
Identification of Critical Molecular Pathways Induced by HDAC11 Overexpression in Cardiac Mesenchymal Stem Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Mouse CMSCs
2.2. Adenovirus Transduction
2.3. RNA Extraction, Next-Generation Sequencing, and Bioinformatic Analysis
2.4. Real Time qRT-PCR
| hHDAC11: |
| F primer: AGGGCTACCATCATTGATCTTG |
| R primer: GTAGATGTGGCGGTTGTAGAC |
| SELP: |
| F primer: AGTTTCCGGTTCCCAGTAAAG |
| R primer: CCAGTAGCCAGGCATCTTATC |
| FABP3: |
| F primer: GACCAAGCCTACTACCATCATC |
| R primer: GTCACCTCGTCGAACTCTATTC |
| GM11266: |
| F primer: GTAATGAAGCCAACCAGAAGAAAG |
| R primer: CCAAGTCCAGGTAACCAGTATG |
| GM16548: |
| F primer: GAGCATTGTGGTTTGCTTTGA |
| R primer: CTGTGTGTGACTCTTGGGTATG |
| GAPDH: |
| F primer: CAAATGGCAGCCCTGGT |
| R primer: CCTCGTCCCGTAGACAAA |
2.5. Western Blot
2.6. Measuring Nuclear Size Using Fluorescent Staining
2.7. Statistical Analysis
3. Results
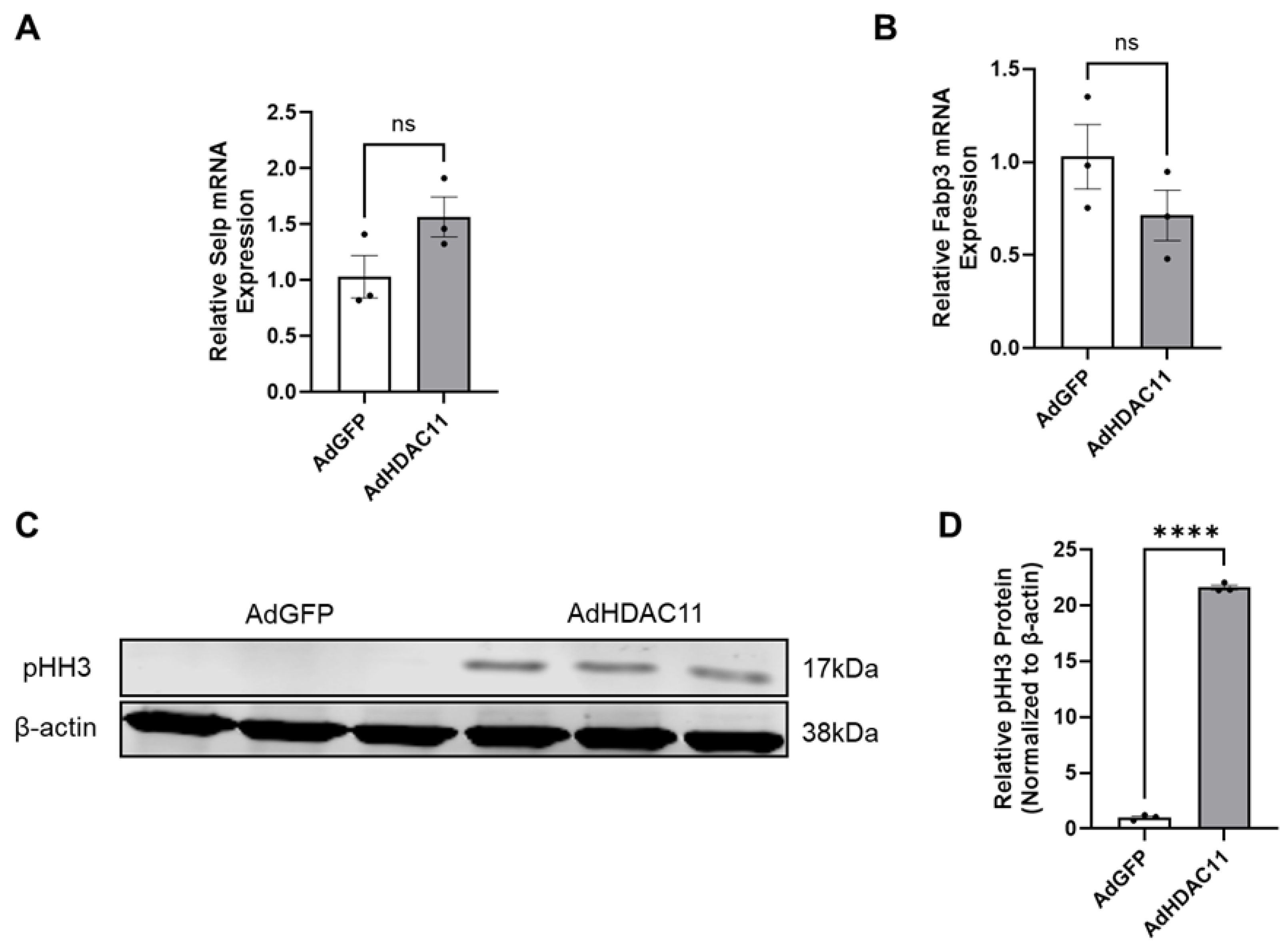
3.1. Confirmation of HDAC11 Overexpression in CMSCs
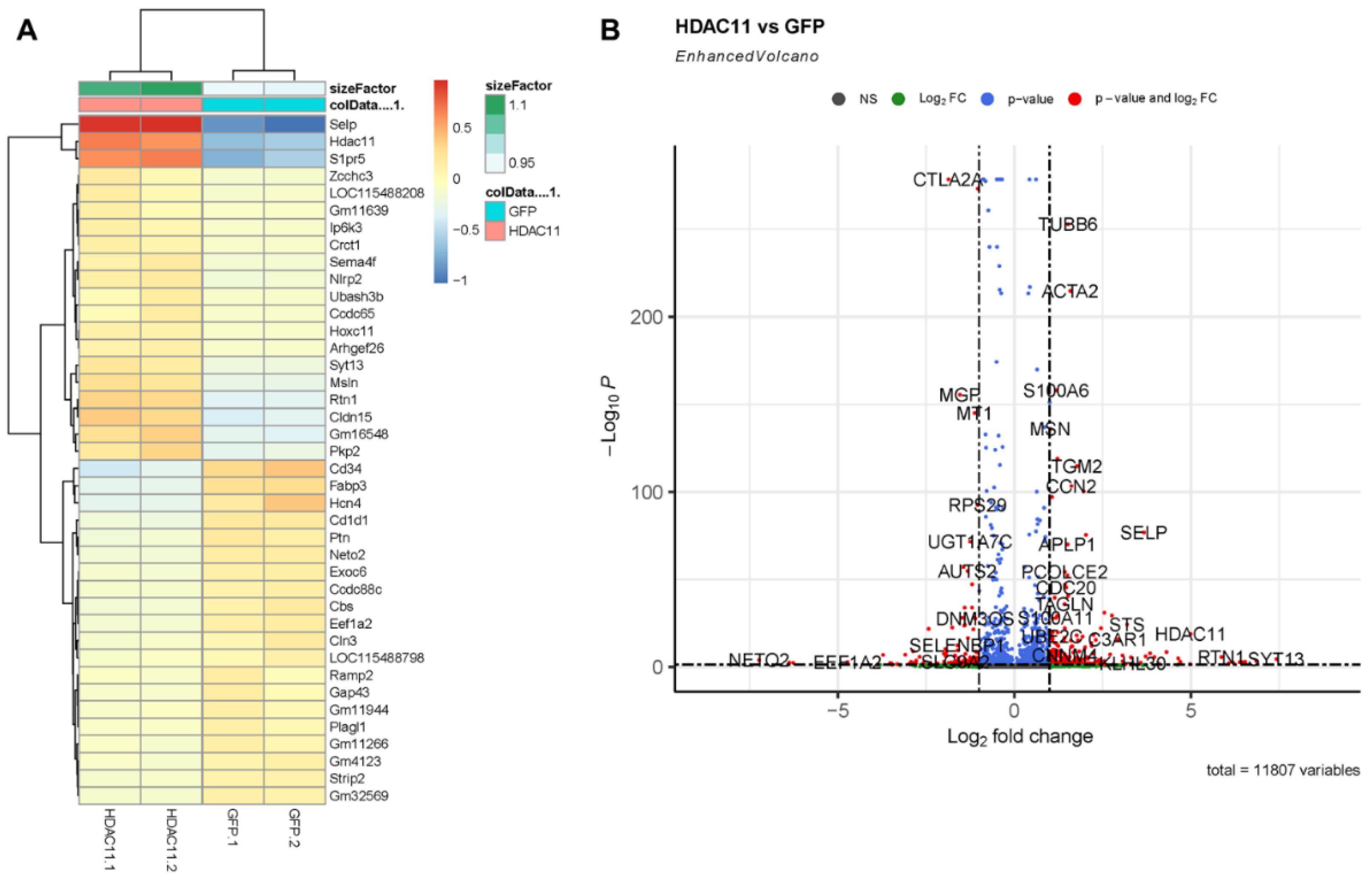
3.2. Differential Gene Expression and Functional Enrichment Analysis of HDAC11-Overexpressing Versus Control CMSCs
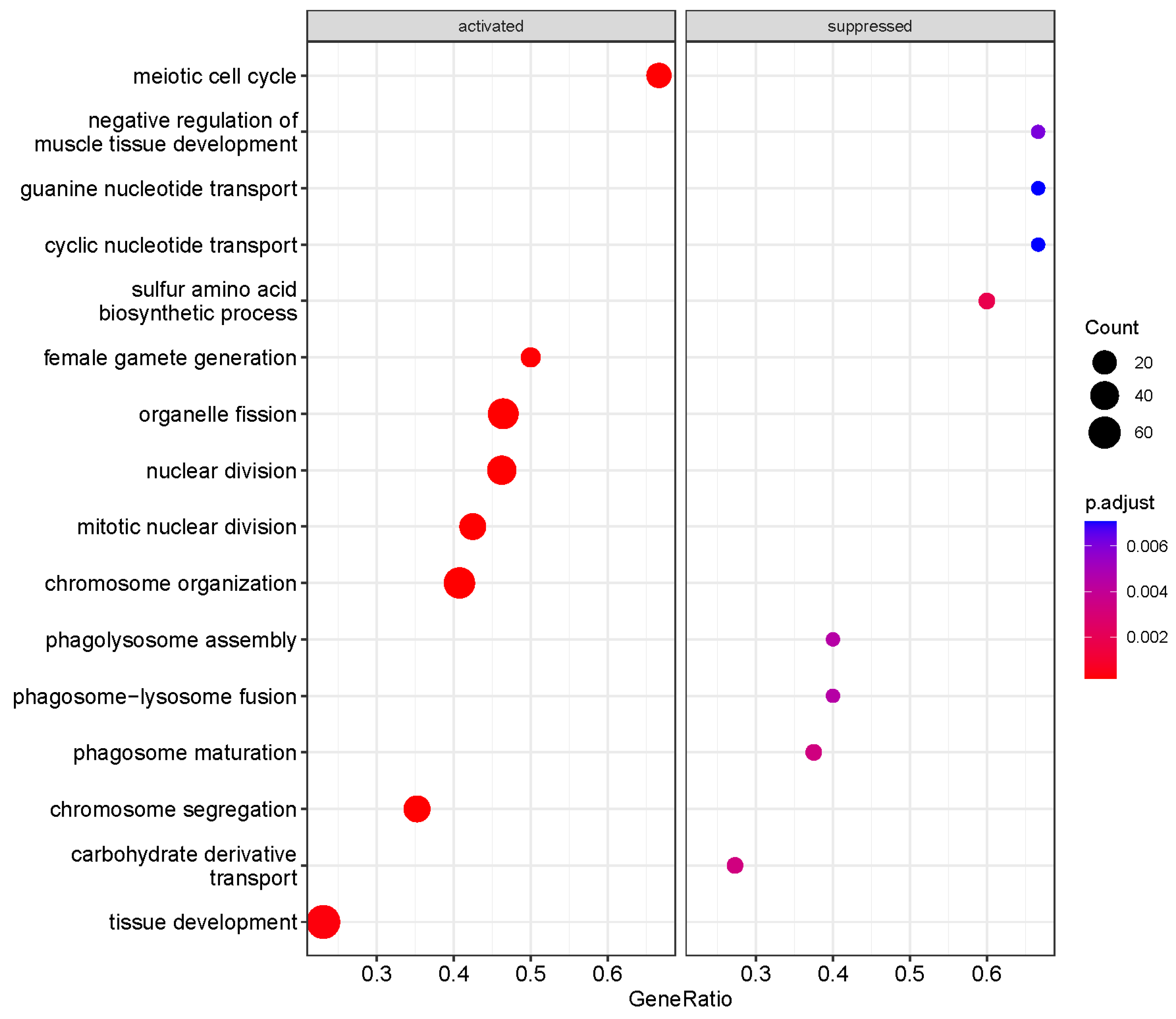
3.3. Functional Enrichment Analysis of GO Biological Processes in HDAC11-Overexpressing CMSCs
3.4. Chromosome Organization Is a Major Activated Pathway in HDAC11-Overexpressing CMSCs
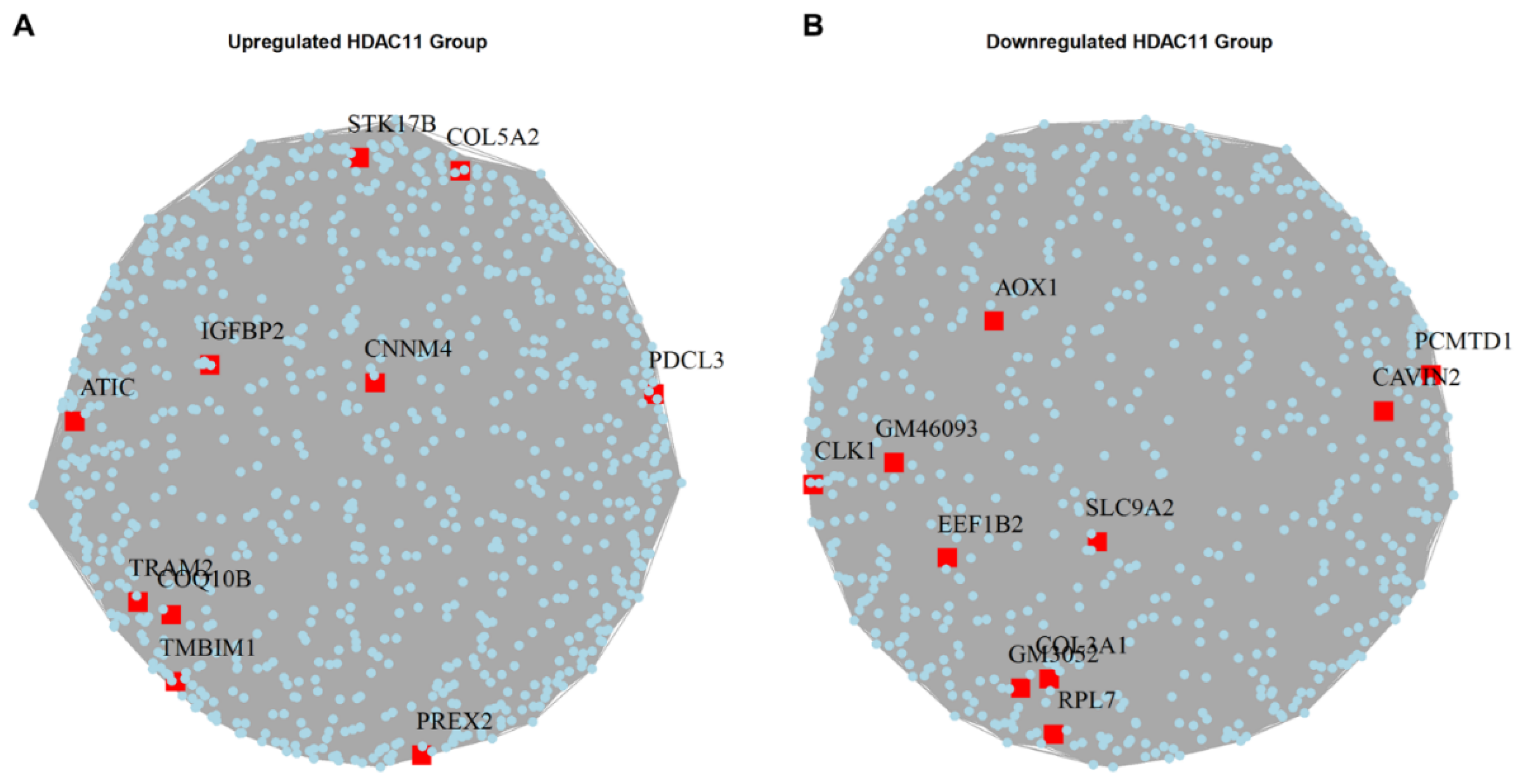
3.5. Identification of Hub Genes in HDAC11-Overexpressing CMSCs
3.6. The Effects of HDAC11 Overexpression on CMSC Proliferation
3.7. HDAC11-Targeted Long Non-Coding RNAs (lncRNAs)
4. Discussion
5. Conclusions
6. Future Perspective
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seto, E.; Yoshida, M. Erasers of histone acetylation: The histone deacetylase enzymes. Cold Spring Harb. Perspect. Biol. 2014, 6, a018713. [Google Scholar] [CrossRef] [PubMed]
