Fein, D. Using the childhood autism rating scale to diagnose autism spectrum disorders. J. Autism Dev. Disord. 2010, 40, 787–799. [Google Scholar] [CrossRef]
- Lord, C.; Elsabbagh, M.; Baird, G.; Veenstra-Vanderweele, J. Autism spectrum disorder. Lancet 2018, 392, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Mandy, W.; Midouhas, E.; Hosozawa, M.; Cable, N.; Sacker, A.; Flouri, E. Mental health and social difficulties of late-diagnosed autistic children, across childhood and adolescence. J. Child Psychol. Psychiatry 2022, 63, 1405–1414. [Google Scholar] [CrossRef] [PubMed]
- Frye, R.E.; Vassall, S.; Kaur, G.; Lewis, C.; Karim, M.; Rossignol, D. Emerging biomarkers in autism spectrum disorder: A systematic review. Ann. Transl. Med. 2019, 7, 792. [Google Scholar] [CrossRef]
- Jensen, A.R.; Lane, A.L.; Werner, B.A.; McLees, S.E.; Fletcher, T.S.; Frye, R.E. Modern Biomarkers for Autism Spectrum Disorder: Future Directions. Mol. Diagn Ther. 2022, 26, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, J.; Religioni, U.; Gellert, B.; Sytnik-Czetwertyński, J.; Pinkas, J. Autism Spectrum Disorders: Etiology, Epidemiology, and Challenges for Public Health. Med. Sci. Monit. 2024, 30, e944161. [Google Scholar] [CrossRef]
- Tick, B.; Bolton, P.; Happé, F.; Rutter, M.; Rijsdijk, F. Heritability of autism spectrum disorders: A meta-analysis of twin studies. J. Child Psychol. Psychiatry 2016, 57, 585–595. [Google Scholar] [CrossRef]
- Wiśniowiecka-Kowalnik, B.; Nowakowska, B.A. Genetics and epigenetics of autism spectrum disorder-current evidence in the field. J. Appl. Genet. 2019, 60, 37–47. [Google Scholar] [CrossRef]
- Rylaarsdam, L.; Guemez-Gamboa, A. Genetic Causes and Modifiers of Autism Spectrum Disorder. Front. Cell. Neurosci. 2019, 13, 385. [Google Scholar] [CrossRef]
- Panisi, C.; Guerini, F.R.; Abruzzo, P.M.; Balzola, F.; Biava, P.M.; Bolotta, A.; Brunero, M.; Burgio, E.; Chiara, A.; Clerici, M.; et al. Autism Spectrum Disorder from the Womb to Adulthood: Suggestions for a Paradigm Shift. J. Pers. Med. 2021, 11, 70. [Google Scholar] [CrossRef]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar]
- Fang, Y.; Fullwood, M.J. Roles, Functions, and Mechanisms of Long Non-coding RNAs in Cancer. Genom. Proteom. Bioinform. 2016, 14, 42–54. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Li, L.; Wen, K. Interactions between long non-coding RNAs and RNA-binding proteins in cancer. Oncol. Rep. 2021, 46, 256. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, J.D.; Wei, Y.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2018, 19, 143–157. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef]
- Dykes, I.M.; Emanueli, C. Transcriptional and Post-transcriptional Gene Regulation by Long Non-coding RNA. Genom. Proteom. Bioinform. 2017, 15, 177–186. [Google Scholar] [CrossRef]
- Ferrer, J.; Dimitrova, N. Transcription regulation by long non-coding RNAs: Mechanisms and disease relevance. Nat. Rev. Mol. Cell Biol. 2024, 25, 396–415. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, H.; Li, L.; Gao, Y.; Yu, B.; Ma, G.; Jin, X.; Sun, Y. New mechanism of LncRNA: In addition to act as a ceRNA. Non-Coding RNA Res. 2024, 9, 1050–1060. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef]
- Yang, C.; Wu, D.; Gao, L.; Liu, X.; Jin, Y.; Wang, D.; Wang, T.; Li, X. Competing endogenous RNA networks in human cancer: Hypothesis, validation, and perspectives. Oncotarget 2016, 7, 13479–13490. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.T.; Lin, J.F.; Li, T.; Li, J.J.; Xu, R.H.; Ju, H.Q. LncRNA-mediated posttranslational modifications and reprogramming of energy metabolism in cancer. Cancer Commun. 2021, 41, 109–120. [Google Scholar] [CrossRef]
- Karakas, D.; Ozpolat, B. The Role of LncRNAs in Translation. Noncoding RNA 2021, 7, 16. [Google Scholar] [CrossRef]
