

Multimodal Function of Mesenchymal Stem Cells in Psoriasis Treatment

 and
and 
Abstract
1. Introduction
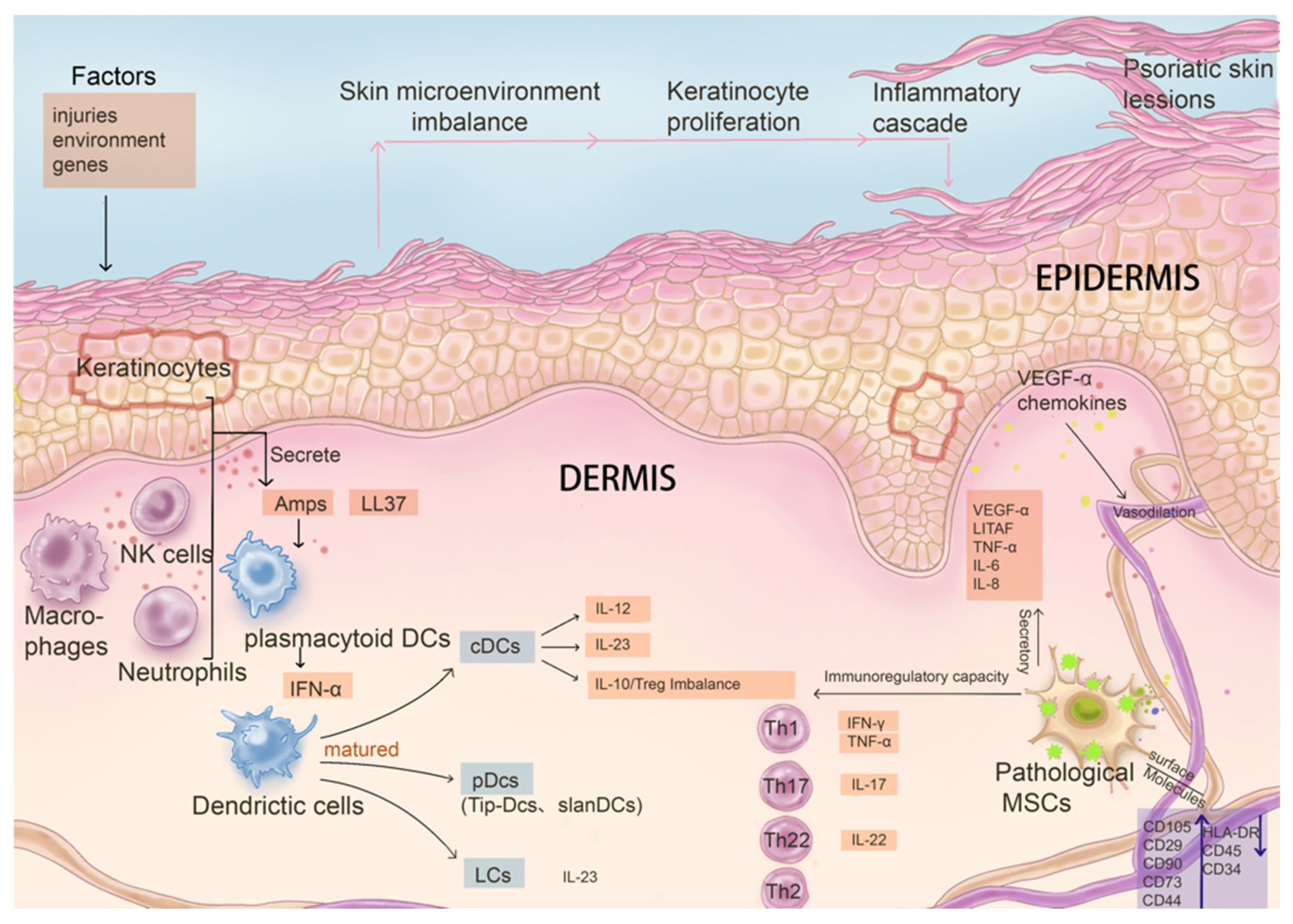
2. The Main Aspects of Psoriasis Pathogenesis
2.1. Keratinocytes
2.2. Dysregulation of Dendritic Cell Function
2.3. Abnormal Activation of T Cells and Key Cytokines in the Inflammatory Cycle
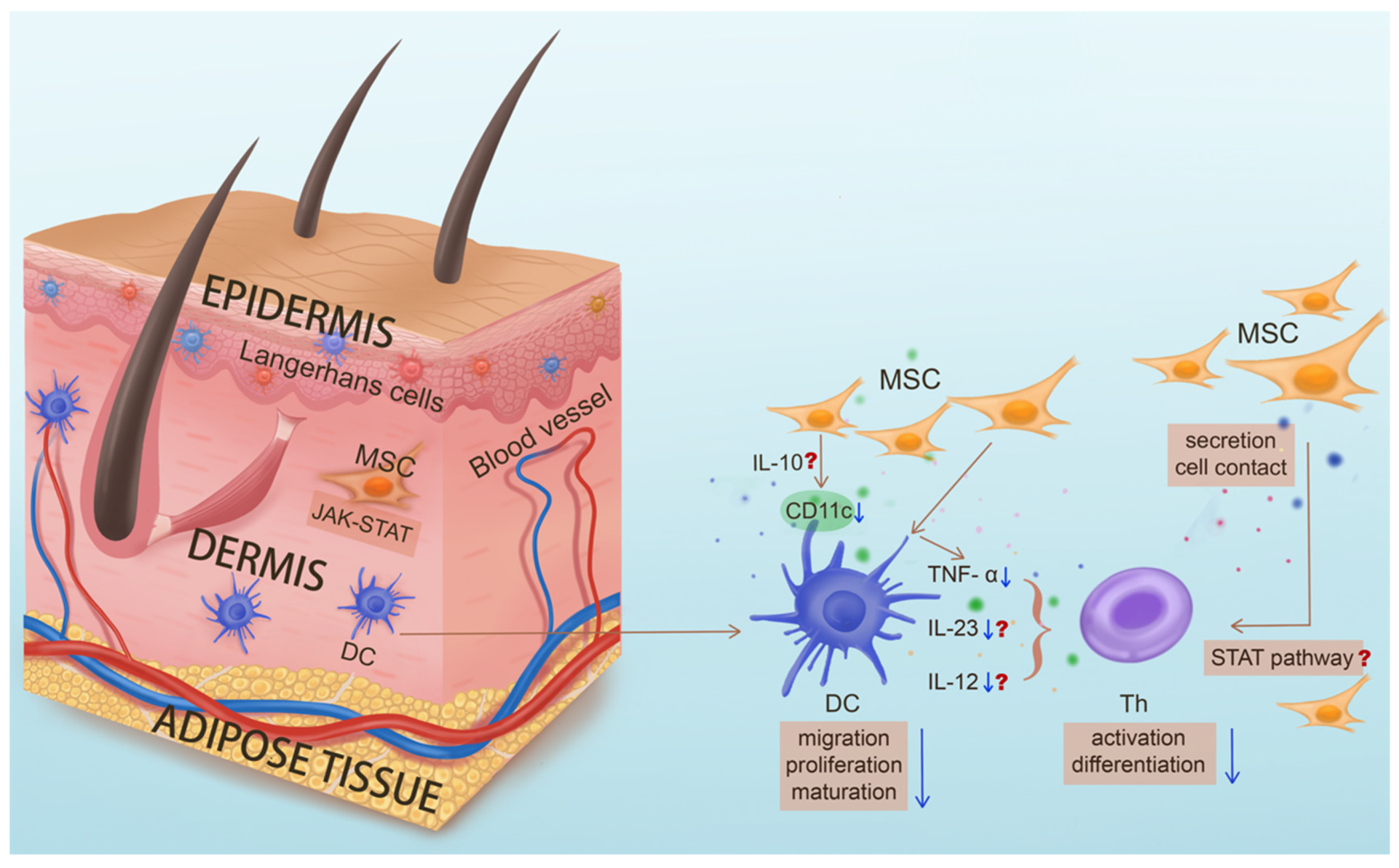
3. Multidimensional Functions of Mesenchymal Stem Cells in Skin Homeostasis and Psoriasis Development
3.1. The Essential Role of MSCs in Skin Homeostasis
3.2. Pathological Changes of Resident MSCs in Psoriatic Skin Lesions
3.2.1. Abnormal Interaction with Psoriatic Keratinocytes
3.2.2. Changes in Immunomodulatory Capacity
3.2.3. Pro-Angiogenesis and Pro-Inflammatory Mediators Increased
3.2.4. Decreased Antioxidant Capacity
4. The Therapeutic Effect of MSCs in Psoriasis
4.1. Adipose-Derived Mesenchymal Stem Cells
4.1.1. Preclinical Studies
4.1.2. Clinical Studies
4.2. Human Umbilical Cord Mesenchymal Stem Cells (hUC-MSCs)
4.2.1. Preclinical Studies
4.2.2. Clinical Studies
4.3. Human Embryonic Mesenchymal Stem Cells (hE-MSCs)
4.3.1. Preclinical Studies
4.3.2. Clinical Studies
4.4. Gingival Mesenchymal Stem Cells (GMSCs)
4.4.1. Preclinical Studies
4.4.2. Clinical Studies
4.5. Human Amniotic Membrane-Derived Mesenchymal Stem Cells (hAMSC)
4.5.1. Preclinical Studies
4.5.2. Clinical Studies
4.6. Human Tonsil-Derived Mesenchymal Stem Cells (T-MSC)
4.6.1. Preclinical Studies
4.6.2. Clinical Studies
4.7. Summary of MSC-Based Psoriasis Treatments
4.8. MSCs Targeting Pathogenic DCs
5. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lowes, M.A.; Suárez-Fariñas, M.; Krueger, J.G. Immunology of psoriasis. Annu. Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef] [PubMed]
- Boehncke, W.H.; Schön, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Bocheńska, K.; Smolińska, E.; Moskot, M.; Jakóbkiewicz-Banecka, J.; Gabig-Cimińska, M. Models in the Research Process of Psoriasis. Int. J. Mol. Sci. 2017, 18, 2514. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Taylor, C.; Kornmehl, H.; Armstrong, A.W. Psoriasis and suicidality: A systematic review and meta-analysis. J. Am. Acad. Dermatol. 2017, 77, 425–440.e422. [Google Scholar] [CrossRef]
- Springate, D.A.; Parisi, R.; Kontopantelis, E.; Reeves, D.; Griffiths, C.E.; Ashcroft, D.M. Incidence, prevalence and mortality of patients with psoriasis: A U.K. population-based cohort study. Br. J. Dermatol. 2017, 176, 650–658. [Google Scholar] [CrossRef]
- Ryan, C.; Sadlier, M.; De Vol, E.; Patel, M.; Lloyd, A.A.; Day, A.; Lally, A.; Kirby, B.; Menter, A. Genital psoriasis is associated with significant impairment in quality of life and sexual functioning. J. Am. Acad. Dermatol. 2015, 72, 978–983. [Google Scholar] [CrossRef]
- Michalek, I.M.; Loring, B.; John, S.M. A systematic review of worldwide epidemiology of psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 205–212. [Google Scholar] [CrossRef]
- Armstrong, A.W.; Read, C. Pathophysiology, Clinical Presentation, and Treatment of Psoriasis: A Review. JAMA 2020, 323, 1945–1960. [Google Scholar] [CrossRef]
- Ghoreschi, K.; Balato, A.; Enerbäck, C.; Sabat, R. Therapeutics targeting the IL-23 and IL-17 pathway in psoriasis. Lancet 2021, 397, 754–766. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Bogatcheva, N.V.; Coleman, M.E. Conditioned Medium of Mesenchymal Stromal Cells: A New Class of Therapeutics. Biochem. Mosc. 2019, 84, 1375–1389. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Scholtemeijer, M.; Shah, K. Mesenchymal Stem Cell Immunomodulation: Mechanisms and Therapeutic Potential. Trends Pharmacol. Sci. 2020, 41, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Chiu, S.M.; Motan, D.A.; Zhang, Z.; Chen, L.; Ji, H.L.; Tse, H.F.; Fu, Q.L.; Lian, Q. Mesenchymal stem cells and immunomodulation: Current status and future prospects. Cell Death Dis. 2016, 7, e2062. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, J.; Li, Z.; Zheng, J.; Sun, Q. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Alleviate Psoriasis-like Skin Inflammation. J. Interferon Cytokine Res. 2022, 42, 8–18. [Google Scholar] [CrossRef]
- Chen, M.; Peng, J.; Xie, Q.; Xiao, N.; Su, X.; Mei, H.; Lu, Y.; Zhou, J.; Dai, Y.; Wang, S.; et al. Mesenchymal Stem Cells Alleviate Moderate-to-Severe Psoriasis by Reducing the Production of Type I Interferon (IFN-I) by Plasmacytoid Dendritic Cells (pDCs). Stem Cells Int. 2019, 2019, 6961052. [Google Scholar] [CrossRef]
- Tokuyama, M.; Mabuchi, T. New Treatment Addressing the Pathogenesis of Psoriasis. Int. J. Mol. Sci. 2020, 21, 7488. [Google Scholar] [CrossRef]
- Nestle, F.O.; Conrad, C.; Tun-Kyi, A.; Homey, B.; Gombert, M.; Boyman, O.; Burg, G.; Liu, Y.J.; Gilliet, M. Plasmacytoid predendritic cells initiate psoriasis through interferon-alpha production. J. Exp. Med. 2005, 202, 135–143. [Google Scholar] [CrossRef]
- Tohyama, M.; Yang, L.; Hanakawa, Y.; Dai, X.; Shirakata, Y.; Sayama, K. IFN-α enhances IL-22 receptor expression in keratinocytes: A possible role in the development of psoriasis. J. Investig. Dermatol. 2012, 132, 1933–1935. [Google Scholar] [CrossRef]
- Rauschenberger, T.; Schmitt, V.; Azeem, M.; Klein-Hessling, S.; Murti, K.; Grän, F.; Goebeler, M.; Kerstan, A.; Klein, M.; Bopp, T.; et al. T Cells Control Chemokine Secretion by Keratinocytes. Front. Immunol. 2019, 10, 1917. [Google Scholar] [CrossRef]
- Albanesi, C.; Madonna, S.; Gisondi, P.; Girolomoni, G. The Interplay Between Keratinocytes and Immune Cells in the Pathogenesis of Psoriasis. Front. Immunol. 2018, 9, 1549. [Google Scholar] [CrossRef]
