
Electrophysiological Characterization of Sex-Dependent Hypnosis by an Endogenous Neuroactive Steroid Epipregnanolone


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
- NAS preparation
- Loss of the righting reflex (LORR)
- Electrophysiological Recordings and Data Acquisition
- Data Analysis
3. Results
3.1. Sex Differences in LORR

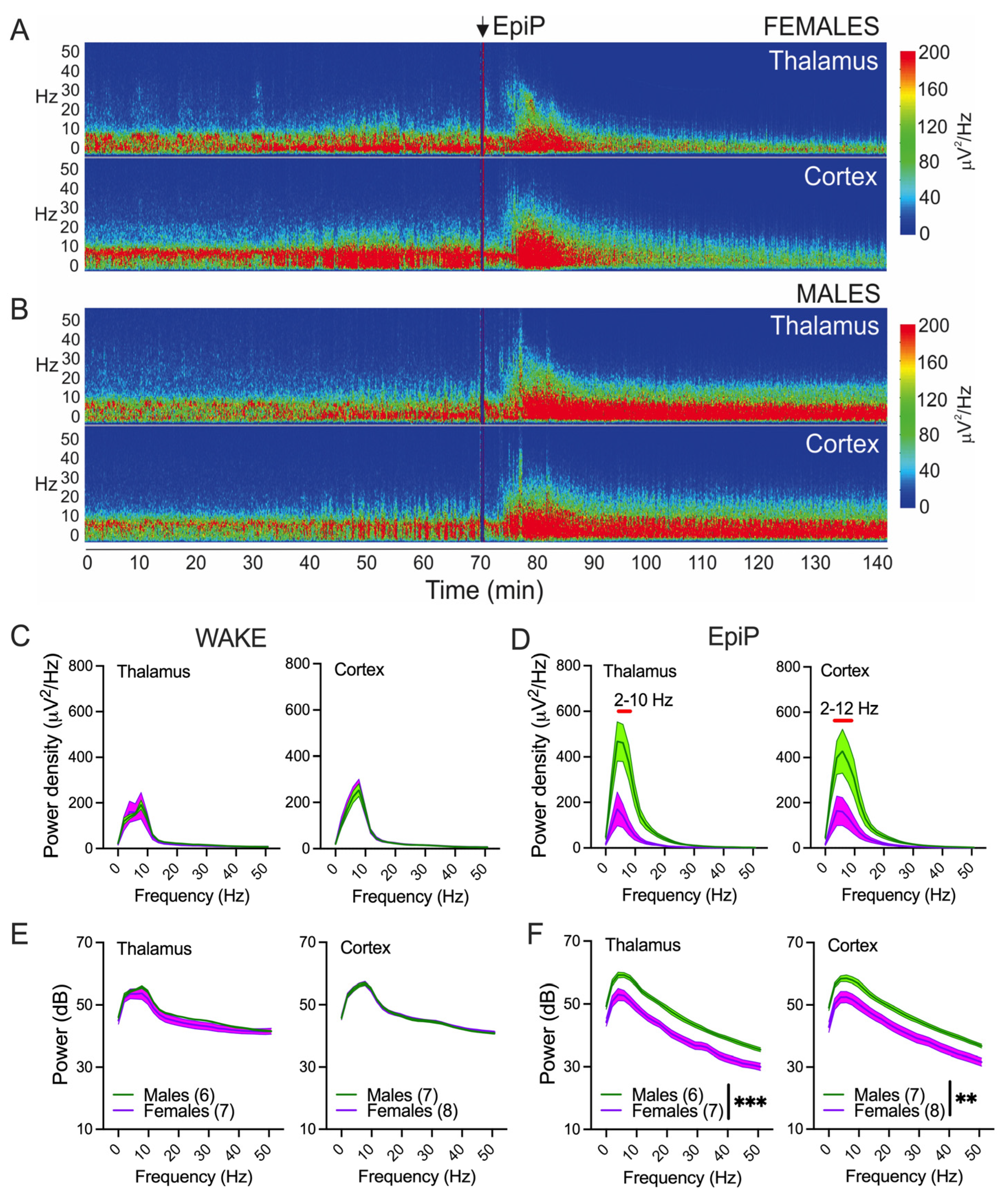
3.2. Sex Differences in Spectral Characteristics
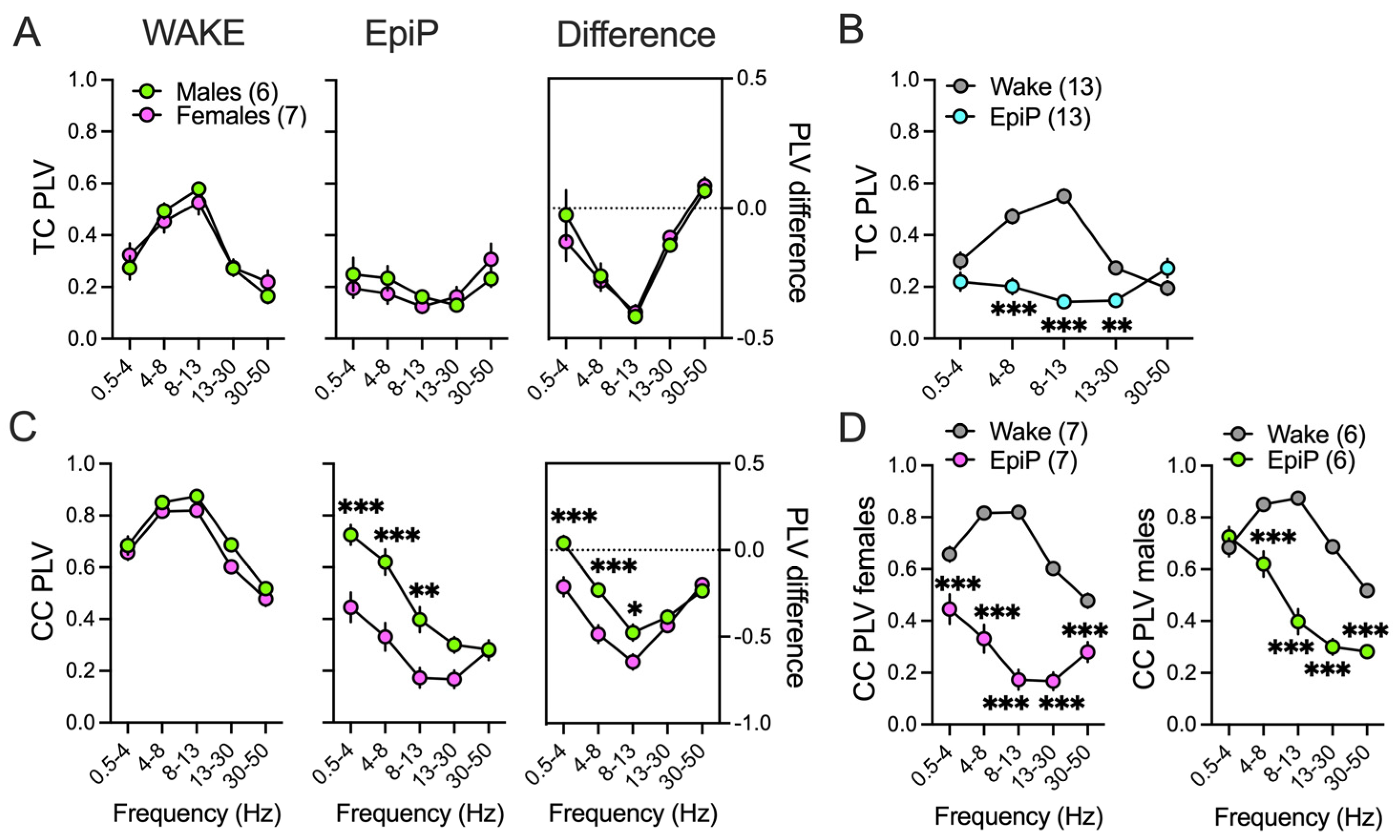
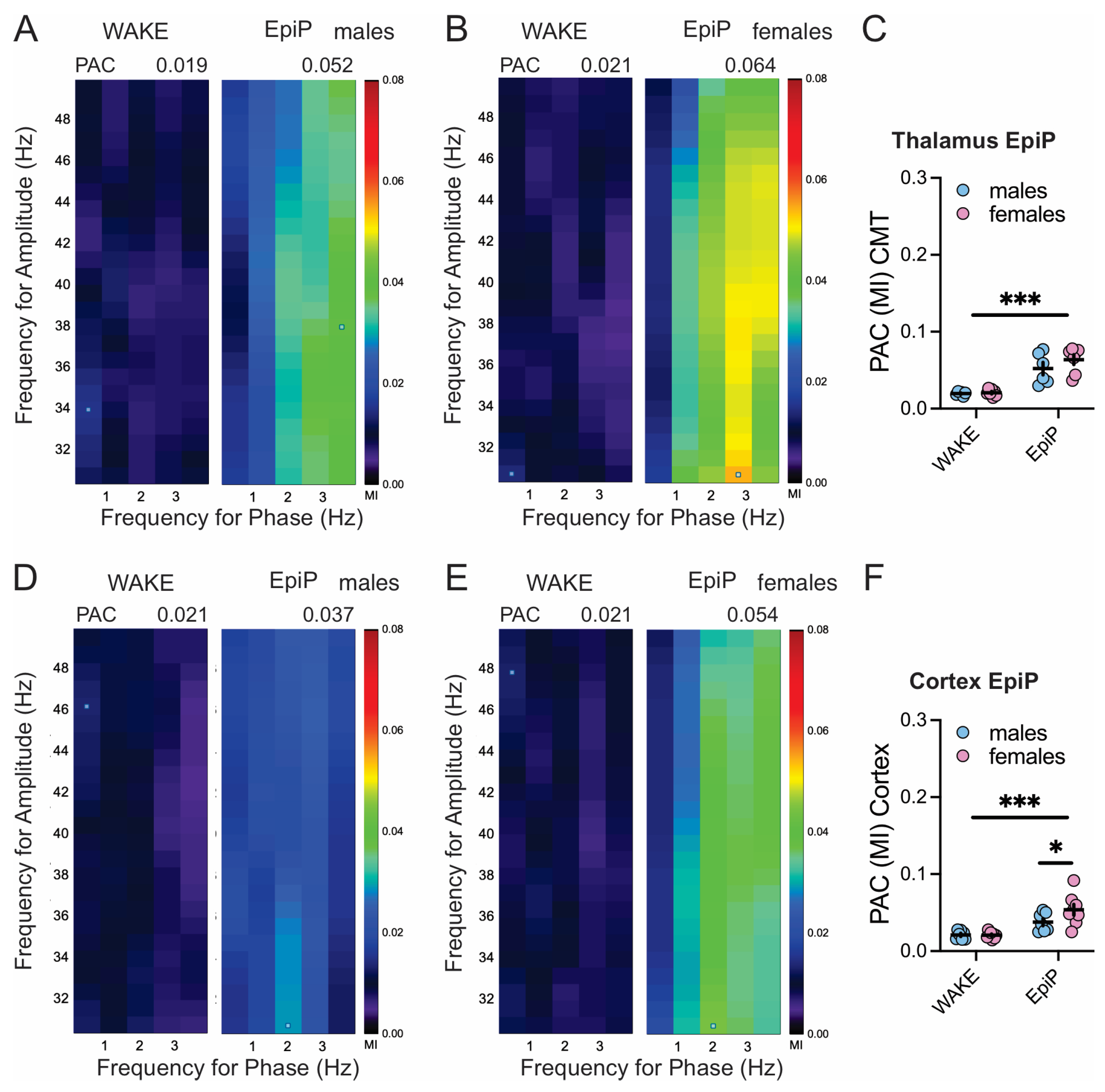
3.3. Sex Differences in Brain Synchronization and Phase–Amplitude Coupling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EpiP | Epipregnanolone |
| LORR | Loss of righting reflex |
| EEG | Electroencephalogram |
| LFP | Local field potential |
| CMT | Central medial nucleus of thalamus |
| GABAA | γ-aminobutyric acid type A receptor |
| GABA | γ-aminobutyric acid type |
| MI | Modulation index |
| PAC | Phase–amplitude coupling |
| TC | Thalamocortical |
| CC | Corticocortical |
| PLV | Phase locking value |
| 3β-HSD | 3β-hydroxysteroid dehydrogenase |
| RM | Repeated measure |
| NAS | Neuroactive steroid |
References
- Reddy, D.S. Neurosteroids: Endogenous Role in the Human Brian and Therapeutic Potentials. Prog. Brain Res. 2010, 186, 113. [Google Scholar] [CrossRef] [PubMed]
- Penning, T.M.; Covey, D.F. 5β-Dihydrosteroids: Formation and Properties. Int. J. Mol. Sci. 2024, 25, 8857. [Google Scholar] [CrossRef] [PubMed]
- Coulter, I.; Timic Stamenic, T.; Eggan, P.; Fine, B.R.; Corrigan, T.; Covey, D.F.; Yang, L.; Pan, J.Q.; Todorovic, S.M. Different Roles of T-Type Calcium Channel Isoforms in Hypnosis Induced by an Endogenous Neurosteroid Epipregnanolone. Neuropharmacology 2021, 197, 108739. [Google Scholar] [CrossRef] [PubMed]