- Heydarnezhad Asl, M.; Pasban Khelejani, F.; Bahojb Mahdavi, S.Z.; Emrahi, L.; Jebelli, A.; Mokhtarzadeh, A. The various regulatory functions of long noncoding RNAs in apoptosis, cell cycle, and cellular senescence. J. Cell Biochem. 2022, 123, 995–1024. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Lim, K.H.; Kim, S.H.; Joo, J.Y. Molecular landscape of long noncoding RNAs in brain disorders. Mol. Psychiatry 2021, 26, 1060–1074. [Google Scholar] [CrossRef] [PubMed]
- Aliperti, V.; Skonieczna, J.; Cerase, A. Long Non-Coding RNA (lncRNA) Roles in Cell Biology, Neurodevelopment and Neurological Disorders. Non-Coding RNA 2021, 7, 36. [Google Scholar] [CrossRef]
- Liaci, C.; Prandi, L.; Pavinato, L.; Brusco, A.; Maldotti, M.; Molineris, I.; Oliviero, S.; Merlo, G.R. The Emerging Roles of Long Non-Coding RNAs in Intellectual Disability and Related Neurodevelopmental Disorders. Int. J. Mol. Sci. 2022, 23, 6118. [Google Scholar] [CrossRef]
- Alammari, F.; Al-Hujaily, E.M.; Alshareeda, A.; Albarakati, N.; Al-Sowayan, B.S. Hidden regulators: The emerging roles of lncRNAs in brain development and disease. Front. Neurosci. 2024, 18, 1392688. [Google Scholar] [CrossRef]
- Anilkumar, A.K.; Vij, P.; Lopez, S.; Leslie, S.M.; Doxtater, K.; Khan, M.M.; Yallapu, M.M.; Chauhan, S.C.; Maestre, G.E.; Tripathi, M.K. Long Non-Coding RNAs: New Insights in Neurodegenerative Diseases. Int. J. Mol. Sci. 2024, 25, 2268. [Google Scholar] [CrossRef]
- Badowski, C.; He, B.; Garmire, L.X. Blood-derived lncRNAs as biomarkers for cancer diagnosis: The Good, the Bad and the Beauty. NPJ Precis. Oncol. 2022, 6, 40. [Google Scholar] [CrossRef]
- Zhang, M.; He, P.; Bian, Z. Long Noncoding RNAs in Neurodegenerative Diseases: Pathogenesis and Potential Implications as Clinical Biomarkers. Front. Mol. Neurosci. 2021, 14, 685143. [Google Scholar] [CrossRef] [PubMed]
- Manna, I.; Quattrone, A.; De Benedittis, S.; Iaccino, E.; Quattrone, A. Roles of Non-Coding RNAs as Novel Diagnostic Biomarkers in Parkinson’s Disease. J. Parkinson’s Dis. 2021, 11, 1475–1489. [Google Scholar] [CrossRef]
- Yang, Y.; Li, Y.; Yang, H.; Guo, J.; Li, N. Circulating MicroRNAs and Long Non-coding RNAs as Potential Diagnostic Biomarkers for Parkinson’s Disease. Front. Mol. Neurosci. 2021, 14, 631553. [Google Scholar] [CrossRef]
- Varesi, A.; Carrara, A.; Pires, V.G.; Floris, V.; Pierella, E.; Savioli, G.; Prasad, S.; Esposito, C.; Ricevuti, G.; Chirumbolo, S.; et al. Blood-Based Biomarkers for Alzheimer’s Disease Diagnosis and Progression: An Overview. Cells 2022, 11, 1367. [Google Scholar] [CrossRef] [PubMed]
- Salloum-Asfar, S.; Elsayed, A.K.; Elhag, S.F.; Abdulla, S.A. Circulating Non-Coding RNAs as a Signature of Autism Spectrum Disorder Symptomatology. Int. J. Mol. Sci. 2021, 22, 6549. [Google Scholar] [CrossRef] [PubMed]
- Bourgeron, T. From the genetic architecture to synaptic plasticity in autism spectrum disorder. Nat. Rev. Neurosci. 2015, 16, 551–563. [Google Scholar] [CrossRef]
- Briggs, J.A.; Wolvetang, E.J.; Mattick, J.S.; Rinn, J.L.; Barry, G. Mechanisms of Long Non-coding RNAs in Mammalian Nervous System Development, Plasticity, Disease, and Evolution. Neuron 2015, 88, 861–877. [Google Scholar] [CrossRef]
- Zimmer-Bensch, G. Emerging Roles of Long Non-Coding RNAs as Drivers of Brain Evolution. Cells 2019, 8, 1399. [Google Scholar] [CrossRef]
- Taylor, M.S.; Devon, R.S.; Millar, J.K.; Porteous, D.J. Evolutionary constraints on the Disrupted in Schizophrenia locus. Genomics 2003, 81, 67–77. [Google Scholar] [CrossRef]
- Millar, J.K.; James, R.; Brandon, N.J.; Thomson, P.A. DISC1 and DISC2: Discovering and dissecting molecular mechanisms underlying psychiatric illness. Ann. Med. 2004, 36, 367–378. [Google Scholar] [CrossRef]
- Lin, M.; Pedrosa, E.; Shah, A.; Hrabovsky, A.; Maqbool, S.; Zheng, D.; Lachman, H.M. RNA-Seq of human neurons derived from iPS cells reveals candidate long non-coding RNAs involved in neurogenesis and neuropsychiatric disorders. PLoS ONE 2011, 6, e23356. [Google Scholar] [CrossRef]
- Williams, J.M.; Beck, T.F.; Pearson, D.M.; Proud, M.B.; Cheung, S.W.; Scott, D.A. A 1q42 deletion involving DISC1, DISC2, and TSNAX in an autism spectrum disorder. Am. J. Med. Genet. A 2009, 149A, 1758–1762. [Google Scholar] [CrossRef] [PubMed]