- Li, N.; Yamasaki, K.; Saito, R.; Fukushi-Takahashi, S.; Shimada-Omori, R.; Asano, M.; Aiba, S. Alarmin function of cathelicidin antimicrobial peptide LL37 through IL-36γ induction in human epidermal keratinocytes. J. Immunol. 2014, 193, 5140–5148. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.T.; Cao, L.; Roberson, E.D.; Duan, S.; Helms, C.A.; Nair, R.P.; Duffin, K.C.; Stuart, P.E.; Goldgar, D.; Hayashi, G.; et al. Rare and common variants in CARD14, encoding an epidermal regulator of NF-kappaB, in psoriasis. Am. J. Hum. Genet. 2012, 90, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, R.; Dainichi, T.; Tsuchiya, S.; Nomura, T.; Kitoh, A.; Hayden, M.S.; Ishii, K.J.; Tanaka, M.; Honda, T.; Egawa, G.; et al. Epithelial TRAF6 drives IL-17-mediated psoriatic inflammation. JCI Insight 2018, 3, e121175. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.R.; Cho, D.H.; Park, H.J. Molecular Mechanisms and Management of a Cutaneous Inflammatory Disorder: Psoriasis. Int. J. Mol. Sci. 2017, 18, 2684. [Google Scholar] [CrossRef]
- Sano, S.; Chan, K.S.; Carbajal, S.; Clifford, J.; Peavey, M.; Kiguchi, K.; Itami, S.; Nickoloff, B.J.; DiGiovanni, J. Stat3 links activated keratinocytes and immunocytes required for development of psoriasis in a novel transgenic mouse model. Nat. Med. 2005, 11, 43–49. [Google Scholar] [CrossRef]
- Ravipati, A.; Nolan, S.; Alphonse, M.; Dikeman, D.; Youn, C.; Wang, Y.; Orlando, N.; Patrick, G.; Lee, S.; Ortines, R.V.; et al. IL-6R/Signal Transducer and Activator of Transcription 3 Signaling in Keratinocytes rather than in T Cells Induces Psoriasis-Like Dermatitis in Mice. J. Investig. Dermatol. 2022, 142, 1126–1135.e1124. [Google Scholar] [CrossRef] [PubMed]
- Lizzul, P.F.; Aphale, A.; Malaviya, R.; Sun, Y.; Masud, S.; Dombrovskiy, V.; Gottlieb, A.B. Differential expression of phosphorylated NF-kappaB/RelA in normal and psoriatic epidermis and downregulation of NF-kappaB in response to treatment with etanercept. J. Investig. Dermatol. 2005, 124, 1275–1283. [Google Scholar] [CrossRef]
- Mose, M.; Kang, Z.; Raaby, L.; Iversen, L.; Johansen, C. TNFα- and IL-17A-mediated S100A8 expression is regulated by p38 MAPK. Exp. Dermatol. 2013, 22, 476–481. [Google Scholar] [CrossRef]
- Sakurai, K.; Dainichi, T.; Garcet, S.; Tsuchiya, S.; Yamamoto, Y.; Kitoh, A.; Honda, T.; Nomura, T.; Egawa, G.; Otsuka, A.; et al. Cutaneous p38 mitogen-activated protein kinase activation triggers psoriatic dermatitis. J. Allergy Clin. Immunol. 2019, 144, 1036–1049. [Google Scholar] [CrossRef]
- Yang, L.; Fan, X.; Cui, T.; Dang, E.; Wang, G. Nrf2 Promotes Keratinocyte Proliferation in Psoriasis through Up-Regulation of Keratin 6, Keratin 16, and Keratin 17. J. Investig. Dermatol. 2017, 137, 2168–2176. [Google Scholar] [CrossRef] [PubMed]
- Zolotarenko, A.; Chekalin, E.; Piruzian, E.; Bruskin, S. FRA1 mediates the activation of keratinocytes: Implications for the development of psoriatic plaques. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3726–3734. [Google Scholar] [CrossRef] [PubMed]
- Gordon, W.M.; Zeller, M.D.; Klein, R.H.; Swindell, W.R.; Ho, H.; Espetia, F.; Gudjonsson, J.E.; Baldi, P.F.; Andersen, B. A GRHL3-regulated repair pathway suppresses immune-mediated epidermal hyperplasia. J. Clin. Investig. 2014, 124, 5205–5218. [Google Scholar] [CrossRef]
- Griffiths, C.E.; Barker, J.N. Pathogenesis and clinical features of psoriasis. Lancet 2007, 370, 263–271. [Google Scholar] [CrossRef]
- Hawkes, J.E.; Chan, T.C.; Krueger, J.G. Psoriasis pathogenesis and the development of novel targeted immune therapies. J. Allergy Clin. Immunol. 2017, 140, 645–653. [Google Scholar] [CrossRef]
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Harden, J.L.; Krueger, J.G.; Bowcock, A.M. The immunogenetics of Psoriasis: A comprehensive review. J. Autoimmun. 2015, 64, 66–73. [Google Scholar] [CrossRef]
- Rendon, A.; Schäkel, K. Psoriasis Pathogenesis and Treatment. Int. J. Mol. Sci. 2019, 20, 1475. [Google Scholar] [CrossRef]
- Steinman, R.M.; Banchereau, J. Taking dendritic cells into medicine. Nature 2007, 449, 419–426. [Google Scholar] [CrossRef]
- Wang, A.; Bai, Y. Dendritic cells: The driver of psoriasis. J. Dermatol. 2020, 47, 104–113. [Google Scholar] [CrossRef]
- Macri, C.; Pang, E.S.; Patton, T.; O’Keeffe, M. Dendritic cell subsets. Semin. Cell Dev. Biol. 2018, 84, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Kolbeck, R.; Sanjuan, M.A. Plasmacytoid dendritic cells in autoimmunity. Curr. Opin. Immunol. 2017, 44, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Kopfnagel, V.; Wagenknecht, S.; Harder, J.; Hofmann, K.; Kleine, M.; Buch, A.; Sodeik, B.; Werfel, T. RNase 7 Strongly Promotes TLR9-Mediated DNA Sensing by Human Plasmacytoid Dendritic Cells. J. Investig. Dermatol. 2018, 138, 872–881. [Google Scholar] [CrossRef]
- Lande, R.; Gilliet, M. Plasmacytoid dendritic cells: Key players in the initiation and regulation of immune responses. Ann. N. Y. Acad. Sci. 2010, 1183, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cao, X. Regulatory dendritic cells in autoimmunity: A comprehensive review. J. Autoimmun. 2015, 63, 1–12. [Google Scholar] [CrossRef]
- Buhl, T.; Saleh, M.M.; Schön, M.P. More tolerance for dendritic cells in psoriasis. Exp. Dermatol. 2017, 26, 335–337. [Google Scholar] [CrossRef]
- Martini, E.; Wikén, M.; Cheuk, S.; Gallais Sérézal, I.; Baharom, F.; Ståhle, M.; Smed-Sörensen, A.; Eidsmo, L. Dynamic Changes in Resident and Infiltrating Epidermal Dendritic Cells in Active and Resolved Psoriasis. J. Investig. Dermatol. 2017, 137, 865–873. [Google Scholar] [CrossRef]
- Casciano, F.; Pigatto, P.D.; Secchiero, P.; Gambari, R.; Reali, E. T Cell Hierarchy in the Pathogenesis of Psoriasis and Associated Cardiovascular Comorbidities. Front. Immunol. 2018, 9, 1390. [Google Scholar] [CrossRef]
- Lee, E.; Trepicchio, W.L.; Oestreicher, J.L.; Pittman, D.; Wang, F.; Chamian, F.; Dhodapkar, M.; Krueger, J.G. Increased expression of interleukin 23 p19 and p40 in lesional skin of patients with psoriasis vulgaris. J. Exp. Med. 2004, 199, 125–130. [Google Scholar] [CrossRef]
- Stritesky, G.L.; Yeh, N.; Kaplan, M.H. IL-23 promotes maintenance but not commitment to the Th17 lineage. J. Immunol. 2008, 181, 5948–5955. [Google Scholar] [CrossRef]
- Ghoreschi, K.; Laurence, A.; Yang, X.P.; Tato, C.M.; McGeachy, M.J.; Konkel, J.E.; Ramos, H.L.; Wei, L.; Davidson, T.S.; Bouladoux, N.; et al. Generation of pathogenic T(H)17 cells in the absence of TGF-β signalling. Nature 2010, 467, 967–971. [Google Scholar] [CrossRef] [PubMed]
- Fotiadou, C.; Lazaridou, E.; Sotiriou, E.; Ioannides, D. Targeting IL-23 in psoriasis: Current perspectives. Psoriasis Targets Ther. 2018, 8, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Beringer, A.; Noack, M.; Miossec, P. IL-17 in Chronic Inflammation: From Discovery to Targeting. Trends Mol. Med. 2016, 22, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Speeckaert, R.; Lambert, J.; Grine, L.; Van Gele, M.; De Schepper, S.; van Geel, N. The many faces of interleukin-17 in inflammatory skin diseases. Br. J. Dermatol. 2016, 175, 892–901. [Google Scholar] [CrossRef]
- Johansen, C.; Usher, P.A.; Kjellerup, R.B.; Lundsgaard, D.; Iversen, L.; Kragballe, K. Characterization of the interleukin-17 isoforms and receptors in lesional psoriatic skin. Br. J. Dermatol. 2009, 160, 319–324. [Google Scholar] [CrossRef]
- Miossec, P.; Korn, T.; Kuchroo, V.K. Interleukin-17 and type 17 helper T cells. N. Engl. J. Med. 2009, 361, 888–898. [Google Scholar] [CrossRef]
- Hawkes, J.E.; Yan, B.Y.; Chan, T.C.; Krueger, J.G. Discovery of the IL-23/IL-17 Signaling Pathway and the Treatment of Psoriasis. J. Immunol. 2018, 201, 1605–1613. [Google Scholar] [CrossRef]
- Perera, G.K.; Di Meglio, P.; Nestle, F.O. Psoriasis. Annu. Rev. Pathol. 2012, 7, 385–422. [Google Scholar] [CrossRef]
- Chiricozzi, A.; Guttman-Yassky, E.; Suárez-Fariñas, M.; Nograles, K.E.; Tian, S.; Cardinale, I.; Chimenti, S.; Krueger, J.G. Integrative responses to IL-17 and TNF-α in human keratinocytes account for key inflammatory pathogenic circuits in psoriasis. J. Investig. Dermatol. 2011, 131, 677–687. [Google Scholar] [CrossRef]
- Sereflican, B.; Goksugur, N.; Bugdayci, G.; Polat, M.; Haydar Parlak, A. Serum Visfatin, Adiponectin, and Tumor Necrosis Factor Alpha (TNF-α) Levels in Patients with Psoriasis and their Correlation with Disease Severity. Acta Dermatovenerol. Croat. 2016, 24, 13–19. [Google Scholar]
- Grine, L.; Dejager, L.; Libert, C.; Vandenbroucke, R.E. An inflammatory triangle in psoriasis: TNF, type I IFNs and IL-17. Cytokine Growth Factor Rev. 2015, 26, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Grän, F.; Kerstan, A.; Serfling, E.; Goebeler, M.; Muhammad, K. Current Developments in the Immunology of Psoriasis. Yale J. Biol. Med. 2020, 93, 97–110. [Google Scholar] [PubMed]
- Hao, J.Q. Targeting interleukin-22 in psoriasis. Inflammation 2014, 37, 94–99. [Google Scholar] [CrossRef]
- Chiricozzi, A.; Romanelli, P.; Volpe, E.; Borsellino, G.; Romanelli, M. Scanning the Immunopathogenesis of Psoriasis. Int. J. Mol. Sci. 2018, 19, 179. [Google Scholar] [CrossRef]
- Iskandar, I.Y.K.; Warren, R.B.; Lunt, M.; Mason, K.J.; Evans, I.; McElhone, K.; Smith, C.H.; Reynolds, N.J.; Ashcroft, D.M.; Griffiths, C.E.M. Differential Drug Survival of Second-Line Biologic Therapies in Patients with Psoriasis: Observational Cohort Study from the British Association of Dermatologists Biologic Interventions Register (BADBIR). J. Investig. Dermatol. 2018, 138, 775–784. [Google Scholar] [CrossRef]
- Topaloğlu Demir, F.; Özkök Akbulut, T.; Kıvanç Altunay, İ.; Aytekin, S.; Oğuz Topal, İ.; Kara Polat, A.; Özkur, E.; Karadağ, A.S. Evaluation of the adverse effects of biological agents used in the treatment of psoriasis: A multicenter retrospective cohort study. Dermatol. Ther. 2020, 33, e14216. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Z.; Sun, J.; Ma, T.; Hua, F.; Shen, Z. A brief review of cytotoxicity of nanoparticles on mesenchymal stem cells in regenerative medicine. Int. J. Nanomed. 2019, 14, 3875–3892. [Google Scholar] [CrossRef]
- Li, H.; Tian, Y.; Xie, L.; Liu, X.; Huang, Z.; Su, W. Mesenchymal stem cells in allergic diseases: Current status. Allergol. Int. 2020, 69, 35–45. [Google Scholar] [CrossRef]
- Auerswald, S.; Schreml, S.; Meier, R.; Blancke Soares, A.; Niyazi, M.; Marschner, S.; Belka, C.; Canis, M.; Haubner, F. Wound monitoring of pH and oxygen in patients after radiation therapy. Radiat. Oncol. 2019, 14, 199. [Google Scholar] [CrossRef]
- Caplan, A.I. Why are MSCs therapeutic? New data: New insight. J. Pathol. 2009, 217, 318–324. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Y.; Li, Q.; Liu, K.; Hou, J.; Shao, C.; Wang, Y. Immunoregulatory mechanisms of mesenchymal stem and stromal cells in inflammatory diseases. Nat. Rev. Nephrol. 2018, 14, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Abe, R.; Fujita, Y.; Ando, S.; Inokuma, D.; Shimizu, H. Mesenchymal stem cells are recruited into wounded skin and contribute to wound repair by transdifferentiation into multiple skin cell type. J. Immunol. 2008, 180, 2581–2587. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.M.; Ritter, T.; Ceredig, R.; Griffin, M.D. Mesenchymal stem cell effects on T-cell effector pathways. Stem Cell Res. Ther. 2011, 2, 34. [Google Scholar] [CrossRef] [PubMed]
- Cheung, T.S.; Galleu, A.; von Bonin, M.; Bornhäuser, M.; Dazzi, F. Apoptotic mesenchymal stromal cells induce prostaglandin E2 in monocytes: Implications for the monitoring of mesenchymal stromal cell activity. Haematologica 2019, 104, e438–e441. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, L.; Zhang, G.; Zhang, B. Umbilical cord-matrix stem cells induce the functional restoration of vascular endothelial cells and enhance skin wound healing in diabetic mice via the polarized macrophages. Stem Cell Res. Ther. 2020, 11, 39. [Google Scholar] [CrossRef]
- Chiossone, L.; Conte, R.; Spaggiari, G.M.; Serra, M.; Romei, C.; Bellora, F.; Becchetti, F.; Andaloro, A.; Moretta, L.; Bottino, C. Mesenchymal Stromal Cells Induce Peculiar Alternatively Activated Macrophages Capable of Dampening Both Innate and Adaptive Immune Responses. Stem Cells 2016, 34, 1909–1921. [Google Scholar] [CrossRef]
- Spaggiari, G.M.; Abdelrazik, H.; Becchetti, F.; Moretta, L. MSCs inhibit monocyte-derived DC maturation and function by selectively interfering with the generation of immature DCs: Central role of MSC-derived prostaglandin E2. Blood 2009, 113, 6576–6583. [Google Scholar] [CrossRef]