- Bukanova, J.; Solntseva, E.; Kondratenko, R.; Kudova, E. Epipregnanolone as a Positive Modulator of GABAA Receptor in Rat Cerebellar and Hippocampus Neurons. Biomolecules 2021, 11, 791. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.S.; Scott, S.; Masiulis, S.; De Colibus, L.; Pardon, E.; Steyaert, J.; Aricescu, A.R. Structural Basis for GABAA Receptor Potentiation by Neurosteroids. Nat. Struct. Mol. Biol. 2017, 24, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.J.; Belelli, D.; Peden, D.R.; Vardy, A.W.; Peters, J.A. Neurosteroid Modulation of GABA A Receptors. Prog. Neurobiol. 2003, 71, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Maguire, J.L.; Mennerick, S. Neurosteroids: Mechanistic Considerations and Clinical Prospects. Neuropsychopharmacology 2024, 49, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; He, Y.; Eisenman, L.N.; Fields, C.; Zeng, C.-M.; Mathews, J.; Benz, A.; Fu, T.; Zorumski, E.; Steinbach, J.H.; et al. 3β-Hydroxypregnane Steroids Are Pregnenolone Sulfate-like GABAA Receptor Antagonists. J. Neurosci. 2002, 22, 3366–3375. [Google Scholar] [CrossRef] [PubMed]
- Strömberg, J.; Haage, D.; Taube, M.; Bäckström, T.; Lundgren, P. Neurosteroid Modulation of Allopregnanolone and GABA Effect on the GABA-A Receptor. Neuroscience 2006, 143, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Atluri, N.; Joksimovic, S.M.; Oklopcic, A.; Milanovic, D.; Klawitter, J.; Eggan, P.; Krishnan, K.; Covey, D.F.; Todorovic, S.M.; Jevtovic-Todorovic, V. A Neurosteroid Analogue with T-Type Calcium Channel Blocking Properties Is an Effective Hypnotic, but Is Not Harmful to Neonatal Rat Brain. Br. J. Anaesth. 2018, 120, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Joksimovic, S.M.; Sampath, D.; Krishnan, K.; Covey, D.F.; Jevtovic-Todorovic, V.; Raol, Y.H.; Todorovic, S.M. Differential Effects of the Novel Neurosteroid Hypnotic (3β,5β,17β)-3-Hydroxyandrostane-17-Carbonitrile on Electroencephalogram Activity in Male and Female Rats. Br. J. Anaesth. 2021, 127, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Timic Stamenic, T.; Manzella, F.M.; Maksimovic, S.; Krishnan, K.; Covey, D.F.; Jevtovic-Todorovic, V.; Todorovic, S.M. Further Evidence That Inhibition of Neuronal Voltage-Gated Calcium Channels Contributes to the Hypnotic Effect of Neurosteroid Analogue, 3β-OH. Front. Pharmacol. 2022, 13, 850658. [Google Scholar] [CrossRef] [PubMed]
