
Past, Present and Future Perspectives of Forensic Genetics

 ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. The Past of Forensic Genetics
2.1. Blood Groups
2.2. Human Leukocyte Antigen (HLA)
2.3. Serum Proteins
2.4. DNA Fingerprinting: Variable Number of Tandem Repeats
3. The Present of Forensic Genetics
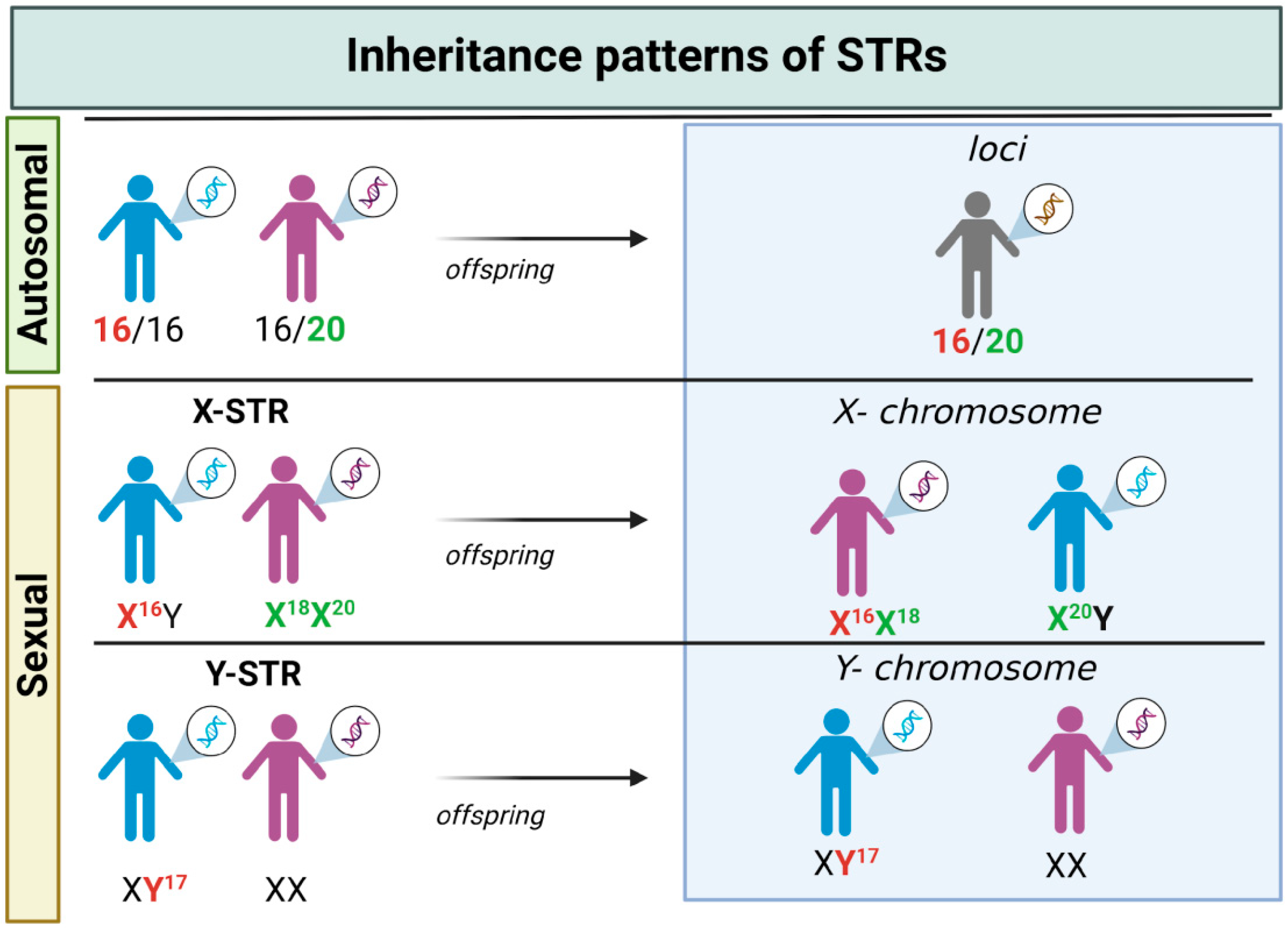
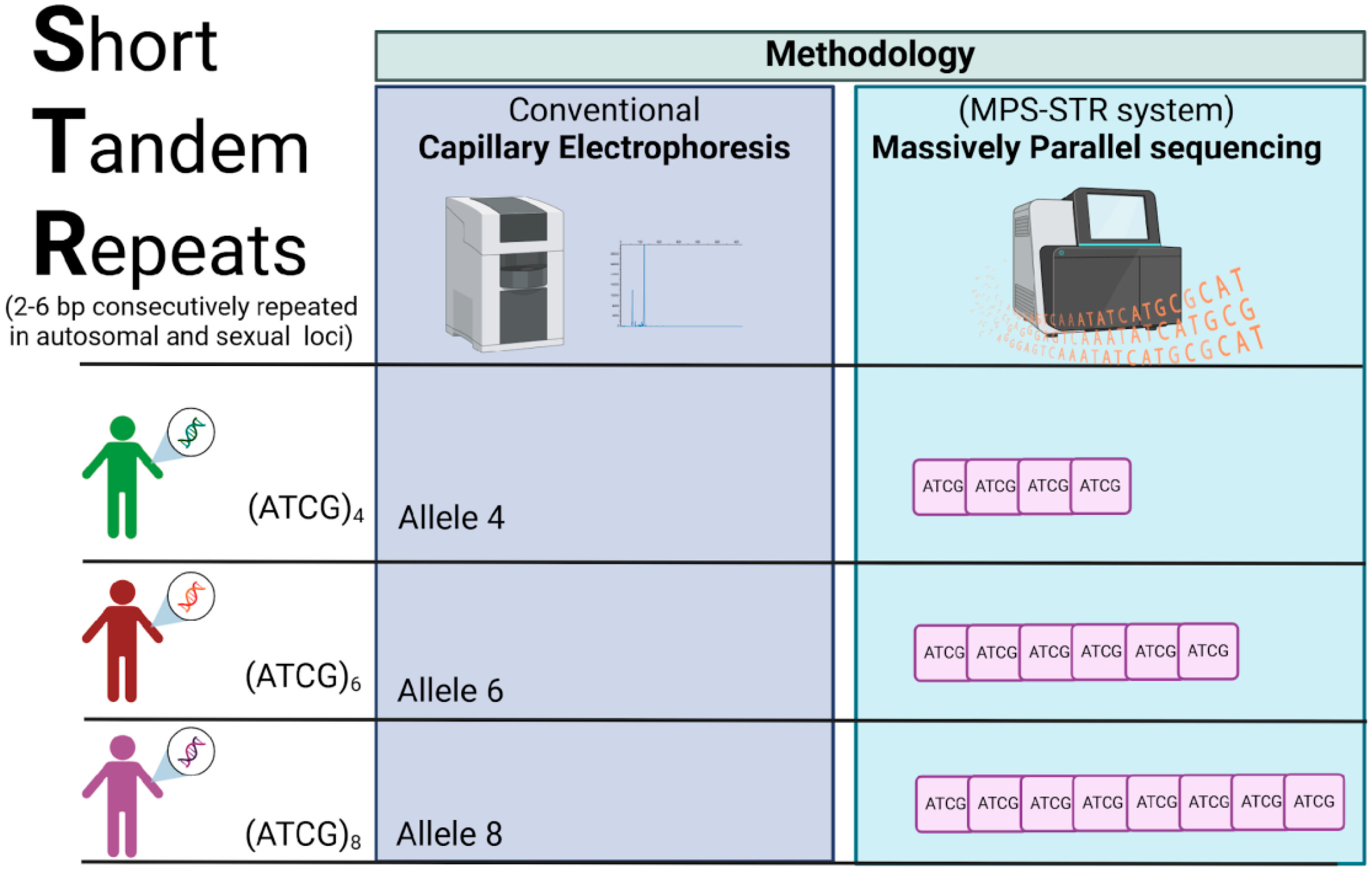
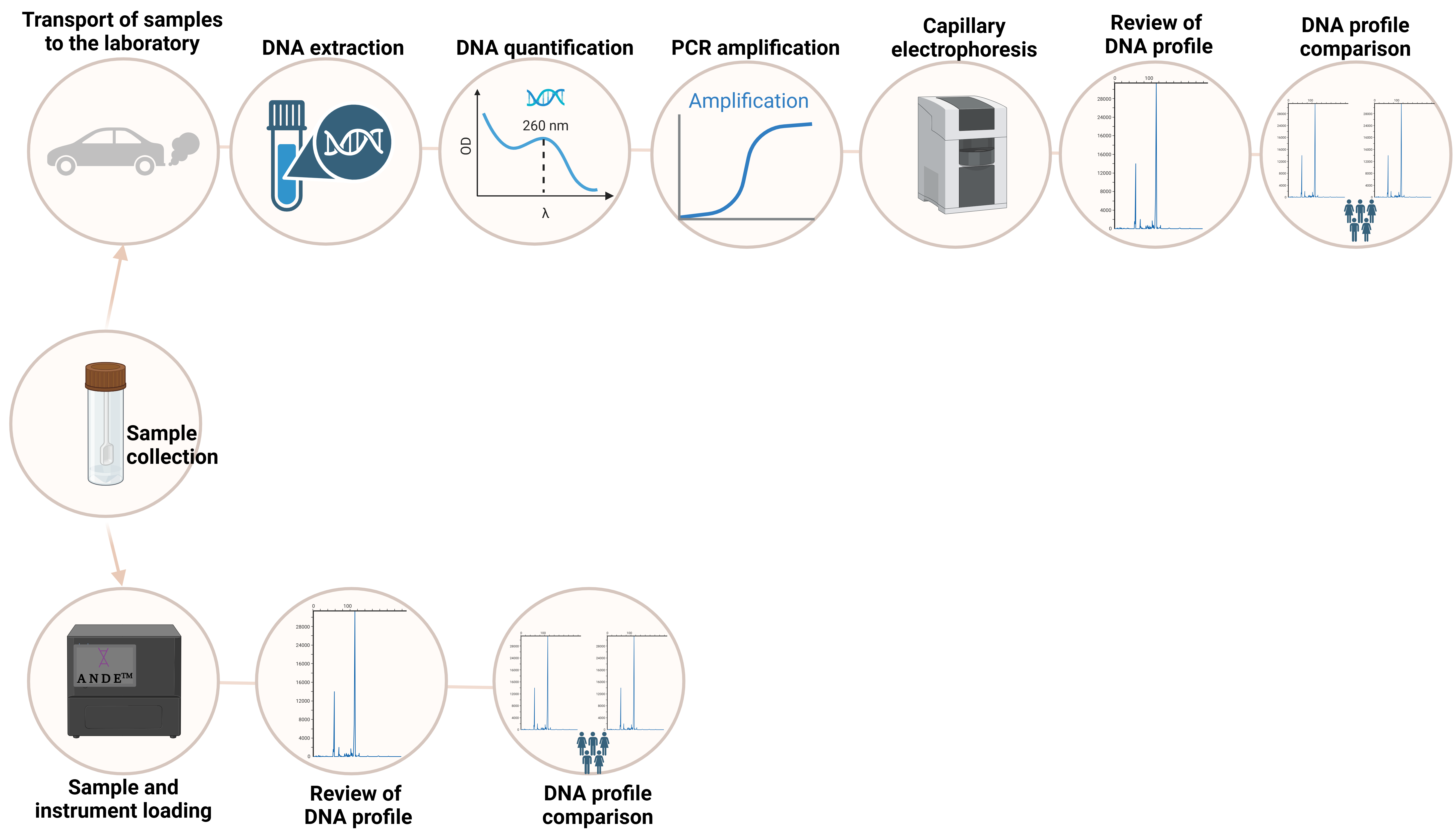
3.1. STRs: The Gold Standard Markers in Forensic Genetics
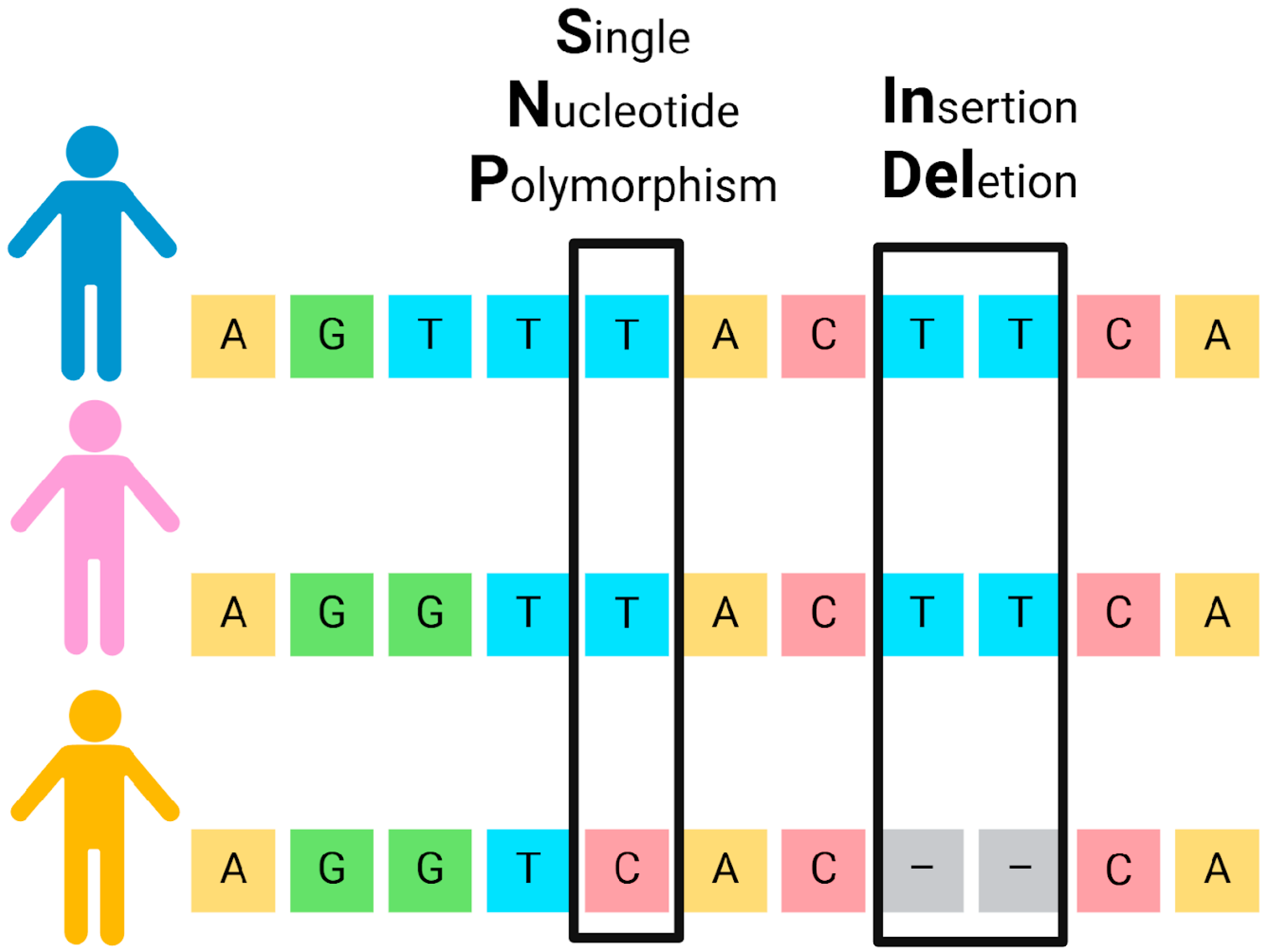
3.2. SNPs and INDELs: Complementary Binary Genetic Markers
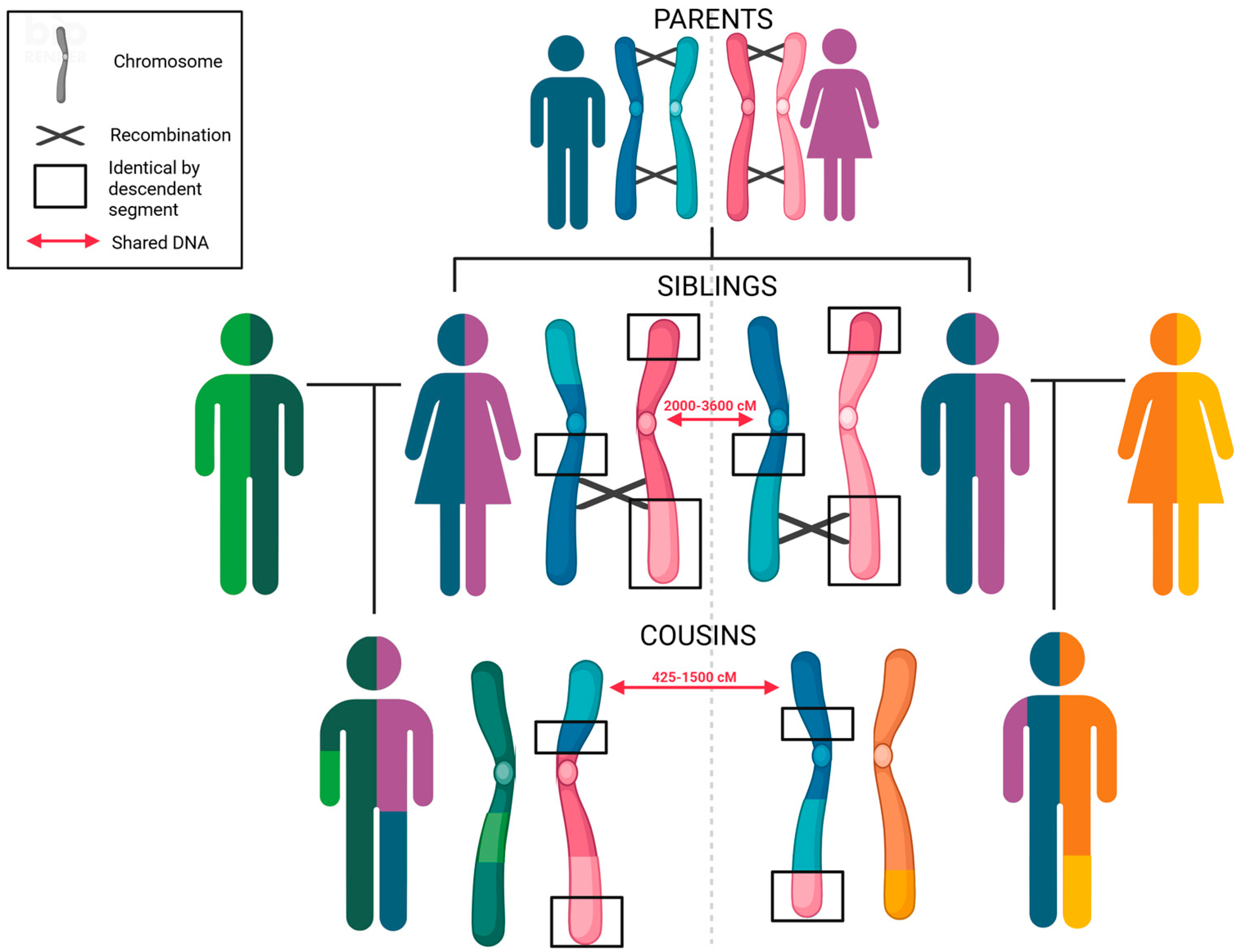
3.3. Statistical Analysis and Interpretation of Autosomal Genetic Evidence
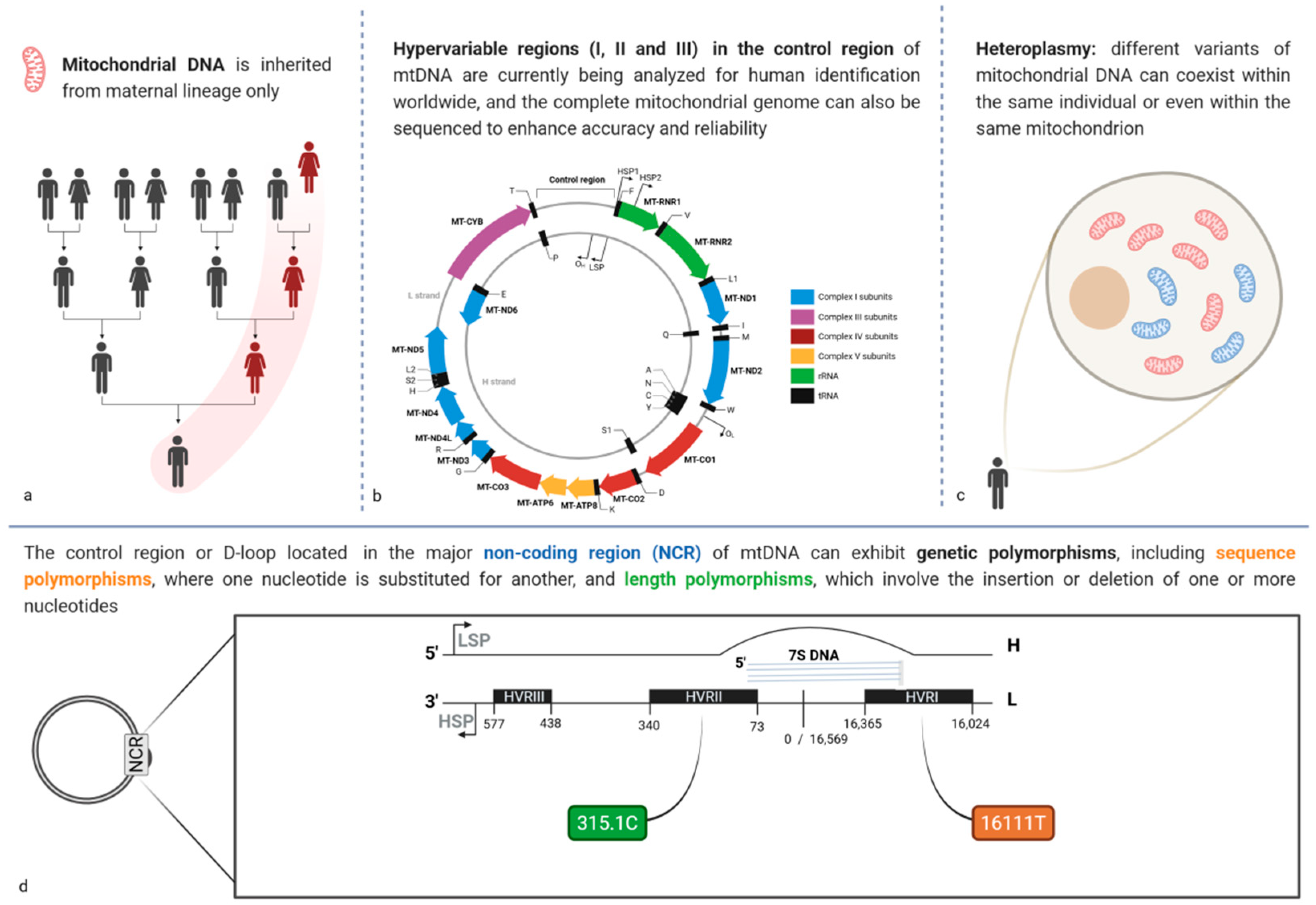
3.4. Mitochondrial DNA Polymorphisms: A Tool of Maternal Heritage
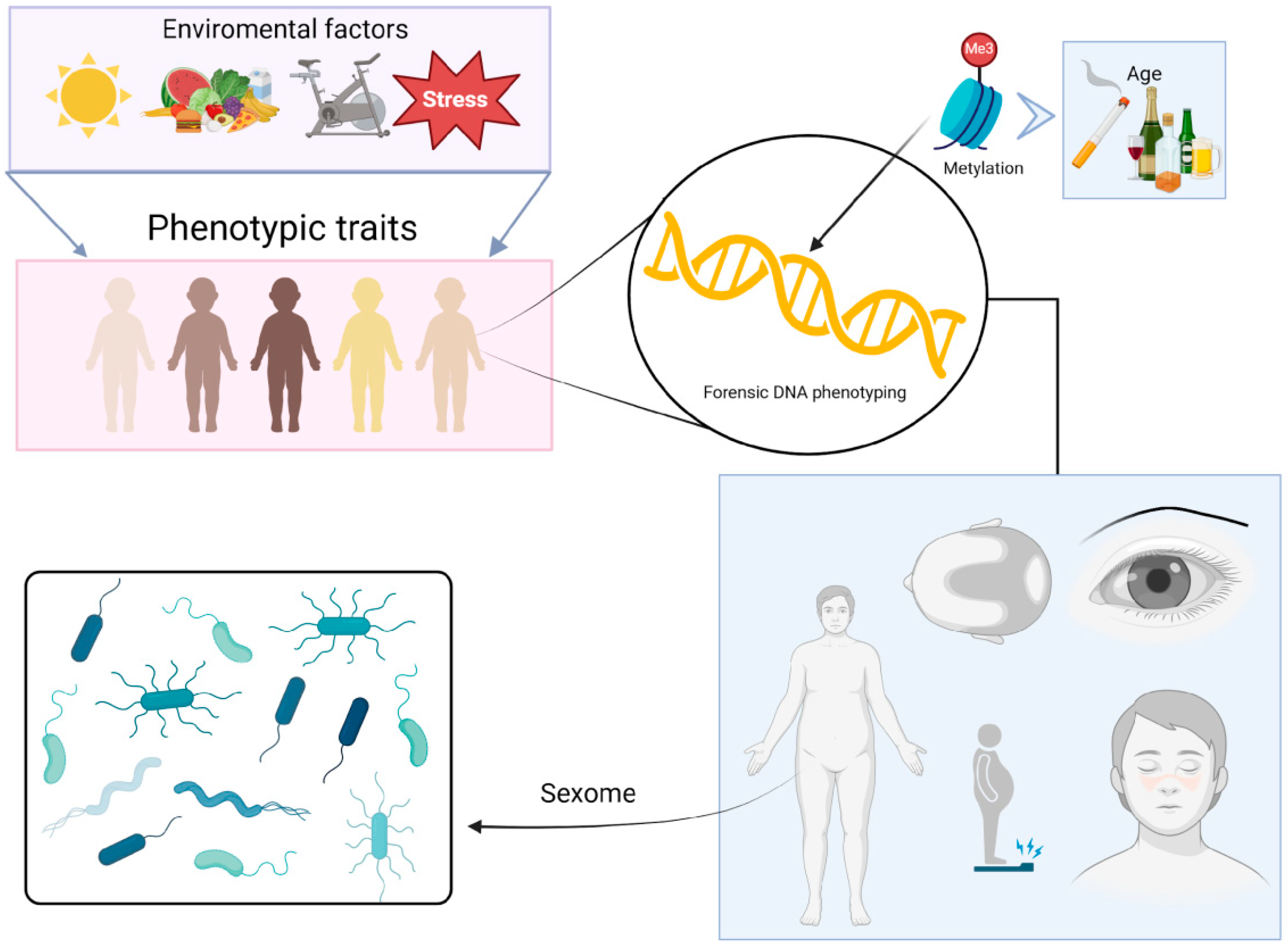
3.5. DNA Phenotyping and Ancestry Inference
3.6. DNA Transfer
3.7. Forensic Genetic Genealogy
3.8. Rapid DNA Profiling
3.9. Non-Human DNA Typing
4. Future Perspectives
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shen, M.; Vieira, D.N. Forensic Science: Defending Justice. Forensic Sci. Res. 2016, 1, 1–2. [Google Scholar] [CrossRef] [PubMed]
- de Knijff, P. From next generation sequencing to now generation sequencing in forensics. Forensic Sci. Int. Genet. 2019, 38, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.M.; Buel, E.; Crivellente, F.; McCord, B.R. Forensic DNA typing by capillary electrophoresis using the ABI Prism 310 and 3100 genetic analyzers for STR analysis. Electrophoresis 2004, 25, 1397–1412. [Google Scholar] [CrossRef]
- MacLean, C.E.; Lamparello, A. Forensic DNA phenotyping in criminal investigations and criminal courts: Assessing and mitigating the dilemmas inherent in the science. Recent Adv. DNA Gene Seq. 2014, 8, 104–112. [Google Scholar]
- Kayser, M. Forensic DNA Phenotyping: DNA testing for externally visible characteristics. In Encyclopedia of Forensic Sciences; Siegel, J.A., Saukko, P.J., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 369–374. [Google Scholar]
- Landsteiner, K. Zur Kenntnis der Anti-Fermentatiren, Lytrschen and Agglutinierenden Wirkungen des Bluteerums und der Lymphe. Zentralbi Bakteriol 1900, 27, 357–362. [Google Scholar]
- Bugert, P.; Klüter, H. 100 Years after von Dungern & Hirschfeld: Kinship Investigation from Blood Groups to SNPs. Transfus. Med. Hemother. Off. Organ Dtsch. Ges. Transfusionsmedizin Immunhamatol. 2012, 39, 161–162. [Google Scholar]
- Landsteiner, K.; Levine, P. On individual differences in human blood. J. Exp. Med. 1928, 47, 757–775. [Google Scholar] [CrossRef]
- Landsteiner, K.; Levine, P. On the inheritance of agglutinogens of human blood demonstrable by immune agglutinins. J. Exp. Med. 1928, 48, 731–749. [Google Scholar] [CrossRef]
- Landsteiner, K.; Wiener, A.S. Studies on an agglutinogen (rh) in human blood reacting with anti-rhesus sera and with human isoantibodies. J. Exp. Med. 1941, 74, 309–320. [Google Scholar] [CrossRef]
- Sussman, L.N. Blood grouping tests in disputed paternity proceedings; studies with ABO, MN, and Rh-Hr factors. J. Am. Med. Assoc. 1954, 155, 1143–1145. [Google Scholar] [CrossRef]
- Sussman, L.N. Blood grouping tests. A review of 1000 cases of disputed paternity. Am. J. Clin. Pathol. 1963, 40, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Wiener, A.S.; Sonn, E.B.; Belkin, R.B. Heredity of the rh blood types. J. Exp. Med. 1944, 79, 235–253. [Google Scholar] [CrossRef] [PubMed]
- Landsteiner, K. Forensic application of serologic individuality tests. J. Am. Med. Assoc. 1934, 103, 1041–1044. [Google Scholar] [CrossRef]
- Boyd, W.C. Use of blood groups in cases of disputed paternity. N. Engl. J. Med. 1949, 241, 759–760. [Google Scholar] [CrossRef]
- Cerda-Flores, R.M.; Barton, S.A.; Marty-Gonzalez, L.F.; Rivas, F.; Chakraborty, R. Estimation of nonpaternity in the Mexican population of Nuevo Leon: A validation study with blood group markers. Am. J. Phys. Anthropol. 1999, 109, 281–293. [Google Scholar] [CrossRef]
- Harley, D.; Lynch, G.R. Blood Tests for Paternity. Br. Med. J. 1937, 1, 163–166. [Google Scholar] [CrossRef]
- Dausset, J. Iso-leuko-antibodies. Acta Haematol. 1958, 20, 156–166. [Google Scholar] [CrossRef]
- Terasaki, P.I. HLA in paternity testing. West. J. Med. 1978, 128, 48. [Google Scholar]
- Lee, C.L.; Lebeck, L.K.; Wong, C. Estimating paternity index from HLA-typing results. Am. J. Clin. Pathol. 1980, 74, 218–223. [Google Scholar] [CrossRef]
- Singh, G.; Johns, M.M.; Paul, G. Paternity testing: Analysis of six blood groups and HLA markers, with particular reference to comparison of races. Am. J. Clin. Pathol. 1982, 78, 748–752. [Google Scholar] [CrossRef]
- Borowsky, R. HLA and the probability of paternity. Am. J. Hum. Genet. 1988, 42, 132–134. [Google Scholar] [PubMed]
- Davey, F.R.; Hubbell, C.A.; Lauenstein, K.J.; Tinnesz, C.; Henry, J.B. Analysis of paternity. The use of HLA and red cell antigens. Transfusion 1984, 24, 340–342. [Google Scholar] [CrossRef] [PubMed]
- Houtz, T.D.; Brooks, M.A.; Wenk, R.E.; Dawson, R.B. Utility of HLA and six erythrocyte antigen systems in excluding paternity among 500 disputed cases. Forensic Sci. Int. 1981, 17, 211–218. [Google Scholar] [CrossRef]
- Reisner, E.G.; MacQueen, J.M. Problems arising from the use of the HLA system in paternity testing. Clin. Lab. Haematol. 1981, 3, 113–119. [Google Scholar] [CrossRef]
- Butler, J.M. Forensic DNA Typing: Biology, Technology, and Genetics of STR Markers; Elsevier: Amsterdam, The Netherlands, 2005; Volume 1. [Google Scholar]
- Smithies, O. Zone electrophoresis in starch gels: Group variations in the serum proteins of normal human adults. Biochem. J. 1955, 61, 629–641. [Google Scholar] [CrossRef]