- Joel, M.D.M.; Yuan, J.; Wang, J.; Yan, Y.; Qian, H.; Zhang, X.; Xu, W.; Mao, F. MSC: Immunoregulatory effects, roles on neutrophils and evolving clinical potentials. Am. J. Transl. Res. 2019, 11, 3890–3904. [Google Scholar]
- Kwon, H.M.; Hur, S.M.; Park, K.Y.; Kim, C.K.; Kim, Y.M.; Kim, H.S.; Shin, H.C.; Won, M.H.; Ha, K.S.; Kwon, Y.G.; et al. Multiple paracrine factors secreted by mesenchymal stem cells contribute to angiogenesis. Vasc. Pharmacol. 2014, 63, 19–28. [Google Scholar] [CrossRef]
- Vieira Paladino, F.; de Moraes Rodrigues, J.; da Silva, A.; Goldberg, A.C. The Immunomodulatory Potential of Wharton’s Jelly Mesenchymal Stem/Stromal Cells. Stem Cells Int. 2019, 2019, 3548917. [Google Scholar] [CrossRef]
- Liu, R.F.; Wang, F.; Wang, Q.; Zhao, X.C.; Zhang, K.M. Research Note Mesenchymal stem cells from skin lesions of psoriasis patients promote proliferation and inhibit apoptosis of HaCaT cells. Genet. Mol. Res. 2015, 14, 17758–17767. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Liang, N.; Cao, Y.; Xing, J.; Li, J.; Li, J.; Zhao, X.; Li, J.; Niu, X.; Hou, R.; et al. The effects of human dermal-derived mesenchymal stem cells on the keratinocyte proliferation and apoptosis in psoriasis. Exp. Dermatol. 2021, 30, 943–950. [Google Scholar] [CrossRef]
- Niu, X.; Li, J.; Zhao, X.; Wang, Q.; Wang, G.; Hou, R.; Li, X.; An, P.; Yin, G.; Zhang, K. Dermal mesenchymal stem cells: A resource of migration-associated function in psoriasis? Stem Cell Res. Ther. 2019, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Yang, Y.; Yan, X.; Zhang, K. Abnormalities in cytokine secretion from mesenchymal stem cells in psoriatic skin lesions. Eur. J. Dermatol. 2013, 23, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.; Chang, W.; Peng, A.; Cao, Y.; Li, J.; Wang, Y.; Jiao, J.; Zhang, K. Dermal Mesenchymal Stem Cells from Psoriatic Lesions Stimulate HaCaT Cell Proliferation, Differentiation, and Migration via Activating the PI3K/AKT Signaling Pathway. Dermatology 2022, 238, 283–291. [Google Scholar] [CrossRef]
- Li, J.; Xing, J.; Lu, F.; Chang, W.; Liang, N.; Li, J.; Wang, Y.; Li, X.; Zhao, X.; Hou, R.; et al. Psoriatic Dermal-derived Mesenchymal Stem Cells Reduce Keratinocyte Junctions, and Increase Glycolysis. Acta Derm. Venereol. 2020, 100, adv00122. [Google Scholar] [CrossRef]
- Cao, Y.; Liang, N.N.; Chang, W.J.; Li, J.Q.; Jiao, J.J.; Hou, R.X.; Li, J.; Zhang, K.M. Role of psoriatic keratinocytes in the metabolic reprogramming of dermal mesenchymal stem cells. Int. J. Dermatol. 2022, 61, 337–345. [Google Scholar] [CrossRef]
- Peng, A.; Lu, F.; Xing, J.; Dou, Y.; Yao, Y.; Li, J.; Li, J.; Hou, R.; Zhang, K.; Yin, G. Psoriatic Dermal-Derived Mesenchymal Stem Cells Induced C3 Expression in Keratinocytes. Clin. Cosmet. Investig. Dermatol. 2022, 15, 1489–1497. [Google Scholar] [CrossRef]
- Castro-Manrreza, M.E.; Bonifaz, L.; Castro-Escamilla, O.; Monroy-García, A.; Cortés-Morales, A.; Hernández-Estévez, E.; Hernández-Cristino, J.; Mayani, H.; Montesinos, J.J. Mesenchymal Stromal Cells from the Epidermis and Dermis of Psoriasis Patients: Morphology, Immunophenotype, Differentiation Patterns, and Regulation of T Cell Proliferation. Stem Cells Int. 2019, 2019, 4541797. [Google Scholar] [CrossRef]
- Liu, R.; Wang, Y.; Zhao, X.; Yang, Y.; Zhang, K. Lymphocyte inhibition is compromised in mesenchymal stem cells from psoriatic skin. Eur. J. Dermatol. 2014, 24, 560–567. [Google Scholar] [CrossRef]
- Zhao, X.; Jiao, J.; Li, X.; Hou, R.; Li, J.; Niu, X.; Liu, R.; Yang, X.; Li, J.; Liang, J.; et al. Immunomodulatory effect of psoriasis-derived dermal mesenchymal stem cells on TH1/TH17 cells. Eur. J. Dermatol. 2021, 31, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Zhao, X.; Wang, Y.; Liang, N.; Li, J.; Yang, X.; Xing, J.; Zhou, L.; Li, J.; Hou, R.; et al. Normal mesenchymal stem cells can improve the abnormal function of T cells in psoriasis via upregulating transforming growth factor-β receptor. J. Dermatol. 2022, 49, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Hou, R.; Li, J.; Zhao, X.; Wang, Q.; Zhang, K.; Li, X. Psoriatic Serum Induce an Abnormal Inflammatory Phenotype and a Decreased Immunosuppressive Function of Mesenchymal Stem Cells. Int. J. Stem Cells 2022, 15, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.W.; Voyles, S.V.; Armstrong, E.J.; Fuller, E.N.; Rutledge, J.C. Angiogenesis and oxidative stress: Common mechanisms linking psoriasis with atherosclerosis. J. Dermatol. Sci. 2011, 63, 1–9. [Google Scholar] [CrossRef]
- Man, X.Y.; Yang, X.H.; Cai, S.Q.; Bu, Z.Y.; Zheng, M. Overexpression of vascular endothelial growth factor (VEGF) receptors on keratinocytes in psoriasis: Regulated by calcium independent of VEGF. J. Cell. Mol. Med. 2008, 12, 649–660. [Google Scholar] [CrossRef]
- Nofal, A.; Al-Makhzangy, I.; Attwa, E.; Nassar, A.; Abdalmoati, A. Vascular endothelial growth factor in psoriasis: An indicator of disease severity and control. J. Eur. Acad. Dermatol. Venereol. 2009, 23, 803–806. [Google Scholar] [CrossRef]
- Hou, R.; Yan, H.; Niu, X.; Chang, W.; An, P.; Wang, C.; Yang, Y.; Yan, X.; Li, J.; Liu, R.; et al. Gene expression profile of dermal mesenchymal stem cells from patients with psoriasis. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 1782–1791. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, J.; Liang, J.; Hou, H.; Li, J.; Li, J.; Cao, Y.; Li, J.; Zhang, K. Psoriatic mesenchymal stem cells stimulate the angiogenesis of human umbilical vein endothelial cells in vitro. Microvasc. Res. 2021, 136, 104151. [Google Scholar] [CrossRef]
- Campanati, A.; Caffarini, M.; Diotallevi, F.; Radi, G.; Lucarini, G.; Di Vincenzo, M.; Orciani, M.; Offidani, A. The efficacy of in vivo administration of Apremilast on mesenchymal stem cells derived from psoriatic patients. Inflamm. Res. 2021, 70, 79–87. [Google Scholar] [CrossRef]
- Hou, R.; Li, J.; Niu, X.; Liu, R.; Chang, W.; Zhao, X.; Wang, Q.; Li, X.; Yin, G.; Zhang, K. Stem cells in psoriasis. J. Dermatol. Sci. 2017, 86, 181–186. [Google Scholar] [CrossRef]
