
The Estimated Intake of S100B Relates to Microbiota Biodiversity in Different Diets

 , , ,
, , ,  , and
, and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Inclusion/Exclusion Criteria
2.3. Data Extraction and Quality Assessment
2.4. Estimation of S100B in Different Dietary Patterns
2.5. Data Preparation for Shannon Index Measurement
2.6. The Relative Risk (RR) of Crohn’s Disease
- RRi is the relative risk of Crohn’s disease for country iii;
- Pi is the prevalence of the disease in country iii;
- Pmin is the lowest observed prevalence among all analyzed countries.
- RRnorm,i is the normalized relative risk for country iii;
- RRmin and RRmax are the minimum and maximum observed RR values, respectively.
3. Results
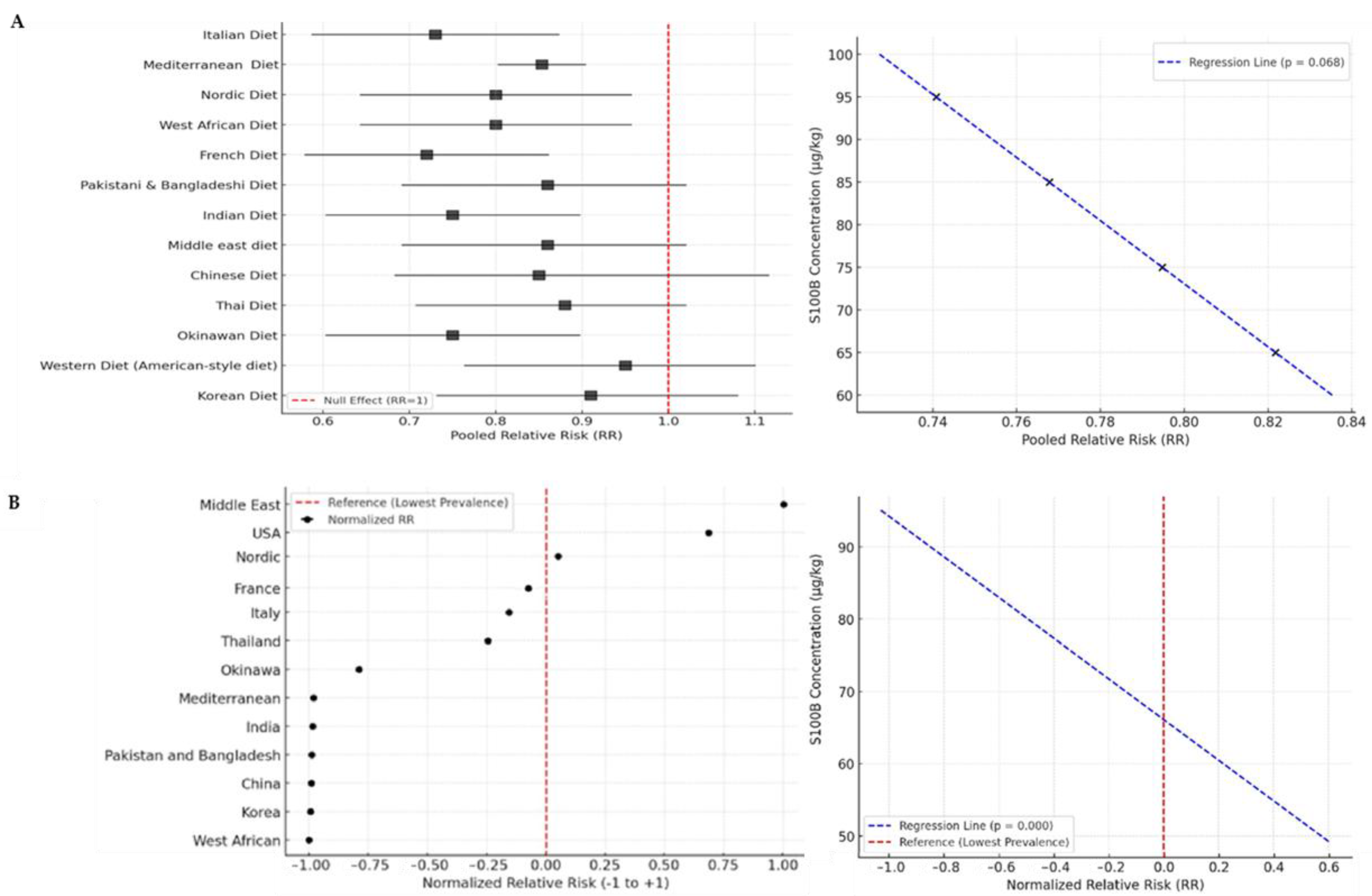
3.1. Estimation of S100B in Different Dietary Patterns
3.2. Epidemiological Investigation into Dietary Patterns with Different Levels of S100B
4. Discussion
5. Mechanistic Considerations and Future Research Directions
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CNS | Central nervous system |
| HLH | Helix–loop–helix |
| ENS | Enteric nervous system |
| RCT | Randomized control trial |
| CI | Confidence interval |
| HR | Hazard ratios |
| RR | Relative risk |
| NOS | Newcastle–Ottawa scale |
| T2DM | Type 2 diabetes mellitus |
| BMI | Body mass index |
| USA | United States of America |
| CVD | Cardiovascular diseases |
References
- Michetti, F.; Romano Spica, V. The “Jekyll Side” of the S100B Protein: Its Trophic Action in the Diet. Nutrients 2025, 17, 881. [Google Scholar] [CrossRef] [PubMed]
- Ferri, G.L.; Probert, L.; Cocchia, D.; Michetti, F.; Marangos, P.J.; Polak, J.M. Evidence for the presence of S-100 protein in the glial component of the human enteric nervous system. Nature 1982, 297, 409–410. [Google Scholar] [CrossRef]
- Santamaria-Kisiel, L.; Rintala-Dempsey, A.C.; Shaw, G.S. Calcium-dependent and-independent interactions of the S100 protein family. Biochem. J. 2006, 396, 201–214. [Google Scholar] [CrossRef]
- Michetti, F.; Clementi, M.E.; Di Liddo, R.; Valeriani, F.; Ria, F.; Rende, M.; Di Sante, G.; Romano Spica, V. The S100B Protein: A Multifaceted Pathogenic Factor More Than a Biomarker. Int. J. Mol. Sci. 2023, 24, 9605. [Google Scholar] [CrossRef]
- Romano Spica, V.; Volpini, V.; Valeriani, F.; Carotenuto, G.; Arcieri, M.; Platania, S.; Castrignanò, T.; Clementi, M.E.; Michetti, F. In Silico Predicting the Presence of the S100B Motif in Edible Plants and Detecting Its Immunoreactive Materials: Perspectives for Functional Foods, Dietary Supplements and Phytotherapies. Int. J. Mol. Sci. 2024, 25, 9813. [Google Scholar] [CrossRef]