- Tamizkar, K.H.; Ghafouri-Fard, S.; Omrani, M.D.; Pouresmaeili, F.; Arsang-Jang, S.; Taheri, M. Altered expression of lncRNAs in autism spectrum disorder. Metab. Brain Dis. 2021, 36, 983–990. [Google Scholar] [CrossRef]
- Rahmani, Z.; Rahmani, D.; Jazi, M.S.; Ghasemi, M.R.; Sadeghi, H.; Miryounesi, M.; Razjouyan, K.; Fayyazi Bordbar, M.R. Altered expression of Csnk1a1p in Autism Spectrum Disorder in Iranian population: Case-control study. Sci. Rep. 2024, 14, 28307. [Google Scholar] [CrossRef] [PubMed]
- Kholghi Oskooei, V.; Geranpayeh, L.; Omrani, M.D.; Ghafouri-Fard, S. Assessment of functional variants and expression of long noncoding RNAs in vitamin D receptor signaling in breast cancer. Cancer Manag. Res. 2018, 10, 3451–3462. [Google Scholar] [CrossRef]
- Eyles, D.W.; Burne, T.H.; McGrath, J.J. Vitamin D, effects on brain development, adult brain function and the links between low levels of vitamin D and neuropsychiatric disease. Front. Neuroendocrinol. 2013, 34, 47–64. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Eghtedarian, R.; Hussen, B.M.; Motevaseli, E.; Arsang-Jang, S.; Taheri, M. Expression Analysis of VDR-Related LncRNAs in Autism Spectrum Disorder. J. Mol. Neurosci. 2021, 71, 1403–1409. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Z.; Shu, Q.; Yuan, S.; Xing, Z.; Song, J. LncRNA SNHG6 functions as a ceRNA to regulate neuronal cell apoptosis by modulating miR-181c-5p/BIM signalling in ischaemic stroke. J. Cell. Mol. Med. 2019, 23, 6120–6130. [Google Scholar] [CrossRef] [PubMed]
- Olde Loohuis, N.F.; Kole, K.; Glennon, J.C.; Karel, P.; Van der Borg, G.; Van Gemert, Y.; Van den Bosch, D.; Meinhardt, J.; Kos, A.; Shahabipour, F.; et al. Elevated microRNA-181c and microRNA-30d levels in the enlarged amygdala of the valproic acid rat model of autism. Neurobiol. Dis. 2015, 80, 42–53. [Google Scholar] [CrossRef]
- Sane, S.; Ebrahimi, V.; Shirvani Farsani, Z.; Ghafouri-Fard, S. Assessment of Expression of lncRNAs in Autistic Patients. J. Mol. Neurosci. 2024, 74, 81. [Google Scholar] [CrossRef]
- Tan, M.C.; Widagdo, J.; Chau, Y.Q.; Zhu, T.; Wong, J.J.; Cheung, A.; Anggono, V. The Activity-Induced Long Non-Coding RNA Meg3 Modulates AMPA Receptor Surface Expression in Primary Cortical Neurons. Front. Cell. Neurosci. 2017, 11, 124. [Google Scholar] [CrossRef] [PubMed]
- Taheri, M.; Honarmand Tamizkar, K.; Omrani, S.; Arsang-Jang, S.; Ghafouri-Fard, S.; Omrani, M.D. MEG3 lncRNA is over-expressed in autism spectrum disorder. Metab. Brain Dis. 2021, 36, 2235–2242. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Z.; Zhang, X.; Zhang, D.; Yang, Q.; Hu, P.; Li, F. LncRNA MEG3 activates CDH2 expression by recruitment of EP300 in valproic acid-induced autism spectrum disorder. Neurosci. Lett. 2022, 783, 136726. [Google Scholar] [CrossRef] [PubMed]
- Guang, S.; Pang, N.; Deng, X.; Yang, L.; He, F.; Wu, L.; Chen, C.; Yin, F.; Peng, J. Synaptopathology Involved in Autism Spectrum Disorder. Front. Cell. Neurosci. 2018, 12, 470. [Google Scholar] [CrossRef]
- Wei, H.; Alberts, I.; Li, X. The apoptotic perspective of autism. Int. J. Dev. Neurosci. 2014, 36, 13–18. [Google Scholar] [CrossRef]
- Eftekharian, M.M.; Komaki, A.; Oskooie, V.K.; Namvar, A.; Taheri, M.; Ghafouri-Fard, S. Assessment of Apoptosis Pathway in Peripheral Blood of Autistic Patients. J. Mol. Neurosci. 2019, 69, 588–596. [Google Scholar] [CrossRef]
- Berkel, S.; Marshall, C.R.; Weiss, B.; Howe, J.; Roeth, R.; Moog, U.; Endris, V.; Roberts, W.; Szatmari, P.; Pinto, D.; et al. Mutations in the SHANK2 synaptic scaffolding gene in autism spectrum disorder and mental retardation. Nat. Genet. 2010, 42, 489–491. [Google Scholar] [CrossRef]
- Won, H.; Lee, H.R.; Gee, H.Y.; Mah, W.; Kim, J.I.; Lee, J.; Ha, S.; Chung, C.; Jung, E.S.; Cho, Y.S.; et al. Autistic-like social behaviour in Shank2-mutant mice improved by restoring NMDA receptor function. Nature 2012, 486, 261–265. [Google Scholar] [CrossRef]