- Dykes, D.D.; Polesky, H.F. The usefulness of serum protein and erythrocyte enzyme polymorphisms in paternity testing. Am. J. Clin. Pathol. 1976, 65, 982–986. [Google Scholar] [CrossRef]
- Spencer, N.; Hopkinson, D.A.; Harris, H. Phosphoglucomutase polymorphism in man. Nature 1964, 204, 742–745. [Google Scholar] [CrossRef]
- Fuhrmann, W.; Lichte, K.H. Human red cell acid phosphatase polymorphism. A study on gene frequency and forensic use of the system in cases of disputed paternity. Humangenetik 1966, 3, 121–126. [Google Scholar]
- Parr, C.W. Erythrocyte phosphogluconate dehydrogenase polymorphism. Nature 1966, 210, 487–489. [Google Scholar] [CrossRef]
- Seppälä, M.; Ruoslahti, E.; Seppälä, I.J. Transferrin system in paternity cases. An immunological verification test for heterozygosity. Ann. Med. Exp. Biol. Fenn. 1967, 45, 14–15. [Google Scholar]
- McCombs, M.L.; Bowman, B.H. Demonstration of inherited ceruloplasmin variants in human Serum by acrylamide electrophoresis. Tex. Rep. Biol. Med. 1969, 27, 769–772. [Google Scholar] [PubMed]
- Harris, H. Protein polymorphism in man. Can. J. Genet. Cytol. J. Can. Genet. Cytol. 1971, 13, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, H.; Fiedler, H.; Niebuhr, R. Value of the adenosine deaminase (ADA) isoenzyme system for forensic evidence: Statistical dependability testing of paternity exclusion claims. Z. Rechtsmed. J. Leg. Med. 1972, 71, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Bender, K.; Volkmann, E. Paternity exclusion using the transferrin system. Z. Immun. Allerg. Klin. Immunol. 1970, 140, 468–471. [Google Scholar]
- Waldinger, D.; Cleve, H. Two-dimensional electrophoresis of human lymphocyte proteins: Two-dimensional polymorphisms and paternity testing. Electrophoresis 1988, 9, 375–379. [Google Scholar] [CrossRef]
- Salmon, D.B.; Brocteur, J. Probability of paternity exclusion when relatives are involved. Am. J. Hum. Genet. 1978, 30, 65–75. [Google Scholar]
- Saad, R. Discovery, development, and current applications of DNA identity testing. Proceedings 2005, 18, 130–133. [Google Scholar] [CrossRef]
- Aronson, J.D. DNA fingerprinting on trial: The dramatic early history of a new forensic technique. Endeavour 2005, 29, 126–131. [Google Scholar] [CrossRef]
- Wyman, A.R.; White, R. A highly polymorphic locus in human DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 6754–6758. [Google Scholar] [CrossRef]
- Jeffreys, A.J.; Wilson, V.; Thein, S.L. Hypervariable ‘minisatellite’ regions in human DNA. Nature 1985, 314, 67–73. [Google Scholar] [CrossRef]
- Jeffreys, A.J.; Wilson, V.; Thein, S.L. Individual-specific ‘fingerprints’ of human DNA. Nature 1985, 316, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Lindblom, B.; Holmlund, G.; Stennek, A. DNA analyses in forensic investigations. Lakartidningen 1991, 88, 2055–2059. [Google Scholar] [PubMed]
- Tautz, D. Notes on the definition and nomenclature of tandemly repetitive DNA sequences. EXS 1993, 67, 21–28. [Google Scholar]
- Katti, M.V.; Ranjekar, P.K.; Gupta, V.S. Differential distribution of simple sequence repeats in eukaryotic genome sequences. Mol. Biol. Evol. 2001, 18, 1161–1167. [Google Scholar] [CrossRef]
- Jeffreys, A.J.; Brookfield, J.F.; Semeonoff, R. Positive identification of an immigration test-case using human DNA fingerprints. Nature 1985, 317, 818–819. [Google Scholar] [CrossRef]
- Kayser, M. Forensic DNA Phenotyping, Predicting human appearance from crime scene material for investigative purposes. Forensic Sci. Int. Genet. 2015, 18, 33–48. [Google Scholar] [CrossRef]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef]
- Collins, J.R.; Stephens, R.M.; Gold, B.; Long, B.; Dean, M.; Burt, S.K. An exhaustive DNA micro-satellite map of the human genome using high performance computing. Genomics 2003, 82, 10–19. [Google Scholar] [CrossRef]
- Panneerchelvam, S.; Norazmi, M.N. Forensic DNA profiling and database. Malays. J. Med. Sci. 2003, 10, 20–26. [Google Scholar]
- Butler, J.M. Chapter 5—Short Tandem Repeat (STR) Loci and Kits. In Advanced Topics in Forensic DNA Typing: Methodology; Butler, J.M., San, D., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 99–139. [Google Scholar]
- Butler, J.M. Genetics and genomics of core short tandem repeat loci used in human identity testing. J. Forensic Sci. 2006, 51, 253–265. [Google Scholar] [CrossRef]
- Budowle, B.; Moretti, T.R.; Niezgoda, S.J.; Brown, B.L. CODIS and PCR-based short tandem repeat loci: Law enforcement tools. In Proceedings of the Second European Symposium on Human Identification, Innsbruck, Austria, 10–12 June 1998; Promega Corporation: Madison, WI, USA, 1998; pp. 73–88. [Google Scholar]
- Nwawuba Stanley, U.; Mohammed Khadija, A.; Bukola, A.T.; Omusi Precious, I.; Ayevbuomwan Davidson, E. Forensic DNA Profiling: Autosomal Short Tandem Repeat as a Prominent Marker in Crime Investigation. Malays. J. Med. Sci. 2020, 27, 22–35. [Google Scholar] [PubMed]
- ENFSI Standing Committee for Quality and Competence (QCC). Validation and Implementation of (New) Methods; 2006. Ref. Code: QCC-VAL-001. Available online: http://www.enfsi.eu (accessed on 4 May 2025).
- Scientific Working Group on DNA Analysis Methods (SWGDAM). Validation Guidelines for DNA Analysis Methods; SWGDAM: online, 2012. [Google Scholar]
- Hares, D.R. Selection and implementation of expanded CODIS core loci in the United States. Forensic Sci. Int. Genet. 2015, 17, 33–34. [Google Scholar] [CrossRef] [PubMed]
- LaFountain, M.J.; Schwartz, M.B.; Svete, P.A.; Walkinshaw, M.A.; Buel, E. TWGDAM validation of the AmpFlSTR Profiler Plus and AmpFlSTR COfiler STR multiplex systems using capillary electrophoresis. J. Forensic Sci. 2001, 46, 1191–1198. [Google Scholar] [CrossRef]