- Timic Stamenic, T.; Feseha, S.; Manzella, F.M.; Wallace, D.; Wilkey, D.; Corrigan, T.; Fiedler, H.; Doerr, P.; Krishnan, K.; Raol, Y.H.; et al. The T-Type Calcium Channel Isoform Cav3.1 Is a Target for the Hypnotic Effect of the Anaesthetic Neurosteroid (3β,5β,17β)-3-Hydroxyandrostane-17-Carbonitrile. Br. J. Anaesth. 2021, 126, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Coulter, I.; Cox, R.; Covey, D.F.; Todorovic, S.M.; Timic Stamenic, T. Comparative Electrophysiological Study of Neuroactive Steroid-Induced Hypnosis in Mice: Sex and Drug-Specific Differences. Exp. Biol. Med. 2025, 250, 10550. [Google Scholar] [CrossRef] [PubMed]
- McCarren, H.S.; Moore, J.T.; Kelz, M.B. Assessing Changes in Volatile General Anesthetic Sensitivity of Mice after Local or Systemic Pharmacological Intervention. J. Vis. Exp. 2013, 80, e51079. [Google Scholar] [CrossRef] [PubMed]
- Tadel, F.; Bock, E.; Niso, G.; Mosher, J.C.; Cousineau, M.; Pantazis, D.; Leahy, R.M.; Baillet, S. MEG/EEG Group Analysis with Brainstorm. Front. Neurosci. 2019, 13, 435877. [Google Scholar] [CrossRef] [PubMed]
- Bastos, A.M.; Schoffelen, J.M. A Tutorial Review of Functional Connectivity Analysis Methods and Their Interpretational Pitfalls. Front. Syst. Neurosci. 2016, 9, 165147. [Google Scholar] [CrossRef] [PubMed]
- Manzella, F.M.; Cabrera, O.H.; Wilkey, D.; Fine-Raquet, B.; Klawitter, J.; Krishnan, K.; Covey, D.F.; Jevtovic-Todorovic, V.; Todorovic, S.M. Sex-Specific Hypnotic Effects of the Neuroactive Steroid (3β,5β,17β)-3-Hydroxyandrostane-17-Carbonitrile Are Mediated by Peripheral Metabolism into an Active Hypnotic Steroid. Br. J. Anaesth. 2023, 130, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Ren, Y.; Yan, J.; Li, D.; Voss, L.J.; Sleigh, J.W.; Li, X. A Comparison of Different Synchronization Measures in Electroencephalogram during Propofol Anesthesia. J. Clin. Monit. Comput. 