- Shahbazian, M.D.; Grunstein, M. Functions of site-specific histone acetylation and deacetylation. Annu. Rev. Biochem. 2007, 76, 75–100. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Cueto, M.A.; Asselbergs, F.; Atadja, P. Cloning and functional characterization of HDAC11, a novel member of the human histone deacetylase family. J. Biol. Chem. 2002, 277, 25748–25755. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Sun, L.; Aramsangtienchai, P.; Spiegelman, N.A.; Zhang, X.; Huang, W.; Seto, E.; Lin, H. HDAC11 regulates type I interferon signaling through defatty-acylation of SHMT2. Proc. Natl. Acad. Sci. USA 2019, 116, 5487–5492. [Google Scholar] [CrossRef]
- Bi, L.; Ren, Y.; Feng, M.; Meng, P.; Wang, Q.; Chen, W.; Jiao, Q.; Wang, Y.; Du, L.; Zhou, F.; et al. HDAC11 Regulates Glycolysis through the LKB1/AMPK Signaling Pathway to Maintain Hepatocellular Carcinoma Stemness. Cancer Res. 2021, 81, 2015–2028. [Google Scholar] [CrossRef]
- Kutil, Z.; Novakova, Z.; Meleshin, M.; Mikesova, J.; Schutkowski, M.; Barinka, C. Histone Deacetylase 11 Is a Fatty-Acid Deacylase. ACS Chem. Biol. 2018, 13, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, R.A.; Robinson, E.L.; Hu, T.; Cao, J.; Hong, J.Y.; Tharp, C.A.; Qasim, H.; Gavin, K.M.; da Silva, J.P.; Major, J.L.; et al. Reversible lysine fatty acylation of an anchoring protein mediates adipocyte adrenergic signaling. Proc. Natl. Acad. Sci. USA 2022, 119, e2119678119. [Google Scholar] [CrossRef]
- Bagchi, R.A.; Ferguson, B.S.; Stratton, M.S.; Hu, T.; Cavasin, M.A.; Sun, L.; Lin, Y.; Liu, D.; Londono, P.; Song, K.; et al. HDAC11 suppresses the thermogenic program of adipose tissue via BRD2. JCI Insight. 2018, 3, e120159. [Google Scholar] [CrossRef]
- Yanginlar, C.; Logie, C. HDAC11 is a regulator of diverse immune functions. Biochim. Biophys. Acta Gene Regul. Mech. 2018, 1861, 54–59. [Google Scholar] [CrossRef]