- Collins, P.J.; Hennessy, L.K.; Leibelt, C.S.; Roby, R.K.; Reeder, D.J.; Foxall, P.A. Developmental validation of a single-tube amplification of the 13 CODIS STR loci, D2S1338, D19S433, and amelogenin: The AmpFlSTR Identifiler PCR Amplification Kit. J. Forensic Sci. 2004, 49, 1265–1277. [Google Scholar] [CrossRef]
- Ludeman, M.J.; Zhong, C.; Mulero, J.J.; Lagacé, R.E.; Hennessy, L.K.; Short, M.L.; Wang, D.Y. Developmental validation of GlobalFiler™ PCR amplification kit: A 6-dye multiplex assay designed for amplification of casework samples. Int. J. Leg. Med. 2018, 132, 1555–1573. [Google Scholar] [CrossRef]
- Mattayat, D.; Kitpipit, T.; Phetpeng, S.; Asawutmangkul, W.; Thanakiatkrai, P. Comparative performance of AmpFLSTR® Identifiler® Plus PCR amplification kit and QIAGEN® Investigator® IDplex Plus kit. Sci. Justice J. Forensic Sci. Soc. 2016, 56, 468–474. [Google Scholar] [CrossRef]
- Levedakou, E.N.; Freeman, D.A.; Budzynski, M.J.; Early, B.E.; Damaso, R.C.; Pollard, A.M.; Townley, A.J.; Gombos, J.L.; Lewis, J.L.; Kist, F.G.; et al. Characterization and validation studies of powerplex 2.1, a nine-locus short tandem repeat (STR) multiplex system and penta D monoplex. J. Forensic Sci. 2002, 47, 757–772. [Google Scholar] [CrossRef]
- Krenke, B.E.; Tereba, A.; Anderson, S.J.; Buel, E.; Culhane, S.; Finis, C.J.; Tomsey, C.S.; Zachetti, J.M.; Masibay, A.; Rabbach, D.R.; et al. Validation of a 16-locus fluorescent multiplex system. J. Forensic Sci. 2002, 47, 773–785. [Google Scholar] [CrossRef]
- Ensenberger, M.G.; Hill, C.R.; McLaren, R.S.; Sprecher, C.J.; Storts, D.R. Developmental validation of the PowerPlex(®) 21 System. Forensic Sci. Int. Genet. 2014, 9, 169–178. [Google Scholar] [CrossRef]
- Oostdik, K.; Lenz, K.; Nye, J.; Schelling, K.; Yet, D.; Bruski, S.; Strong, J.; Buchanan, C.; Sutton, J.; Linner, J.; et al. Developmental validation of the PowerPlex® Fusion System for analysis of casework and reference samples: A 24-locus multiplex for new database standards. Forensic Sci. Int. Genet. 2014, 12, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Ensenberger, M.G.; Lenz, K.A.; Matthies, L.K.; Hadinoto, G.M.; Schienman, J.E.; Przech, A.J.; Morganti, M.W.; Renstrom, D.T.; Baker, V.M.; Gawrys, K.M.; et al. Developmental validation of the PowerPlex® Fusion 6C System. Forensic Sci. Int. Genet. 2016, 21, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, P.; Dixit, S.; Kumawat, R.K.; Srivastava, A. Efficiency analysis of VersaPlex™ 27PY system in Central Indian Population: First report from Indian population. Leg. Med. 2022, 54, 101983. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.; Prochnow, A.; Bussmann, M.; Scherer, M.; Peist, R.; Steffen, C. Developmental validation of QIAGEN Investigator® 24plex QS Kit and Investigator® 24plex GO! Kit: Two 6-dye multiplex assays for the extended CODIS core loci. Forensic Sci. Int. Genet. 2017, 29, 9–20. [Google Scholar] [CrossRef]
- Martin, P.D.; Schmitter, H.; Schneider, P.M. A brief history of the formation of DNA databases in forensic science within Europe. Forensic Sci. Int. 2001, 119, 225–231. [Google Scholar] [CrossRef]
- Ruitberg, C.M.; Reeder, D.J.; Butler, J.M. STRBase: A short tandem repeat DNA database for the human identity testing community. Nucleic Acids Res. 2001, 29, 320–322. [Google Scholar] [CrossRef]
- Roewer, L.; Krawczak, M.; Willuweit, S.; Nagy, M.; Alves, C.; Amorim, A.; Anslinger, K.; Augustin, C.; Betz, A.; Bosch, E.; et al. Online reference database of European Y-chromosomal short tandem repeat (STR) haplotypes. Forensic Sci. Int. 2001, 118, 106–113. [Google Scholar] [CrossRef]
- Bodner, M.; Bastisch, I.; Butler, J.M.; Fimmers, R.; Gill, P.; Gusmão, L.; Morling, N.; Phillips, C.; Prinz, M.; Schneider, P.M.; et al. Recommendations of the DNA Commission of the International Society for Forensic Genetics (ISFG) on quality control of autosomal Short Tandem Repeat allele frequency databasing (STRidER). Forensic Sci. Int. Genet. 2016, 24, 97–102. [Google Scholar] [CrossRef]
- Sobrino, B.; Carracedo, A. SNP typing in forensic genetics: A review. Methods Mol. Biol. 2005, 297, 107–126. [Google Scholar]
- Butler, J.M. Single Nucleotide polymorphisms and Applications. In Advanced Topics in Forensic DNA Typing: Methodology; Butler, J., Walthan, M.A., Eds.; Elsevier/Academic Press: Amsterdam, The Netherlands, 2012; pp. 347–370. [Google Scholar]
- Phillips, C. Application of Autosomal SNPs and Indels in Forensic Analysis. Forensic Sci. Rev. 2012, 24, 43–62. [Google Scholar]
- Budowle, B.; van Daal, A. Forensically relevant SNP classes. Biotechniques 2008, 44, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Malkki, M.; Petersdorf, E.W. Genotyping of single nucleotide polymorphisms by 5′ nuclease allelic discrimination. Methods Mol. Biol. 2012, 882, 173–182. [Google Scholar] [PubMed]
- Mehta, B.; Daniel, R.; Phillips, C.; McNevin, D. Forensically relevant SNaPshot(®) assays for human DNA SNP analysis: A review. Int. J. Leg. Med. 2017, 131, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.B.; King, J.L.; Warshauer, D.H.; Davis, C.P.; Ge, J.; Budowle, B. Single nucleotide polymorphism typing with massively parallel sequencing for human identification. Int. J. Leg. Med. 2013, 127, 1079–1086. [Google Scholar] [CrossRef]
- Alonso, A.; Barrio, P.A.; Müller, P.; Köcher, S.; Berger, B.; Martin, P.; Bodner, M.; Willuweit, S.; Parson, W.; Roewer, L.; et al. Current state-of-art of STR sequencing in forensic genetics. Electrophoresis 2018, 39, 2655–2668. [Google Scholar] [CrossRef]
- Zidkova, A.; Horinek, A.; Kebrdlova, V.; Korabecna, M. Application of the new insertion-deletion polymorphism kit for forensic identification and parentage testing on the Czech population. Int. J. Leg. Med. 2013, 127, 7–10. [Google Scholar] [CrossRef]
