

The Impact of PET Imaging on Translational Medicine: Insights from Large-Animal Disease Models

 and
and
Abstract
1. Introduction
2. The PET Scanner for Large Animals
2.1. The Design of the PET Scanner
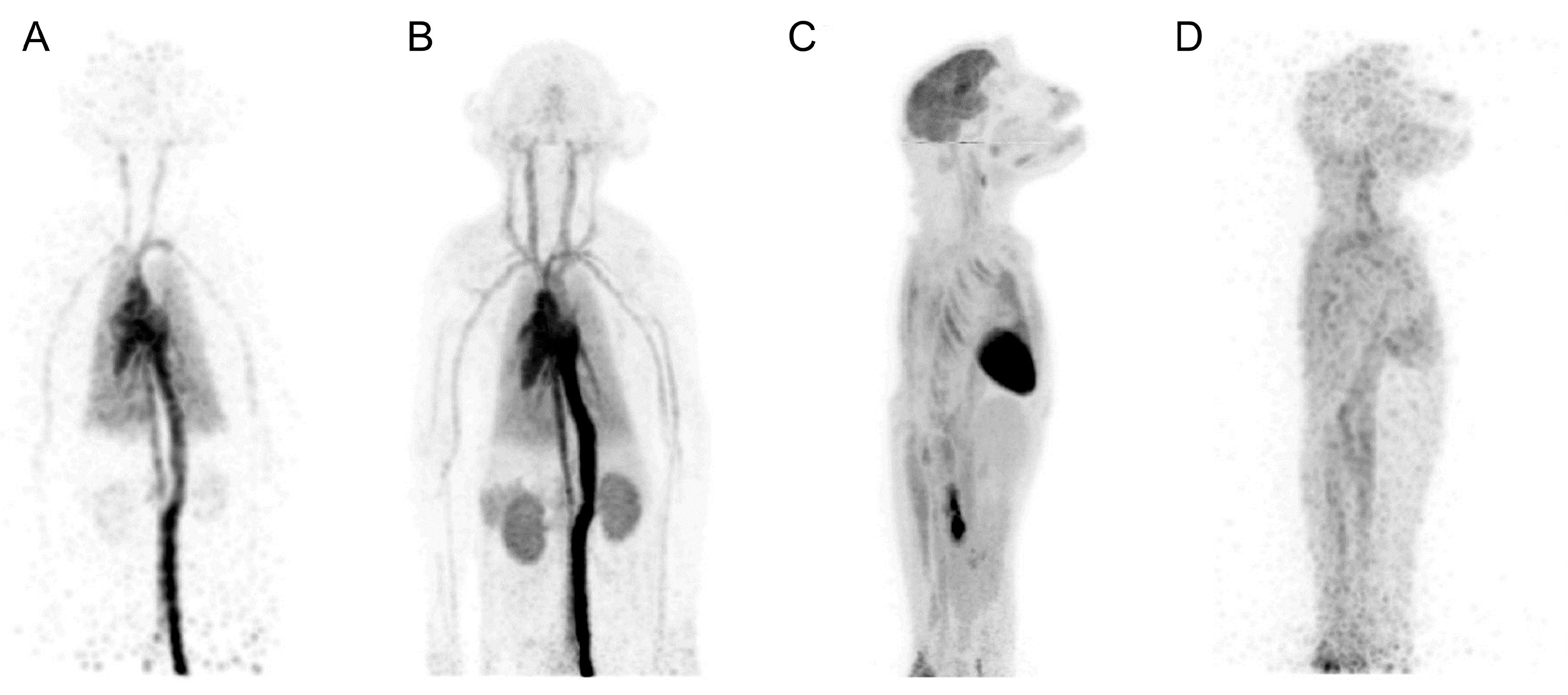
2.2. Dedicated Large-Animal PET
3. Large-Animal Model
3.1. Pig Model
3.2. Non-Human Primate Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | Phenotype | Potential Applications and Significance |
|---|---|---|
| Simian immunodeficiency virus (SIV) [61] | High levels of virus replication associated with a high magnitude of cytokine/chemokine response | Difference between progressive and non-progressive disease courses |
| Simian–human immunodeficiency virus (SHIV) [62] | Lower viral burden and viral control during cART; exhibited less peripheral CD4 depletion and lower gut immune dysfunction and immune activation | SHIV macaque model may be better for identifying initial vaccine candidates |
| Sudan virus [63] | Early stage: viremia, granulocytosis, lymphopenia, albuminemia, thrombocytopenia, and decreased expression of HLA-class transcripts; mid-to-late stage: fever and petechial rashes, high levels of pro-inflammatory mediators and pro-thrombotic factors; end stage: shock and multi-organ failure | The development of vaccines and therapeutics |
| Mycobacterium tuberculosis (Mtb) [64] | Complement C1q increased after Mtb infection; C1q increased after Bacillus Calmette Guérin (BCG) vaccination | C1q can serve as a marker of progressive TB disease |
| Severe acute respiratory syndrome–coronavirus 2 (SARS-CoV-2) [65,66] | Increased levels of monocytes and chemokines; interstitial macrophages accumulate in the lungs | The immune events of the host response and viral replication and disease progression; therapeutic strategies |
| Both intranasal and intragastric inoculation with SARS-CoV-2 caused pneumonia and GI dysfunction | Inflammatory cytokines are possible connections for the pathogenesis of SARS-CoV-2 between the respiratory and digestive systems |
3.3. Sheep Model
4. Application
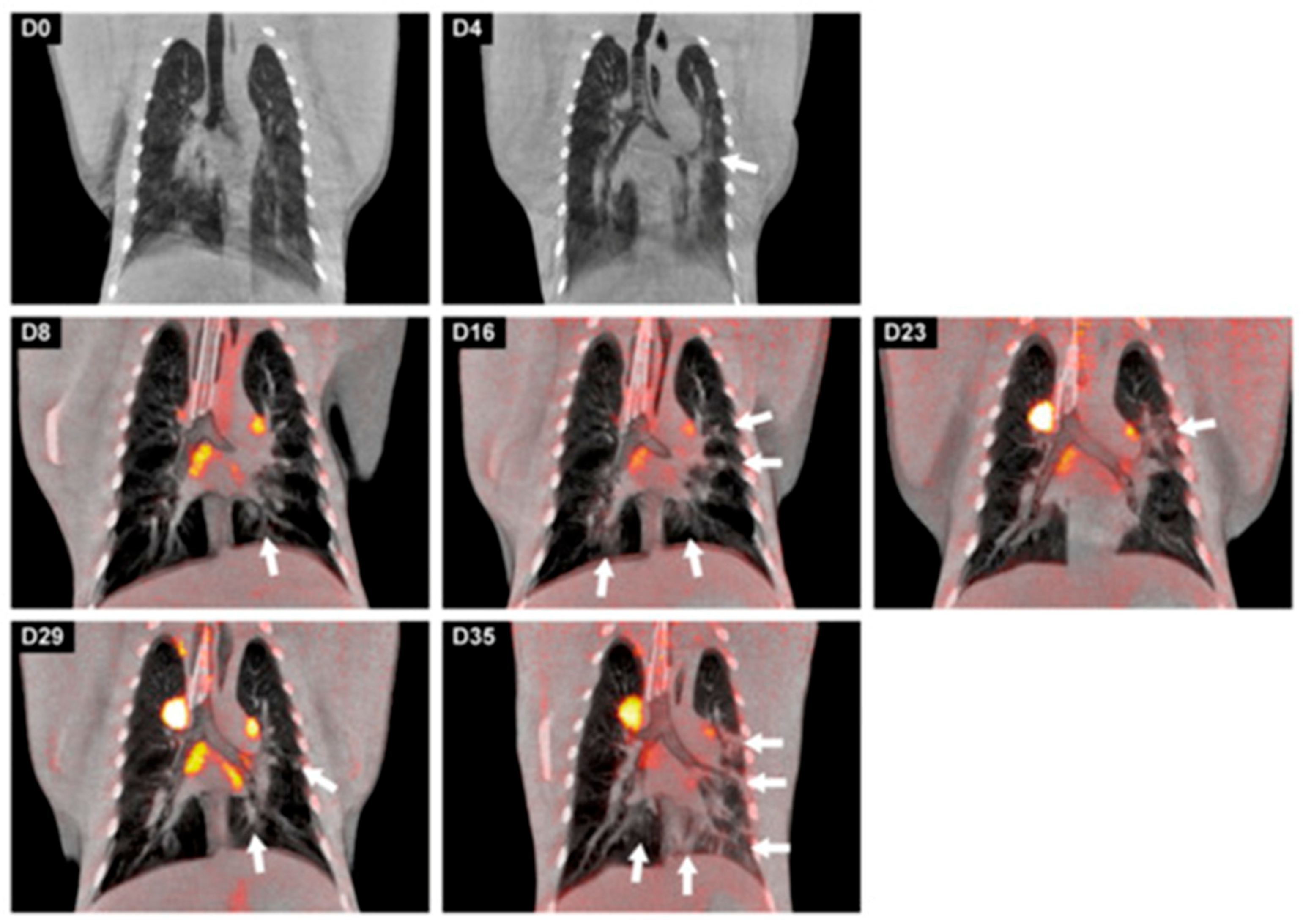
4.1. Infection
4.2. Chronic Heart Disease
4.3. Diabetes
4.4. Cancer
4.5. Central Nervous System Disease
4.5.1. α-Synucleinopathies
4.5.2. Alzheimer’s Disease
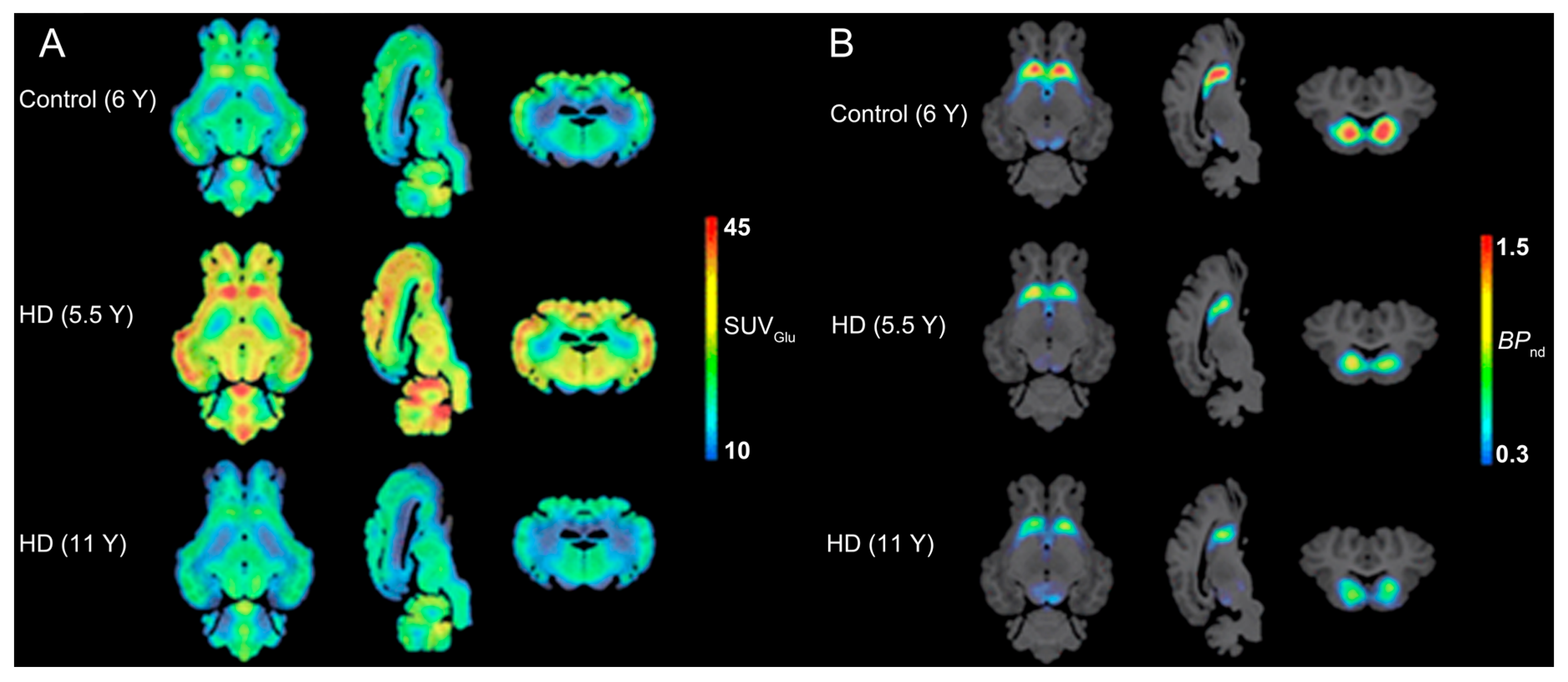
4.5.3. Huntington’s Disease
4.5.4. Psychiatric Disorders
4.6. Addiction
| Tracer | Application | Mechanism | Reference |
|---|---|---|---|
| [11C]acetate | Myocardial perfusion imaging (MPI) | [11C]acetate is rapidly metabolized in cardiomyocytes to acetyl-coenzyme A; acetyl-CoA is a key intermediate in fatty acid oxidation and glucose metabolism in cardiomyocytes | [93] |
| [11C]dihydrotetrabenazine ([11C]DTBZ) | Parkinson’s disease (PD) | [11C]DTBZ binds to the VMAT2 of synaptic vesicles in monoaminergic neurons, which reflects the density and functional state of VMAT2 | [116,117] |
| 11C-labeled Pittsburgh compound B ([11C]PIB) | Alzheimer’s disease (AD) | [11C]PIB is specifically designed to bind to β-amyloid plaques in the brain | [120,121] |
| [11C]raclopride | Addiction | D2R sensitivity to reward decreases in addiction; [11C]raclopride can Quantify dopamine D2R availability in the brain | [141,144] |
| [11C]AZ12204657 and [11C]MK-7246 | Diabetes | Target to GPR44 to β-cell imaging | [96] |
| [N-methyl-11C]-cholylsarcosine ([11C]CSar) | Cancer | [11C]CSar can assess bile acid excretion in the liver | [110] |
| [11C]MPC-6827 | AD | MPC-6827 is a microtubule-targeting agent that binds to tubulin sites with high affinity | [122] |
| [11C]GSK215083 | AD | [11C]GSK215083 is a selective 5-HT6 tracer | [124] |
| [11C]BIO-1819578 | AD | BIO-1819578 is an O-GlcNAcase PET ligand that is an enzyme associated with the development of τ | [126] |
| [11C]PHNO | Addiction (nicotine) | [11C]PHNO has a higher affinity for D3 vs. D2 DA receptors, which allows for regional interpretation of D3 and D2 receptors | [144] |
| [11C]CHDI-180R, [11C]CHDI-626 and [11C]CHDI-650 | Huntington disease (HD) | [11C]CHDI-180R, [11C]CHDI-626, and [11C]CHDI-650 are mHTT aggregate-specific PET ligands | [131,132,133] |
| [13N]NH3 | MPI | [13N]NH3 can be taken up by cardiomyocytes via an amino acid transport system, and its uptake is proportional to myocardial blood flow | [90,94] |
| [15O]H2O | MPI | [15O]H2O is a freely diffusible and metabolically inert tracer; can be used to establish myocardial blood flow | [91,92,93] |
| [18F]FDG | Cancer, tuberculosis, COVID-19 infection, and psychiatric disorders | Increased glucose metabolism in tumor tissue or inflammation | [79,81,82,83,136,138] |
| Myocardial viability imaging, AD, HD, and α-syn disease | Decreased glucose metabolism in damaged cardiomyocytes, AD brain, or α-syn disease brain | [36,114,115,116,129] | |
| [18F]F-DOPA | PD | Striatal uptake of [18F]F-DOPA reflects aromatic amino acid decarboxylase activity | [113] |
| [18F]Flurpiridaz | MPI | [18F]Flurpiridaz works by binding to mitochondrial complex 1 in the heart | [94,95] |
| [18F]KS1 | Cancer | [18F]KS1 is a fluoroethoxy furanose ring-containing ascorbate derivative, to track ROS in prostate tumor | [103] |
| [18F]TTDP | Cancer | Target to T cell immunoglobulin and ITIM domain | [105] |
| [18F]BMS-986229 | Cancer | A 18F-labeled macrocyclic peptide-based PET ligand for imaging PD-L1 | [106] |
| [18F]fluoro-2-deoxy-D-galactose ([18F]FDGal) | Cancer | [18F]FDGal can assess galactose metabolism | [110] |
| [18F]fluoro-l-m-tyrosine | Schizophrenia | L-m-tyrosine is an analogue of L-DOPA | [134] |
| [18F]1 | HD | An mHTT aggregate-specific PET ligand | [130] |
| [18F]3 | AD | A BACE1 PET ligand | [125] |
| [18F]92 | AD | A new tracer that is designed to combined Aβ | [123] |
| [18F]MPPF (4-(2′-methoxyphenyl)-1-[2′-(N-2′′-pyridinyl)-p-fluorobenzamido]ethylpiperazine) | Psychiatric disorders | A radioligand employed for imaging 5-HT1A receptors | [135] |
| [18F]FECNT | Addiction (cocaine) | Target to dopamine transporter | [141] |
| [18F]fluoroclebopride | Addiction (cocaine) | An 18F-labeled raclopride | [142] |
| [18F]Fallypride | Psychiatric disorders and addiction (nicotine) | A high-affinity dopamine D2/D3 receptor antagonist | [137,145] |
| 64Cu labeled to Photoactivatable-Green Fluorescent Protein-HIV-BaL | HIV infection | PET/CT and fluorescent microscopy dual-modal imaging | [87] |
| 64Cu labeled to vaccine-loaded nanoparticle | HIV infection | A combination of PET and fluorescence imaging | [88] |
| [68Ga]Ga-DOTATATE | Cancer | Based on somatostatin receptor (SSTR) type 2 | [102] |
| [68Ga]Ga-DO3A-VS-Cys(40)-Exendin-4 | Diabetes | Target to GLP1-R | [32,101] |
| [89Zr]hu5A10 | Cancer | A humanized uncomplexed and catalytically active prostate-specific antigen-targeting IgG1-mAb | [104] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Camacho, P.; Fan, H.; Liu, Z.; He, J.Q. Large Mammalian Animal Models of Heart Disease. J. Cardiovasc Dev. Dis. 2016, 3, 30. [Google Scholar] [CrossRef]
- Pirovano, G.; Roberts, S.; Kossatz, S.; Reiner, T. Optical Imaging Modalities: Principles and Applications in Preclinical Research and Clinical Settings. J. Nucl. Med. 2020, 61, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Ter-Pogossian, M.M.; Phelps, M.E.; Hoffman, E.J.; Mullani, N.A. A positron-emission transaxial tomograph for nuclear imaging (PETT). Radiology 1975, 114, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Catana, C.; Procissi, D.; Wu, Y.; Judenhofer, M.S.; Qi, J.; Pichler, B.J.; Jacobs, R.E.; Cherry, S.R. Simultaneous in vivo positron emission tomography and magnetic resonance imaging. Proc. Natl. Acad. Sci. USA 2008, 105, 3705–3710. [Google Scholar] [CrossRef] [PubMed]
- Zanzonico, P. Positron emission tomography: A review of basic principles, scanner design and performance, and current systems. Semin. Nucl. Med. 2004, 34, 87–111. [Google Scholar] [CrossRef]
- Berg, E.; Cherry, S.R. Innovations in Instrumentation for Positron Emission Tomography. Semin. Nucl. Med. 2018, 48, 311–331. [Google Scholar] [CrossRef]
- Cherry, S.R.; Sorenson, J.A.; Phelps, M.E. chapter 18-Positron Emission Tomography. In Physics in Nuclear Medicine, 4th ed.; Cherry, S.R., Sorenson, J.A., Phelps, M.E., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2012; pp. 307–343. [Google Scholar] [CrossRef]
- Li, G.; Ma, W.; Li, X.; Yang, W.; Quan, Z.; Ma, T.; Wang, J.; Wang, Y.; Kang, F.; Wang, J. Performance Evaluation of the uMI Panorama PET/CT System in Accordance with the National Electrical Manufacturers Association NU 2-2018 Standard. J. Nucl. Med. 2024, 65, 652–658. [Google Scholar] [CrossRef]
- van Sluis, J.; de Jong, J.; Schaar, J.; Noordzij, W.; van Snick, P.; Dierckx, R.; Borra, R.; Willemsen, A.; Boellaard, R. Performance Characteristics of the Digital Biograph Vision PET/CT System. J. Nucl. Med. 2019, 60, 1031–1036. [Google Scholar] [CrossRef]
- Prenosil, G.A.; Sari, H.; Fürstner, M.; Afshar-Oromieh, A.; Shi, K.; Rominger, A.; Hentschel, M. Performance Characteristics of the Biograph Vision Quadra PET/CT System with a Long Axial Field of View Using the NEMA NU 2-2018 Standard. J. Nucl. Med. 2022, 63, 476–484. [Google Scholar] [CrossRef]
- Spencer, B.A.; Berg, E.; Schmall, J.P.; Omidvari, N.; Leung, E.K.; Abdelhafez, Y.G.; Tang, S.; Deng, Z.; Dong, Y.; Lv, Y.; et al. Performance Evaluation of the uEXPLORER Total-Body PET/CT Scanner Based on NEMA NU 2-2018 with Additional Tests to Characterize PET Scanners with a Long Axial Field of View. J. Nucl. Med. 2021, 62, 861–870. [Google Scholar] [CrossRef]
- Nagy, K.; Tóth, M.; Major, P.; Patay, G.; Egri, G.; Häggkvist, J.; Varrone, A.; Farde, L.; Halldin, C.; Gulyás, B. Performance Evaluation of the Small-Animal nanoScan PET/MRI System. J. Nucl. Med. 2013, 54, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Taschereau, R.; Vu, N.T.; Prout, D.L.; Lee, J.; Chatziioannou, A.F. Performance evaluation of HiPET, a high sensitivity and high resolution preclinical PET tomograph. Phys. Med. Biol. 2020, 65, 045009. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Z.; Wang, X.; Ren, N.; Wu, S.; Gao, J.; Zeng, T.; Gao, D.; Zhang, C.; Sang, Z.; Hu, Z.; et al. Design and performance of SIAT aPET: A uniform high-resolution small animal PET scanner using dual-ended readout detectors. Phys. Med. Biol. 2020, 65, 235013. [Google Scholar] [CrossRef]
- Sarnyai, Z.; Nagy, K.; Patay, G.; Molnar, M.; Rosenqvist, G.; Toth, M.; Takano, A.; Gulyas, B.; Major, P.; Halldin, C.; et al. Performance Evaluation of a High-Resolution Nonhuman Primate PET/CT System. J. Nucl. Med. 2019, 60, 1818–1824. [Google Scholar] [CrossRef]
- Chai, P.; Feng, B.T.; Zhang, Z.M.; Tang, H.H.; Liu, S.Q.; Sun, X.L.; Wang, P.L.; Wang, X.M.; Zhao, X.D.; Wei, L. NEMA NU-4 performance evaluation of a non-human primate animal PET. Phys. Med. Biol. 2019, 64, 105018. [Google Scholar] [CrossRef]
- Naidoo-Variawa, S.; Hey-Cunningham, A.J.; Lehnert, W.; Kench, P.L.; Kassiou, M.; Banati, R.; Meikle, S.R. High-resolution imaging of the large non-human primate brain using microPET: A feasibility study. Phys. Med. Biol. 2007, 52, 6627–6638. [Google Scholar] [CrossRef]
- Lv, Y.; Lv, X.Y.; Liu, W.P.; Judenhofer, M.S.; Zwingenberger, A.; Wisner, E.; Berg, E.; McKenney, S.; Leung, E.; Spencer, B.A.; et al. Mini EXPLORER II: A prototype high-sensitivity PET/CT scanner for companion animal whole body and human brain scanning. Phys. Med. Biol. 2019, 64, 075004. [Google Scholar] [CrossRef] [PubMed]
- Boisson, F.; Serriere, S.; Cao, L.J.; Bodard, S.; Pilleri, A.; Thomas, L.; Sportelli, G.; Vercouillie, J.; Emond, P.; Tauber, C.; et al. Performance evaluation of the IRIS XL-220 PET/CT system, a new camera dedicated to non-human primates. Nucl. Med. Biol. 2022, 108, S173. [Google Scholar]
- Berg, E.; Zhang, X.Z.; Bec, J.; Judenhofer, M.S.; Patel, B.; Peng, Q.Y.; Kapusta, M.; Schmand, M.; Casey, M.E.; Tarantal, A.F.; et al. Development and Evaluation of mini-EXPLORER: A Long Axial Field-of-View PET Scanner for Nonhuman Primate Imaging. J. Nucl. Med. 2018, 59, 993–998. [Google Scholar] [CrossRef]
- Curtasu, M.V.; Knudsen, K.E.B.; Callesen, H.; Purup, S.; Stagsted, J.; Hedemann, M.S. Obesity Development in a Miniature Yucatan Pig Model: A Multi-compartmental Metabolomics Study on Cloned and Normal Pigs Fed Restricted or Ad Libitum High-Energy Diets. J. Proteome Res. 2019, 18, 30–47. [Google Scholar] [CrossRef]
- Lunney, J.K.; Van Goor, A.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the pig as a human biomedical model. Sci. Transl. Med. 2021, 13, eabd5758. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Song, K.D.; Kim, H.J.; Park, W.; Kim, J.; Lee, T.; Shin, D.H.; Kwak, W.; Kwon, Y.J.; Sung, S.; et al. Exploring the genetic signature of body size in Yucatan miniature pig. PLoS ONE 2015, 10, e0121732. [Google Scholar] [CrossRef]
- Oberle, R.L.; Das, H.; Wong, S.L.; Chan, K.K.; Sawchuk, R.J. Pharmacokinetics and metabolism of diclofenac sodium in Yucatan miniature pigs. Pharm. Res. 1994, 11, 698–703. [Google Scholar] [CrossRef]
- Tang, H.; Mayersohn, M. Porcine Prediction of Pharmacokinetic Parameters in People: A Pig in a Poke? Drug Metab. Dispos. 2018, 46, 1712–1724. [Google Scholar] [CrossRef]
- Simianer, H.; Kohn, F. Genetic management of the Gottingen Minipig population. J. Pharmacol. Toxicol. Methods 2010, 62, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Reimer, C.; Ha, N.T.; Sharifi, A.R.; Geibel, J.; Mikkelsen, L.F.; Schlather, M.; Weigend, S.; Simianer, H. Assessing breed integrity of Gottingen Minipigs. BMC Genom. 2020, 21, 308. [Google Scholar] [CrossRef] [PubMed]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 738. [Google Scholar] [CrossRef]
- Lignet, F.; Sherbetjian, E.; Kratochwil, N.; Jones, R.; Suenderhauf, C.; Otteneder, M.B.; Singer, T.; Parrott, N. Characterization of Pharmacokinetics in the Gottingen Minipig with Reference Human Drugs: An In Vitro and In Vivo Approach. Pharm. Res. 2016, 33, 2565–2579. [Google Scholar] [CrossRef]
- von Wilmowsky, C.; Stockmann, P.; Metzler, P.; Harsch, I.A.; Amann, K.; Schlegel, K.A. Establishment of a streptozotocin-induced diabetic domestic pig model and a systematic evaluation of pathological changes in the hard and soft tissue over a 12-month period. Clin. Oral. Implant. Res. 2010, 21, 709–717. [Google Scholar] [CrossRef]
- Jensen-Waern, M.; Andersson, M.; Kruse, R.; Nilsson, B.; Larsson, R.; Korsgren, O.; Essén-Gustavsson, B. Effects of streptozotocin-induced diabetes in domestic pigs with focus on the amino acid metabolism. Lab. Anim. 2009, 43, 249–254. [Google Scholar] [CrossRef]
- Nalin, L.; Selvaraju, R.K.; Velikyan, I.; Berglund, M.; Andreasson, S.; Wikstrand, A.; Ryden, A.; Lubberink, M.; Kandeel, F.; Nyman, G.; et al. Positron emission tomography imaging of the glucagon-like peptide-1 receptor in healthy and streptozotocin-induced diabetic pigs. Eur. J. Nucl. Med. Mol. Imaging 2014, 41, 1800–1810. [Google Scholar] [CrossRef]
- Bellinger, D.A.; Merricks, E.P.; Nichols, T.C. Swine models of type 2 diabetes mellitus: Insulin resistance, glucose tolerance, and cardiovascular complications. Ilar J. 2006, 47, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Milani-Nejad, N.; Janssen, P.M. Small and large animal models in cardiac contraction research: Advantages and disadvantages. Pharmacol. Ther. 2014, 141, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Millard, R.W. Induction of functional coronary collaterals in the swine heart. Basic. Res. Cardiol. 1981, 76, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, N.; Koshino, K.; Yokoyama, I.; Miyagawa, S.; Zeniya, T.; Hirano, Y.; Fukuda, H.; Enmi, J.; Sawa, Y.; Knuuti, J.; et al. Experimental pig model of old myocardial infarction with long survival leading to chronic left ventricular dysfunction and remodeling as evaluated by PET. J. Nucl. Med. 2011, 52, 761–768. [Google Scholar] [CrossRef]
- Renner, S.; Fehlings, C.; Herbach, N.; Hofmann, A.; von Waldthausen, D.C.; Kessler, B.; Ulrichs, K.; Chodnevskaja, I.; Moskalenko, V.; Amselgruber, W.; et al. Glucose intolerance and reduced proliferation of pancreatic beta-cells in transgenic pigs with impaired glucose-dependent insulinotropic polypeptide function. Diabetes 2010, 59, 1228–1238. [Google Scholar] [CrossRef]
- Renner, S.; Braun-Reichhart, C.; Blutke, A.; Herbach, N.; Emrich, D.; Streckel, E.; Wunsch, A.; Kessler, B.; Kurome, M.; Bahr, A.; et al. Permanent neonatal diabetes in INS(C94Y) transgenic pigs. Diabetes 2013, 62, 1505–1511. [Google Scholar] [CrossRef]
- Goldfracht, I.; Efraim, Y.; Shinnawi, R.; Kovalev, E.; Huber, I.; Gepstein, A.; Arbel, G.; Shaheen, N.; Tiburcy, M.; Zimmermann, W.H.; et al. Engineered heart tissue models from hiPSC-derived cardiomyocytes and cardiac ECM for disease modeling and drug testing applications. Acta Biomater. 2019, 92, 145–159. [Google Scholar] [CrossRef]
- Liu, S.; Li, K.; Wagner Florencio, L.; Tang, L.; Heallen, T.R.; Leach, J.P.; Wang, Y.; Grisanti, F.; Willerson, J.T.; Perin, E.C.; et al. Gene therapy knockdown of Hippo signaling induces cardiomyocyte renewal in pigs after myocardial infarction. Sci. Transl. Med. 2021, 13, eabd6892. [Google Scholar] [CrossRef]
- Tilemann, L.; Lee, A.; Ishikawa, K.; Aguero, J.; Rapti, K.; Santos-Gallego, C.; Kohlbrenner, E.; Fish, K.M.; Kho, C.; Hajjar, R.J. SUMO-1 gene transfer improves cardiac function in a large-animal model of heart failure. Sci. Transl. Med. 2013, 5, 211ra159. [Google Scholar] [CrossRef]
- Takano, A.; Varrone, A.; Gulyás, B.; Salvadori, P.; Gee, A.; Windhorst, A.; Vercouillie, J.; Bormans, G.; Lammertsma, A.A.; Halldin, C. Guidelines to PET measurements of the target occupancy in the brain for drug development. Eur. J. Nucl. Med. Mol. Imaging 2016, 43, 2255–2262. [Google Scholar] [CrossRef] [PubMed]
- Capuano, S.V., 3rd; Croix, D.A.; Pawar, S.; Zinovik, A.; Myers, A.; Lin, P.L.; Bissel, S.; Fuhrman, C.; Klein, E.; Flynn, J.L. Experimental Mycobacterium tuberculosis infection of cynomolgus macaques closely resembles the various manifestations of human M. tuberculosis infection. Infect. Immun. 2003, 71, 5831–5844. [Google Scholar] [CrossRef]
- Flynn, J.L.; Capuano, S.V.; Croix, D.; Pawar, S.; Myers, A.; Zinovik, A.; Klein, E. Non-human primates: A model for tuberculosis research. Tuberculosis 2003, 83, 116–118. [Google Scholar] [CrossRef]
- Russell, D.G.; Barry, C.E., 3rd; Flynn, J.L. Tuberculosis: What we don’t know can, and does, hurt us. Science 2010, 328, 852–856. [Google Scholar] [CrossRef]
- Chu, X.; Bleasby, K.; Evers, R. Species differences in drug transporters and implications for translating preclinical findings to humans. Expert. Opin. Drug Metab. Toxicol. 2013, 9, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Shalgunov, V.; Xiong, M.; L’Estrade, E.T.; Raval, N.R.; Andersen, I.V.; Edgar, F.G.; Speth, N.R.; Baerentzen, S.L.; Hansen, H.D.; Donovan, L.L.; et al. Blocking of efflux transporters in rats improves translational validation of brain radioligands. EJNMMI Res. 2020, 10, 124. [Google Scholar] [CrossRef]
- Syvänen, S.; Lindhe, O.; Palner, M.; Kornum, B.R.; Rahman, O.; Långström, B.; Knudsen, G.M.; Hammarlund-Udenaes, M. Species differences in blood-brain barrier transport of three positron emission tomography radioligands with emphasis on P-glycoprotein transport. Drug Metab. Dispos. 2009, 37, 635–643. [Google Scholar] [CrossRef]
- Bakken, T.E.; Jorstad, N.L.; Hu, Q.; Lake, B.B.; Tian, W.; Kalmbach, B.E.; Crow, M.; Hodge, R.D.; Krienen, F.M.; Sorensen, S.A.; et al. Comparative cellular analysis of motor cortex in human, marmoset and mouse. Nature 2021, 598, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Lal, R.; Singh, A.; Watts, S.; Chopra, K. Experimental models of Parkinson’s disease: Challenges and Opportunities. Eur. J. Pharmacol. 2024, 980, 176819. [Google Scholar] [CrossRef]
- Le, W.; Sayana, P.; Jankovic, J. Animal Models of Parkinson’s Disease: A Gateway to Therapeutics? Neurotherapeutics 2014, 11, 92–110. [Google Scholar] [CrossRef]
- Deffains, M.; Canron, M.H.; Teil, M.; Li, Q.; Dehay, B.; Bezard, E.; Fernagut, P.O. L-DOPA regulates α-synuclein accumulation in experimental parkinsonism. Neuropathol. Appl. Neurobiol. 2021, 47, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Lillethorup, T.P.; Noer, O.; Alstrup, A.K.O.; Real, C.C.; Stokholm, K.; Thomsen, M.B.; Zaer, H.; Orlowski, D.; Mikkelsen, T.W.; Glud, A.N.; et al. Spontaneous partial recovery of striatal dopaminergic uptake despite nigral cell loss in asymptomatic MPTP-lesioned female minipigs. Neurotoxicology 2022, 91, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Gough, M.; Singh, D.K.; Singh, B.; Kaushal, D.; Mehra, S. System-wide identification of myeloid markers of TB disease and HIV-induced reactivation in the macaque model of Mtb infection and Mtb/SIV co-infection. Front. Immunol. 2022, 13, 777733. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Cheng, P.H.; Banta, H.; Piotrowska-Nitsche, K.; Yang, J.J.; Cheng, E.C.; Snyder, B.; Larkin, K.; Liu, J.; Orkin, J.; et al. Towards a transgenic model of Huntington’s disease in a non-human primate. Nature 2008, 453, 921–924. [Google Scholar] [CrossRef]
- Yang, W.; Liu, Y.; Tu, Z.; Xiao, C.; Yan, S.; Ma, X.; Guo, X.; Chen, X.; Yin, P.; Yang, Z.; et al. CRISPR/Cas9-mediated PINK1 deletion leads to neurodegeneration in rhesus monkeys. Cell Res. 2019, 29, 334–336. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, Y.; Kang, Y.; Yang, W.; Niu, Y.; Guo, X.; Tu, Z.; Si, C.; Wang, H.; Xing, R.; et al. Functional disruption of the dystrophin gene in rhesus monkey using CRISPR/Cas9. Hum. Mol. Genet. 2015, 24, 3764–3774. [Google Scholar] [CrossRef]
- Liu, Z.; Li, X.; Zhang, J.T.; Cai, Y.J.; Cheng, T.L.; Cheng, C.; Wang, Y.; Zhang, C.C.; Nie, Y.H.; Chen, Z.F.; et al. Autism-like behaviours and germline transmission in transgenic monkeys overexpressing MeCP2. Nature 2016, 530, 98–102. [Google Scholar] [CrossRef]
- Mennicken, F.; Zhang, J.; Hoffert, C.; Ahmad, S.; Beaudet, A.; O’Donnell, D. Phylogenetic changes in the expression of delta opioid receptors in spinal cord and dorsal root ganglia. J. Comp. Neurol. 2003, 465, 349–360. [Google Scholar] [CrossRef]
- Porrino, L.J.; Smith, H.R.; Nader, M.A.; Beveridge, T.J. The effects of cocaine: A shifting target over the course of addiction. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2007, 31, 1593–1600. [Google Scholar] [CrossRef]
- Keating, S.M.; Heitman, J.W.; Wu, S.; Deng, X.; Stacey, A.R.; Zahn, R.C.; de la Rosa, M.; Finstad, S.L.; Lifson, J.D.; Piatak, M., Jr.; et al. Magnitude and Quality of Cytokine and Chemokine Storm during Acute Infection Distinguish Nonprogressive and Progressive Simian Immunodeficiency Virus Infections of Nonhuman Primates. J. Virol. 2016, 90, 10339–10350. [Google Scholar] [CrossRef]
- O’Connor, M.A.; Munson, P.V.; Dross, S.E.; Tunggal, H.C.; Lewis, T.B.; Osborn, J.; Peterson, C.W.; Huang, M.L.W.; Moats, C.; Smedley, J.; et al. A Gut Reaction to SIV and SHIV Infection: Lower Dysregulation of Mucosal T Cells during Acute Infection Is Associated with Greater Viral Suppression during cART. Viruses 2021, 13, 1609. [Google Scholar] [CrossRef]
- Woolsey, C.; Fears, A.C.; Borisevich, V.; Agans, K.N.; Dobias, N.S.; Prasad, A.N.; Deer, D.J.; Geisbert, J.B.; Fenton, K.A.; Geisbert, T.W.; et al. Natural history of Sudan ebolavirus infection in rhesus and cynomolgus macaques. Emerg. Microbes Infect. 2022, 11, 1635–1646. [Google Scholar] [CrossRef]
- Dijkman, K.; Lubbers, R.; Borggreven, N.V.; Ottenhoff, T.H.M.; Joosten, S.A.; Trouw, L.A.; Verreck, F.A.W. Systemic and pulmonary C1q as biomarker of progressive disease in experimental non-human primate tuberculosis. Sci. Rep. 2020, 10, 6290. [Google Scholar] [CrossRef]
- Fahlberg, M.D.; Blair, R.V.; Doyle-Meyers, L.A.; Midkiff, C.C.; Zenere, G.; Russell-Lodrigue, K.E.; Monjure, C.J.; Haupt, E.H.; Penney, T.P.; Lehmicke, G.; et al. Cellular events of acute, resolving or progressive COVID-19 in SARS-CoV-2 infected non-human primates. Nat. Commun. 2020, 11, 6078. [Google Scholar] [CrossRef]
- Jiao, L.; Li, H.Y.; Xu, J.W.; Yang, M.L.; Ma, C.X.; Li, J.M.; Zhao, S.W.; Wang, H.X.; Yang, Y.; Yu, W.H.; et al. The Gastrointestinal Tract Is an Alternative Route for SARS-CoV-2 Infection in a Nonhuman Primate Model. Gastroenterology 2021, 160, 1647–1661. [Google Scholar] [CrossRef] [PubMed]
- McCracken, J.M.; Calderon, G.A.; Robinson, A.J.; Sullivan, C.N.; Cosgriff-Hernandez, E.; Hakim, J.C.E. Animal Models and Alternatives in Vaginal Research: A Comparative Review. Reprod. Sci. 2021, 28, 1759–1773. [Google Scholar] [CrossRef] [PubMed]
- Hympanova, L.; Rynkevic, R.; Urbankova, I.; Blacher, S.; de Landsheere, L.; Mackova, K.; Krofta, L.; Deprest, J. Morphological and Functional Changes in the Vagina following Critical Lifespan Events in the Ewe. Gynecol. Obstet. Investig. 2019, 84, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.J.; Mitchell, N.L. The Translational Benefits of Sheep as Large Animal Models of Human Neurological Disorders. Front. Vet. Sci. 2022, 9, 831838. [Google Scholar] [CrossRef] [PubMed]
- Morton, A.J. Large-Brained Animal Models of Huntington’s Disease: Sheep. Methods Mol. Biol. 2018, 1780, 221–239. [Google Scholar] [CrossRef]
- Mageed, M.; Berner, D.; Jülke, H.; Hohaus, C.; Brehm, W.; Gerlach, K. Morphometrical dimensions of the sheep thoracolumbar vertebrae as seen on digitised CT images. Lab. Anim. Res. 2013, 29, 138–147. [Google Scholar] [CrossRef]
- Jacobsen, J.C.; Bawden, C.S.; Rudiger, S.R.; McLaughlan, C.J.; Reid, S.J.; Waldvogel, H.J.; MacDonald, M.E.; Gusella, J.F.; Walker, S.K.; Kelly, J.M.; et al. An ovine transgenic Huntington’s disease model. Human. Mol. Genet. 2010, 19, 1873–1882. [Google Scholar] [CrossRef] [PubMed]
- McKean, N.; McMurray, C.; Handley, R.; Rudiger, S.; Verma, P.; Kelly, J.; Reid, S.; Pearson, J.; Hardy, J.; Gusella, J.; et al. A Sheep Model of Alzheimer’s Disease; Centre for Brain Research: Bengaluru, India, 2023. [Google Scholar] [CrossRef]
- Pouladi, M.A.; Morton, A.J.; Hayden, M.R. Choosing an animal model for the study of Huntington’s disease. Nat. Rev. Neurosci. 2013, 14, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Rockx, B.; Kuiken, T.; Herfst, S.; Bestebroer, T.; Lamers, M.M.; Oude Munnink, B.B.; de Meulder, D.; van Amerongen, G.; van den Brand, J.; Okba, N.M.A.; et al. Comparative pathogenesis of COVID-19, MERS, and SARS in a nonhuman primate model. Science 2020, 368, 1012–1015. [Google Scholar] [CrossRef]
- Singh, D.K.; Singh, B.; Ganatra, S.R.; Gazi, M.; Cole, J.; Thippeshappa, R.; Alfson, K.J.; Clemmons, E.; Gonzalez, O.; Escobedo, R.; et al. Responses to acute infection with SARS-CoV-2 in the lungs of rhesus macaques, baboons and marmosets. Nat. Microbiol. 2021, 6, 73–86. [Google Scholar] [CrossRef]
- Baum, A.; Ajithdoss, D.; Copin, R.; Zhou, A.; Lanza, K.; Negron, N.; Ni, M.; Wei, Y.; Mohammadi, K.; Musser, B.; et al. REGN-COV2 antibodies prevent and treat SARS-CoV-2 infection in rhesus macaques and hamsters. Science 2020, 370, 1110–1115. [Google Scholar] [CrossRef]
- Vogel, A.B.; Kanevsky, I.; Che, Y.; Swanson, K.A.; Muik, A.; Vormehr, M.; Kranz, L.M.; Walzer, K.C.; Hein, S.; Guler, A.; et al. BNT162b vaccines protect rhesus macaques from SARS-CoV-2. Nature 2021, 592, 283–289. [Google Scholar] [CrossRef]
- Böszörményi, K.P.; Stammes, M.A.; Fagrouch, Z.C.; Kiemenyi-Kayere, G.; Niphuis, H.; Mortier, D.; van Driel, N.; Nieuwenhuis, I.; Vervenne, R.A.W.; Haaksma, T.; et al. The Post-Acute Phase of SARS-CoV-2 Infection in Two Macaque Species Is Associated with Signs of Ongoing Virus Replication and Pathology in Pulmonary and Extrapulmonary Tissues. Viruses 2021, 13, 1673. [Google Scholar] [CrossRef] [PubMed]
- Naninck, T.; Kahlaoui, N.; Lemaitre, J.; Maisonnasse, P.; De Mori, A.; Pascal, Q.; Contreras, V.; Marlin, R.; Relouzat, F.; Delache, B.; et al. Computed tomography and [18F]-FDG PET imaging provide additional readouts for COVID-19 pathogenesis and therapies evaluation in non-human primates. iScience 2022, 25, 104101. [Google Scholar] [CrossRef]
- Darrah, P.A.; Zeppa, J.J.; Maiello, P.; Hackney, J.A.; Wadsworth, M.H., 2nd; Hughes, T.K.; Pokkali, S.; Swanson, P.A., 2nd; Grant, N.L.; Rodgers, M.A.; et al. Prevention of tuberculosis in macaques after intravenous BCG immunization. Nature 2020, 577, 95–102. [Google Scholar] [CrossRef]
- White, A.G.; Maiello, P.; Coleman, M.T.; Tomko, J.A.; Frye, L.J.; Scanga, C.A.; Lin, P.L.; Flynn, J.L. Analysis of 18FDG PET/CT Imaging as a Tool for Studying Mycobacterium tuberculosis Infection and Treatment in Non-human Primates. JoVE-J. Vis. Exp. 2017, 127, e56375. [Google Scholar] [CrossRef]
- Ganchua, S.K.C.; Cadena, A.M.; Maiello, P.; Gideon, H.P.; Myers, A.J.; Junecko, B.F.; Klein, E.C.; Lin, P.L.; Mattila, J.T.; Flynn, J.L. Lymph nodes are sites of prolonged bacterial persistence during Mycobacterium tuberculosis infection in macaques. PLoS Pathog. 2018, 14, e1007337. [Google Scholar] [CrossRef] [PubMed]
- Apetrei, C.; Robertson, D.L.; Marx, P.A. The history of SIVS and AIDS: Epidemiology, phylogeny and biology of isolates from naturally SIV infected non-human primates (NHP) in Africa. Front. Biosci. 2004, 9, 225–254. [Google Scholar] [CrossRef]
- Schmitz, J.E.; Korioth-Schmitz, B. Immunopathogenesis of simian immunodeficiency virus infection in nonhuman primates. Curr. Opin. Hiv. Aids 2013, 8, 273–279. [Google Scholar] [CrossRef]
- Kim, I.; Srinivasula, S.; DeGrange, P.; Long, B.; Jang, H.; Carrasquillo, J.A.; Lane, H.C.; Di Mascio, M. Quantitative PET imaging of the CD4 pool in nonhuman primates. Eur. J. Nucl. Med. Mol. Imaging 2022, 50, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.A.; McRaven, M.D.; Carias, A.M.; Anderson, M.R.; Matias, E.; Arainga, M.; Allen, E.J.; Rogers, K.A.; Gupta, S.; Kulkarni, V.; et al. Localization of infection in neonatal rhesus macaques after oral viral challenge. PLoS Pathog. 2021, 17, e1009855. [Google Scholar] [CrossRef]
- Martin, J.T.; Hartwell, B.L.; Kumarapperuma, S.C.; Melo, M.B.; Carnathan, D.G.; Cossette, B.J.; Adams, J.; Gong, S.; Zhang, W.; Tokatlian, T.; et al. Combined PET and whole-tissue imaging of lymphatic-targeting vaccines in non-human primates. Biomaterials 2021, 275, 120868. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xiang, Z.; Wang, F.; Pan, X.; Zhang, Q.; Wang, P.; Jiang, L.; Yuan, H. 13N-NH3 myocardial perfusion imaging with reduced scan duration: A feasibility study in the era of total-body PET/CT. EJNMMI Phys. 2025, 12, 18. [Google Scholar] [CrossRef]
- Wang, J.; Mpharm, S.L.; Liu, T.W.; Zhang, J.M.; Chen, Y.; Li, J.M.; Xu, W.G. Preliminary and Comparative Experiment Study Between 18F-Flurpiridaz and 13N-NH3·H2O Myocardial Perfusion Imaging with PET/CT in Miniature Pigs. Mol. Imaging 2020, 19, 1536012120947506. [Google Scholar] [CrossRef]
- Maaniitty, T.; Knuuti, J.; Saraste, A. 15O-Water PET MPI: Current Status and Future Perspectives. Semin. Nucl. Med. 2020, 50, 238–247. [Google Scholar] [CrossRef]
- Gronman, M.; Tarkia, M.; Stark, C.; Vahasilta, T.; Kiviniemi, T.; Lubberink, M.; Halonen, P.; Kuivanen, A.; Saunavaara, V.; Tolvanen, T.; et al. Assessment of myocardial viability with [O-15]water PET: A validation study in experimental myocardial infarction. J. Nucl. Cardiol. 2021, 28, 1271–1280. [Google Scholar] [CrossRef]
- Ballo, H.; Tarkia, M.; Haavisto, M.; Stark, C.; Strandberg, M.; Vahasilta, T.; Saunavaara, V.; Tolvanen, T.; Teras, M.; Hynninen, V.V.; et al. Determinants of Myocardial Strain in Experimental Chronic Myocardial Infarction. Ultrasound Med. Biol. 2019, 45, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, S.; Mizoguchi, H.; Fukushima, S.; Imanishi, Y.; Watabe, T.; Harada, A.; Sakai, Y.; Sawa, Y. New regional drug delivery system by direct epicardial placement of slow-release prostacyclin agonist promise therapeutic angiogenesis in a porcine chronic myocardial infarction. J. Artif. Organs 2021, 24, 465–472. [Google Scholar] [CrossRef]
- Werner, R.A.; Koshino, K.; Arimitsu, K.; Lapa, C.; Javadi, M.S.; Rowe, S.P.; Nose, N.; Kimura, H.; Fukushima, K.; Higuchi, T. Stability of Distribution of F18 Flurpiridaz After Transient Coronary Occlusion in Pigs. JACC Cardiovasc. Imaging 2019, 12, 2269–2271. [Google Scholar] [CrossRef]
- Eriksson, O. GPR44 as a Target for Imaging Pancreatic Beta-Cell Mass. Curr. Diabetes Rep. 2019, 19, 49. [Google Scholar] [CrossRef]
- Meier, J.J.; Menge, B.A.; Breuer, T.G.; Muller, C.A.; Tannapfel, A.; Uhl, W.; Schmidt, W.E.; Schrader, H. Functional assessment of pancreatic beta-cell area in humans. Diabetes 2009, 58, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, H.; Takahashi, K.; Inaba, W.; Tsuboi, K.; Osonoi, S.; Yoshida, T.; Yagihashi, S. Involvement of oxidative stress-induced DNA damage, endoplasmic reticulum stress, and autophagy deficits in the decline of beta-cell mass in Japanese type 2 diabetic patients. Diabetes Care 2014, 37, 1966–1974. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.J.; Breuer, T.G.; Bonadonna, R.C.; Tannapfel, A.; Uhl, W.; Schmidt, W.E.; Schrader, H.; Menge, B.A. Pancreatic diabetes manifests when beta cell area declines by approximately 65% in humans. Diabetologia 2012, 55, 1346–1354. [Google Scholar] [CrossRef]
- Inaishi, J.; Saisho, Y. Beta-Cell Mass in Obesity and Type 2 Diabetes, and Its Relation to Pancreas Fat: A Mini-Review. Nutrients 2020, 12, 3846. [Google Scholar] [CrossRef]
- Malbert, C.H.; Chauvin, A.; Horowitz, M.; Jones, K.L. Glucose Sensing Mediated by Portal Glucagon-Like Peptide 1 Receptor Is Markedly Impaired in Insulin-Resistant Obese Animals. Diabetes 2021, 70, 99–110. [Google Scholar] [CrossRef]
- Deppen, S.A.; Blume, J.; Bobbey, A.J.; Shah, C.; Graham, M.M.; Lee, P.; Delbeke, D.; Walker, R.C. 68Ga-DOTATATE Compared with 111In-DTPA-Octreotide and Conventional Imaging for Pulmonary and Gastroenteropancreatic Neuroendocrine Tumors: A Systematic Review and Meta-Analysis. J. Nucl. Med. 2016, 57, 872–878. [Google Scholar] [CrossRef]
- Damuka, N.; Bashetti, N.; Mintz, A.; Bansode, A.H.; Miller, M.; Krizan, I.; Furdui, C.; Bhoopal, B.; Gollapelli, K.K.; Shanmukha Kumar, J.V.; et al. [18F]KS1, a novel ascorbate-based ligand images ROS in tumor models of rodents and nonhuman primates. Biomed. Pharmacother. 2022, 156, 113937. [Google Scholar] [CrossRef] [PubMed]
- Veach, D.R.; Storey, C.M.; Lückerath, K.; Braun, K.; von Bodman, C.; Lamminmäki, U.; Kalidindi, T.; Strand, S.E.; Strand, J.; Altai, M.; et al. PSA-Targeted Alpha-, Beta-, and Positron-Emitting Immunotheranostics in Murine Prostate Cancer Models and Nonhuman Primates. Clin. Cancer Res. 2021, 27, 2050–2060. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, X.; Wang, Y.; Li, X.; Sun, Y.; Guan, Z.; Li, X.; Wu, Y.; Wang, J.; Zhao, F.; et al. Development and characterisation of [18F]TTDP, a novel T cell immunoglobulin and ITIM domain tracer, in humanised mice and non-human primates. Eur. J. Nucl. Med. Mol. Imaging 2025, 52, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, D.J.; Kim, J.; Tran, T.; Scola, P.M.; Tenney, D.; Pena, A.; Petrone, T.; Zhang, Y.; Boy, K.M.; Poss, M.A.; et al. The discovery and evaluation of [18F]BMS-986229, a novel macrocyclic peptide PET radioligand for the measurement of PD-L1 expression and in-vivo PD-L1 target engagement. Eur. J. Nucl. Med. Mol. Imaging 2024, 51, 978–990. [Google Scholar] [CrossRef]
- Brauer, J.; Tumani, M.; Frey, N.; Lehmann, L.H. The cardio-oncologic burden of breast cancer: Molecular mechanisms and importance of preclinical models. Basic. Res. Cardiol. 2025, 120, 91–112. [Google Scholar] [CrossRef]
- Wang, L.; Piao, Y.; Guo, F.; Wei, J.; Chen, Y.; Dai, X.; Zhang, X. Current progress of pig models for liver cancer research. Biomed. Pharmacother. 2023, 165, 115256. [Google Scholar] [CrossRef]
- Choen, S.; Kent, M.S.; Loucks, F.A.; Winger, J.A.; Zwingenberger, A.L. Assessment of tumor hypoxia in spontaneous canine tumors after treatment with OMX, a novel H-NOX oxygen carrier, with [18F]FMISO PET/CT. BMC Vet. Res. 2024, 20, 196. [Google Scholar] [CrossRef]
- Kjaergaard, K.; Weber, B.; Alstrup, A.K.O.; Petersen, J.B.B.; Hansen, R.; Hamilton-Dutoit, S.J.; Mortensen, F.V.; Sorensen, M. Hepatic regeneration following radiation-induced liver injury is associated with increased hepatobiliary secretion measured by PET in Gottingen minipigs. Sci. Rep. 2020, 10, 10858. [Google Scholar] [CrossRef]
- Xiang, J.; Zhang, Z.; Wu, S.; Ye, K. Positron emission tomography tracers for synucleinopathies. Mol. Neurodegener. 2025, 20, 1. [Google Scholar] [CrossRef]
- Yuan, X.; Nie, S.; Yang, Y.; Liu, C.; Xia, D.; Meng, L.; Xia, Y.; Su, H.; Zhang, C.; Bu, L.; et al. Propagation of pathologic α-synuclein from kidney to brain may contribute to Parkinson’s disease. Nat. Neurosci. 2025, 28, 577–588. [Google Scholar] [CrossRef]
- Pavese, N.; Rivero-Bosch, M.; Lewis, S.J.; Whone, A.L.; Brooks, D.J. Progression of monoaminergic dysfunction in Parkinson’s disease: A longitudinal 18F-dopa PET study. Neuroimage 2011, 56, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Fayard, A.; Fenyi, A.; Lavisse, S.; Dovero, S.; Bousset, L.; Bellande, T.; Lecourtois, S.; Jouy, C.; Guillermier, M.; Jan, C.; et al. Functional and neuropathological changes induced by injection of distinct alpha-synuclein strains: A pilot study in non-human primates. Neurobiol. Dis. 2023, 180, 106086. [Google Scholar] [CrossRef] [PubMed]
- Sawamura, M.; Onoe, H.; Tsukada, H.; Isa, K.; Yamakado, H.; Okuda, S.; Ikuno, M.; Hatanaka, Y.; Murayama, S.; Uemura, N.; et al. Lewy Body Disease Primate Model with alpha-Synuclein Propagation from the Olfactory Bulb. Mov. Disord. 2022, 37, 2033–2044. [Google Scholar] [CrossRef]
- Molinet-Dronda, F.; Blesa, J.; Del Rey, N.L.; Juri, C.; Collantes, M.; Pineda-Pardo, J.A.; Trigo-Damas, I.; Iglesias, E.; Hernandez, L.F.; Rodriguez-Rojas, R.; et al. Cerebral metabolic pattern associated with progressive parkinsonism in non-human primates reveals early cortical hypometabolism. Neurobiol. Dis. 2022, 167, 105669. [Google Scholar] [CrossRef]
- López-Ornelas, A.; Escobedo-Avila, I.; Ramírez-García, G.; Lara-Rodarte, R.; Meléndez-Ramírez, C.; Urrieta-Chavez, B.; Barrios-García, T.; Cáceres-Chávez, V.A.; Flores-Ponce, X.; Carmona, F.; et al. Human Embryonic Stem Cell-Derived Immature Midbrain Dopaminergic Neurons Transplanted in Parkinsonian Monkeys. Cells 2023, 12, 2738. [Google Scholar] [CrossRef]
- Pokrzyk, J.; Kulczyńska-Przybik, A.; Guzik-Makaruk, E.; Winkel, I.; Mroczko, B. Clinical Importance of Amyloid Beta Implication in the Detection and Treatment of Alzheimer’s Disease. Int. J. Mol. Sci. 2025, 26, 1935. [Google Scholar] [CrossRef]
- Nabizadeh, F. Connectomics and neurotransmitter receptor profile explain regional tau pathology in Alzheimer’s disease. Cereb. Cortex 2025, 35, bhaf053. [Google Scholar] [CrossRef] [PubMed]
- Maetzler, W.; Reimold, M.; Liepelt, I.; Solbach, C.; Leyhe, T.; Schweitzer, K.; Eschweiler, G.W.; Mittelbronn, M.; Gaenslen, A.; Uebele, M.; et al. [11C]PIB binding in Parkinson’s disease dementia. Neuroimage 2008, 39, 1027–1033. [Google Scholar] [CrossRef]
- Cohen, A.D.; Klunk, W.E. Early detection of Alzheimer’s disease using PiB and FDG PET. Neurobiol. Dis. 2014, 72 Pt A, 117–122. [Google Scholar] [CrossRef]
- Bhoopal, B.; Gollapelli, K.K.; Damuka, N.; Miller, M.; Krizan, I.; Bansode, A.; Register, T.; Frye, B.M.; Kim, J.; Mintz, A.; et al. Preliminary PET Imaging of Microtubule-Based PET Radioligand [11C]MPC-6827 in a Nonhuman Primate Model of Alzheimer’s Disease. Acs Chem. Neurosci. 2023, 14, 3745–3751. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, K.; Zhang, X.; Zhao, H.; Wang, X.; Dong, R.; Wang, Y.; Chen, B.; Yan, X.X.; Dai, J.; et al. Fluorine-18-Labeled Diaryl-azines as Improved beta-Amyloid Imaging Tracers: From Bench to First-in-Human Studies. J. Med. Chem. 2023, 66, 4603–4616. [Google Scholar] [CrossRef] [PubMed]
- Sawant-Basak, A.; Chen, L.; Lockwood, P.; Boyden, T.; Doran, A.C.; Mancuso, J.; Zasadny, K.; McCarthy, T.; Morris, E.D.; Carson, R.E.; et al. Investigating CNS distribution of PF-05212377, a P-glycoprotein substrate, by translation of 5-HT(6) receptor occupancy from non-human primates to humans. Biopharm. Drug Dispos. 2023, 44, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, L.; Dutra, J.K.; Beck, E.M.; Nag, S.; Takano, A.; Amini, N.; Arakawa, R.; Brodney, M.A.; Buzon, L.M.; et al. Identification of a Novel Positron Emission Tomography (PET) Ligand for Imaging beta-Site Amyloid Precursor Protein Cleaving Enzyme 1 (BACE-1) in Brain. J. Med. Chem. 2018, 61, 3296–3308. [Google Scholar] [CrossRef] [PubMed]
- Cook, B.E.; Nag, S.; Arakawa, R.; Lin, E.Y.; Stratman, N.; Guckian, K.; Hering, H.; Lulla, M.; Choi, J.; Salinas, C.; et al. Development of a PET Tracer for OGA with Improved Kinetics in the Living Brain. J. Nucl. Med. 2023, 64, 1588–1593. [Google Scholar] [CrossRef]
- Stoker, T.B.; Mason, S.L.; Greenland, J.C.; Holden, S.T.; Santini, H.; Barker, R.A. Huntington’s disease: Diagnosis and management. Pract. Neurol. 2022, 22, 32–41. [Google Scholar] [CrossRef]
- Aqel, S.; Ahmad, J.; Saleh, I.; Fathima, A.; Al Thani, A.A.; Mohamed, W.M.Y.; Shaito, A.A. Advances in Huntington’s Disease Biomarkers: A 10-Year Bibliometric Analysis and a Comprehensive Review. Biology 2025, 14, 129. [Google Scholar] [CrossRef]
- Williams, G.K.; Akkermans, J.; Lawson, M.; Syta, P.; Staelens, S.; Adhikari, M.H.; Morton, A.J.; Nitzsche, B.; Boltze, J.; Christou, C.; et al. Imaging Glucose Metabolism and Dopaminergic Dysfunction in Sheep (Ovis aries) Brain Using Positron Emission Tomography Imaging Reveals Abnormalities in OVT73 Huntington’s Disease Sheep. Acs Chem. Neurosci. 2024, 15, 4082–4091. [Google Scholar] [CrossRef]
- Kaur, T.; Brooks, A.F.; Lapsys, A.; Desmond, T.J.; Stauff, J.; Arteaga, J.; Winton, W.P.; Scott, P.J.H. Synthesis and Evaluation of a Fluorine-18 Radioligand for Imaging Huntingtin Aggregates by Positron Emission Tomographic Imaging. Front. Neurosci. 2021, 15, 766176. [Google Scholar] [CrossRef]
- Liu, L.; Prime, M.E.; Lee, M.R.; Khetarpal, V.; Brown, C.J.; Johnson, P.D.; Miranda-Azpiazu, P.; Chen, X.; Clark-Frew, D.; Coe, S.; et al. Imaging Mutant Huntingtin Aggregates: Development of a Potential PET Ligand. J. Med. Chem. 2020, 63, 8608–8633. [Google Scholar] [CrossRef]
- Liu, L.; Johnson, P.D.; Prime, M.E.; Khetarpal, V.; Lee, M.R.; Brown, C.J.; Chen, X.; Clark-Frew, D.; Coe, S.; Conlon, M.; et al. [(11)C]CHDI-626, a PET Tracer Candidate for Imaging Mutant Huntingtin Aggregates with Reduced Binding to AD Pathological Proteins. J. Med. Chem. 2021, 64, 12003–12021. [Google Scholar] [CrossRef]
- Liu, L.B.; Johnson, P.D.; Prime, M.E.; Khetarpal, V.; Brown, C.J.; Anzillotti, L.; Bertoglio, D.; Chen, X.M.; Coe, S.; Davis, R.; et al. Design and Evaluation of [18F]CHDI-650 as a Positron Emission Tomography Ligand to Image Mutant Huntingtin Aggregates. J. Med. Chem. 2023, 66, 641–656. [Google Scholar] [CrossRef] [PubMed]
- Smucny, J.; Vlasova, R.M.; Lesh, T.A.; Rowland, D.J.; Wang, G.; Chaudhari, A.J.; Chen, S.; Iosif, A.M.; Hogrefe, C.E.; Bennett, J.L.; et al. Increased Striatal Presynaptic Dopamine in a Nonhuman Primate Model of Maternal Immune Activation: A Longitudinal Neurodevelopmental Positron Emission Tomography Study With Implications for Schizophrenia. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2023, 8, 505–513. [Google Scholar] [CrossRef]
- Wakeford, A.; Nye, J.A.; Grieb, Z.A.; Voisin, D.A.; Mun, J.; Huhman, K.L.; Albers, E.; Michopoulos, V. Sex influences the effects of social status on socioemotional behavior and serotonin neurochemistry in rhesus monkeys. Biol. Sex. Differ. 2023, 14, 75. [Google Scholar] [CrossRef]
- Birn, R.M.; Shackman, A.J.; Oler, J.A.; Williams, L.E.; McFarlin, D.R.; Rogers, G.M.; Shelton, S.E.; Alexander, A.L.; Pine, D.S.; Slattery, M.J.; et al. Evolutionarily conserved prefrontal-amygdalar dysfunction in early-life anxiety. Mol. Psychiatry 2014, 19, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.S.; Oler, J.A.; Shackman, A.J.; Shelton, S.E.; Raveendran, M.; McKay, D.R.; Converse, A.K.; Alexander, A.; Davidson, R.J.; Blangero, J.; et al. Intergenerational neural mediators of early-life anxious temperament. Proc. Natl. Acad. Sci. USA 2015, 112, 9118–9122. [Google Scholar] [CrossRef] [PubMed]
- Shackman, A.J.; Fox, A.S.; Oler, J.A.; Shelton, S.E.; Oakes, T.R.; Davidson, R.J.; Kalin, N.H. Heightened extended amygdala metabolism following threat characterizes the early phenotypic risk to develop anxiety-related psychopathology. Mol. Psychiatry 2017, 22, 724–732. [Google Scholar] [CrossRef]
- Belin, D.; Dalley, J.W. Animal Models in Addiction Research. In Drug Abuse and Addiction in Medical Illness: Causes, Consequences and Treatment; Verster, J.C., Brady, K., Galanter, M., Conrod, P., Eds.; Springer: New York, NY, USA, 2012; pp. 73–93. [Google Scholar] [CrossRef]
- Trifilieff, P.; Ducrocq, F.; van der Veldt, S.; Martinez, D. Blunted Dopamine Transmission in Addiction: Potential Mechanisms and Implications for Behavior. Semin. Nucl. Med. 2017, 47, 64–74. [Google Scholar] [CrossRef]
- Allen, M.I.; Duke, A.N.; Nader, S.H.; Adler-Neal, A.; Solingapuram Sai, K.K.; Reboussin, B.A.; Gage, H.D.; Voll, R.J.; Mintz, A.; Goodman, M.M.; et al. PET imaging of dopamine transporters and D2/D3 receptors in female monkeys: Effects of chronic cocaine self-administration. Neuropsychopharmacology 2023, 48, 1436–1445. [Google Scholar] [CrossRef]
- Nader, M.A.; Morgan, D.; Gage, H.D.; Nader, S.H.; Calhoun, T.L.; Buchheimer, N.; Ehrenkaufer, R.; Mach, R.H. PET imaging of dopamine D2 receptors during chronic cocaine self-administration in monkeys. Nat. Neurosci. 2006, 9, 1050–1056. [Google Scholar] [CrossRef]
- Gould, R.W.; Porrino, L.J.; Nader, M.A. Nonhuman primate models of addiction and PET imaging: Dopamine system dysregulation. Curr. Top. Behav. Neurosci. 2012, 11, 25–44. [Google Scholar] [CrossRef]
- Gallezot, J.D.; Kloczynski, T.; Weinzimmer, D.; Labaree, D.; Zheng, M.Q.; Lim, K.; Rabiner, E.A.; Ridler, K.; Pittman, B.; Huang, Y.; et al. Imaging nicotine- and amphetamine-induced dopamine release in rhesus monkeys with [(11)C]PHNO vs [(11)C]raclopride PET. Neuropsychopharmacology 2014, 39, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Naylor, J.E.; Hiranita, T.; Matazel, K.S.; Zhang, X.; Paule, M.G.; Goodwin, A.K. Positron emission tomography (PET) imaging of nicotine-induced dopamine release in squirrel monkeys using [18F]Fallypride. Drug Alcohol. Depend. 2017, 179, 254–259. [Google Scholar] [CrossRef] [PubMed]
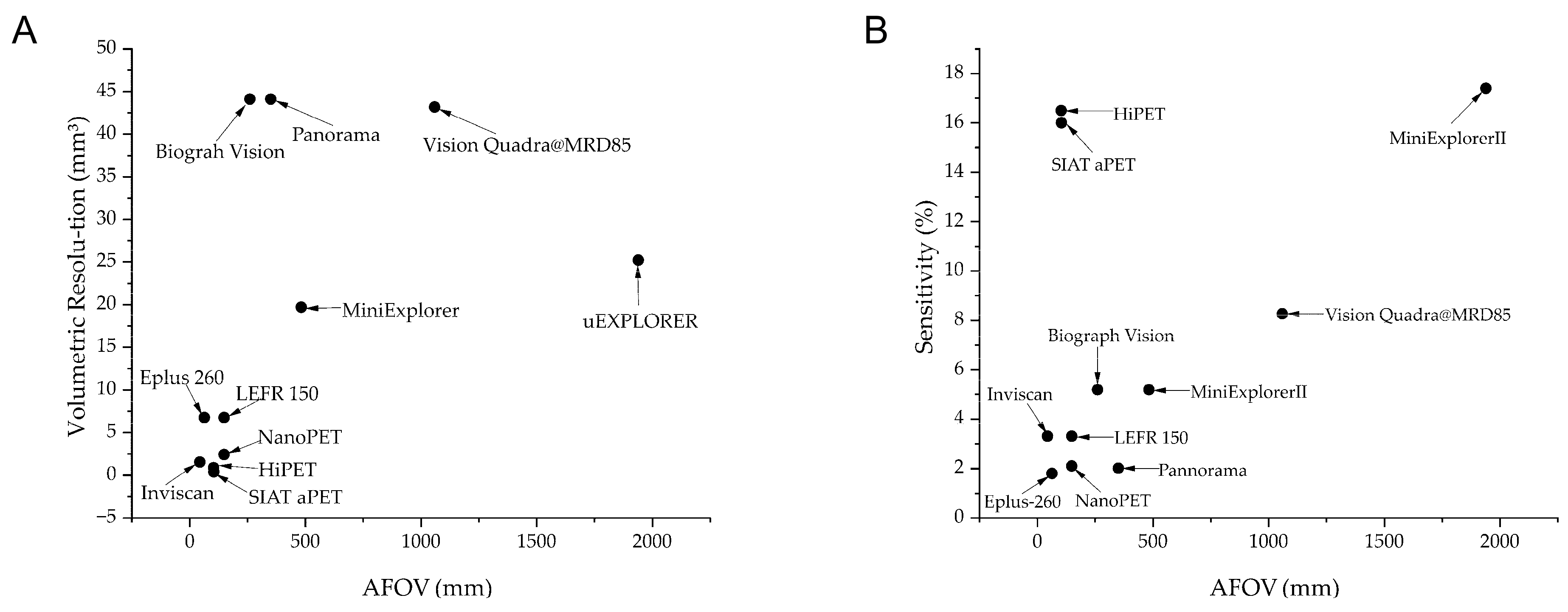
| Name of Scanner | Crystal Size (mm3) | Transaxial/Axial Field of View (mm) | Volumetric Resolution (mm3)/(Radial × Tangential × Axial) | Reconstruction Method | Sensitivity (%) | Energy Window (keV) | Coincidence Window (ns) | Time-of-Flight Resolution (ps) | Depth of Interaction Information | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Animal PET Scanner | Small-animal PET | NanoPET [12] | 1.12 × 1.12 × 13 | 123/9.4~150 | 2.40 (1.49 × 1.39 × 1.16) | Filtered Back Projection (FBP) | 2.1 | 250–750 | 5 | -- | No |
| HiPET [13] | 1.01 × 1.01 × 6.1 & 1.55 × 1.55 × 8.9 | 131/104 | 0.882 (1.0 × 0.98 × 0.9) | FBP | 16.5 | 150–750 | 6 | Yes | |||
| SIAT aPET [14] | 1.0 × 1.0 × 20 | <111/105.6 | 0.37 (0.68 × 0.8 × 0.69) | Ordered Subset Expectation Maximization | 16 | 250–750 | -- | -- | Yes | ||
| Large-animal PET | Inviscan [19] | 1.6 × 1.6 × 16 | 170/45 | 1.53 (1.05 × 0.97 × 1.51) | FBP | 3.3 | 250–750 | 5.2 | -- | Yes | |
| LFER 150 [15] | 1.51 × 1.51 × 10 | 200/150 | 6.73 (2 × 1.86 × 1.81) | FBP | 3.3 | 400–600 | 5 | -- | No | ||
| Eplus-260 [16] | 1.9 × 1.9 × 10 | 190/64 | 6.72 (2.4 × 2 × 1.4) | FBP | 1.8 | 460–660 | 2 | -- | No | ||
| MiniExplorer II [18] | 2.76 × 2.76 × 18 | 520/483 | 19.69 (2.62 × 2.88 × 2.61) @1/2 AFOV | FBP | 5.18 | -- | -- | 409 | No | ||
| Clinical PET Scanner | Short-axial FOV | Panorama [8] | 2.76 × 2.76 × 18.1 | 760/351 | 44.1 (2.72 × 3.02 × 2.74) | FBP | 2.01 | -- | -- | 189 | No |
| Biograph Vison [9] | 3.2 × 3.2 × 20 | 780/261 | 44.1 (3.5 × 3.6 × 3.5) | FBP | 5.18 | -- | -- | 203 | No | ||
| Long-axial FOV | Vision Quadra@MRD85 [10] | 3.2 × 3.2 × 20 | 780/1060 | 43.16 (3.19 × 3.58 × 3.78) | FBP | 8.26 | -- | -- | 228 | No | |
| uExplorer [11] | 2.76 × 2.76 × 18.1 | 760/1940 | 25.2 (3.0 × 3.0 × 2.8) | FBP | 17.4 | -- | -- | 500 | No |
| Disease | Genetic Modification | Phenotype | Potential Applications |
|---|---|---|---|
| Diabetes [37,38] | Knockin dominant-negative glucose-dependent insulinotropic polypeptide (GIP) receptor | Reduced insulinotropic effect of GIP; reduced glucose tolerance and insulin secretion | Development and preclinical evaluation of incretin-based therapeutic strategies |
| Knockin INS-C94Y | Reduced β-cell mass and insulin level; dilation of the endoplasmic reticulum in β cell; cataract development | Preclinical testing of novel treatment strategies | |
| Congenital heart disease [39] | Combine patient-specific human-induced pluripotent stem cell-derived cardiomyocytes with pig hearts’ extracellular-matrix hydrogel | Abnormal phenotype of the long QT syndrome and catecholaminergic polymorphic ventricular tachycardia; occurrence of reentrant arrhythmia | Study of inherited and acquired cardiac disorders and drug development and testing |
| Ischemia/reperfusion-induced myocardial infarction [40] | Knockout the Hippo pathway gene Salvador | Improved ejection fraction; reduced scar sizes; increased capillary density and reduced cardiomyocyte ploidy | Treating heart failure |
| Heart failure (HF) [41] | Knockout the small ubiquitin-related modifier 1 gene | Improved cardiac function and stabilized left ventricular volumes | An approach to human HF therapy |
| Disease | Genetic Modification | Phenotype | Potential Applications |
|---|---|---|---|
| Huntington’s disease (HD) [55] | Knockin polyglutamine-expanded human huntingtin (HTT) gene | Observed nuclear inclusions and neuropil aggregates; dystonia and chorea | Underlying biology of HD and the development of potential therapies |
| Parkinson’s disease (PD) [56] | Knockout PINK1 gene | Reduced neuronal cells in cortex and striatum; decreased gray matter density in the cortex | A tool to investigate the diverse functions of PINK1 and the pathogenesis related to PINK1 dysfunction |
| Duchenne muscular dystrophy (DMD) [57] | Snip out a section of the dystrophin gene | Loss of dystrophin | A tool to understand the pathogenesis of DMD |
| Rett Syndrome [58] | Knockin MECP2 gene | Observed increase in frequency of repetitive circular locomotion; increase in anxiety; reduced social interaction; relatively weak cognitive phenotypes | -- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Z.; Xi, P.; Zheng, D.; Xie, Z.; Meng, X.; Ren, Q. The Impact of PET Imaging on Translational Medicine: Insights from Large-Animal Disease Models. Biomolecules 2025, 15, 919. https://doi.org/10.3390/biom15070919
Deng Z, Xi P, Zheng D, Xie Z, Meng X, Ren Q. The Impact of PET Imaging on Translational Medicine: Insights from Large-Animal Disease Models. Biomolecules. 2025; 15(7):919. https://doi.org/10.3390/biom15070919
Chicago/Turabian StyleDeng, Zhengyan, Peng Xi, Dongye Zheng, Zhaoheng Xie, Xiangxi Meng, and Qiushi Ren. 2025. "The Impact of PET Imaging on Translational Medicine: Insights from Large-Animal Disease Models" Biomolecules 15, no. 7: 919. https://doi.org/10.3390/biom15070919
APA StyleDeng, Z., Xi, P., Zheng, D., Xie, Z., Meng, X., & Ren, Q. (2025). The Impact of PET Imaging on Translational Medicine: Insights from Large-Animal Disease Models. Biomolecules, 15(7), 919. https://doi.org/10.3390/biom15070919