- Hou, R.X.; Liu, R.F.; Zhao, X.C.; Jia, Y.R.; An, P.; Hao, Z.P.; Li, J.Q.; Li, X.H.; Yin, G.H.; Zhang, K.M. Increased miR-155-5p expression in dermal mesenchymal stem cells of psoriatic patients: Comparing the microRNA expression profile by microarray. Genet. Mol. Res. 2016, 15, gmr.15038631. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hou, R.; Niu, X.; Liu, R.; Wang, Q.; Wang, C.; Li, X.; Hao, Z.; Yin, G.; Zhang, K. Comparison of microarray and RNA-Seq analysis of mRNA expression in dermal mesenchymal stem cells. Biotechnol. Lett. 2016, 38, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, X.; Li, J.; Zhou, L.; Chang, W.; Li, J.; Hou, R.; Li, J.; Yin, G.; Li, X.; et al. MiR-155 inhibits TP53INP1 expression leading to enhanced glycolysis of psoriatic mesenchymal stem cells. J. Dermatol. Sci. 2022, 105, 142–151. [Google Scholar] [CrossRef]
- Zhao, X.; Xing, J.; Li, J.; Hou, R.; Niu, X.; Liu, R.; Jiao, J.; Yang, X.; Li, J.; Liang, J.; et al. Dysregulated Dermal Mesenchymal Stem Cell Proliferation and Differentiation Interfered by Glucose Metabolism in Psoriasis. Int. J. Stem Cells 2021, 14, 85–93. [Google Scholar] [CrossRef]
- Orciani, M.; Campanati, A.; Salvolini, E.; Lucarini, G.; Di Benedetto, G.; Offidani, A.; Di Primio, R. The mesenchymal stem cell profile in psoriasis. Br. J. Dermatol. 2011, 165, 585–592. [Google Scholar] [CrossRef]
- Sah, S.K.; Park, K.H.; Yun, C.O.; Kang, K.S.; Kim, T.Y. Effects of Human Mesenchymal Stem Cells Transduced with Superoxide Dismutase on Imiquimod-Induced Psoriasis-Like Skin Inflammation in Mice. Antioxid. Redox Signal. 2016, 24, 233–248. [Google Scholar] [CrossRef]
- Ribeiro, A.; Laranjeira, P.; Mendes, S.; Velada, I.; Leite, C.; Andrade, P.; Santos, F.; Henriques, A.; Grãos, M.; Cardoso, C.M.; et al. Mesenchymal stem cells from umbilical cord matrix, adipose tissue and bone marrow exhibit different capability to suppress peripheral blood B, natural killer and T cells. Stem Cell Res. Ther. 2013, 4, 125. [Google Scholar] [CrossRef]
- English, K. Mechanisms of mesenchymal stromal cell immunomodulation. Immunol. Cell Biol. 2013, 91, 19–26. [Google Scholar] [CrossRef]
- Rokunohe, A.; Matsuzaki, Y.; Rokunohe, D.; Sakuraba, Y.; Fukui, T.; Nakano, H.; Sawamura, D. Immunosuppressive effect of adipose-derived stromal cells on imiquimod-induced psoriasis in mice. J. Dermatol. Sci. 2016, 82, 50–53. [Google Scholar] [CrossRef]
- Shi, F.; Guo, L.C.; Zhu, W.D.; Cai, M.H.; Chen, L.L.; Wu, L.; Chen, X.J.; Zhu, H.Y.; Wu, J. Human adipose tissue-derived MSCs improve psoriasis-like skin inflammation in mice by negatively regulating ROS. J. Dermatol. Treat. 2022, 33, 2129–2136. [Google Scholar] [CrossRef]
- De Jesus, M.M.; Santiago, J.S.; Trinidad, C.V.; See, M.E.; Semon, K.R.; Fernandez, M.O., Jr.; Chung, F.S. Autologous Adipose-Derived Mesenchymal Stromal Cells for the Treatment of Psoriasis Vulgaris and Psoriatic Arthritis: A Case Report. Cell Transplant. 2016, 25, 2063–2069. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Ye, S.; He, Z.; Huang, Y.; Deng, J.; Wen, Z.; Chen, X.; Li, H.; Han, Q.; Deng, H.; et al. Adipose-derived mesenchymal stem cells (AD-MSCs) in the treatment for psoriasis: Results of a single-arm pilot trial. Ann. Transl. Med. 2021, 9, 1653. [Google Scholar] [CrossRef] [PubMed]
- Attia, S.S.; Rafla, M.; El-Nefiawy, N.E.; Abdel Hamid, H.F.; Amin, M.A.; Fetouh, M.A. A potential role of mesenchymal stem cells derived from human umbilical cord blood in ameliorating psoriasis-like skin lesion in the rats. Folia Morphol. 2022, 81, 614–631. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Sah, S.K.; Lee, J.H.; Seo, K.W.; Kang, K.S.; Kim, T.Y. Human umbilical cord blood-derived mesenchymal stem cells ameliorate psoriasis-like skin inflammation in mice. Biochem. Biophys. Rep. 2017, 9, 281–288. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, H.; Jiang, C.; Chen, C.; Shen, D.; Xie, F.; Zhang, H.; Yang, J.; Wang, H. Effects of different concentrations of human umbilical cord mesenchymal stem cells to ameliorate psoriasis-like skin lesions in BALB/c mice. Ann. Transl. Med. 2022, 10, 86. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, Y.; Zhou, X.; Zhao, Z.; Yu, Q.; Chen, Z.; Wang, Y.; Xu, P.; Yu, Z.; Guo, C.; et al. Human umbilical cord-derived mesenchymal stem cells ameliorate psoriasis-like dermatitis by suppressing IL-17-producing γδ T cells. Cell Tissue Res. 2022, 388, 549–563. [Google Scholar] [CrossRef]
- Chen, H.; Niu, J.W.; Ning, H.M.; Pan, X.; Li, X.B.; Li, Y.; Wang, D.H.; Hu, L.D.; Sheng, H.X.; Xu, M.; et al. Treatment of Psoriasis with Mesenchymal Stem Cells. Am. J. Med. 2016, 129, e13–e14. [Google Scholar] [CrossRef]
- Ahn, H.; Lee, S.Y.; Jung, W.J.; Pi, J.; Lee, K.H. Psoriasis treatment using minimally manipulated umbilical cord-derived mesenchymal stem cells: A case report. World J. Clin. Cases 2021, 9, 6798–6803. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, S.; Peng, C.; Zou, X.; Yang, C.; Mei, H.; Li, C.; Su, X.; Xiao, N.; Ouyang, Q.; et al. Human umbilical cord mesenchymal stem cells for psoriasis: A phase 1/2a, single-arm study. Signal Transduct. Target. Ther. 2022, 7, 263. [Google Scholar] [CrossRef]
- Kim, C.H.; Lim, C.Y.; Lee, J.H.; Kim, K.C.; Ahn, J.Y.; Lee, E.J. Human Embryonic Stem Cells-Derived Mesenchymal Stem Cells Reduce the Symptom of Psoriasis in Imiquimod-Induced Skin Model. Tissue Eng. Regen. Med. 2019, 16, 93–102. [Google Scholar] [CrossRef]
- Ye, Z.; Liang, Y.; Lin, B.; Li, Y.; Chai, X.; Lian, J.; Zhang, X.; Che, Z.; Zeng, J. Gingiva-Derived Mesenchymal Stem Cells Attenuate Imiquimod- (IMQ-) Induced Murine Psoriasis-Like Skin Inflammation. Stem Cells Int. 2022, 2022, 6544514. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.G.; Hsu, N.C.; Wang, S.M.; Wang, F.N. Successful Treatment of Plaque Psoriasis with Allogeneic Gingival Mesenchymal Stem Cells: A Case Study. Case Rep. Dermatol. Med. 2020, 2020, 4617520. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Yamahara, K.; Hamada, A.; Fujimori, Y.; Yamanishi, K. Human amnion-derived mesenchymal stem cells ameliorate imiquimod-induced psoriasiform dermatitis in mice. J. Dermatol. 2019, 46, 276–278. [Google Scholar] [CrossRef]