- Romano Spica, V.; Valeriani, F.; Orsini, M.; Clementi, M.E.; Seguella, L.; Gianfranceschi, G.; Di Liddo, R.; Di Sante, G.; Ubaldi, F.; Ria, F.; et al. S100B affects gut microbiota biodiversity. Int. J. Mol. Sci. 2023, 24, 2248. [Google Scholar] [CrossRef]
- Bienenstock, J.; Kunze, W.; Forsythe, P. Microbiota and the gut–brain axis. Nutr. Rev. 2015, 73 (Suppl. S1), 28–31. [Google Scholar] [CrossRef]
- Freedman, L.S.; Potischman, N.; Kipnis, V.; Midthune, D.; Schatzkin, A.; Thompson, F.E.; Troiano, R.P.; Prentice, R.; Patterson, R.; Carroll, R.; et al. A comparison of two dietary instruments for evaluating the fat–breast cancer relationship. Int. J. Epidemiol. 2010, 39, 370–378. [Google Scholar] [CrossRef]
- Godala, M.; Gaszyńska, E.; Zatorski, H.; Małecka-Wojciesko, E. Dietary Interventions in Inflammatory Bowel Disease. Nutrients 2022, 14, 4261. [Google Scholar] [CrossRef]
- Barros, A.J.; Hirakata, V.N. Alternatives for logistic regression in cross-sectional studies: An empirical comparison of models that directly estimate the prevalence ratio. BMC Med. Res. Methodol. 2003, 3, 21. [Google Scholar] [CrossRef]
- Li, Y.; Pan, A.; Wang, D.D.; Liu, X.; Dhana, K.; Franco, O.H.; Kaptoge, S.; Di Angelantonio, E.; Stampfer, M.J.; Willett, W.C.; et al. Impact of Healthy Lifestyle Factors on Life Expectancies in the US Population. JAMA Intern. Med. 2018, 178, 601–608. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Hoffmann, G. Does a Mediterranean-Type Diet Reduce Cancer Risk? Curr. Nutr. Rep. 2016, 5, 9–17. [Google Scholar] [CrossRef]
- Reedy, J.; Krebs-Smith, S.M.; Miller, P.E.; Liese, A.D.; Subar, A.F.; Kahle, L.L.; Boushey, C.J. Higher diet quality is associated with decreased risk of all-cause, cardiovascular disease, and cancer mortality among older adults. J. Nutr. 2014, 144, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Heidemann, C.; Schulze, M.B.; Franco, O.H.; van Dam, R.M.; Mantzoros, C.S.; Hu, F.B. Dietary patterns and risk of mortality from cardiovascular disease, cancer, and all causes in a prospective cohort of women. Circulation 2008, 118, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Shu, W.; Liu, L.; Wang, X. Dietary patterns and hepatocellular carcinoma risk: A systematic review and meta-analysis of cohort and case–control studies. Nutr. Metab. 2024, 21, 47. [Google Scholar] [CrossRef] [PubMed]
- Tognon, G.; Nilsson, L.M.; Lissner, L.; Johansson, I.; Hallmans, G.; Lindahl, B.; Winkvist, A. The Mediterranean diet score and mortality are inversely associated in adults living in the subarctic region. J. Nutr. 2012, 142, 1547–1553. [Google Scholar] [CrossRef]
- Liese, A.D.; Krebs-Smith, S.M.; Subar, A.F.; George, S.M.; Harmon, B.E.; Neuhouser, M.L.; Reedy, J. The Dietary Patterns Methods Project: Synthesis of findings across cohorts and relevance to dietary guidance. J. Nutr. 2015, 145, 393–402. [Google Scholar] [CrossRef]