- Luo, T.; Liu, P.; Wang, X.Y.; Li, L.Z.; Zhao, L.P.; Huang, J.; Li, Y.M.; Ou, J.L.; Peng, X.Q. Effect of the autism-associated lncRNA Shank2-AS on architecture and growth of neurons. J. Cell Biochem. 2019, 120, 1754–1762. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, X.; Ju, W.; Flory, M.; Zhong, J.; Jiang, S.; Wang, P.; Dong, X.; Tao, X.; Chen, Q.; et al. Genome-wide differential expression of synaptic long noncoding RNAs in autism spectrum disorder. Transl. Psychiatry 2015, 5, e660. [Google Scholar] [CrossRef]
- Sayad, A.; Omrani, M.D.; Fallah, H.; Taheri, M.; Ghafouri-Fard, S. Aberrant Expression of Long Non-coding RNAs in Peripheral Blood of Autistic Patients. J. Mol. Neurosci. 2019, 67, 276–281. [Google Scholar] [CrossRef]
- Peng, W.; Wang, Z.; Fan, H. LncRNA NEAT1 Impacts Cell Proliferation and Apoptosis of Colorectal Cancer via Regulation of Akt Signaling. Pathol. Oncol. Res. 2017, 23, 651–656. [Google Scholar] [CrossRef]
- Li, K.; Wang, Z. lncRNA NEAT1: Key player in neurodegenerative diseases. Ageing Res. Rev. 2023, 86, 101878. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Zhou, H.; Chen, L.; Liu, Z. NEAT1 Promotes Valproic Acid-Induced Autism Spectrum Disorder by Recruiting YY1 to Regulate UBE3A Transcription. Mol. Neurobiol. 2025, 62, 846–860. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J. Long non-coding RNA nuclear paraspeckle assembly transcript 1 inhibits the apoptosis of retina Müller cells after diabetic retinopathy through regulating miR-497/brain-derived neurotrophic factor axis. Diab. Vasc. Dis. Res. 2018, 15, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Saghazadeh, A.; Rezaei, N. Brain-Derived Neurotrophic Factor Levels in Autism: A Systematic Review and Meta-Analysis. J. Autism Dev. Disord. 2017, 47, 1018–1029. [Google Scholar] [CrossRef]
- Chen, S.; Wang, M.; Yang, H.; Mao, L.; He, Q.; Jin, H.; Ye, Z.M.; Luo, X.Y.; Xia, Y.P.; Hu, B. LncRNA TUG1 sponges microRNA-9 to promote neurons apoptosis by up-regulated Bcl2l11 under ischemia. Biochem. Biophys Res. Commun. 2017, 485, 167–173. [Google Scholar] [CrossRef]
- Xiang, P.; Hu, J.; Wang, H.; Luo, Y.; Gu, C.; Tan, X.; Tu, Y.; Guo, W.; Chen, L.; Gao, L.; et al. miR-204-5p is sponged by TUG1 to aggravate neuron damage induced by focal cerebral ischemia and reperfusion injury through upregulating COX2. Cell Death Discov. 2022, 8, 89. [Google Scholar] [CrossRef]
- Wu, H.; Li, Y.; Wang, X.; Zhang, Z.; Huang, Y. Long non-coding RNA TUG1 knockdown prevents neurons from death to alleviate acute spinal cord injury via the microRNA-338/BIK axis. Bioengineered 2021, 12, 5566–5582. [Google Scholar] [CrossRef]
- He, Z.; Zhao, Y.; Zhu, Y.; Wang, W.; Liu, X.; Lu, F. Interfering TUG1 Attenuates Cerebrovascular Endothelial Apoptosis and Inflammatory injury After Cerebral Ischemia/Reperfusion via TUG1/miR-410/FOXO3 ceRNA Axis. Neurotox. Res. 2022, 40, 1–13. [Google Scholar] [CrossRef]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Rivea Morales, D.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [PubMed]
- Zucchini, C.; Serpe, C.; De Sanctis, P.; Ghezzo, A.; Visconti, P.; Posar, A.; Facchin, F.; Marini, M.; Abruzzo, P.M. TLDc Domain-Containing Genes in Autism Spectrum Disorder: New Players in the Oxidative Stress Response. Int. J. Mol. Sci. 2023, 24, 15802. [Google Scholar] [CrossRef]
- Bjørklund, G.; Meguid, N.A.; El-Bana, M.A.; Tinkov, A.A.; Saad, K.; Dadar, M.; Hemimi, M.; Skalny, A.V.; Hosnedlová, B.; Kizek, R.; et al. Oxidative Stress in Autism Spectrum Disorder. Mol. Neurobiol. 2020, 57, 2314–2332. [Google Scholar] [CrossRef]
- Liu, X.; Lin, J.; Zhang, H.; Khan, N.U.; Zhang, J.; Tang, X.; Cao, X.; Shen, L. Oxidative Stress in Autism Spectrum Disorder-Current Progress of Mechanisms and Biomarkers. Front. Psychiatry 2022, 13, 813304. [Google Scholar] [CrossRef]
- Kuźniar-Pałka, A. The Role of Oxidative Stress in Autism Spectrum Disorder Pathophysiology, Diagnosis and Treatment. Biomedicines 2025, 13, 388. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gong, H.Y.; Xu, L. PVT1 protects diabetic peripheral neuropathy via PI3K/AKT pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6905–6911. [Google Scholar] [PubMed]