- Glozak, M.A.; Seto, E. Acetylation/deacetylation modulates the stability of DNA replication licensing factor Cdt1. J. Biol. Chem. 2009, 284, 11446–11453. [Google Scholar] [CrossRef]
- Blixt, N.C.; Faulkner, B.K.; Astleford, K.; Lelich, R.; Schering, J.; Spencer, E.; Gopalakrishnan, R.; Jensen, E.D.; Mansky, K.C. Class II and IV HDACs function as inhibitors of osteoclast differentiation. PLoS ONE 2017, 12, e0185441. [Google Scholar] [CrossRef] [PubMed]
- Bhaskara, S. Histone deacetylase 11 as a key regulator of metabolism and obesity. eBioMedicine 2018, 35, 27–28. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Marin de Evsikova, C.; Bian, K.; Achille, A.; Telles, E.; Pei, H.; Seto, E. Programming and Regulation of Metabolic Homeostasis by HDAC11. eBioMedicine 2018, 33, 157–168. [Google Scholar] [CrossRef]
- Liu, Y.; Tong, X.; Hu, W.; Chen, D. HDAC11: A novel target for improved cancer therapy. Biomed. Pharmacother. 2023, 166, 115418. [Google Scholar] [CrossRef]
- Kim, J.I.; Jung, K.J.; Jang, H.S.; Park, K.M. Gender-specific role of HDAC11 in kidney ischemia- and reperfusion-induced PAI-1 expression and injury. Am. J. Physiol. Renal Physiol. 2013, 305, F61–F70. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, X.M.; Xiao, S.; Wu, Z.R.; Shi, Y.J.; Xie, M.J. HDAC11 is related to breast cancer prognosis and inhibits invasion and proliferation of breast cancer cells. Int. J. Clin. Exp. Pathol. 2023, 16, 172–183. [Google Scholar]
- Bagui, T.K.; Sharma, S.S.; Ma, L.; Pledger, W.J. Proliferative status regulates HDAC11 mRNA abundance in nontransformed fibroblasts. Cell Cycle 2013, 12, 3433–3441. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zibetti, C.; Shang, P.; Sripathi, S.R.; Zhang, P.; Cano, M.; Hoang, T.; Xia, S.; Ji, H.; Merbs, S.L.; et al. ATAC-Seq analysis reveals a widespread decrease of chromatin accessibility in age-related macular degeneration. Nat. Commun. 2018, 9, 1364. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhao, K.; Yao, Y.; Liu, C.; Chen, Y.; Li, J.; Wang, Y.; Pei, R.; Chen, J.; Hu, X.; et al. HDAC11 restricts HBV replication through epigenetic repression of cccDNA transcription. Antivir. Res. 2019, 172, 104619. [Google Scholar] [CrossRef]