- Yang, M.; Wang, L.; Chen, Z.; Hao, W.; You, Q.; Lin, J.; Tang, J.; Zhao, X.; Gao, W.Q.; Xu, H. Topical administration of the secretome derived from human amniotic epithelial cells ameliorates psoriasis-like skin lesions in mice. Stem Cell Res. Ther. 2022, 13, 393. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, M.; Kim, Y.H.; Ryu, K.H.; Lee, K.H.; Cho, K.A.; Woo, S.Y. Tonsil-derived mesenchymal stem cells (T-MSCs) prevent Th17-mediated autoimmune response via regulation of the programmed death-1/programmed death ligand-1 (PD-1/PD-L1) pathway. J. Tissue Eng. Regen. Med. 2018, 12, e1022–e1033. [Google Scholar] [CrossRef]
- Francisco, L.M.; Sage, P.T.; Sharpe, A.H. The PD-1 pathway in tolerance and autoimmunity. Immunol. Rev. 2010, 236, 219–242. [Google Scholar] [CrossRef]
- Kim, J.; Krueger, J.G. The immunopathogenesis of psoriasis. Dermatol. Clin. 2015, 33, 13–23. [Google Scholar] [CrossRef]
- Nauta, A.J.; Kruisselbrink, A.B.; Lurvink, E.; Willemze, R.; Fibbe, W.E. Mesenchymal stem cells inhibit generation and function of both CD34+-derived and monocyte-derived dendritic cells. J. Immunol. 2006, 177, 2080–2087. [Google Scholar] [CrossRef]
- Zhang, W.; Ge, W.; Li, C.; You, S.; Liao, L.; Han, Q.; Deng, W.; Zhao, R.C. Effects of mesenchymal stem cells on differentiation, maturation, and function of human monocyte-derived dendritic cells. Stem Cells Dev. 2004, 13, 263–271. [Google Scholar] [CrossRef]
- Jiang, X.X.; Zhang, Y.; Liu, B.; Zhang, S.X.; Wu, Y.; Yu, X.D.; Mao, N. Human mesenchymal stem cells inhibit differentiation and function of monocyte-derived dendritic cells. Blood 2005, 105, 4120–4126. [Google Scholar] [CrossRef]
- Rasmusson, I.; Ringdén, O.; Sundberg, B.; Le Blanc, K. Mesenchymal stem cells inhibit lymphocyte proliferation by mitogens and alloantigens by different mechanisms. Exp. Cell Res. 2005, 305, 33–41. [Google Scholar] [CrossRef]
- Ghannam, S.; Pène, J.; Moquet-Torcy, G.; Jorgensen, C.; Yssel, H. Mesenchymal stem cells inhibit human Th17 cell differentiation and function and induce a T regulatory cell phenotype. J. Immunol. 2010, 185, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Chen, X.; Shao, H.; Bai, L.; Li, X.; Zhang, X. Mesenchymal Stem Cells Inhibited Dendritic Cells Via the Regulation of STAT1 and STAT6 Phosphorylation in Experimental Autoimmune Uveitis. Curr. Mol. Med. 2018, 17, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, R.; Fazekasova, H.; Lam, E.W.; Soeiro, I.; Lombardi, G.; Dazzi, F. Mesenchymal stem cells inhibit dendritic cell differentiation and function by preventing entry into the cell cycle. Transplantation 2007, 83, 71–76. [Google Scholar] [CrossRef]
- Li, Y.P.; Paczesny, S.; Lauret, E.; Poirault, S.; Bordigoni, P.; Mekhloufi, F.; Hequet, O.; Bertrand, Y.; Ou-Yang, J.P.; Stoltz, J.F.; et al. Human mesenchymal stem cells license adult CD34+ hemopoietic progenitor cells to differentiate into regulatory dendritic cells through activation of the Notch pathway. J. Immunol. 2008, 180, 1598–1608. [Google Scholar] [CrossRef]
- Chen, H.W.; Chen, H.Y.; Wang, L.T.; Wang, F.H.; Fang, L.W.; Lai, H.Y.; Chen, H.H.; Lu, J.; Hung, M.S.; Cheng, Y.; et al. Mesenchymal stem cells tune the development of monocyte-derived dendritic cells toward a myeloid-derived suppressive phenotype through growth-regulated oncogene chemokines. J. Immunol. 2013, 190, 5065–5077. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, Y.; Ye, L.; Zhang, T.; Cheng, J.; Chen, G.; Zhang, Q.; Yang, Y. Umbilical Cord-derived Mesenchymal Stem Cells Instruct Monocytes Towards an IL10-producing Phenotype by Secreting IL6 and HGF. Sci. Rep. 2016, 6, 37566. [Google Scholar] [CrossRef]
- Liu, X.; Ren, S.; Ge, C.; Cheng, K.; Zenke, M.; Keating, A.; Zhao, R.C. Sca-1+Lin-CD117- mesenchymal stem/stromal cells induce the generation of novel IRF8-controlled regulatory dendritic cells through Notch-RBP-J signaling. J. Immunol. 2015, 194, 4298–4308. [Google Scholar] [CrossRef]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef]
- Darlan, D.M.; Raga, A.; Muhar, A.M.; Putra, A.; Alif, I. Mesenchymal Stem Cells Suppress Dendritic Cells and Modulate Proinflammatory Milieu Through Interleukin-10 Expression in Peripheral Blood Mononuclear Cells of Human Systemic Lupus Erythematosus. Acta Inform. Med. 2023, 31, 20–25. [Google Scholar] [CrossRef]
- Antoon, R.; Overdevest, N.; Saleh, A.H.; Keating, A. Mesenchymal stromal cells as cancer promoters. Oncogene 2024, 43, 3545–3555. [Google Scholar] [CrossRef] [PubMed]
- Khakoo, A.Y.; Pati, S.; Anderson, S.A.; Reid, W.; Elshal, M.F.; Rovira, I.I.; Nguyen, A.T.; Malide, D.; Combs, C.A.; Hall, G.; et al. Human mesenchymal stem cells exert potent antitumorigenic effects in a model of Kaposi’s sarcoma. J. Exp. Med. 2006, 203, 1235–1247. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Xu, Z.; Zhao, T.; Zhao, Z.; Shi, M.; Zhao, R.C.; Ye, L.; Zhang, X. Suppression of tumorigenesis by human mesenchymal stem cells in a hepatoma model. Cell Res. 2008, 18, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Xu, Z.L.; Zhao, T.J.; Ye, L.H.; Zhang, X.D. Dkk-1 secreted by mesenchymal stem cells inhibits growth of breast cancer cells via depression of Wnt signalling. Cancer Lett. 2008, 269, 67–77. [Google Scholar] [CrossRef]
- Otsu, K.; Das, S.; Houser, S.D.; Quadri, S.K.; Bhattacharya, S.; Bhattacharya, J. Concentration-dependent inhibition of angiogenesis by mesenchymal stem cells. Blood 2009, 113, 4197–4205. [Google Scholar] [CrossRef]
- Gao, P.; Ding, Q.; Wu, Z.; Jiang, H.; Fang, Z. Therapeutic potential of human mesenchymal stem cells producing IL-12 in a mouse xenograft model of renal cell carcinoma. Cancer Lett. 2010, 290, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Guan, Y.; Liu, H.; Hao, N.; Liu, T.; Meng, X.; Fu, C.; Li, Y.; Qu, Q.; Zhang, Y.; et al. Silica nanorattle-doxorubicin-anchored mesenchymal stem cells for tumor-tropic therapy. ACS Nano 2011, 5, 7462–7470. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Type of Cell | Animal Model (Sex, Age) | Disease Model | Injection Method | Treatment | Effectiveness | Follow-Up | Reference |
|---|---|---|---|---|---|---|---|
| AD-MSCs | —— | IMQ-induced psoriasis-like inflammation | intravenous injection | On days 0, 2, and 4, either 1.0 × 106 cells or PBS were injected into IMQ-treated skin | Inhibits the production of Th17-related cytokines (such as IL-17A and TNFα) and minimizes IMQ-induced skin changes in psoriasis | 6 days | [78] |
| hAD-MSCs | C57BL/6 mice (male, 8–10 weeks) | IMQ-induced psoriasis-like inflammation (every day for 7 days) | subcutaneous injection | On days 1 and 4, 1.0 × 106 hAD-MSCs were injected. Control mice received an equal volume of PBS. | hAD-MSCs might treat inflammatory disease by suppressing ROS production | 8 days | [79] |
| UC-MSCs | C57BL/6 mice (male, 6 weeks) | IMQ-induced psoriasis-like inflammation | subcutaneous injection | On days 0 and 6, the control group and the model group received a 200 μL injection of normal saline, and the hUC-MSCs group received an injection of 2.0 × 106 cells. | hUC MSCs may play an immunomodulatory role by regulating IL-6 and alleviating the symptoms of psoriasis | 12 days | [85] |
| hUCB-MSCs | adult albino rats (male, —) | IMQ-induced psoriasis-like inflammation (for 12 consecutive days) | subcutaneous injection | On day 6, the rats received an MSC injection (2.0 × 106 cells per injection in 2 mL of media) at four corners around the edge of the inflamed area of the skin. | The erythema, scale, and thickening of psoriasis were significantly reduced | 12 days | [83] |
| hUCB-MSCs | C57BL/6 mice (female, 8–10 weeks) | IMQ-induced psoriasis-like inflammation (every day for 6 days) | intravenous injection | 1.5 × 106 cells were injected through the tail vein on the first and fourth days | It alleviated psoriasis-like skin inflammation in the mouse model, which may be related to the inhibition of Th17 cells and TNFα production | 7 days | [84] |
| hUCB-MSCs | C57BL/6 mice (male, 8–12 weeks) | IL-23-mediated psoriasis-like skin inflammation in mice; IMQ-induced psoriasis-like inflammation | subcutaneous injection | IL-23 mediated model: 2.0 × 106 cells were injected on days 1 and 7 after induction of psoriasis-like skin. IMQ-induced model: mice received an injection of 2.0 × 106 cells every day for 5 days. Control group: an equal volume of phosphate-buffered saline was given at the same time points | Treatment inhibited pro-inflammatory cytokines (IL-6, IL-17, and TNFα) and the expression of chemokines (CCL17, CCL20, and CCL27) in mouse skin, providing prevention and treatment of psoriasis-like skin inflammation in mice | 15 days | [87] |
| hUCB-MSCs | BALB/C mice (male, 6–8 weeks) | IMQ-induced psoriasis-like inflammation (for consecutive 7 days), plus subcutaneous injection of 5 μL of rmIL-12 and LPS once at the beginning of model creation | intravenous injection | The intervention group was divided into four intervention conditions: UC stem cells 1.0 × 107/kg, 2 × 107/kg, and 4 × 107/kg and fresh UC stem cells 2 × 107/kg | It had an obvious effect on the treatment of psoriasis in mice, and the high concentration of hUC-MSCs had the best therapeutic effect. | 7 days | [88] |
| hUCB-MSCs | BALB/C mice (female, 6–8 weeks) | IMQ-induced psoriasis-like inflammation (for 6 consecutive days) | intravenous injection | IMQ group: 100 μL sterile PBS solution, IMQ+MSC group: 1.0 × 106/cells MSC suspension in the same volume | MSCs could alleviate the severity of psoriasis-like dermatitis in mice by inhibiting TYK2 phosphorylation | 8 days | [86] |
| hUCB-MSCs | C57BL/6 mice (—, 6–8 weeks) | IMQ-induced psoriasis-like inflammation (for 6 consecutive days) | subcutaneous injection | Subcutaneous injection with hUC-MSCs (2 × 106/cells) in 200 μL PBS. Control mice received an equal volume of PBS | hUC-MSCs suppressed psoriasis-like skin inflammation by suppressing γδ T cells, specifically IL-17-producing γδ T cells | 6 days | [89] |
| hE-MSCs | C57BL/6 mice (female, 8 weeks) | IMQ-induced psoriasis-like inflammation (for 6 consecutive days) | subcutaneous injection | On the second and fourth days after the start of local application of IMQ, 2.5 × 106/cells of MSCs in 200 uL PBS were injected into mice | Inhibition of Th1- and Th17-related cytokines and IMQ-induced psoriasis skin changes | 7 days | [90] |
| GMSCs | C57BL/6 mice (female, 8 weeks) | IMQ-induced psoriasis-like inflammation (for 7 consecutive days) | intravenous injection | 2 × 106 2D-GMSCs or 3D-GMSCs in 200 μL PBS via the mouse tail vein on day 1 and day 4, with consecutive IMQ treatment. The IMQ control group received an equal volume of PBS at the same time points | The proportion of Treg cells increased, while the proportion of Th17 cells decreased, indicating that GMSCs play an immunomodulatory and anti-inflammatory role | 8 days | [92] |
| hAMSC | C57BL/6 mice (—) | IMQ-induced psoriasis-like inflammation (for 5 consecutive days) | intravenous injection | On days 0 and 3, the MSC treatment group was injected with hAMSCs (2 × 105 cells) suspended in 100 μL mouse serum. The control group was injected with 100 μL mouse serum. | hAMSCs decreased both IL-17A and IL-22 production by cutaneous γδ-low T cells in IMQ-treated skin | 5 days | [93] |
| T-MSCs | C57BL/6 mice (female, 8 weeks) | IMQ-induced psoriasis-like inflammation (for 6 consecutive days) | intravenous injection | 1 × 106 T-MSCs were injected into the mouse tail vein on days 1 and 3 of the IMQ application period. Control mice received an equal volume of PBS at the same time points | T-MSCs could prevent autoimmune disease (psoriasis) by enhancing the PD-1/PD-L1 pathway | 7 days | [95] |
| Type of Cell | Clinical Study | Number of Male/Female Patients | Age (Years) | Treatment | Safety (Treatment-Related Adverse Event) | Inclusion Criteria | Effectiveness | Follow-Up | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Autologous AD-MSCs | Case report | 2 (1/1) | 58, 28 | Two to three intravenous infusions of AD-MSCs at a dose of 0.5–3.1 × 106 cells/kg | No adverse reaction was recorded in the first year of follow-up | Psoriasis vulgaris and psoriatic arthritis | PASI scores improved significantly | 12 months | [76] |