- Li, C.; Li, R.; Bai, X.; Jiang, H. PVT1 regulates hippocampal neuron apoptosis and inflammation in epilepsy by miR-206-3p-dependent regulation of CAMK4. Gen. Physiol. Biophys. 2024, 43, 423–434. [Google Scholar] [CrossRef]
- Safari, M.R.; Komaki, A.; Arsang-Jang, S.; Taheri, M.; Ghafouri-Fard, S. Expression Pattern of Long Non-coding RNAs in Schizophrenic Patients. Cell. Mol. Neurobiol. 2019, 39, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Li, J.; Guo, M.; Bai, R.; Lei, G.; Sun, H.; Tong, S.; He, K.; He, L. Aberrant expressions of MIAT and PVT1 in serum exosomes of schizophrenia patients. Schizophr. Res. 2022, 240, 71–72. [Google Scholar] [CrossRef]
- Jiang, M.; Chen, G. Investigation of LncRNA PVT1 and MiR-21-5p Expression as Promising Novel Biomarkers for Autism Spectrum Disorder. J. Mol. Neurosci. 2023, 73, 865–873. [Google Scholar] [CrossRef]
- Wu, F.; Huang, W.; Tan, Q.; Guo, Y.; Cao, Y.; Shang, J.; Ping, F.; Wang, W.; Li, Y. ZFP36L2 regulates myocardial ischemia/reperfusion injury and attenuates mitochondrial fusion and fission by LncRNA PVT1. Cell Death Dis. 2021, 12, 614. [Google Scholar] [CrossRef] [PubMed]
- Angelova, P.R.; Abramov, A.Y. Interplay of mitochondrial calcium signalling and reactive oxygen species production in the brain. Biochem. Soc. Trans. 2024, 52, 1939–1946. [Google Scholar] [CrossRef]
- San-Millán, I. The Key Role of Mitochondrial Function in Health and Disease. Antioxidants 2023, 12, 782. [Google Scholar] [CrossRef] [PubMed]
- Citrigno, L.; Muglia, M.; Qualtieri, A.; Spadafora, P.; Cavalcanti, F.; Pioggia, G.; Cerasa, A. The Mitochondrial Dysfunction Hypothesis in Autism Spectrum Disorders: Current Status and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 5785. [Google Scholar] [CrossRef]
- Frye, R.E. Mitochondrial Dysfunction in Autism Spectrum Disorder: Unique Abnormalities and Targeted Treatments. Semin. Pediatr. Neurol. 2020, 35, 100829. [Google Scholar] [CrossRef]
- Khaliulin, I.; Hamoudi, W.; Amal, H. The multifaceted role of mitochondria in autism spectrum disorder. Mol. Psychiatry 2025, 30, 629–650. [Google Scholar] [CrossRef]
- Nguyen, R.L.; Medvedeva, Y.V.; Ayyagari, T.E.; Schmunk, G.; Gargus, J.J. Intracellular calcium dysregulation in autism spectrum disorder: An analysis of converging organelle signaling pathways. Biochim. Biophys Acta Mol. Cell Res. 2018, 1865, 1718–1732. [Google Scholar] [CrossRef]
- Pourtavakoli, A.; Ghafouri-Fard, S.; Eslami, S.; Brand, S.; Taheri, M. Expression assay of calcium signaling related lncRNAs in autism. Mol. Biol. Rep. 2024, 51, 185. [Google Scholar] [CrossRef] [PubMed]
- Soueid, J.; Kourtian, S.; Makhoul, N.J.; Makoukji, J.; Haddad, S.; Ghanem, S.S.; Kobeissy, F.; Boustany, R.M. RYR2, PTDSS1 and AREG genes are implicated in a Lebanese population-based study of copy number variation in autism. Sci. Rep. 2016, 6, 19088. [Google Scholar] [CrossRef]
- Morris, G.; Gevezova, M.; Sarafian, V.; Maes, M. Redox regulation of the immune response. Cell. Mol. Immunol. 2022, 19, 1079–1101. [Google Scholar] [CrossRef]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxidative Med. Cell. Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxidative Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Chen, J.; Li, Y. Microglia and astrocytes underlie neuroinflammation and synaptic susceptibility in autism spectrum disorder. Front. Neurosci. 2023, 17, 1125428. [Google Scholar] [CrossRef]
- Matta, S.M.; Hill-Yardin, E.L.; Crack, P.J. The influence of neuroinflammation in Autism Spectrum Disorder. Brain Behav. Immun. 2019, 79, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Saghazadeh, A.; Ataeinia, B.; Keynejad, K.; Abdolalizadeh, A.; Hirbod-Mobarakeh, A.; Rezaei, N. A meta-analysis of pro-inflammatory cytokines in autism spectrum disorders: Effects of age, gender, and latitude. J. Psychiatr. Res. 2019, 115, 90–102. [Google Scholar] [CrossRef]
- Eissa, N.; Sadeq, A.; Sasse, A.; Sadek, B. Role of Neuroinflammation in Autism Spectrum Disorder and the Emergence of Brain Histaminergic System. Lessons Also for BPSD? Front. Pharmacol. 2020, 11, 886. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, H.; Liu, S.; Luo, W.; Jiang, Y.; Gao, J. Association of Peripheral Blood Levels of Cytokines With Autism Spectrum Disorder: A Meta-Analysis. Front. Psychiatry 2021, 12, 670200. [Google Scholar] [CrossRef]
- Pangrazzi, L.; Cerilli, E.; Balasco, L.; Khurshid, C.; Tobia, C.; Dall’O’, G.M.; Chelini, G.; Perini, S.; Filosi, M.; Barbieri, A.; et al. The interplay between oxidative stress and inflammation supports autistic-related behaviors in Cntnap2 knockout mice. Brain Behav. Immun. 2025, 127, 5–71. [Google Scholar] [CrossRef]
- Than, U.T.T.; Nguyen, L.T.; Nguyen, P.H.; Nguyen, X.H.; Trinh, D.P.; Hoang, D.H.; Nguyen, P.A.T.; Dang, V.D. Inflammatory mediators drive neuroinflammation in autism spectrum disorder and cerebral palsy. Sci. Rep. 2023, 13, 22587. [Google Scholar] [CrossRef]
- Kordulewska, N.K.; Kostyra, E.; Piskorz-Ogórek, K.; Moszyńska, M.; Cieślińska, A.; Fiedorowicz, E.; Jarmołowska, B. Serum cytokine levels in children with spectrum autism disorder: Differences in pro- and anti-inflammatory balance. J. Neuroimmunol. 2019, 337, 577066. [Google Scholar] [CrossRef]
- Lochman, I.; Švachová, V.; Mílková Pavlíková, K.; Medřická, H.; Novák, V.; Trilecová, L.; Pavliska, L.; Procházka, V. Serum Cytokine and Growth Factor Levels in Children with Autism Spectrum Disorder. Med. Sci. Monit. 2018, 24, 2639–2646. [Google Scholar] [CrossRef] [PubMed]
- Abruzzo, P.M.; Matté, A.; Bolotta, A.; Federti, E.; Ghezzo, A.; Guarnieri, T.; Marini, M.; Posar, A.; Siciliano, A.; De Franceschi, L.; et al. Plasma peroxiredoxin changes and inflammatory cytokines support the involvement of neuro-inflammation and oxidative stress in Autism Spectrum Disorder. J. Transl. Med. 2019, 17, 332. [Google Scholar] [CrossRef]
- Petrelli, F.; Pucci, L.; Bezzi, P. Astrocytes and Microglia and Their Potential Link with Autism Spectrum Disorders. Front. Cell. Neurosci. 2016, 10, 21. [Google Scholar] [CrossRef]
- Xie, J.; Huang, L.; Li, X.; Li, H.; Zhou, Y.; Zhu, H.; Pan, T.; Kendrick, K.M.; Xu, W. Immunological cytokine profiling identifies TNF-α as a key molecule dysregulated in autistic children. Oncotarget 2017, 8, 82390–82398. [Google Scholar] [CrossRef]
- Li, Z.; Chao, T.C.; Chang, K.Y.; Lin, N.; Patil, V.S.; Shimizu, C.; Head, S.R.; Burns, J.C.; Rana, T.M. The long noncoding RNA THRIL regulates TNFα expression through its interaction with hnRNPL. Proc. Natl. Acad. Sci. USA 2014, 111, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.A.; Wapinski, O.L.; Yang, Y.W.; Bureau, J.F.; Gopinath, S.; Monack, D.M.; Chang, H.Y.; Brahic, M.; Kirkegaard, K. The NeST long ncRNA controls microbial susceptibility and epigenetic activation of the interferon-γ locus. Cell 2013, 152, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Peng, H.; Tian, J.; Ma, J.; Tang, X.; Rui, K.; Tian, X.; Wang, Y.; Chen, J.; Lu, L.; et al. Upregulation of long noncoding RNA TMEVPG1 enhances T helper type 1 cell response in patients with Sjögren syndrome. Immunol. Res. 2016, 64, 489–496. [Google Scholar] [CrossRef]
- Spurlock, C.F.; Shaginurova, G.; Tossberg, J.T.; Hester, J.D.; Chapman, N.; Guo, Y.; Crooke, P.S.; Aune, T.M. Profiles of Long Noncoding RNAs in Human Naive and Memory T Cells. J. Immunol. 2017, 199, 547–558. [Google Scholar] [CrossRef]
- De Giacomo, A.; Gargano, C.D.; Simone, M.; Petruzzelli, M.G.; Pedaci, C.; Giambersio, D.; Margari, L.; Ruggieri, M. B and T Immunoregulation: A New Insight of B Regulatory Lymphocytes in Autism Spectrum Disorder. Front. Neurosci. 2021, 15, 732611. [Google Scholar] [CrossRef]
- Li, H.; Hao, Y.; Zhang, D.; Fu, R.; Liu, W.; Zhang, X.; Xue, F.; Yang, R. Aberrant expression of long noncoding RNA TMEVPG1 in patients with primary immune thrombocytopenia. Autoimmunity 2016, 49, 496–502. [Google Scholar] [CrossRef]
- Fu, Y.; Zhang, Y.L.; Liu, R.Q.; Xu, M.M.; Xie, J.L.; Zhang, X.L.; Xie, G.M.; Han, Y.T.; Zhang, X.M.; Zhang, W.T.; et al. Exosome lncRNA IFNG-AS1 derived from mesenchymal stem cells of human adipose ameliorates neurogenesis and ASD-like behavior in BTBR mice. J. Nanobiotechnol. 2024, 22, 66. [Google Scholar] [CrossRef]
- Fallah, H.; Sayad, A.; Ranjbaran, F.; Talebian, F.; Ghafouri-Fard, S.; Taheri, M. IFNG/IFNG-AS1 expression level balance: Implications for autism spectrum disorder. Metab. Brain Dis. 2020, 35, 327–333. [Google Scholar] [CrossRef]
- Shih, R.H.; Wang, C.Y.; Yang, C.M. NF-kappaB Signaling Pathways in Neurological Inflammation: A Mini Review. Front. Mol. Neurosci. 2015, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Young, A.M.; Campbell, E.; Lynch, S.; Suckling, J.; Powis, S.J. Aberrant NF-kappaB expression in autism spectrum condition: A mechanism for neuroinflammation. Front. Psychiatry 2011, 2, 27. [Google Scholar] [CrossRef] [PubMed]
- Honarmand Tamizkar, K.; Badrlou, E.; Aslani, T.; Brand, S.; Arsang-Jang, S.; Ghafouri-Fard, S.; Taheri, M. Dysregulation of NF-κB-Associated LncRNAs in Autism Spectrum Disorder. Front. Mol. Neurosci. 2021, 14, 747785. [Google Scholar] [CrossRef] [PubMed]
- Safa, A.; Badrlou, E.; Arsang-Jang, S.; Sayad, A.; Taheri, M.; Ghafouri-Fard, S. Expression of NF-κB associated lncRNAs in schizophrenia. Sci. Rep. 2020, 10, 18105. [Google Scholar] [CrossRef]
- Safa, A.; Noroozi, R.; Taheri, M.; Ghafouri-Fard, S. Association Analysis of ANRIL Polymorphisms and Haplotypes with Autism Spectrum Disorders. J. Mol. Neurosci. 2021, 71, 187–192. [Google Scholar] [CrossRef]
- Taheri, M.; Younesi, Z.; Moradi, S.; Tamizkar, K.H.; Razjouyan, K.; Arsang-Jang, S.; Ghafouri-Fard, S.; Neishabouri, S.M. Altered expression of CCAT1 and CCAT2 lncRNAs in autism spectrum disorder. Gene Rep. 2021, 23, 101172. [Google Scholar] [CrossRef]
- Kaur, A.; Lee, L.H.; Chow, S.C.; Fang, C.M. IRF5-mediated immune responses and its implications in immunological disorders. Int. Rev. Immunol. 2018, 37, 229–248. [Google Scholar] [CrossRef]
- Zhao, C.; Qiao, C.; Zong, L.; Chen, Y. Long non-coding RNA-CCAT2 promotes the occurrence of non-small cell lung cancer by regulating the Wnt/β-catenin signaling pathway. Oncol. Lett. 2018, 16, 4600–4606. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, C.; Tang, X.; Tang, Q.; Lou, L.; Yu, Y.; Zheng, F.; Wu, J.; Yang, X.B.; Wang, W.; et al. The Reciprocal Interaction Between LncRNA CCAT1 and miR-375-3p Contribute to the Downregulation of IRF5 Gene Expression by Solasonine in HepG2 Human Hepatocellular Carcinoma Cells. Front. Oncol. 2019, 9, 1081. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xiang, Y.; Li, F.; Yin, C.; Li, B.; Ke, X. WNT/β-Catenin Signaling Pathway Regulating T Cell-Inflammation in the Tumor Microenvironment. Front. Immunol. 2019, 10, 2293. [Google Scholar] [CrossRef]
- Fang, Y.; Wan, C.; Wen, Y.; Wu, Z.; Pan, J.; Zhong, M.; Zhong, N. Autism-associated synaptic vesicle transcripts are differentially expressed in maternal plasma exosomes of physiopathologic pregnancies. J. Transl. Med. 2021, 19, 154. [Google Scholar] [CrossRef] [PubMed]
- Sabaie, H.; Dehghani, H.; Shiva, S.; Asadi, M.R.; Rezaei, O.; Taheri, M.; Rezazadeh, M. Mechanistic Insight Into the Regulation of Immune-Related Genes Expression in Autism Spectrum Disorder. Front. Mol. Biosci. 2021, 8, 754296. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef]
- Sun, J.J.; Chen, B.; Yu, T. Construction of an immune-related ceRNA network to screen for potential diagnostic markers for autism spectrum disorder. Front. Genet. 2022, 13, 1025813. [Google Scholar] [CrossRef]
- Balasubramanian, R.; Vinod, P.K. Inferring miRNA sponge modules across major neuropsychiatric disorders. Front. Mol. Neurosci. 2022, 15, 1009662. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, Z.; Lu, T.; Li, X.; Yang, K.; Zhao, L.; Zhang, D.; Li, J.; Wang, L. Integrative analysis of long noncoding RNAs dysregulation and synapse-associated ceRNA regulatory axes in autism. Transl. Psychiatry 2023, 13, 375. [Google Scholar] [CrossRef]
- Parellada, M.; Andreu-Bernabeu, Á.; Burdeus, M.; San José Cáceres, A.; Urbiola, E.; Carpenter, L.L.; Kraguljac, N.V.; McDonald, W.M.; Nemeroff, C.B.; Rodriguez, C.I.; et al. In Search of Biomarkers to Guide Interventions in Autism Spectrum Disorder: A Systematic Review. Am. J. Psychiatry 2023, 180, 23–40. [Google Scholar] [CrossRef]
- Qin, Y.; Cao, L.; Zhang, J.; Zhang, H.; Cai, S.; Guo, B.; Wu, F.; Zhao, L.; Li, W.; Ni, L.; et al. Whole-Transcriptome Analysis of Serum L1CAM-Captured Extracellular Vesicles Reveals Neural and Glycosylation Changes in Autism Spectrum Disorder. J. Mol. Neurosci. 2022, 72, 1274–1292. [Google Scholar] [CrossRef]
- Metz, C.E. Basic principles of ROC analysis. Semin. Nucl. Med. 1978, 8, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Perlis, R.H. Translating biomarkers to clinical practice. Mol. Psychiatry 2011, 16, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Abruzzo, P.M.; Ghezzo, A.; Bolotta, A.; Ferreri, C.; Minguzzi, R.; Vignini, A.; Visconti, P.; Marini, M. Perspective Biological Markers for Autism Spectrum Disorders: Advantages of the Use of Receiver Operating Characteristic Curves in Evaluating Marker Sensitivity and Specificity. Dis. Markers 2015, 2015, 329607. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zheng, X.; Liu, P.; Li, M. Clinical value of lncRNA TUG1 in temporal lobe epilepsy and its role in the proliferation of hippocampus neuron via sponging miR-199a-3p. Bioengineered 2021, 12, 10666–10673. [Google Scholar] [CrossRef]
- Mirzajani, S.; Ghafouri-Fard, S.; Habibabadi, J.M.; Arsang-Jang, S.; Omrani, M.D.; Fesharaki, S.S.H.; Sayad, A.; Taheri, M. Expression Analysis of lncRNAs in Refractory and Non-Refractory Epileptic Patients. J. Mol. Neurosci. 2020, 70, 689–698. [Google Scholar] [CrossRef]
- Awwad, D.A. Beyond classic editing: Innovative CRISPR approaches for functional studies of long non-coding RNA. Biol. Methods Protoc. 2019, 4, bpz017. [Google Scholar] [CrossRef]
- Hazan, J.; Bester, A.C. CRISPR-Based Approaches for the High-Throughput Characterization of Long Non-Coding RNAs. Non-Coding RNA 2021, 7, 79. [Google Scholar] [CrossRef]
- Zibitt, M.S.; Hartford, C.C.R.; Lal, A. Interrogating lncRNA functions via CRISPR/Cas systems. RNA Biol 2021, 18, 2097–2106. [Google Scholar] [CrossRef]
- Ross, P.J.; Zhang, W.B.; Mok, R.S.F.; Zaslavsky, K.; Deneault, E.; D’Abate, L.; Rodrigues, D.C.; Yuen, R.K.C.; Faheem, M.; Mufteev, M. Synaptic Dysfunction in Human Neurons With Autism-Associated Deletions in PTCHD1-AS. Biol. Psychiatry 2020, 87, 139–149. [Google Scholar] [CrossRef]
- Zhang, T.N.; Wen, R.; Yang, Y.H.; Yang, N.; Liu, C.F. Integration of transcriptomic, proteomic, and metabolomic data to identify lncRNA rPvt1 associations in lipopolysaccharide-treated H9C2 cardiomyocytes. Front. Genet. 2023, 14, 1278830. [Google Scholar] [CrossRef]
- Teng, P.; Li, Y.; Ku, L.; Wang, F.; Goldsmith, D.R.; Wen, Z.; Yao, B.; Feng, Y. The human lncRNA GOMAFU suppresses neuronal interferon response pathways affected in neuropsychiatric diseases. Brain Behav. Immun. 2023, 112, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Deng, S.; Ge, T.; Ye, M.; Yu, J.; Lin, S.; Ma, W.; Songyang, Z. Live cell imaging and proteomic profiling of endogenous NEAT1 lncRNA by CRISPR/Cas9-mediated knock-in. Protein Cell 2020, 11, 641–660. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serpe, C.; De Sanctis, P.; Marini, M.; Canaider, S.; Abruzzo, P.M.; Zucchini, C. Human Blood-Derived lncRNAs in Autism Spectrum Disorder. Biomolecules 2025, 15, 937. https://doi.org/10.3390/biom15070937
Serpe C, De Sanctis P, Marini M, Canaider S, Abruzzo PM, Zucchini C. Human Blood-Derived lncRNAs in Autism Spectrum Disorder. Biomolecules. 2025; 15(7):937. https://doi.org/10.3390/biom15070937
Chicago/Turabian StyleSerpe, Carmela, Paola De Sanctis, Marina Marini, Silvia Canaider, Provvidenza Maria Abruzzo, and Cinzia Zucchini. 2025. "Human Blood-Derived lncRNAs in Autism Spectrum Disorder" Biomolecules 15, no. 7: 937. https://doi.org/10.3390/biom15070937
APA StyleSerpe, C., De Sanctis, P., Marini, M., Canaider, S., Abruzzo, P. M., & Zucchini, C. (2025). Human Blood-Derived lncRNAs in Autism Spectrum Disorder. Biomolecules, 15(7), 937. https://doi.org/10.3390/biom15070937