- Zhang, R.; Pan, Y.; Feng, W.; Zhao, Y.; Yang, Y.; Wang, L.; Zhang, Y.; Cheng, J.; Jiang, Q.; Zheng, Z.; et al. HDAC11 Regulates the Proliferation of Bovine Muscle Stem Cells through the Notch Signaling Pathway and Inhibits Muscle Regeneration. J. Agric. Food Chem. 2022, 70, 9166–9178. [Google Scholar] [CrossRef]
- Nunez-Alvarez, Y.; Hurtado, E.; Munoz, M.; Garcia-Tunon, I.; Rech, G.E.; Pluvinet, R.; Sumoy, L.; Pendas, A.M.; Peinado, M.A.; Suelves, M. Loss of HDAC11 accelerates skeletal muscle regeneration in mice. FEBS J. 2021, 288, 1201–1223. [Google Scholar] [CrossRef]
- Yang, H.; Li, C.; Che, M.; Liang, J.; Tian, X.; Yang, G.; Sun, C. HDAC11 deficiency resists obesity by converting adipose-derived stem cells into brown adipocyte-like cells. Int. J. Biol. Macromol. 2024, 258 Pt 1, 128852. [Google Scholar] [CrossRef]
- Villagra, A.; Cheng, F.; Wang, H.W.; Suarez, I.; Glozak, M.; Maurin, M.; Nguyen, D.; Wright, K.L.; Atadja, P.W.; Bhalla, K.; et al. The histone deacetylase HDAC11 regulates the expression of interleukin 10 and immune tolerance. Nat. Immunol. 2009, 10, 92–100. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Y.; Pan, Y.; Zhang, L.; Shen, C.; Qin, G.; Ashraf, M.; Weintraub, N.; Tang, Y. Cardiac progenitor-derived exosomes protect ischemic myocardium from acute ischemia/reperfusion injury. Biochem. Biophys. Res. Commun. 2013, 431, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Zhao, Q.; Zhang, Y.C.; Cheng, L.; Liu, M.; Shi, J.; Yang, Y.Z.; Pan, C.; Ge, J.; Phillips, M.I. Autologous mesenchymal stem cell transplantation induce VEGF and neovascularization in ischemic myocardium. Regul. Pept. 2004, 117, 3–10. [Google Scholar] [CrossRef]
- Tang, Y.L.; Zhu, W.; Cheng, M.; Chen, L.; Zhang, J.; Sun, T.; Kishore, R.; Phillips, M.I.; Losordo, D.W.; Qin, G. Hypoxic preconditioning enhances the benefit of cardiac progenitor cell therapy for treatment of myocardial infarction by inducing CXCR4 expression. Circ. Res. 2009, 104, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Ju, C.; Shen, Y.; Ma, G.; Liu, Y.; Cai, J.; Kim, I.M.; Weintraub, N.L.; Liu, N.; Tang, Y. Transplantation of Cardiac Mesenchymal Stem Cell-Derived Exosomes Promotes Repair in Ischemic Myocardium. J. Cardiovasc. Transl. Res. 2018, 11, 420–428. [Google Scholar] [CrossRef]
- Mouquet, F.; Pfister, O.; Jain, M.; Oikonomopoulos, A.; Ngoy, S.; Summer, R.; Fine, A.; Liao, R. Restoration of cardiac progenitor cells after myocardial infarction by self-proliferation and selective homing of bone marrow-derived stem cells. Circ. Res. 2005, 97, 1090–1092. [Google Scholar] [CrossRef]
- Su, X.; Jin, Y.; Shen, Y.; Kim, I.M.; Weintraub, N.L.; Tang, Y. RNAase III-Type Enzyme Dicer Regulates Mitochondrial Fatty Acid Oxidative Metabolism in Cardiac Mesenchymal Stem Cells. Int. J. Mol. Sci. 2019, 20, 5554. [Google Scholar] [CrossRef]
- Chen, L.; Pan, Y.; Zhang, L.; Wang, Y.; Weintraub, N.; Tang, Y. Two-step protocol for isolation and culture of cardiospheres. Methods Mol Biol. 2013, 1036, 75–80. [Google Scholar]
- Chen, L.; Ashraf, M.; Wang, Y.; Zhou, M.; Zhang, J.; Qin, G.; Rubinstein, J.; Weintraub, N.L.; Tang, Y. The role of notch 1 activation in cardiosphere derived cell differentiation. Stem Cells Dev. 2012, 21, 2122–2129. [Google Scholar]
- Zhang, H.; Shen, Y.; Kim, I.M.; Liu, Y.; Cai, J.; Berman, A.E.; Nilsson, K.R.; Weintraub, N.L.; Tang, Y. Electrical Stimulation Increases the Secretion of Cardioprotective Extracellular Vesicles from Cardiac Mesenchymal Stem Cells. Cells 2023, 12, 875. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Dekker, B.; Liu, Z.; Cabrera, H.; Ryan, J.; Dekker, J.; Heald, R. Mitotic chromosomes scale to nuclear-cytoplasmic ratio and cell size in Xenopus. eLife 2023, 12, e84360. [Google Scholar] [CrossRef]
- Murphy, C.M.; Michael, W.M. Control of DNA replication by the nucleus/cytoplasm ratio in Xenopus. J. Biol. Chem. 2013, 288, 29382–29393. [Google Scholar] [CrossRef]
- Tang, A.; Gao, K.; Chu, L.; Zhang, R.; Yang, J.; Zheng, J. Aurora kinases: Novel therapy targets in cancers. Oncotarget 2017, 8, 23937–23954. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Magana, A.; Blanco, F.J. Human PCNA Structure, Function and Interactions. Biomolecules 2020, 10, 570. [Google Scholar] [CrossRef] [PubMed]
- Gerhold, C.B.; Hauer, M.H.; Gasser, S.M. INO80-C and SWR-C: Guardians of the genome. J. Mol. Biol. 2015, 427, 637–651. [Google Scholar]
- Jiang, Y.; Mu, H.; Zhao, H. HMBOX1, a member of the homeobox family: Current research progress. Cent. Eur. J. Immunol. 2023, 48, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Csárdi, G.; Nepusz, T. The Igraph Software Package for Complex Network Research. InterJournal Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Wu, Q.W.; Kapfhammer, J.P. The Emerging Key Role of the mGluR1-PKCgamma Signaling Pathway in the Pathogenesis of Spinocerebellar Ataxias: A Neurodevelopmental Viewpoint. Int. J. Mol. Sci. 2022, 23, 9169. [Google Scholar] [CrossRef]
- Malfait, F.; Wenstrup, R.J.; De Paepe, A. Clinical and genetic aspects of Ehlers-Danlos syndrome, classic type. Genet. Med. 2010, 12, 597–605. [Google Scholar] [CrossRef]
- Li, T.; Forbes, M.E.; Fuller, G.N.; Li, J.; Yang, X.; Zhang, W. IGFBP2: Integrative hub of developmental and oncogenic signaling network. Oncogene 2020, 39, 2243–2257. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, D.; Funato, Y.; Miura, J.; Sato, S.; Toyosawa, S.; Furutani, K.; Kurachi, Y.; Omori, Y.; Furukawa, T.; Tsuda, T.; et al. Basolateral Mg2+ extrusion via CNNM4 mediates transcellular Mg2+ transport across epithelia: A mouse model. PLoS Genet. 2013, 9, e1003983. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Meyer, R.D.; Lugo, R.; Rahimi, N. Identification of PDCL3 as a novel chaperone protein involved in the generation of functional VEGF receptor 2. J. Biol. Chem. 2013, 288, 23171–23181. [Google Scholar] [CrossRef]
- Liu, X.; Paila, U.D.; Teraoka, S.N.; Wright, J.A.; Huang, X.; Quinlan, A.R.; Gatti, R.A.; Concannon, P. Identification of ATIC as a Novel Target for Chemoradiosensitization. Int. J. Radiat. Oncol. Biol. Phys. 2018, 100, 162–173. [Google Scholar] [CrossRef]
- Li, L.; Ugalde, A.P.; Scheele, C.L.G.J.; Dieter, S.M.; Nagel, R.; Ma, J.; Pataskar, A.; Korkmaz, G.; Elkon, R.; Chien, M.; et al. A comprehensive enhancer screen identifies TRAM2 as a key and novel mediator of YAP oncogenesis. Genome Biol. 2021, 22, 54. [Google Scholar] [CrossRef]
- Sood, B.; Patel, P.; Keenaghan, M. Coenzyme Q10; StatPearls: Treasure Island, FL, USA, 2025. [Google Scholar]
- Zhao, G.N.; Zhang, P.; Gong, J.; Zhang, X.J.; Wang, P.X.; Yin, M.; Jiang, Z.; Shen, L.; Ji, Y.; Tong, J.; et al. Tmbim1 is a multivesicular body regulator that protects against non-alcoholic fatty liver disease in mice and monkeys by targeting the lysosomal degradation of Tlr4. Nat. Med. 2017, 23, 742–752. [Google Scholar] [CrossRef]
- Li, M.; Xiao, J.; Song, S.; Han, F.; Liu, H.; Lin, Y.; Ni, Y.; Zeng, S.; Zou, X.; Wu, J.; et al. PREX2 contributes to radiation resistance by inhibiting radiotherapy-induced tumor immunogenicity via cGAS/STING/IFNs pathway in colorectal cancer. BMC Med. 2024, 22, 154. [Google Scholar] [CrossRef]
- Vantaku, V.; Putluri, V.; Bader, D.A.; Maity, S.; Ma, J.; Arnold, J.M.; Rajapakshe, K.; Donepudi, S.R.; von Rundstedt, F.; Devarakonda, V.; et al. Epigenetic loss of AOX1 expression via EZH2 leads to metabolic deregulations and promotes bladder cancer progression. Oncogene 2020, 39, 6265–6285. [Google Scholar] [CrossRef]
- Warmack, R.A.; Pang, E.Z.; Peluso, E.; Lowenson, J.D.; Ong, J.Y.; Torres, J.Z.; Clarke, S.G. Human Protein-l-isoaspartate O-Methyltransferase Domain-Containing Protein 1 (PCMTD1) Associates with Cullin-RING Ligase Proteins. Biochemistry 2022, 61, 879–894. [Google Scholar] [CrossRef]
- Peng, M.; Ye, L.; Yang, L.; Liu, X.; Chen, Y.; Huang, G.; Jiang, Y.; Wang, Y.; Li, D.; He, J.; et al. CAVIN2 is frequently silenced by CpG methylation and sensitizes lung cancer cells to paclitaxel and 5-FU. Epigenomics 2020, 12, 1793–1810. [Google Scholar] [CrossRef] [PubMed]
- Auld, D.; Shen, M.; Thomas, C. A High-Throughput Screen for Pre-mRNA Splicing Modulators; Probe Reports from the NIH Molecular Libraries Program; NIH: Bethesda, MD, USA, 2010. [Google Scholar]
- Huang, Z.; Li, N.; Ji, X.; Zhou, D.; Wang, R.; Zhao, X.; Wang, S.; Zhang, H.; Huang, C.; Lin, G. EEF1B2 regulates the proliferation and apoptosis of human spermatogonial stem cell lines through TAF4B. Heliyon 2024, 10, e36467. [Google Scholar] [CrossRef]
- Han, J.; Li, S.; Cao, J.; Han, H.; Lu, B.; Wen, T.; Bian, W. SLC9A2, suppressing by the transcription suppressor ETS1, restrains growth and invasion of osteosarcoma via inhibition of aerobic glycolysis. Environ. Toxicol. 2024, 39, 238–251. [Google Scholar] [CrossRef]
- Cray, J.J., Jr.; Gallo, P.H.; Durham, E.L.; Losee, J.E.; Mooney, M.P.; Kathju, S.; Cooper, G.M. Molecular analysis of coronal perisutural tissues in a craniosynostotic rabbit model using polymerase chain reaction suppression subtractive hybridization. Plast. Reconstr. Surg. 2011, 128, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Yang, Y.; Fan, X.; Tang, Y.; Tang, Z. Sp1-Mediated circRNA circHipk2 Regulates Myogenesis by Targeting Ribosomal Protein Rpl7. Genes 2021, 12, 696. [Google Scholar] [CrossRef]
- Yabuki, Y.; Fukunaga, K. Clinical Therapeutic Strategy and Neuronal Mechanism Underlying Post-Traumatic Stress Disorder (PTSD). Int. J. Mol. