

Baicalin Alleviates Piglet Immunosuppression Induced by Glaesserella parasuis via Promoting CD163/Tumor Necrosis Factor-like Weak Inducer of Apoptosis-Mediated Autophagy
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacterial
2.3. Drugs
2.4. Animal Experiment Design 1
2.5. Histopathological Analysis
2.6. Flow Cytometry
2.7. Proteome Analysis
2.8. Animal Experiment Design 2
2.9. RT-PCR(Reverse Transcription-Polymerase Chain Reaction)
2.10. Western Blot
2.11. Detection of CD163 Expression by Immunohistochemistry
2.12. Statistical Analysis
3. Results
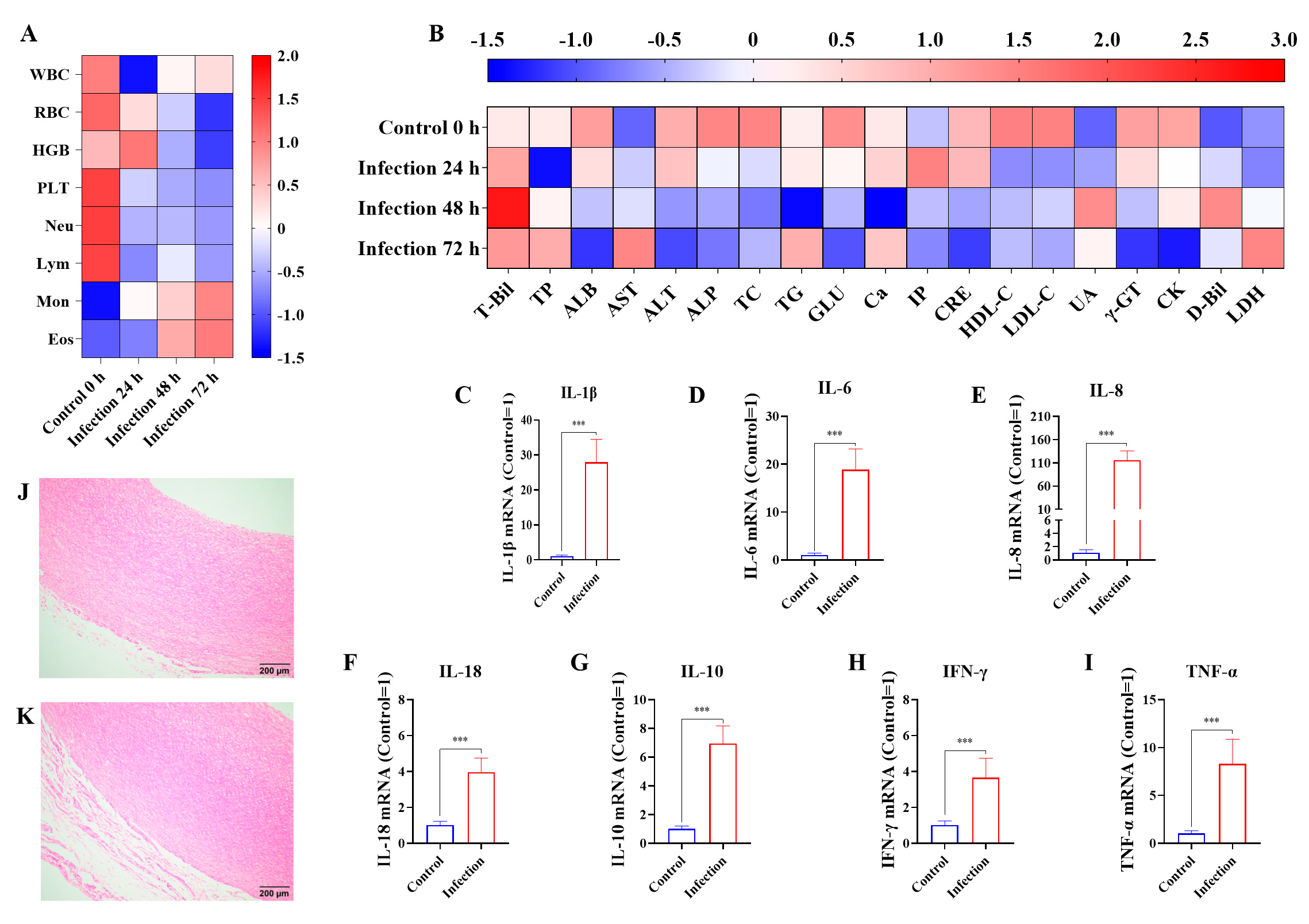
3.1. G. parasuis Altered the Routine Blood Indicators and Biochemical Parameters, Increased Cytokines Production, and Induced Blood Vessel Tissue Damage
3.2. G. parasuis Reduced the Proportion of CD3+, CD3+CD4+, and CD3+CD8+ T Cells in the Blood of Piglets
3.3. G. parasuis Infection Dysregulated Proteins in the Blood Vessels of Piglets
3.4. Baicalin Regulated CD163/TWEAK Axis Expression in Blood Vessels from Piglets Infected by G. parasuis
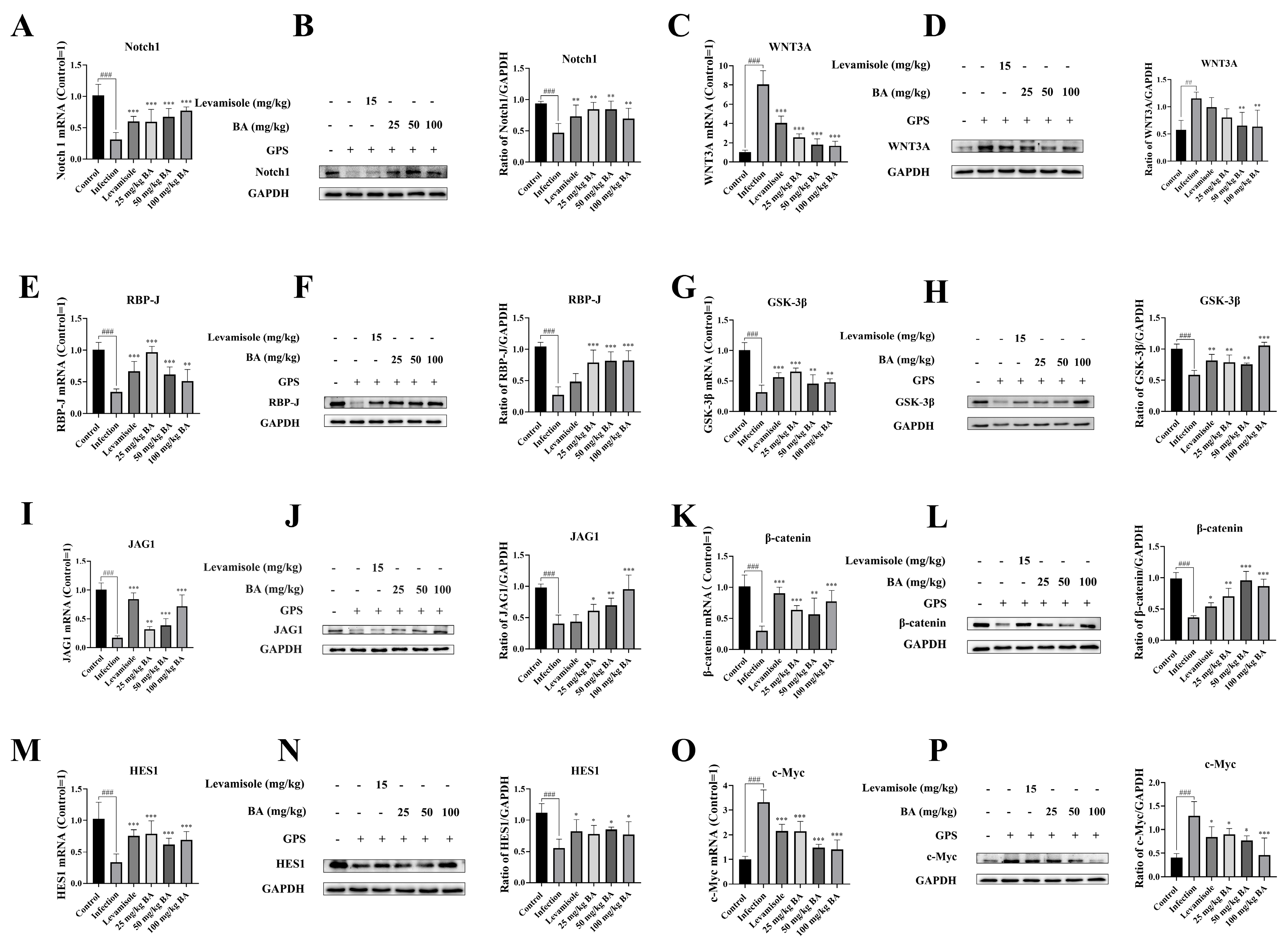
3.5. Baicalin Inhibited Notch/Wnt Signaling Pathways Activation in the Blood Vessels from Piglets Challenged by G. parasuis
3.6. Baicalin Promoted Autophagy in the Blood Vessels of G. parasuis-Infected Piglets
3.7. Baicalin Inhibited NLRP3/Caspase 1 Signaling Pathway Activation in the Blood Vessels of G. parasuis-Infected Piglets
3.8. Baicalin Decreased Cytokine Production and Diminished Pathological Tissue Damage in Blood Vessels of G. parasuis-Infected Piglets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gong, X.; Cui, Q.; Zhang, W.; Shi, Y.; Zhang, P.; Zhang, C.; Hu, G.; Sahin, O.; Wang, L.; Shen, Z.; et al. Genomic insight into the diversity of Glaesserella parasuis isolates from 19 countries. mSphere 2024, 9, e0023124. [Google Scholar] [CrossRef]
- González-Fernández, A.; Mencía-Ares, O.; García-Iglesias, M.J.; Petrocchi-Rilo, M.; Miguélez-Pérez, R.; Perelló-Jiménez, A.; Herencia-Lagunar, E.; Acebes-Fernández, V.; Gutiérrez-Martín, C.B.; Martínez-Martínez, S. TbpB-based oral mucosal vaccine provides heterologous protection against Glässer’s disease caused by different serovars of Spanish field isolates of Glaesserella parasuis. Porcine Health Manag. 