| Allogeneic AD-MSCs | A single-center, single-arm pilot study (NCT03265613) | 7 (6/1) | 18–65 | At weeks 0, 4, and 8, 0.5 × 106 cells/kg body weight and 2–3 mL/min infusion rate were used for intravenous infusion of AD MSCs | 16 adverse events, mild (level 1, 94%) or moderate (level 2, 6%). The most common adverse event was transient fever (31.2%) | Moderate to severe psoriasis vulgaris (PASI > 10 or BSA > 10%) | The PASI score decreased gradually from baseline to week 12 | 12 weeks | [77] |
| Allogeneic UC-MSCs | Case report | 2 (1/1) | 35, 26 | Dose of 1 × 106 cells at different frequencies | None | Psoriasis vulgaris | Psoriasis was relieved without recurrence | 4–5 years | [80] |
| Allogeneic UC-MSCs | Case report | 1 (1/0) | 47 | Three rounds of treatment were given within 2 weeks. In the first round, intravenous transplantation (3 × 106 cells) and local transplantation (1 × 106 cells) were given. The second and third rounds included only local transplantation, and they were conducted once a week | None | Psoriasis vulgaris | Psoriasis was improved, and there was no side effect or recurrence during the follow-up | 5 months | [81] |
| Allogeneic UC-MSCs | A phase 1/2a, single-arm study (NCT03765957) | 12 | 18–65 | There were four groups (A, B, C, and D). Groups A and B received intravenous 1.5 × 106/kg and 2 × 106/kg. Groups C and D received intravenous 2.5 × 106/kg and 3 × 106/kg (mesenchymal stem cells at baseline and every 4 weeks, two times) | N/A | Psoriasis vulgaris | PASI improvement ≥ 75%, n = 6. One patient had improvement in PASI 90 at the 6-month follow-up and remained relapse-free for 1 year without using any other traditional drugs. | 6 months | [82] |
| Allogeneic GMSCs | Case report | 1 (1/0) | 19 | The dose for two consecutive weeks was 3 × 106/kg for infusion treatment. After five weeks, MSC infusion was conducted three times a week | None | Severe plaque psoriasis | The plaque was gradually cleared, and no adverse reaction was reported. One week after the last injection, the psoriasis lesions completely disappeared. There was no recurrence in the three-year monitoring. | 3 years | [91] |
| Allogeneic UC-MSCs | A phase 1/2a, single-arm study | 30 | 18–65 | Subjects in this arm received six UC-MSC infusions (each time 1 × 106/kg) within 8 weeks. The first time to the fourth time were given once a week for 4 successive weeks; then, the last two times were given once every 2 weeks. | N/A | Psoriasis vulgaris | N/A | 12 months | Safety and efficacy of UC-MSCs in patients with psoriasis vulgaris (NCT02491658) ClinicalTrials.gov |
| Allogeneic UC-MSCs | A randomized, positive controlled phase 1 trial | 57 | 18–60 | Low-dose: 1 × 106 cells/kg; high-dose: 3 × 106 cells/kg For the first to the fourth time, the subjects were given treatment once a week for 4 successive weeks, and the last two times were given once every 2 weeks. Active comparator: methotrexate (each time 5–25 mg orally once a week for 16 successive weeks) | N/A | Moderate and severe plaque psoriasis | N/A | 52 weeks | Safety and efficacy of UC-MSCs in patients with plaque psoriasis (NCT03424629) ClinicalTrials.gov |
| Allogeneic AD-MSCs | A phase 1/2, single-group study | 8 | 18–65 | Drug: calcipotriol ointment (twice daily for 12 weeks) AD-MSCs: 2 × 106 cells/kg | N/A | Moderate to severe psoriasis vulgaris (PASI > 10 or BSA >10%) | N/A | 12 weeks | Efficacy and safety of AD-MSCs plus calcipotriol ointment in patients with moderate to severe psoriasis (NCT03392311) ClinicalTrials.gov |
| Allogeneic AD-MSCs | A single-group study | 8 | 18–65 | Drug: calcipotriol ointment (twice daily for 12 weeks). AD-MSCs: 2 × 106 cells/kg; drug: PSORI-CM01 granule 5.5 g orally once a day for 12 weeks | N/A | Moderate to severe psoriasis (PASI > 10 or BSA >10%) | N/A | 12 weeks | Efficacy and safety of AD-MSCs plus calcipotriol ointment and PSORI-CM01 granule in psoriasis patients (NCT04275024) ClinicalTrials.gov |
| Allogeneic AD-MSCs | A randomized controlled phase 1/2 trial | 16 | 18–65 | Gu Ben Hua Yu group: Gu Ben Hua Yu formula was orally administrated once a day for 12 weeks (except the day of the infusion of AD-MSCs) + AD-MSCs: 2 × 106 cells/kg (at weeks 0, 2, 4, 6, and 8) | N/A | Psoriasis vulgaris (PASI > 7 or BSA >10%) | N/A | 12 weeks | Comparison of PSORI-CM01 formula vs. Gu Ben Hua Yu formula combined with AD-MSCs in psoriasis (NCT04785027) ClinicalTrials.gov |
| hUCB-MSCs | A phase 1, single-center, randomized, open-label study | 9 | 19–65 | Patients were treated by FURESTEM-CD Inj. Subcutaneous injection: 5.0 × 107 cells; 1.0 × 107 cells; 2.0 × 108 cells | N/A | Moderate to severe plaque-type psoriasis | N/A | 144 weeks | Safety of FURESTEM-CD inj. in patients with moderate to severe plaque-type psoriasis (NCT02918123) ClinicalTrials.gov |
| Allogeneic UC-MSCs | A phase 1/2, single-group study | 5 | 18–65 | UC-MSC intravenous injection at a dose of 2.0 × 106 cells at weeks 0, 2, 4, 6, and 8 for 12 weeks | N/A | Moderate to severe psoriasis vulgaris (PASI > 7 or BSA > 10%) | N/A | 12 weeks | Efficacy and safety of expanded UCMSCs on patients with moderate to severe psoriasis (NCT03745417) ClinicalTrials.gov |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ou, J.; Li, Z.; Yao, D.; Lu, C.; Zeng, X. Multimodal Function of Mesenchymal Stem Cells in Psoriasis Treatment. Biomolecules 2025, 15, 737. https://doi.org/10.3390/biom15050737
Ou J, Li Z, Yao D, Lu C, Zeng X. Multimodal Function of Mesenchymal Stem Cells in Psoriasis Treatment. Biomolecules. 2025; 15(5):737. https://doi.org/10.3390/biom15050737
Chicago/Turabian StyleOu, Jiaxin, Ziqing Li, Danni Yao, Chuanjian Lu, and Xiang Zeng. 2025. "Multimodal Function of Mesenchymal Stem Cells in Psoriasis Treatment" Biomolecules 15, no. 5: 737. https://doi.org/10.3390/biom15050737
APA StyleOu, J., Li, Z., Yao, D., Lu, C., & Zeng, X. (2025). Multimodal Function of Mesenchymal Stem Cells in Psoriasis Treatment. Biomolecules, 15(5), 737. https://doi.org/10.3390/biom15050737