2016, 30, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, M.; Seppänen, T.; Tuukkanen, J.; Yli-Hankala, A.; Jäntti, V. Propofol Anesthesia Induces Phase Synchronization Changes in EEG. Clin. Neurophysiol. 2001, 112, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Steyn-Ross, M.L.; Steyn-Ross, D.A.; Wilson, M.T.; Sleigh, J.W. EEG Slow-Wave Coherence Changes in Propofol-Induced General Anesthesia: Experiment and Theory. Front. Syst. Neurosci. 2014, 8, 107602. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, N.; Georgiou, J. Spatial Analytic Phase Difference of EEG Activity during Anesthetic-Induced Unconsciousness. Clin. Neurophysiol. 2014, 125, 2122–2131. [Google Scholar] [CrossRef] [PubMed]
- Tsai, F.F.; Fan, S.Z.; Cheng, H.L.; Yeh, J.R. Multi-Timescale Phase-Amplitude Couplings in Transitions of Anesthetic-Induced Unconsciousness. Sci. Rep. 2019, 9, 7815. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; Silverstein, B.H.; Sharba, L.; Li, D.; Hambrecht-Wiedbusch, V.S.; Hudetz, A.G.; Mashour, G.A. Propofol, Sevoflurane, and Ketamine Induce a Reversible Increase in Delta-Gamma and Theta-Gamma Phase-Amplitude Coupling in Frontal Cortex of Rat. Front. Syst. Neurosci. 2017, 11, 258424. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wu, D.; Bahuri, N.F.A.; Wang, S.; Hyam, J.A.; Yarrow, S.; FitzGerald, J.J.; Aziz, T.Z.; Green, A.L. Spectral and Phase-Amplitude Coupling Signatures in Human Deep Brain Oscillations during Propofol-Induced Anaesthesia. Br. J. Anaesth. 2018, 121, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Zhang, D.; Wei, Q.; Wang, G.; Huang, F.; Chen, X.; Muhammad, K.G.; Sun, Y.; Liu, J. Intrinsic Phase–Amplitude Coupling on Multiple Spatial Scales during the Loss and Recovery of Consciousness. Comput. Biol. Med. 2022, 147, 105687. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, L.; Desowska, A.; Berde, C.B.; Cornelissen, L. Electroencephalographic Delta and Alpha Oscillations Reveal Phase-Amplitude Coupling in Paediatric Patients Undergoing Sevoflurane-Based General Anaesthesia. Br. J. Anaesth. 2023, 130, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Sze, Y.; Gill, A.C.; Brunton, P.J. Sex-Dependent Changes in Neuroactive Steroid Concentrations in the Rat Brain Following Acute Swim Stress. J. Neuroendocrinol. 2018, 30, e12644. [Google Scholar] [CrossRef] [PubMed]
- Caruso, D.; D’Intino, G.; Giatti, S.; Maschi, O.; Pesaresi, M.; Calabrese, D.; Garcia-Segura, L.M.; Calza, L.; Melcangi, R.C. Sex-Dimorphic Changes in Neuroactive Steroid Levels after Chronic Experimental Autoimmune Encephalomyelitis. J. Neurochem. 2010, 114, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Caruso, D.; Pesaresi, M.; Maschi, O.; Giatti, S.; Garcia-Segura, L.M.; Melcangi, R.C. Effect of Short-and Long-Term Gonadectomy on Neuroactive Steroid Levels in the Central and Peripheral Nervous System of Male and Female Rats. J. Neuroendocrinol. 2010, 22, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Pisu, M.G.; Concas, L.; Siddi, C.; Serra, M.; Porcu, P. The Allopregnanolone Response to Acute Stress in Females: Preclinical and Clinical Studies. Biomolecules 2022, 12, 1262. [Google Scholar] [CrossRef] [PubMed]
- Vallée, M.; Rivera, J.D.; Koob, G.F.; Purdy, R.H.; Fitzgerald, R.L. Quantification of Neurosteroids in Rat Plasma and Brain Following Swim Stress and Allopregnanolone Administration Using Negative Chemical Ionization Gas Chromatography/Mass Spectrometry. Anal. Biochem. 2000, 287, 153–166. [Google Scholar] [CrossRef] [PubMed]
- van Broekhoven, F.; Bäckström, T.; van Luijtelaar, G.; Buitelaar, J.K.; Smits, P.; Verkes, R.J. Effects of Allopregnanolone on Sedation in Men, and in Women on Oral Contraceptives. Psychoneuroendocrinology 2007, 32, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Diviccaro, S.; Cioffi, L.; Falvo, E.; Giatti, S.; Melcangi, R.C. Allopregnanolone: An Overview on Its Synthesis and Effects. J. Neuroendocrinol. 2022, 34, e12996. [Google Scholar] [CrossRef] [PubMed]