- Holt, C.L.; Stephens, K.M.; Walichiewicz, P.; Fleming, K.D.; Forouzmand, E.; Wu, S.F. Human Mitochondrial Control Region and mtGenome: Design and Forensic Validation of NGS Multiplexes, Sequencing and Analytical Software. Genes 2021, 12, 599. [Google Scholar] [CrossRef]
- Venables, S.J.; Mehta, B.; Daniel, R.; Walsh, S.J.; van Oorschot, R.A.; McNevin, D. Assessment of high resolution melting analysis as a potential SNP genotyping technique in forensic casework. Electrophoresis 2014, 35, 3036–3043. [Google Scholar] [CrossRef]
- Palencia-Madrid, L.; Xavier, C.; de la Puente, M.; Hohoff, C.; Phillips, C.; Kayser, M.; Parson, W. Evaluation of the VISAGE Basic Tool for Appearance and Ancestry Prediction Using PowerSeq Chemistry on the MiSeq FGx System. Genes 2020, 11, 708. [Google Scholar] [CrossRef]
- Breslin, K.; Wills, B.; Ralf, A.; Ventayol Garcia, M.; Kukla-Bartoszek, M.; Pospiech, E.; Freire-Aradas, A.; Xavier, C.; Ingold, S.; de La Puente, M.; et al. HIrisPlex-S system for eye, hair, and skin color prediction from DNA: Massively parallel sequencing solutions for two common forensically used platforms. Forensic Sci. Int. Genet. 2019, 43, 102152. [Google Scholar] [CrossRef]
- Kiesler, K.M.; Borsuk, L.A.; Steffen, C.R.; Vallone, P.M.; Gettings, K.B. US Population Data for 94 Identity-Informative SNP Loci. Genes 2023, 14, 1071. [Google Scholar] [CrossRef] [PubMed]
- Fondevila, M.; Pereira, R.; Gusmão, L.; Phillips, C.; Lareu, M.V.; Carracedo, A. Forensic performance of insertion-deletion marker systems. Forensic Sci. Int. Genet. Suppl. Ser. 2011, 3, e443–e444. [Google Scholar] [CrossRef]
- Manta, F.; Caiafa, A.; Pereira, R.; Silva, D.; Amorim, A.; Carvalho, E.F.; Gusmão, L. Indel markers: Genetic diversity of 38 polymorphisms in Brazilian populations and application in a paternity investigation with post mortem material. Forensic Sci. Int. Genet. 2012, 6, 658–661. [Google Scholar] [CrossRef]
- LaRue, B.L.; Ge, J.; King, J.L.; Budowle, B. A validation study of the Qiagen Investigator DIPplex® kit; an INDEL-based assay for human identification. Int. J. Leg. Med. 2012, 126, 533–540. [Google Scholar] [CrossRef]
- Casals, F.; Rasal, R.; Anglada, R.; Tormo, M.; Bonet, N.; Rivas, N.; Vásquez, P.; Calafell, F. A forensic population database in El Salvador: 58 STRs and 94 SNPs. Forensic Sci. Int. Genet. 2022, 57, 102646. [Google Scholar] [CrossRef]
- Aguilar-Velázquez, J.A.; Duran-Salazar, M.; Córdoba-Mercado, M.F.; Coronado-Avila, C.E.; Salas-Salas, O.; Martinez-Cortés, G.; Casals, F.; Calafell, F.; Ramos-González, B.; Rangel-Villalobos, H. Characterization of 58 STRs and 94 SNPs with the ForenSeq™ DNA signature prep kit in Mexican-Mestizos from the Monterrey city (Northeast, Mexico). Mol. Biol. Rep. 2022, 49, 7601–7609. [Google Scholar] [CrossRef]
- Guevara, E.K.; Palo, J.U.; King, J.L.; Buś, M.M.; Guillén, S.; Budowle, B.; Sajantila, A. Autosomal STR and SNP characterization of populations from the Northeastern Peruvian Andes with the ForenSeqTM; DNA Signature Prep Kit. Forensic Sci. Int. Genet. 2021, 52, 102487. [Google Scholar] [CrossRef]
- Peng, D.; Zhang, Y.; Ren, H.; Li, H.; Li, R.; Shen, X.; Wang, N.; Huang, E.; Wu, R.; Sun, H. Identification of sequence polymorphisms at 58 STRs and 94 iiSNPs in a Tibetan population using massively parallel sequencing. Sci. Rep. 2020, 10, 12225. [Google Scholar] [CrossRef]
- Delest, A.; Godfrin, D.; Chantrel, Y.; Ulus, A.; Vannier, J.; Faivre, M.; Hollard, C.; Laurent, F.-X. Sequenced-based French population data from 169 unrelated individuals with Verogen’s ForenSeq DNA signature prep kit. Forensic Sci. Int. Genet. 2020, 47, 102304. [Google Scholar] [CrossRef]
- Hussing, C.; Bytyci, R.; Huber, C.; Morling, N.; Børsting, C. The Danish STR sequence database: Duplicate typing of 363 Danes with the ForenSeq™ DNA Signature Prep Kit. Int. J. Leg. Med. 2019, 133, 325–334. [Google Scholar] [CrossRef]
- Butler, J.M. Relationship Testing: Kinship Statistics. In Advanced Topics in Forensic DNA Typing: Interpretation, 1st ed.; Butler, J.M., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 363–401. [Google Scholar]
- Wein, S. Forensic DNA analysis and statistics. In Forensic DNA Applications: An Interdisciplinary Perspective, 2nd ed.; Scott, M.S., Ed.; CRC Press: Boca Raton, FL, USA, 2024; pp. 29–56. [Google Scholar]
- Shrivastava, P. Calculation of paternity index in paternity dispute and identification cases. In Principles and Practices of DNA Analysis: A Laboratory Manual for Forensic DNA Typing, 1st ed.; Trivedi, V.B., Ed.; Springer: Singapore, 2023; pp. 287–296. [Google Scholar]
- Butler, J.M. DNA profile frequency estimates and match probabilities. In Advanced Topics in Forensic DNA Typing: Interpretation, 1st ed.; Butler, J.M., Ed.; Academic Press: San Diego, CA, USA, 2015; pp. 265–298. [Google Scholar]
- Prinz, M.; Carracedo, A.; Mayr, W.R.; Morling, N.; Parsons, T.J.; Sajantila, A.; Scheithauer, R.; Schmitter, H.; Schneider, P.M. DNA Commission of the International Society for Forensic Genetics (ISFG): Recommendations regarding the role of forensic genetics for disaster victim identification (DVI). Forensic Sci. Int. Genet. 2007, 1, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Gusmão, L.; Butler, J.M.; Carracedo, A.; Gill, P.; Kayser, M.; Mayr, W.R.; Morling, N.; Prinz, M.; Roewer, L.; Tyler-Smith, C.; et al. DNA Commission of the International Society of Forensic Genetics (ISFG): An update of the recommendations on the use of Y-STRs in forensic analysis. Forensic Sci. Int. 2006, 157, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Tillmar, A.O.; Kling, D.; Butler, J.M.; Parson, W.; Prinz, M.; Schneider, P.M.; Egeland, T.; Gusmão, L. DNA Commission of the International Society for Forensic Genetics (ISFG): Guidelines on the use of X-STRs in kinship analysis. Forensic Sci. Int. Genet. 2017, 29, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Gjertson, D.W.; Brenner, C.H.; Baur, M.P.; Carracedo, A.; Guidet, F.; Luque, J.A.; Lessig, R.; Mayr, W.R.; Pascali, V.L.; Prinz, M.; et al. ISFG: Recommendations on biostatistics in paternity testing. Forensic Sci. Int. Genet. 2007, 1, 223–231. [Google Scholar] [CrossRef]
- Yan, C.; Duanmu, X.; Zeng, L.; Liu, B.; Song, Z. Mitochondrial DNA: Distribution, Mutations, and Elimination. Cells 2019, 8, 379. [Google Scholar] [CrossRef]
- Budowle, B.; Allard, M.W.; Wilson, M.R.; Chakraborty, R. Forensics and mitochondrial DNA: Applications, debates, and foundations. Annu. Rev. Genom. Hum. Genet. 2003, 4, 119–141. [Google Scholar] [CrossRef]
- Amorim, A.; Fernandes, T.; Taveira, N. Mitochondrial DNA in human identification: A review. PeerJ 2019, 7, e7314. [Google Scholar] [CrossRef]
- Ohuchi, T.; Guan, X.; Funayama, M. Evaluation of the Utility of Mitochondrial DNA Testing in Personal Identification Work in the Great East Japan Earthquake of 2011. Tohoku J. Exp. Med. 2021, 255, 275–281. [Google Scholar] [CrossRef]
- Parson, W.; Bandelt, H.J. Extended guidelines for mtDNA typing of population data in forensic science. Forensic Sci. Int. Genet. 2007, 1, 13–19. [Google Scholar] [CrossRef]
- Bär, W.; Brinkmann, B.; Budowle, B.; Carracedo, A.; Gill, P.; Holland, M.; Lincoln, P.J.; Mayr, W.; Morling, N.; Olaisen, B.; et al. Guidelines for mitochondrial DNA typing. DNA Commission of the International Society for Forensic Genetics. Vox Sang. 2000, 79, 121–125. [Google Scholar] [CrossRef]
- SWGDAM. Interpretation Guidelines for Mitochondrial DNA Analysis by Forensic DNA Testing Laboratories; SWGDAM: online, 2019. [Google Scholar]
- Parson, W.; Gusmão, L.; Hares, D.R.; Irwin, J.A.; Mayr, W.R.; Morling, N.; Pokorak, E.; Prinz, M.; Salas, A.; Schneider, P.M.; et al. DNA Commission of the International Society for Forensic Genetics: Revised and extended guidelines for mitochondrial DNA typing. Forensic Sci. Int. Genet. 2014, 13, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, K.; Kasai, K.; Levin, B.C. Inter- and intragenerational transmission of a human mitochondrial DNA heteroplasmy among 13 maternally-related individuals and differences between and within tissues in two family members. Mitochondrion 2003, 2, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Tsai, L.C.; Lin, C.Y.; Lee, J.C.; Chang, J.G.; Linacre, A.; Goodwin, W. Sequence polymorphism of mitochondrial D-loop DNA in the Taiwanese Han population. Forensic Sci. Int. 2001, 119, 239–247. [Google Scholar] [CrossRef]
- Guardado-Estrada, M.; Juarez-Torres, E.; Medina-Martinez, I.; Wegier, A.; Macías, A.; Gomez, G.; Cruz-Talonia, F.; Roman-Bassaure, E.; Piñero, D.; Kofman-Alfaro, S.; et al. A great diversity of Amerindian mitochondrial DNA ancestry is present in the Mexican mestizo population. J. Hum. Genet. 2009, 54, 695–705. [Google Scholar] [CrossRef]
- Nishimaki, Y.; Sato, K.; Fang, L.; Ma, M.; Hasekura, H.; Boettcher, B. Sequence polymorphism in the mtDNA HV1 region in Japanese and Chinese. Leg. Med. 1999, 1, 238–249. [Google Scholar] [CrossRef]
- Alvarez, J.C.; Johnson, D.L.; Lorente, J.A.; Martinez-Espin, E.; Martinez-Gonzalez, L.J.; Allard, M.; Wilson, M.R.; Budowle, B. Characterization of human control region sequences for Spanish individuals in a forensic mtDNA data set. Leg. Med. 2007, 9, 293–304. [Google Scholar] [CrossRef]
- Prieto, L.; Zimmermann, B.; Goios, A.; Rodriguez-Monge, A.; Paneto, G.G.; Alves, C.; Alonso, A.; Fridman, C.; Cardoso, S.; Lima, G.; et al. The GHEP-EMPOP collaboration on mtDNA population data—A new resource for forensic casework. Forensic Sci. Int. Genet. 2011, 5, 146–151. [Google Scholar] [CrossRef]
- Parson, W.; Dür, A. EMPOP—A forensic mtDNA database. Forensic Sci. Int. Genet. 2007, 1, 88–92. [Google Scholar] [CrossRef]
- Woerner, A.E.; Ambers, A.; Wendt, F.R.; King, J.L.; Moura-Neto, R.S.; Silva, R.; Budowle, B. Evaluation of the precision ID mtDNA whole genome panel on two massively parallel sequencing systems. Forensic Sci. Int. Genet. 2018, 36, 213–224. [Google Scholar] [CrossRef]
- Wood, M.R.; Sturk-Andreaggi, K.; Ring, J.D.; Huber, N.; Bodner, M.; Crawford, M.H.; Parson, W.; Marshall, C. Resolving mitochondrial haplogroups B2 and B4 with next-generation mitogenome sequencing to distinguish Native American from Asian haplotypes. Forensic Sci. Int. Genet. 2019, 43, 102143. [Google Scholar] [CrossRef]
- Kayser, M.; de Knijff, P. Improving human forensics through advances in genetics, genomics and molecular biology. Nat. Rev. Genet. 2011, 12, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Tozzo, P.; Politi, C.; Delicati, A.; Gabbin, A.; Caenazzo, L. External visible characteristics prediction through SNPs analysis in the forensic setting: A review. FBL 2021, 26, 828–850. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Liu, F.; Ballantyne, K.N.; van Oven, M.; Lao, O.; Kayser, M. IrisPlex: A sensitive DNA tool for accurate prediction of blue and brown eye colour in the absence of ancestry information. Forenic Sci. Int. Genet. 2011, 5, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Chaitanya, L.; Clarisse, L.; Wirken, L.; Draus-Barini, J.; Kovatsi, L.; Maeda, H.; Ishikawa, T.; Sijen, T.; de Knijff, P.; et al. Developmental validation of the HIrisPlex system: DNA-based eye and hair colour prediction for forensic and anthropological usage. Forensic Sci. Int. Genet. 2014, 9, 150–161. [Google Scholar] [CrossRef]
- Peng, F.; Zhu, G.; Hysi, P.G.; Eller, R.J.; Chen, Y.; Li, Y.; Hamer, M.A.; Zeng, C.; Hopkins, R.L.; Jacobus, C.L.; et al. Genome-Wide Association Studies Identify Multiple Genetic Loci Influencing Eyebrow Color Variation in Europeans. J. Investig. Dermatol. 2019, 139, 1601–1605. [Google Scholar] [CrossRef]
- Hernando, B.; Ibañez, M.V.; Deserio-Cuesta, J.A.; Soria-Navarro, R.; Vilar-Sastre, I.; Martinez-Cadenas, C. Genetic determinants of freckle occurrence in the Spanish population: Towards ephelides prediction from human DNA samples. Forensic Sci. Int. Genet. 2018, 33, 38–47. [Google Scholar] [CrossRef]
- Pośpiech, E.; Karłowska-Pik, J.; Marcińska, M.; Abidi, S.; Andersen, J.D.; Berge, M.V.D.; Carracedo, Á.; Eduardoff, M.; Freire-Aradas, A.; Morling, N.; et al. Evaluation of the predictive capacity of DNA variants associated with straight hair in Europeans. Forensic Sci. Int. Genet. 2015, 19, 280–288. [Google Scholar] [CrossRef]
- Marcińska, M.; Pośpiech, E.; Abidi, S.; Andersen, J.D.; van den Berge, M.; Carracedo, Á.; Eduardoff, M.; Marczakiewicz-Lustig, A.; Morling, N.; Sijen, T.; et al. Evaluation of DNA variants associated with androgenetic alopecia and their potential to predict male pattern baldness. PLoS ONE 2015, 10, e0127852. [Google Scholar] [CrossRef]
- Liu, F.; Zhong, K.; Jing, X.; Uitterlinden, A.G.; Hendriks, A.E.J.; Drop, S.L.S.; Kayser, M. Update on the predictability of tall stature from DNA markers in Europeans. Forensic Sci. Int. Genet. 2019, 42, 8–13. [Google Scholar] [CrossRef]
- Ambroa-Conde, A.; Casares de Cal, M.A.; Gómez-Tato, A.; Robinson, O.; Mosquera-Miguel, A.; de la Puente, M.; Ruiz-Ramírez, J.; Phillips, C.; Lareu, M.V.; Freire-Aradas, A. Inference of tobacco and alcohol consumption habits from DNA methylation analysis of blood. Forensic Sci. Int. Genet. 2024, 70, 103022. [Google Scholar] [CrossRef]
- Dixon, R.; Egan, S.; Hughes, S.; Chapman, B. The Sexome—A proof of concept study into microbial transfer between heterosexual couples after sexual intercourse. Forensic Sci. Int. 2023, 348, 111711. [Google Scholar] [CrossRef] [PubMed]
- Kayser, M.; Branicki, W.; Parson, W.; Phillips, C. Recent advances in Forensic DNA Phenotyping of appearance, ancestry and age. Forensic Sci. Int. Genet. 2023, 65, 102870. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.M.; Wagner, W. Epigenetic-aging-signature to determine age in different tissues. Aging 2011, 3, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Jung, S.E.; Hong, S.R.; Lee, E.H.; Lee, J.H.; Lee, S.D.; Lee, H.Y. Independent validation of DNA-based approaches for age prediction in blood. Forensic Sci. Int. Genet. 2017, 29, 250–256. [Google Scholar] [CrossRef]
- Freire-Aradas, A.; Phillips, C.; Girón-Santamaría, L.; Mosquera-Miguel, A.; Gómez-Tato, A.; Casares de Cal, M.; Álvarez-Dios, J.; Lareu, M.V. Tracking age-correlated DNA methylation markers in the young. Forensic Sci. Int. Genet. 2018, 36, 50–59. [Google Scholar] [CrossRef]
- McEwen, L.M.; O’Donnell, K.J.; McGill, M.G.; Edgar, R.D.; Jones, M.J.; MacIsaac, J.L.; Lin, D.T.S.; Ramadori, K.; Morin, A.; Gladish, N.; et al. The PedBE clock accurately estimates DNA methylation age in pediatric buccal cells. Proc. Natl. Acad. Sci. USA 2020, 117, 23329–23335. [Google Scholar] [CrossRef]
- Koop, B.E.; Reckert, A.; Becker, J.; Han, Y.; Wagner, W.; Ritz-Timme, S. Epigenetic clocks may come out of rhythm-implications for the estimation of chronological age in forensic casework. Int. J. Leg. Med. 2020, 134, 2215–2228. [Google Scholar] [CrossRef]
- Freire-Aradas, A.; Girón-Santamaría, L.; Mosquera-Miguel, A.; Ambroa-Conde, A.; Phillips, C.; Casares de Cal, M.; Gómez-Tato, A.; Álvarez-Dios, J.; Pospiech, E.; Aliferi, A.; et al. A common epigenetic clock from childhood to old age. Forensic Sci. Int. Genet. 2022, 60, 102743. [Google Scholar] [CrossRef]
- Phipps, M.; Petricevic, S. The tendency of individuals to transfer DNA to handled items. Forensic Sci. Int. 2007, 168, 162–168. [Google Scholar] [CrossRef]
- Cavanaugh, S.E.; Bathrick, A.S. Direct PCR amplification of forensic touch and other challenging DNA samples: A review. Forensic Sci. Int. Genet. 2018, 32, 40–49. [Google Scholar] [CrossRef]
- Tozzo, P.; Mazzobel, E.; Marcante, B.; Delicati, A.; Caenazzo, L. Touch DNA Sampling Methods: Efficacy Evaluation and Systematic Review. Int. J. Mol. Sci. 2022, 23, 15541. [Google Scholar] [CrossRef] [PubMed]
- Ladd, C.; Adamowicz, M.S.; Bourke, M.T.; Scherczinger, C.A.; Lee, H.C. A systematic analysis of secondary DNA transfer. J. Forensic Sci. 1999, 44, 1270–1272. [Google Scholar] [CrossRef] [PubMed]
- van den Berge, M.; Ozcanhan, G.; Zijlstra, S.; Lindenbergh, A.; Sijen, T. Prevalence of human cell material: DNA and RNA profiling of public and private objects and after activity scenarios. Forensic Sci. Int. Genet. 2016, 21, 81–89. [Google Scholar] [CrossRef]
- Alessandrini, F.; Cecati, M.; Pesaresi, M.; Turchi, C.; Carle, F.; Tagliabracci, A. Fingerprints as evidence for a genetic profile: Morphological study on fingerprints and analysis of exogenous and individual factors affecting DNA typing. J. Forensic Sci. 2003, 48, 586–592. [Google Scholar] [CrossRef]
- Burrill, J.; Daniel, B.; Frascione, N. A review of trace "Touch DNA" deposits: Variability factors and an exploration of cellular composition. Forensic Sci. Int. Genet. 2019, 39, 8–18. [Google Scholar] [CrossRef]
- Bonsu, D.O.M.; Higgins, D.; Austin, J.J. Forensic touch DNA recovery from metal surfaces—A review. Sci. Justice J. Forensic Sci. Soc. 2020, 60, 206–215. [Google Scholar] [CrossRef]
- Martin, B.; Linacre, A. Direct PCR: A review of use and limitations. Sci. Justice J. Forensic Sci. Soc. 2020, 60, 303–310. [Google Scholar] [CrossRef]
- Federal Bureau of Investigation (FBI). Quality Assurance Standards for Forensic DNA Testing Laboratories. U.S. Department of Justice. 2020. Available online: https://www.swgdam.org/_files/ugd/4344b0_d73afdd0007c4ed6a0e7e2ffbd6c4eb8.pdf (accessed on 3 March 2025).
- van Oorschot, R.A.H.; Phelan, D.G.; Furlong, S.; Scarfo, G.M.; Holding, N.L.; Cummins, M.J. Are you collecting all the available DNA from touched objects? Int. Congr. Ser. 2003, 1239, 803–807. [Google Scholar] [CrossRef]
- Harbison, S.; Marita, F.; Bushell, D. An analysis of the success rate of 908 trace DNA samples submitted to the Crime Sample Database Unit in New Zealand. Aust. J. Forensic Sci. 2008, 40, 49–53. [Google Scholar] [CrossRef]
- Quinones, I.; Daniel, B. Cell free DNA as a component of forensic evidence recovered from touched surfaces. Forensic Sci. Int. Genet. 2012, 6, 26–30. [Google Scholar] [CrossRef]
- Ottens, R.; Templeton, J.; Paradiso, V.; Taylor, D.; Abarno, D.; Linacre, A. Application of direct PCR in forensic casework. Forensic Sci. Int. Genet. Suppl. Ser. 2013, 4, e47–e48. [Google Scholar] [CrossRef]
- Buckingham, A.K.; Harvey, M.L.; van Oorschot, R.A.H. The origin of unknown source DNA from touched objects. Forensic Sci. Int. Genet. 2016, 25, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Lowe, A.; Murray, C.; Whitaker, J.; Tully, G.; Gill, P. The propensity of individuals to deposit DNA and secondary transfer of low level DNA from individuals to inert surfaces. Forensic Sci. Int. 2002, 129, 25–34. [Google Scholar] [CrossRef] [PubMed]
- ISO 18385:2016; Minimizing the Risk of Human DNA Contamination in Products Used to Collect, Store and Analyze Biological Material for Forensic Purposes. ISO: Geneva, Switzerland, 2016.
- Wilson-Wilde, L.; Yakovchyts, D.; Neville, S.; Maynard, P.; Gunn, P. Investigation into ethylene oxide treatment and residuals on DNA and downstream DNA analysis. Sci. Justice 2017, 57, 13–20. [Google Scholar] [CrossRef] [PubMed]
- European DNA Profiling Group (EDNAP); European Network of Forensic Science Institutes (ENFSI). Guideline for DNA Contamination Minimization in DNA Laboratories (ENFSI DNA-GDL-003); European Network of Forensic Science Institutes: Wiesbaden, Germany, 2023. [Google Scholar]
- Vaněk, D.; Sasková, L.; Votrubová, J. Does the new ISO 18385:2016 standard for forensic DNA-grade products need a revision? Forensic Sci. Int. Genet. Suppl. Ser. 2017, 6, e148–e149. [Google Scholar] [CrossRef]
- Dowdeswell, T.L. Forensic genetic genealogy: A profile of cases solved. Forensic Sci. Int. Genet. 2022, 58, 102679. [Google Scholar] [CrossRef]
- Glynn, C.L. Bridging Disciplines to Form a New One: The Emergence of Forensic Genetic Genealogy. Genes 2022, 13, 1381. [Google Scholar] [CrossRef]
- Kling, D.; Phillips, C.; Kennett, D.; Tillmar, A. Investigative genetic genealogy: Current methods, knowledge and practice. Forensic Sci. Int. Genet. 2021, 52, 102474. [Google Scholar] [CrossRef]
- Wang, Z.; Fang, Y.; Liu, Z.; Hao, N.; Zhang, H.H.; Sun, X.; Que, J.; Ding, H. Adapting nanopore sequencing basecalling models for modification detection via incremental learning and anomaly detection. Nat. Commun. 2024, 15, 7148. [Google Scholar] [CrossRef]
- Greytak, E.M.; Moore, C.; Armentrout, S.L. Genetic genealogy for cold case and active investigations. Forensic Sci. Int. 2019, 299, 103–113. [Google Scholar] [CrossRef]
- García, Ó. Genealogía forense. Implicaciones sociales, éticas, legales y científicas. Rev. Española Med. Leg. 2021, 47, 112–119. [Google Scholar] [CrossRef]
- Katsanis, S.H. Pedigrees and Perpetrators: Uses of DNA and Genealogy in Forensic Investigations. Annu. Rev. Genom. Hum. Genet. 2020, 21, 535–564. [Google Scholar] [CrossRef] [PubMed]
- Kennett, D. Using genetic genealogy databases in missing persons cases and to develop suspect leads in violent crimes. Forensic Sci. Int. 2019, 301, 107–117. [Google Scholar] [CrossRef]
- Wickenheiser, R.A. Forensic genealogy, bioethics and the Golden State Killer case. Forensic Sci. Int. Synerg. 2019, 1, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Tillmar, A.; Sturk-Andreaggi, K.; Daniels-Higginbotham, J.; Thomas, J.T.; Marshall, C. The FORCE Panel: An All-in-One SNP Marker Set for Confirming Investigative Genetic Genealogy Leads and for General Forensic Applications. Genes 2021, 12, 1968. [Google Scholar] [CrossRef]
- Minervino, A.C.; Silva Júnior, R.C.; Corte-Real, F. Advancing justice: The impact of Brazil’s convict genetic profile identification project after 5 years. Sci. Justice 2024, 64, 660–664. [Google Scholar] [CrossRef]
- Bowman, Z.; Daniel, R.; Gerostamoulos, D.; Woodford, N.; Hartman, D. Rapid DNA from a disaster victim identification perspective: Is it a game changer? Forensic Sci. Int. Genet. 2022, 58, 102684. [Google Scholar] [CrossRef]
- Turingan, R.S.; Vasantgadkar, S.; Palombo, L.; Hogan, C.; Jiang, H.; Tan, E.; Selden, R.F. Rapid DNA analysis for automated processing and interpretation of low DNA content samples. Investig. Genet. 2016, 7, 2. [Google Scholar] [CrossRef]
- Foley, M.M. Rapid DNA Profile Development with Applied Biosystems RapidHIT™ ID System. Methods Mol. Biol. 2023, 2685, 367–396. [Google Scholar]
- Turingan, R.S.; Tan, E.; Jiang, H.; Brown, J.; Estari, Y.; Krautz-Peterson, G.; Selden, R.F. Developmental Validation of the ANDE 6C System for Rapid DNA Analysis of Forensic Casework and DVI Samples. J. Forensic Sci. 2020, 65, 1056–1071. [Google Scholar] [CrossRef]
- Gupta, G.Y.; Gupta, Y.A. From Genes to Justice: Ethical Dilemmas and Scientific Advancements in DNA Profiling for Legal Cases. Int. J. Law Manag. Humanit. 2024, 7, 125–140. [Google Scholar]
- Ogden, R.; Dawnay, N.; McEwing, R. Wildlife DNA Forensics—Bridging the Gap Between Conservation Genetics and Law Enforcement. Endanger. Species Res. 2009, 9, 179–195. [Google Scholar] [CrossRef]
- Moore, M.; Baker, B.; Bauman, T.; Curtis, M.; Espinoza, E.; Ferrell, C.; Frankham, G.; Frazier, K.; Giles, J.; Hawk, D.; et al. The Society for Wildlife Forensic Science standards and guidelines. Forensic Sci. Int. Anim. Environ. 2021, 1, 100015. [Google Scholar] [CrossRef]
- Johnson, R.N.; Wilson-Wilde, L.; Linacre, A. Current and future directions of DNA in wildlife forensic science. Forensic Sci. Int. Genet. 2014, 10, 1–11. [Google Scholar] [CrossRef]
- Parson, W.; Pegoraro, K.; Niederstätter, H.; Föger, M.; Steinlechner, M. Species identification by means of the cytochrome b gene. Int. J. Leg. Med. 2000, 114, 23–28. [Google Scholar] [CrossRef]
- Parson, W.; Brandstätter, A.; Alonso, A.; Brandt, N.; Brinkmann, B.; Carracedo, A.; Corach, D.; Froment, O.; Furac, I.; Grzybowski, T.; et al. The EDNAP mitochondrial DNA population database (EMPOP) collaborative exercises: Organisation, results and perspectives. Forensic Sci. Int. 2004, 139, 215–226. [Google Scholar] [CrossRef]
- Vasiljevic, N.; Lim, M.; Humble, E.; Seah, A.; Kratzer, A.; Morf, N.V.; Prost, S.; Ogden, R. Developmental validation of Oxford Nanopore Technology MinION sequence data and the NGSpeciesID bioinformatic pipeline for forensic genetic species identification. Forensic Sci. Int. Genet. 2021, 53, 102493. [Google Scholar] [CrossRef]
- Ogden, R.; Vasiljevic, N.; Prost, S. Nanopore sequencing in non-human forensic genetics. Emerg. Top. Life Sci. 2021, 5, 465–473. [Google Scholar]
- Ogden, R.; Linacre, A. Wildlife forensic science: A review of genetic geographic origin assignment. Forensic Sci. Int. Genet. 2015, 18, 152–159. [Google Scholar] [CrossRef]
- Berger, B.; Berger, C.; Hecht, W.; Hellmann, A.; Rohleder, U.; Schleenbecker, U.; Parson, W. Validation of two canine STR multiplex-assays following the ISFG recommendations for non-human DNA analysis. Forensic Sci. Int. Genet. 2014, 8, 90–100. [Google Scholar] [CrossRef]
- Morf, N.; Kopps, A.; Nater, A.; Lendvay, B.; Vasiljevic, N.; Webster, L.; Fautley, R.; Ogden, R.; Kratzer, A. STRoe deer: A validated forensic STR profiling system for the European roe deer (Capreolus Capreolus). Forensic Sci. Int. Anim. Environ. 2021, 1, 100023. [Google Scholar] [CrossRef]
- Dormontt, E.E.; Jardine, D.I.; van Dijk, K.J.; Dunker, B.F.; Dixon, R.R.M.; Hipkins, V.D.; Tobe, S.; Linacre, A.; Lowe, A.J. Forensic validation of a SNP and INDEL panel for individualisation of timber from bigleaf maple (Acer macrophyllum Pursch). Forensic Sci. Int. Genet. 2020, 46, 102252. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, L.; Gooch, J.; Wolff, K.; Daniel, B.; Frascione, N. Fingermarks in wildlife forensics: A review. Forensic Sci. Int. 2023, 350, 111781. [Google Scholar] [CrossRef]
- Baxter, J.R.; Kotze, A.; de Bruyn, M.; Matlou, K.; Labuschagne, K.; Mwale, M. DNA barcoding of southern African mammal species and construction of a reference library for forensic application. Genome 2024, 67, 378–391. [Google Scholar] [CrossRef]
- ANSI/ASB Standard 180; Standard for the Selection and Evaluation of GenBank® Results for Taxonomic Assignment of Wildlife, 1st ed. ASB Academy Standard Board: Colorado Springs, CO, USA, 2024.
- Frankham, G.J.; Ogden, R.; Baker, B.W.; Ewart, K.M.; Johnson, R.N.; Kuiper, I.; Lindquist, C.D.; Moore, M.K.; Ndiaye, A. Webster LMI: Standards in wildlife forensic science, with a focus on non-human DNA analysis. Anim. Genet. 2025, 56, e70005. [Google Scholar] [CrossRef]
- García-Aceves, M.E.; Romero Rentería, O.; Díaz-Navarro, X.X.; Rangel-Villalobos, H. Paternity tests in Mexico: Results obtained in 3005 cases. J. Forensic Leg. Med. 2018, 55, 1–7. [Google Scholar] [CrossRef]
- Turrina, S.; Bortoletto, E.; Giannini, G.; De Leo, D. Monozygotic twins: Identical or distinguishable for science and law? Med. Sci. law 2021, 61, 62–66. [Google Scholar] [CrossRef]
- Chen, A.; Tao, R.; Li, C.; Zhang, S. Investigation on the genetic-inconsistent paternity cases using the MiSeq FGx system. Forensic Sci. Res. 2022, 7, 702–707. [Google Scholar] [CrossRef]
- Gettings, K.B.; Bodner, M.; Borsuk, L.A.; King, J.L.; Ballard, D.; Parson, W.; Benschop, C.C.G.; Børsting, C.; Budowle, B.; Butler, J.M.; et al. Recommendations of the DNA Commission of the International Society for Forensic Genetics (ISFG) on short tandem repeat sequence nomenclature. Forensic Sci. Int. Genet. 2024, 68, 102946. [Google Scholar] [CrossRef]
- Churchill, J.D.; Schmedes, S.E.; King, J.L.; Budowle, B. Evaluation of the Illumina (R) beta version ForenSeqTMDNA signature prep kit for use in genetic profiling. Forensic Sci. Int. Genet. 2016, 20, 20–29. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, D.; Wang, H.; Jia, Z.; Liu, J.; Qian, X.; Li, C.; Hou, Y. Massively parallel sequencing of 32 forensic markers using the Precision ID GlobalFiler™ NGS STR Panel and the Ion PGM™ System. Forensic Sci. Int. Genet. 2017, 31, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Montano, E.A.; Bush, J.M.; Garver, A.M.; Larijani, M.M.; Wiechman, S.M.; Baker, C.H.; Wilson, M.R.; Guerrieri, R.A.; Benzinger, E.A.; Gehres, D.N.; et al. Optimization of the Promega PowerSeq™ Auto/Y system for efficient integration within a forensic DNA laboratory. Forensic Sci. Int. Genet. 2018, 32, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Riman, S.; Iyer, H.; Borsuk, L.A.; Vallone, P.M. Understanding the characteristics of sequence-based single-source DNA profiles. Forensic Sci. Int. Genet. 2020, 44, 102192. [Google Scholar] [CrossRef] [PubMed]
- Connell, J.R.; Benton, M.C.; Lea, R.A.; Sutherland, H.G.; Haupt, L.M.; Wright, K.M.; Griffiths, L.R. Evaluating the suitability of current mitochondrial DNA interpretation guidelines for multigenerational whole mitochondrial genome comparisons. J. Forensic Sci. 2022, 67, 1766–1775. [Google Scholar] [CrossRef]
- Liu, G.; Zheng, Y.; Wu, Q.; Feng, T.; Xia, Y.; Chen, D.; Ren, L.; Bai, X.; Li, Q.; Chen, D.; et al. Assessment of ForenSeq mtDNA Whole Genome Kit for forensic application. Int. J. Leg. Med. 2023, 137, 1693–1703. [Google Scholar] [CrossRef]
- Verogen, Inc. ForenSeq Imagen Kit Reference Guide [Internet]. San Diego, CA: Verogen; 2022 [Revised 15th April 2025]. Available online: https://verogen.com/wp-content/uploads/2022/08/forenseq-imagen-reference-guide-PCR1-vd2022008-a.pdf (accessed on 15 April 2025).
- Garneau, J.E.; Dupuis, M.-È.; Villion, M.; Romero, D.A.; Barrangou, R.; Boyaval, P.; Fremaux, C.; Horvath, P.; Magadán, A.H.; Moineau, S. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 2010, 468, 67–71. [Google Scholar] [CrossRef]
- Dash, H.R.; Arora, M. CRISPR-CasB technology in forensic DNA analysis: Challenges and solutions. Appl. Microbiol. Biotechnol. 2022, 106, 4367–4374. [Google Scholar] [CrossRef]
- Sobral, A.F.; Dinis-Oliveira, R.J.; Barbosa, D.J. CRISPR-Cas technology in forensic investigations: Principles, applications, and ethical considerations. Forensic Sci. Int. Genet. 2025, 74, 103163. [Google Scholar] [CrossRef]
- Barash, M.; McNevin, D.; Fedorenko, V.; Giverts, P. Machine learning applications in forensic DNA profiling: A critical review. Forensic Sci. Int. Genet. 2024, 69, 102994. [Google Scholar] [CrossRef]
- Afkanpour, M.; Momeni, M.; Tabrizi, A.A.; Tabesh, H. A haplogroup-based methodology for assigning individuals to geographical regions using Y-STR data. Forensic Sci. Int. 2024, 365, 112260. [Google Scholar] [CrossRef]
- Sessa, F.; Esposito, M.; Cocimano, G.; Sablone, S.; Karaboue, M.A.A.; Chisari, M.; Albano, D.G.; Salerno, M. Artificial Intelligence and Forensic Genetics: Current Applications and Future Perspectives. Appl. Sci. 2024, 14, 2113. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Source | STRs and Other Markers Included | Reference |
|---|---|---|---|
| AmpFlSTR® COfiler™ | Applied Biosystems | D3S1358, D16S539, Amelogenin, TH01, TPOX, CSF1PO, D7S820 | [59] |
| AmpFlSTR® Profiler™ | Applied Biosystems | D3S1358, vWA, FGA, Amelogenin, TH01, TPOX, CSF1PO, D5S818, D13S317, D7S820 | [59] |
| AmpFISTR® Identifiler™ | Applied Biosystems | D8S1179, D21S11, D7S820, CSF1PO, D3S1358, TH01, D13S317, D16S539, D2S1338, D19S433, vWA, TPOX, D18S51, Amelogenin, D5S818, FGA | [60] |
| GlobalFiler™ | Applied Biosystems | D13S317, D7S820, D5S818, CSF1PO, D1S1656, D12S391, D2S441, D10S1248, D18S51, FGA, D21S11, D8S1179, vWA, D16S539, TH01, D3S1358, AMEL, D2S1338, D19S433, DYS391, TPOX, D22S1045, SE33 and a Y-specific insertion/deletion locus (Y-indel). | [61] |
| Investigator IDplex Plus Kit | QIAGEN | CSF1PO, FGA, TH01, TPOX, VWA, D3S1358, D5S818, D7S820, D8S1179, D13S317, D16S539, D18S51, and D21S1, D2S1338, D19S433 and Amelogenin. | [62] |
| PowerPlex® 1.2 | Promega | D16S539, D7S820, D13S317, D5S818, CSF1PO, TPOX, TH01, vWA. | [63] |
| PowerPlex® 16 Monoplex System | Promega | D3S1358, TH01, D21S11, D18S51, Penta E, D5S818, D13S317, D7S820, D16S539, CSF1PO, Penta D, Amelogenin, vWA, D8S1179, TPOX, FGA. | [64] |
| The PowerPlex(®) 21 System | Promega | D1S1656, D2S1338, D3S1358, D5S818, D6S1043, D7S820, D8S1179, D12S391, D13S317, D16S539, D18S51, D19S433, D21S11, Amelogenin, CSF1PO, FGA, Penta D, Penta E, TH01, TPOX, and vWA. | [65] |
| PowerPlex® fusion 5C | Promega | Amelogenin, D3S1358, D1S1656, D2S441, D10S1248, D13S317, Penta E, D16S539, D18S51, D2S1338, CSF1PO, Penta D, TH01, vWA, D21S11, D7S820, D5S818, TPOX, DYS391, D8S1179, D12S391, D19S433, FGA, and D22S1045. | [66] |
| PowerPlex® fusion 6C | Promega | CSF1PO, FGA, TH01, TPOX, vWA, D1S1656, D2S1338, D2S441, D3S1358, D5S818, D7S820, D8S1179, D10S1248, D12S391, D13S317, D16S539, D18S51, D19S433, D21S11 and D22S1045, Amelogenin and DYS391 for gender determination, Penta D, Penta E and SE33. | [67] |
| VersaPlex™ 27PY | Applied Biosystems | CSF1PO, FGA, TH01, TPOX, vWA, D1S1656, D2S1338, D2S441, D3S1358, D5S818, D7S820, D8S1179, D10S1248, D12S391, D13S317, D16S539, D18S51, D19S433, D21S11, D22S1045, Amelogenin, DYS391, Penta D, Penta E, D6S1043, DYS570 and DYS576. | [68] |
| Investigator 24plex QS | QIAGEN | D1S1656, D2S441, D2S1338, D3S1358, D5S818, D7S820, D8S1179, D10S1248, D12S391, D13S317, D16S539, D18S51, D19S433, D21S11, D22S1045, CSF1PO, FGA, TH01, TPOX, vWA, SE33, DYS391, Amelogenin and the Quality Sensor. | [69] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Hurtado, I.A.; García-Acéves, M.E.; Puga-Carrillo, Y.; Guardado-Estrada, M.; Becerra-Loaiza, D.S.; Carrillo-Rodríguez, V.D.; Plazola-Zamora, R.; Godínez-Rubí, J.M.; Rangel-Villalobos, H.; Aguilar-Velázquez, J.A. Past, Present and Future Perspectives of Forensic Genetics. Biomolecules 2025, 15, 713. https://doi.org/10.3390/biom15050713
Gutiérrez-Hurtado IA, García-Acéves ME, Puga-Carrillo Y, Guardado-Estrada M, Becerra-Loaiza DS, Carrillo-Rodríguez VD, Plazola-Zamora R, Godínez-Rubí JM, Rangel-Villalobos H, Aguilar-Velázquez JA. Past, Present and Future Perspectives of Forensic Genetics. Biomolecules. 2025; 15(5):713. https://doi.org/10.3390/biom15050713
Chicago/Turabian StyleGutiérrez-Hurtado, Itzae Adonai, Mayra Elizabeth García-Acéves, Yolanda Puga-Carrillo, Mariano Guardado-Estrada, Denisse Stephania Becerra-Loaiza, Víctor Daniel Carrillo-Rodríguez, Reynaldo Plazola-Zamora, Juliana Marisol Godínez-Rubí, Héctor Rangel-Villalobos, and José Alonso Aguilar-Velázquez. 2025. "Past, Present and Future Perspectives of Forensic Genetics" Biomolecules 15, no. 5: 713. https://doi.org/10.3390/biom15050713
APA StyleGutiérrez-Hurtado, I. A., García-Acéves, M. E., Puga-Carrillo, Y., Guardado-Estrada, M., Becerra-Loaiza, D. S., Carrillo-Rodríguez, V. D., Plazola-Zamora, R., Godínez-Rubí, J. M., Rangel-Villalobos, H., & Aguilar-Velázquez, J. A. (2025). Past, Present and Future Perspectives of Forensic Genetics. Biomolecules, 15(5), 713. https://doi.org/10.3390/biom15050713