2024, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yu, B.; Su, F.; Ye, S.; Xu, L.; Yuan, X.; Wu, S.; Zhang, H.; Li, J. Ribosomal protein L32 contributes to the growth, antibiotic resistance and virulence of Glaesserella parasuis. Front. Vet. Sci. 2024, 11, 1361023. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, Q.; Wei, H.; Wen, X.; Cao, S.; Huang, X.; Wu, R.; Yan, Q.; Huang, Y.; Wen, Y. Prevalence and seroepidemiology of Haemophilus parasuis in Sichuan province, China. PeerJ 2017, 5, e3379. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Jia, Y.C.; Zhang, X.L.; Zhou, Y.Y.; Guo, Y.; Yin, R.L.; Yuan, J.; Wang, L.X.; Guo, Z.B.; Wang, J.Y.; et al. Virulence assessment of four Glaesserella parasuis strains isolated in Liaoning province of China. Res. Vet. Sci. 2023, 158, 226–234. [Google Scholar] [CrossRef]
- Siwan, E.; Wong, J.; Brooks, B.A.; Shinko, D.; Baker, C.J.; Deshpande, N.; McLennan, S.V.; Twigg, S.M.; Min, D. Deep Immune and RNA Profiling Revealed Distinct Circulating CD163+ Monocytes in Diabetes-Related Complications. Int. J. Mol. Sci. 2024, 25, 10094. [Google Scholar] [CrossRef]
- Schleh, M.W.; Ameka, M.; Rodriguez, A.; Hasty, A.H. Deficiency of the hemoglobin-haptoglobin receptor, CD163, worsens insulin sensitivity in obese male mice. Diabetes 2024, 73, 1990–2002. [Google Scholar] [CrossRef]
- Gobejishvili, L.; Vatsalya, V.; Avila, D.V.; Feygin, Y.B.; McClain, C.J.; Mokshagundam, S.; Barve, S. Association of Circulating Markers of Microbial Translocation and Hepatic Inflammation with Liver Injury in Patients with Type 2 Diabetes. Biomedicines 2024, 12, 1227. [Google Scholar] [CrossRef]
- Zhu, Y.J.; Chen, H.L.; Huang, J.K.; Cai, X.J.; Zhan, B.L. TWEAK increases angiogenesis to promote diabetic skin wound healing by regulating Fn14/EGFR signaling. J. Cosmet. Dermatol. 2024, 23, 4230–4238. [Google Scholar] [CrossRef]
- Akahori, H.; Karmali, V.; Polavarapu, R.; Lyle, A.N.; Weiss, D.; Shin, E.; Husain, A.; Naqvi, N.; Van Dam, R.; Habib, A.; et al. CD163 interacts with TWEAK to regulate tissue regeneration after ischaemic injury. Nat. Commun. 2015, 6, 7792. [Google Scholar] [CrossRef]
- Kowal-Bielecka, O.; Bielecki, M.; Guiducci, S.; Trzcinska-Butkiewicz, B.; Michalska-Jakubus, M.; Matucci-Cerinic, M.; Brzosko, M.; Krasowska, D.; Chyczewski, L.; Kowal, K. High serum sCD163/sTWEAK ratio is associated with lower risk of digital ulcers but more severe skin disease in patients with systemic sclerosis. Arthritis Res. Ther. 2013, 15, R69. [Google Scholar] [CrossRef]
- Gorvel, L.; Olive, D. Tumor associated macrophage in HPV(+) tumors: Between immunosuppression and inflammation. Semin. Immunol. 2023, 65, 101671. [Google Scholar] [PubMed]
- Campbell, S.; Michaelson, J.; Burkly, L.; Putterman, C. The role of TWEAK/Fn14 in the pathogenesis of inflammation and systemic autoimmunity. Front. Biosci. 2004, 9, 2273–2284. [Google Scholar] [CrossRef]
- Winkles, J.A. The TWEAK-Fn14 cytokine-receptor axis: Discovery, biology and therapeutic targeting. Nat. Rev. Drug Discov. 2008, 7, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, L.; Dave, M.; Menghini, P.; Xin, W.; Arseneau, K.O.; Pizarro, T.T.; Cominelli, F. Protective Role for TWEAK/Fn14 in Regulating Acute Intestinal Inflammation and Colitis-Associated Tumorigenesis. Cancer Res. 2016, 76, 6533–6542. [Google Scholar] [CrossRef]
- Shan, C.; Wang, Y.; Wang, Y. The Crosstalk between Autophagy and Nrf2 Signaling in Cancer: From Biology to Clinical Applications. Int. J. Biol. Sci. 2024, 20, 6181–6206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lindstrom, A.; Kim, E.J.; Hwang, C.I.; Hall, M.L.; Lin, T.Y.; Li, Y. SEMA3C Supports Pancreatic Cancer Progression by Regulating the Autophagy Process and Tumor Immune Microenvironment. Front. Oncol. 2022, 12, 890154. [Google Scholar]
- Carr, D.; Lau, R.; Hnatykiw, A.D.; Ward, G.C.D.; Daneshmand, M.; Cabrita, M.A.; Pratt, M.A.C. cIAP2 Is an Independent Signaling and Survival Factor during Mammary Lactational Involution and Tumorigenesis. J. Mammary Gland. Biol. Neoplasia 2018, 23, 109–123. [Google Scholar] [CrossRef]
- Zhang, Q.; Guo, S.; Wang, H. The Protective Role of Baicalin in the Regulation of NLRP3 Inflammasome in Different Diseases. Cell Biochem. Biophys. 2024. [Google Scholar] [CrossRef]
- Wen, Y.; Wang, Y.; Zhao, C.; Zhao, B.; Wang, J. The Pharmacological Efficacy of Baicalin in Inflammatory Diseases. Int. J. Mol. Sci. 2023, 24, 9317. [Google Scholar]
- Song, D.; Wei, W.; Zhang, J.; Zhang, L.; Huo, J.; Wang, W. The mechanism of baicalin in improving pulmonary inflammatory response and injury and regulating intestinal flora in Mycoplasma pneumoniae pneumonia mice. Cell Signal 2025, 126, 111530. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yu, M.; Xu, M.; Ji, X.; Zong, X.; Zhang, Z.; Shang, W.; Zhang, L.; Fang, P. Baicalin suppresses macrophage JNK-mediated adipose tissue inflammation to mitigate insulin resistance in obesity. J. Ethnopharmacol. 2024, 332, 118355. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Liu, Z.; Wang, Y.; Li, X.; Yu, X.; Li, Y.; Yu, Z.; Qiu, Y.; Mei, Z.; Xu, L. Baicalin inhibits monosodium urate crystal-induced pyroptosis in renal tubular epithelial cell line through Panx-1/P2 × 7 pathways: Molecular docking, molecular dynamics, and in vitro experiments. Chem. Biol. Drug Des. 2024, 103, e14522. [Google Scholar] [CrossRef]
- Yang, X.; Li, J.; Shan, C.; Song, X.; Yang, J.; Xu, H.; Ou, D. Baicalin reduced injury of and autophagy-related gene expression in RAW264.7 cells infected with H6N6 avian influenza virus. Heliyon 2024, 10, e32645. [Google Scholar] [CrossRef]
- Dong, X.; Liu, X.; Lin, D.; Zhang, L.; Wu, Y.; Chang, Y.; Jin, M.; Huang, G. Baicalin induces cell death of non-small cell lung cancer cells via MCOLN3-mediated lysosomal dysfunction and autophagy blockage. Phytomedicine 2024, 133, 155872. [Google Scholar] [CrossRef]
- Fu, S.; Liu, S.; Li, J.; Dong, Q.; Fu, Y.; Luo, R.; Sun, Y.; Tian, X.; Liu, W.; Zong, B.; et al. Baicalin and probenecid protect against Glaesserella parasuis challenge in a piglet model. Vet. Res. 2024, 55, 96. [Google Scholar] [CrossRef]
- Fu, S.; Li, J.; You, J.; Liu, S.; Dong, Q.; Fu, Y.; Luo, R.; Sun, Y.; Tian, X.; Liu, W.; et al. Correction: Baicalin attenuates PD-1/PD-L1 axis-induced immunosuppression in piglets challenged with Glaesserella parasuis by inhibiting the PI3K/Akt/mTOR and RAS/MEK/ERK signalling pathways. Vet. Res. 2024, 55, 127. [Google Scholar] [CrossRef]
- Huang, B.; Li, F.; You, D.; Deng, L.; Xu, T.; Lai, S.; Ai, Y.; Huang, J.; Zhou, Y.; Ge, L.; et al. Porcine reproductive and respiratory syndrome virus infects the reproductive system of male piglets and impairs development of the blood-testis barrier. Virulence 2024, 15, 2384564. [Google Scholar] [CrossRef]
- Li, J.; Liu, S.; Dong, Q.; Fu, Y.; Sun, Y.; Luo, R.; Tian, X.; Guo, L.; Liu, W.; Qiu, Y.; et al. PD-1/PD-L1 axis induced host immunosuppression via PI3K/Akt/mTOR signalling pathway in piglets infected by Glaesserella parasuis. BMC Vet. Res. 2024, 20, 141. [Google Scholar] [CrossRef]
- Fu, S.; Yin, R.; Zuo, S.; Liu, J.; Zhang, Y.; Guo, L.; Qiu, Y.; Ye, C.; Liu, Y.; Wu, Z.; et al. The effects of baicalin on piglets challenged with Glaesserella parasuis. Vet. Res. 2020, 51, 102. [Google Scholar] [CrossRef]
- Hu, X.; Zheng, Y.; Fang, M.; Liang, Z.; Wen, C.; Lin, J.; Lin, Z.; Chen, S. Knockdown of the long noncoding RNA VSIG2-1:1 promotes the angiogenic ability of human pulmonary microvascular endothelial cells by activating the VEGF/PI3K/AKT pathway. Respir. Res. 2024, 25, 412. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhu, N.; Liu, J.; Wen, S.; Xu, Y.; Xu, X.; Cai, X. The role of cytolethal distending toxin in Glaesserella parasuis JS0135 strain infection: Cytotoxicity, phagocytic resistance and pathogenicity. Vet. Microbiol. 2024, 295, 110168. [Google Scholar] [CrossRef]
- Shenton, J.M.; Teranishi, M.; Abu-Asab, M.S.; Yager, J.A.; Uetrecht, J.P. Characterization of a potential animal model of an idiosyncratic drug reaction: Nevirapine-induced skin rash in the rat. Chem. Res. Toxicol. 2003, 16, 1078–1089. [Google Scholar] [CrossRef]
- Yuan, S.; Zeng, Y.; Li, J.; Wang, C.; Li, W.; He, Z.; Ye, J.; Li, F.; Chen, Y.; Lin, X.; et al. Phenotypical changes and clinical significance of CD4(+)/CD8(+) T cells in SLE. Lupus Sci. Med. 2022, 9, e000660. [Google Scholar] [CrossRef]
- Nagatani, Y.; Kiyota, N.; Imamura, Y.; Koyama, T.; Funakoshi, Y.; Komatsu, M.; Itoh, T.; Teshima, M.; Nibu, K.I.; Sakai, K.; et al. Different characteristics of the tumor immune microenvironment among subtypes of salivary gland cancer. Asia Pac. J. Clin. Oncol. 2024, 20, 779–788. [Google Scholar] [CrossRef]
- Wu, Y.; Xiao, Y.; Ding, Y.; Ran, R.; Wei, K.; Tao, S.; Mao, H.; Wang, J.; Pang, S.; Shi, J.; et al. Colorectal cancer cell-derived exosomal miRNA-372-5p induces immune escape from colorectal cancer via PTEN/AKT/NF-κB/PD-L1 pathway. Int. Immunopharmacol. 2024, 143, 113261. [Google Scholar] [CrossRef]
- Qian, J.K.; Ma, Y.; Huang, X.; Li, X.R.; Xu, Y.F.; Liu, Z.Y.; Gu, Y.; Shen, K.; Tian, L.J.; Wang, Y.T.; et al. The CD163/TWEAK/Fn14 axis: A potential therapeutic target for alleviating inflammatory bone loss. J. Orthop. Translat 2024, 49, 82–95. [Google Scholar] [CrossRef]
- Liu, M.; Ren, Y.; Zhou, Z.; Yang, J.; Shi, X.; Cai, Y.; Arreola, A.X.; Luo, W.; Fung, K.M.; Xu, C.; et al. The crosstalk between macrophages and cancer cells potentiates pancreatic cancer cachexia. Cancer Cell 2024, 42, 885–903.e884. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, W.; Qiao, S.; Zou, H.; Yu, X.J.; Yang, Y.; Li, Z.; Wang, J.; Chen, M.S.; Xu, J.; et al. Lipid droplet accumulation mediates macrophage survival and Treg recruitment via the CCL20/CCR6 axis in human hepatocellular carcinoma. Cell Mol. Immunol. 2024, 21, 1120–1130. [Google Scholar] [CrossRef] [PubMed]
- Davidsson, S.; Huotilainen, S.; Carlsson, J.; Sundqvist, P. Soluble Levels of CD163, PD-L1, and IL-10 in Renal Cell Carcinoma Patients. Diagnostics 2022, 12, 336. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, W.; Atkinson, S.D.; Kelly, C. The TWEAK/Fn14/CD163 axis-implications for metabolic disease. Rev. Endocr. Metab. Disord. 2022, 23, 449–462. [Google Scholar] [CrossRef]
- Xu, L.; Liu, W.; Huang, X.; Sun, T.; Mei, L.; Liu, M.; Ren, Z.; Wang, M.; Zheng, H.; Wang, Q.; et al. Sinomenine hydrochloride improves DSS-induced colitis in mice through inhibition of the Notch signaling pathway. BMC Gastroenterol. 2024, 24, 451. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Li, Y.; Li, D.; Dou, M.; Fu, C.; Chen, T.; Cui, X.; Zhang, Q.; Yang, P.; Hou, Y.; et al. SRCAP is involved in porcine reproductive and respiratory syndrome virus activated Notch signaling pathway. J. Virol. 2024, 98, e0121624. [Google Scholar] [CrossRef]
- Adams, J.M.; Jafar-Nejad, H. The Roles of Notch Signaling in Liver Development and Disease. Biomolecules 2019, 9, 608. [Google Scholar] [CrossRef]
- Mukherjee, M.; Fogarty, E.; Janga, M.; Surendran, K. Notch Signaling in Kidney Development, Maintenance, and Disease. Biomolecules 2019, 9, 692. [Google Scholar] [CrossRef] [PubMed]
- Zou, G.; Park, J.I. Wnt signaling in liver regeneration, disease, and cancer. Clin. Mol. Hepatol. 2023, 29, 33–50. [Google Scholar] [CrossRef]
- Akoumianakis, I.; Polkinghorne, M.; Antoniades, C. Non-canonical WNT signalling in cardiovascular disease: Mechanisms and therapeutic implications. Nat. Rev. Cardiol. 2022, 19, 783–797. [Google Scholar] [CrossRef]
- Ren, S.Y.; Xu, X. Role of autophagy in metabolic syndrome-associated heart disease. Biochim. Biophys. Acta 2015, 1852, 225–231. [Google Scholar] [CrossRef]
- Kolahdouzmohammadi, M.; Kolahdouz-Mohammadi, R.; Tabatabaei, S.A.; Franco, B.; Totonchi, M. Revisiting the Role of Autophagy in Cardiac Differentiation: A Comprehensive Review of Interplay with Other Signaling Pathways. Genes 2023, 14, 1328. [Google Scholar] [CrossRef]
- Carballo, G.B.; Ribeiro, J.H.; Lopes, G.P.F.; Ferrer, V.P.; Dezonne, R.S.; Pereira, C.M.; Spohr, T. GANT-61 Induces Autophagy and Apoptosis in Glioblastoma Cells despite their heterogeneity. Cell Mol. Neurobiol. 2021, 41, 1227–1244. [Google Scholar] [CrossRef]
- Zhang, C.; Li, W.; Wen, J.; Yang, Z. Autophagy is involved in mouse kidney development and podocyte differentiation regulated by Notch signalling. J. Cell Mol. Med. 2017, 21, 1315–1328. [Google Scholar] [CrossRef]
- Bai, R.; Miao, M.Z.; Li, H.; Wang, Y.; Hou, R.; He, K.; Wu, X.; Jin, H.; Zeng, C.; Cui, Y.; et al. Increased Wnt/β-catenin signaling contributes to autophagy inhibition resulting from a dietary magnesium deficiency in injury-induced osteoarthritis. Arthritis Res. Ther. 2022, 24, 165. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Nucleotide Sequence (5′-3′) | Tm (°C) | Length (bp) | |
|---|---|---|---|---|
| IL-1β | Forward | TCTGCATGAGCTTTGTGCAAG | 59.7 | 155 |
| Reverse | ACAGGGCAGACTCGAATTCAAC | 60.9 | ||
| IL-6 | Forward | CTTCTGGTGATGGCTACTG | 52.7 | 134 |
| Reverse | TTGCCGAGGATGTACTTAA | 50 | ||
| IL-8 | Forward | ACAGCAGTAACAACAACAAG | 50.2 | 117 |
| Reverse | GACCAGCACAGGAATGAG | 53.2 | ||
| IL-10 | Forward | CGTGGAGGAGGTGAAGAGTG | 55.4 | 178 |
| Reverse | TTAGTAGAGTCGTCATCCTGGAAG | 55.6 | ||
| IL-18 | Forward | AGTAACCATATCTGTGCAGTGT | 54 | 155 |
| Reverse | TCTTATCACCATGTCCAGGAAC | 53 | ||
| TNF-α | Forward | CGCTCTTCTGCCTACTGCACTTC | 60.7 | 164 |
| Reverse | CTGTCCCTCGGCTTTGACATT | 57.8 | ||
| Caspase1 | Forward | TACAAGAATCCCAGGCGGTG | 57.5 | 128 |
| Reverse | CCTTTGGGCTATGTCTGGGG | 58.6 | ||
| NLRP3 | Forward | CAGGCTTCTGGGACACCTTT | 59.9 | 110 |
| Reverse | GTGCAGCCCTAGTCAGAGTC | 58.8 | ||
| TWEAK | Forward | AGGCCAAGGCAGGCCAGCG | 68.1 | 100 |
| Reverse | GTCGGAGCCAGAGGCGGAGG | 66.8 | ||
| HES1 | Forward | GTGAGTGCATGAACGAGGT | 59.2 | 118 |
| Reverse | GTCATGGCGTTGATCTGGGT | 60.4 | ||
| JAG1 | Forward | TTTCAGGGCGACCTTGCATC | 61 | 121 |
| Reverse | CCACACCACACCTTCGAGC | 61 | ||
| RBP-J | Forward | CTTGAACTTACAGGACAGAAT | 53.2 | 105 |
| Reverse | GACGACACAGAGCATACT | 53.7 | ||
| GSK-3β | Forward | CAGAACCACCTCCTTTGCG | 58.8 | 100 |
| Reverse | GGTCACCTTGCTGCCATCC | 61.1 | ||
| GAPDH | Forward | GGCACAGTCAAGGCGGAGAAC | 61.9 | 105 |
| Reverse | AGCACCAGCATCACCCCATTTG | 61 | ||
| c-Myc | Forward | CTGCCAAGAGGGCTAAGTT | 54.6 | 135 |
| Reverse | TCTGGCGTTCCAAGACATT | 56 | ||
| WNT3A | Forward | TACTCCTCTGCAGCCTGAAGCA | 63.4 | 322 |
| Reverse | ATGGCGTGGACAAAGGCCGAC | 65.8 | ||
| Notch1 | Forward | GATGGCATCAATTCCTTTAC | 52.2 | 149 |
| Reverse | TGAGGGCAGGTACACTTGT | 53 | ||
| β-catenin | Forward | AAGGCAATCCTGAAGAAGA | 52 | 148 |
| Reverse | ATAGCAGCTCGTACCCTCT | 51.4 | ||
| CD163 | Forward | TGCTGTAGTCGCTGTTCT | 55.9 | 117 |
| Reverse | ACTTTCACCTCCACTCTTC | 54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, S.; Luo, R.; Li, J.; Fu, Y.; Dong, Q.; Liu, S.; Sun, Y.; Guo, L.; Hu, J.; Qiu, Y. Baicalin Alleviates Piglet Immunosuppression Induced by Glaesserella parasuis via Promoting CD163/Tumor Necrosis Factor-like Weak Inducer of Apoptosis-Mediated Autophagy. Biomolecules 2025, 15, 722. https://doi.org/10.3390/biom15050722
Fu S, Luo R, Li J, Fu Y, Dong Q, Liu S, Sun Y, Guo L, Hu J, Qiu Y. Baicalin Alleviates Piglet Immunosuppression Induced by Glaesserella parasuis via Promoting CD163/Tumor Necrosis Factor-like Weak Inducer of Apoptosis-Mediated Autophagy. Biomolecules. 2025; 15(5):722. https://doi.org/10.3390/biom15050722
Chicago/Turabian StyleFu, Shulin, Ronghui Luo, Jingyang Li, Yunjian Fu, Qiaoli Dong, Siyu Liu, Yamin Sun, Ling Guo, Jin Hu, and Yinsheng Qiu. 2025. "Baicalin Alleviates Piglet Immunosuppression Induced by Glaesserella parasuis via Promoting CD163/Tumor Necrosis Factor-like Weak Inducer of Apoptosis-Mediated Autophagy" Biomolecules 15, no. 5: 722. https://doi.org/10.3390/biom15050722
APA StyleFu, S., Luo, R., Li, J., Fu, Y., Dong, Q., Liu, S., Sun, Y., Guo, L., Hu, J., & Qiu, Y. (2025). Baicalin Alleviates Piglet Immunosuppression Induced by Glaesserella parasuis via Promoting CD163/Tumor Necrosis Factor-like Weak Inducer of Apoptosis-Mediated Autophagy. Biomolecules, 15(5), 722. https://doi.org/10.3390/biom15050722