- Skilbeck, K.J.; Hinton, T.; Johnston, G.A.R. Sex-Differences and Stress: Effects on Regional High and Low Affinity [3H]GABA Binding. Neurochem. Int. 2008, 52, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Giatti, S.; Diviccaro, S.; Garcia-Segura, L.M.; Melcangi, R.C. Sex Differences in the Brain Expression of Steroidogenic Molecules under Basal Conditions and after Gonadectomy. J. Neuroendocrinol. 2019, 31, e12736. [Google Scholar] [CrossRef] [PubMed]
- Giatti, S.; Garcia-Segura, L.M.; Barreto, G.E.; Melcangi, R.C. Neuroactive Steroids, Neurosteroidogenesis and Sex. Prog. Neurobiol. 2019, 176, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Samba Reddy, D. Sex Differences in the Anticonvulsant Activity of Neurosteroids. J. Neurosci. Res. 2017, 95, 661. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timic Stamenic, T.; Coulter, I.; Covey, D.F.; Todorovic, S.M. Electrophysiological Characterization of Sex-Dependent Hypnosis by an Endogenous Neuroactive Steroid Epipregnanolone. Biomolecules 2025, 15, 1033. https://doi.org/10.3390/biom15071033
Timic Stamenic T, Coulter I, Covey DF, Todorovic SM. Electrophysiological Characterization of Sex-Dependent Hypnosis by an Endogenous Neuroactive Steroid Epipregnanolone. Biomolecules. 2025; 15(7):1033. https://doi.org/10.3390/biom15071033
Chicago/Turabian StyleTimic Stamenic, Tamara, Ian Coulter, Douglas F. Covey, and Slobodan M. Todorovic. 2025. "Electrophysiological Characterization of Sex-Dependent Hypnosis by an Endogenous Neuroactive Steroid Epipregnanolone" Biomolecules 15, no. 7: 1033. https://doi.org/10.3390/biom15071033
APA StyleTimic Stamenic, T., Coulter, I., Covey, D. F., & Todorovic, S. M. (2025). Electrophysiological Characterization of Sex-Dependent Hypnosis by an Endogenous Neuroactive Steroid Epipregnanolone. Biomolecules, 15(7), 1033. https://doi.org/10.3390/biom15071033