- Myint, P.K.; Luben, R.N.; Wareham, N.J.; Khaw, K.T. Combined work and leisure physical activity and risk of stroke in men and women in the European Prospective Investigation into Cancer-Norfolk prospective population study. Neuroepidemiology 2009, 32, 219–226. [Google Scholar] [CrossRef]
- Shin, D.; Lee, K.W.; Song, W.O. Dietary patterns during pregnancy are associated with risk of gestational diabetes mellitus. Nutrients 2023, 15, 857. [Google Scholar] [CrossRef]
- Castello, A.; Pollán, M.; Buijsse, B.; Ruiz, A.; Casas, A.M.; Baena-Cañada, J.M.; Amiano, P. Spanish Mediterranean diet and other dietary patterns and breast cancer risk: Case–control EpiGEICAM study. Br. J. Cancer 2014, 111, 1454–1462. [Google Scholar] [CrossRef]
- Esposito, K.; Kastorini, C.M.; Panagiotakos, D.B.; Giugliano, D. Prevention of type 2 diabetes by dietary patterns: A systematic review of prospective studies and meta-analysis. Metab. Syndr. Relat. Disord. 2010, 8, 471–476. [Google Scholar] [CrossRef]
- Kurotani, K.; Nanri, A.; Goto, A.; Mizoue, T.; Noda, M.; Kato, M.; Japan Public Health Center-based Prospective Study Group. Red meat consumption is associated with the risk of type 2 diabetes in men but not in women: A Japan Public Health Center-based Prospective Study. Br. J. Nutr. 2013, 110, 1910–1918. [Google Scholar] [CrossRef]
- Kaluza, J.; Åkesson, A.; Wolk, A. Diet quality and risk of hypertension: A prospective cohort study of men. J. Hum. Hypertens. 2014, 28, 590–595. [Google Scholar]
- Chan, R.; Wong, V.W.S.; Chu, W.C.W.; Wong, G.L.H.; Li, L.S.; Leung, J.; Woo, J. Diet-quality scores and prevalence of nonalcoholic fatty liver disease: A population study using proton-magnetic resonance spectroscopy. PLoS ONE 2015, 10, e0139310. [Google Scholar] [CrossRef]
- Kesse-Guyot, E.; Andreeva, V.A.; Lassale, C.; Hercberg, S.; Galan, P. Clustering of midlife lifestyle behaviors is associated with subsequent cognitive function: A longitudinal study. Am. J. Prev. Med. 2013, 45, 376–384. [Google Scholar]
- Heidemann, C.; Boeing, H.; Pischon, T.; Nöthlings, U.; Joost, H.G.; Schulze, M.B. Association of a Diabetes Risk Score with Risk of Myocardial Infarction, Stroke, Specific Types of Cancer, and Mortality: A Prospective Study in the European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Cohort. Eur. J. Epidemiol. 2009, 24, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Kurotani, K.; Nanri, A.; Goto, A.; Mizoue, T.; Noda, M.; Kato, M.; Inoue, M.; Tsugane, S.; Japan Public Health Center-Based Prospective Study Group. Vegetable and Fruit Intake and Risk of Type 2 Diabetes: Japan Public Health Center-Based Prospective Study. Br. J. Nutr. 2013, 109, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B.; Rimm, E.B.; Stampfer, M.J.; Ascherio, A.; Spiegelman, D.; Willett, W.C. Prospective study of major dietary patterns and risk of coronary heart disease in men. Am. J. Clin. Nutr. 2000, 72, 912–921. [Google Scholar] [CrossRef] [PubMed]
- Trichopoulou, A.; Costacou, T.; Bamia, C.; Trichopoulos, D. Adherence to a Mediterranean diet and survival in a Greek population. N. Engl. J. Med. 2003, 348, 2599–2608. [Google Scholar] [CrossRef]
- Fung, T.T.; Willett, W.C.; Stampfer, M.J.; Manson, J.E.; Hu, F.B. Dietary patterns and the risk of coronary heart disease in women. Arch. Intern. Med. 2001, 161, 1857–1862. [Google Scholar] [CrossRef]
- Knoops, K.T.B.; de Groot, L.C.P.G.M.; Kromhout, D.; Perrin, A.E.; Moreiras-Varela, O.; Menotti, A.; van Staveren, W.A. Mediterranean diet, lifestyle factors, and 10-year mortality in elderly European men and women: The HALE project. JAMA 2004, 292, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Esposito, K.; Marfella, R.; Ciotola, M.; Di Palo, C.; Giugliano, F.; Giugliano, G.; Giugliano, D. Effect of a Mediterranean-style diet on endothelial dysfunction and markers of vascular inflammation in the metabolic syndrome: A randomized trial. JAMA 2004, 292, 1440–1446. [Google Scholar] [CrossRef] [PubMed]
- Tucker, K.L.; Hallfrisch, J.; Qiao, N.; Muller, D.; Andres, R.; Fleg, J.L. The combination of high fruit and vegetable and low saturated fat intakes is more protective against mortality in aging men than is either alone: The Baltimore Longitudinal Study of Aging. J. Nutr. 2005, 135, 556–561. [Google Scholar] [CrossRef]
- Nöthlings, U.; Schulze, M.B.; Weikert, C.; Boeing, H.; van der Schouw, Y.T.; Bamia, C.; Trichopoulou, A. Intake of vegetables, legumes, and fruit, and risk for all-cause, cardiovascular, and cancer mortality in a European diabetic population. J. Nutr. 2008, 138, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Mente, A.; de Koning, L.; Shannon, H.S.; Anand, S.S. A systematic review of the evidence supporting a causal link between dietary factors and coronary heart disease. Arch. Intern. Med. 2009, 169, 659–669. [Google Scholar] [CrossRef]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N. Engl. J. Med. 2018, 378, e34. [Google Scholar] [CrossRef]
- Tognon, G.; Lissner, L.; Sääksjärvi, K.; Karlsson, A.K. The Mediterranean diet in relation to mortality and CVD: A Danish cohort study. Br. J. Nutr. 2014, 111, 151–159. [Google Scholar] [CrossRef]
- Kokkinos, P.; Panagiotakos, D.B.; Polychronopoulos, E.; Pitsavos, C.; Lentzas, Y.; Stefanadis, C. Dietary habits in relation to blood pressure in elderly men and women: The MEDIS study. Int. J. Food Sci. Nutr. 2005, 56, 85–92. [Google Scholar]
- Panagiotakos, D.B.; Pitsavos, C.; Chrysohoou, C.; Stefanadis, C. The epidemiology of Type 2 diabetes mellitus in Greek adults: The ATTICA study. Diabet. Med. 2008, 25, 1181–1188. [Google Scholar] [CrossRef]
- Panagiotakos, D.B.; Pitsavos, C.; Chrysohoou, C.; Stefanadis, C. The role of traditional Mediterranean-type of diet and physical activity in the development of acute coronary syndromes in adult Greek men: The CARDIO2000 case–control study. J. Cardiovasc. Risk 2009, 9, 119–125. [Google Scholar]
- Kastorini, C.M.; Milionis, H.J.; Goudevenos, J.A.; Panagiotakos, D.B. Mediterranean diet and coronary heart disease: Is obesity a link?—A systematic review. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Panagiotakos, D.B.; Pitsavos, C.; Chrysohoou, C.; Stefanadis, C. The adoption of Mediterranean diet attenuates the development of acute coronary syndromes in people with the metabolic syndrome. Nutrition 2007, 23, 677–684. [Google Scholar]
- Panagiotakos, D.B.; Pitsavos, C.; Chrysohoou, C.; Stefanadis, C. Dietary patterns and 5-year incidence of cardiovascular disease: A multivariate analysis of the ATTICA study. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Kaplan, G.G. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef] [PubMed]
- Permpoon, V.; Pongpirul, K.; Anuras, S. Five-year clinical outcomes of Crohn’s disease: A report of 287 multiethnic cases from an International Hospital in Thailand. Clin. Exp. Gastroenterol. 2019, 12, 203–208. [Google Scholar] [CrossRef]
- Kappelman, M.D.; Moore, K.R.; Allen, J.K.; Cook, S.F. Recent trends in the prevalence of Crohn’s disease and ulcerative colitis in a commercially insured US population. Dig. Dis. Sci. 2013, 58, 519–525. [Google Scholar] [CrossRef]
- Crocetti, E.; Bergamaschi, W.; Russo, A.G. Population-based incidence and prevalence of inflammatory bowel diseases in Milan (Northern Italy), and estimates for Italy. Eur. J. Gastroenterol. Hepatol. 2021, 33, e383–e389. [Google Scholar] [CrossRef]
- Genin, M.; Fumery, M.; Occelli, F.; Savoye, G.; Pariente, B.; Dauchet, L.; Ficheur, G. Fine-scale geographical distribution and ecological risk factors for Crohn’s disease in France (2007–2014). Aliment. Pharmacol. Ther. 2020, 51, 139–148. [Google Scholar] [CrossRef]
- Jacobsen, B.A.; Fallingborg, J.; Rasmussen, H.H.; Nielsen, K.R.; Drewes, A.M.; Puho, E.; Sørensen, H.T. Increase in incidence and prevalence of inflammatory bowel disease in northern Denmark: A population-based study, 1978–2002. Eur. J. Gastroenterol. Hepatol. 2006, 18, 601–606. [Google Scholar] [CrossRef]
- Park, J.; Cheon, J.H. Incidence and prevalence of inflammatory bowel disease across Asia. Yonsei Med. J. 2021, 62, 99–108. [Google Scholar] [CrossRef]
- Zheng, J.J.; Zhu, X.S.; Huangfu, Z.; Shi, X.H.; Guo, Z.R. Prevalence and incidence rates of Crohn’s disease in mainland China: A meta-analysis of 55 years of research. J. Dig. Dis. 2010, 11, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Lee, J.M. Crohn’s disease in Korea: Past, present, and future. Korean J. Intern. Med. 2014, 29, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Makharia, G.K.; Ramakrishna, B.S.; Abraham, P.; Choudhuri, G.; Misra, S.P.; Ahuja, V.; Indian Society of Gastroenterology Task Force on Inflammatory Bowel Disease. Survey of inflammatory bowel diseases in India. Indian J. Gastroenterol. 2012, 31, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Tessier, A.J.; Wang, F.; Korat, A.A.; Eliassen, A.H.; Chavarro, J.; Grodstein, F.; Li, J.; Liang, L.; Willett, W.C.; Sun, Q.; et al. Optimal Dietary Patterns for Healthy Aging. Nat. Med. 2025, 31, 1644–1652. [Google Scholar] [CrossRef]
- Fetissov, S.O. Role of the gut microbiota in host appetite control: Bacterial growth to animal feeding behaviour. Nat. Rev. Endocrinol. 2017, 13, 11–25. [Google Scholar] [CrossRef]
- Kaplan, G.G.; Windsor, J.W. The four epidemiological stages in the global evolution of inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 56–66. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef]
- Sandhu, K.V.; Sherwin, E.; Schellekens, H.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Feeding the Microbiota–Gut–Brain Axis: Diet, Microbiome, and Neuropsychiatry. Transl. Res. 2017, 179, 223–244. [Google Scholar] [CrossRef]
- Seguella, L.; Chiarito, M.; Scarpellini, E.; Giuliano, C.; Cicala, M.; Pallotta, L.; Esposito, G. Mini-Review: Interaction between Intestinal Microbes and Enteric Glia in Health and Disease. Neurosci. Lett. 2023, 806, 137221. [Google Scholar] [CrossRef]
- Buckman, L.B.; Thompson, M.M.; Lippert, R.N.; Blackwell, T.S.; Yull, F.E.; Ellacott, K.L.J. Regulation of S100B in Adipose Tissue Inflammation and Insulin Resistance. Endocrinology 2015, 156, 2503–2513. [Google Scholar] [CrossRef]
- Camberos-Barraza, J.; Guadrón-Llanos, A.M.; De la Herrán-Arita, A.K. The Gut Microbiome–Neuroglia Axis: Implications for Brain Health, Inflammation, and Disease. Neuroglia 2024, 5, 254–273. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. Gut Instincts: Microbiota as a Key Regulator of Brain Development, Ageing and Neurodegeneration. J. Physiol. 2017, 595, 489–503. [Google Scholar] [CrossRef]
- Donato, R.; Sorci, G.; Riuzzi, F.; Arcuri, C.; Bianchi, R.; Brozzi, F.; Tubaro, C.; Giambanco, I. S100B’s Double Life: Intracellular Regulator and Extracellular Signal. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 1008–1022. [Google Scholar] [CrossRef]
- Rojas, A.; Lindner, C.; Schneider, I.; Gonzalez, I.; Uribarri, J. The RAGE Axis: A Relevant Inflammatory Hub in Human Diseases. Biomolecules 2024, 14, 412. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Liu, H.; Brown, M.A.; Qiao, S. Dietary Protein and Gut Microbiota Composition and Function. Curr. Protein Pept. Sci. 2019, 20, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Galvano, F.; Frigiola, A.; Gagliardi, L.; Ciotti, S.; Bognanno, M.; Iacopino, A.M.; Nigro, F.; Tina, G.L.; Cavallaro, D.; Mussap, M.; et al. S100B milk concentration in mammalian species. Front. Biosci. (Elite Ed.) 2009, 1, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Albenzio, M.; Santillo, A.; Ciliberti, M.G.; Figliola, L.; Caroprese, M.; Marino, R.; Polito, A.N. Milk from Different Species: Relationship between Protein Fractions and Inflammatory Response in Infants Affected by Generalized Epilepsy. J. Dairy Sci. 2016, 99, 5032–5038. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, T.; Fox, P.F.; de Kruif, K.G.; Kelly, A.L. High Pressure-Induced Changes in Bovine Milk Proteins: A Review. Biochim. Biophys. Acta 2006, 1764, 593–598. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Diet | S100B Concentration (µg/kg) | Shannon Index |
|---|---|---|
| Italian | 95.08 ± 0.23 | 3.1 |
| Mediterranean | 80.03 ± 0.39 | 3.0 |
| Nordic | 76.53 ± 0.32 | 2.4 |
| West African | 75.54 ± 0.32 | 2.9 |
| French | 65.03 ± 0.18 | 2.9 |
| Pakistani and Bangladeshi | 63.03 ± 0.37 | 2.2 |
| Indian | 63.03 ± 0.37 | 2.2 |
| Middle east | 60.08 ± 0.22 | 1.5 |
| Chinese | 54.03 ± 0.16 | 2.1 |
| Thai | 53.03 ± 0.16 | 1.9 |
| Okinawan | 51.53 ± 0.46 | 2.7 |
| Western (American style) | 49.54 ± 0.43 | 1.0 |
| Korean | 49.23 ± 0.02 | 1.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghaffar, T.; Volpini, V.; Platania, S.; Vassioukovitch, O.; Valle, A.; Valeriani, F.; Michetti, F.; Romano Spica, V. The Estimated Intake of S100B Relates to Microbiota Biodiversity in Different Diets. Biomolecules 2025, 15, 1047. https://doi.org/10.3390/biom15071047
Ghaffar T, Volpini V, Platania S, Vassioukovitch O, Valle A, Valeriani F, Michetti F, Romano Spica V. The Estimated Intake of S100B Relates to Microbiota Biodiversity in Different Diets. Biomolecules. 2025; 15(7):1047. https://doi.org/10.3390/biom15071047
Chicago/Turabian StyleGhaffar, Tehreema, Veronica Volpini, Serena Platania, Olga Vassioukovitch, Alessandra Valle, Federica Valeriani, Fabrizio Michetti, and Vincenzo Romano Spica. 2025. "The Estimated Intake of S100B Relates to Microbiota Biodiversity in Different Diets" Biomolecules 15, no. 7: 1047. https://doi.org/10.3390/biom15071047
APA StyleGhaffar, T., Volpini, V., Platania, S., Vassioukovitch, O., Valle, A., Valeriani, F., Michetti, F., & Romano Spica, V. (2025). The Estimated Intake of S100B Relates to Microbiota Biodiversity in Different Diets. Biomolecules, 15(7), 1047. https://doi.org/10.3390/biom15071047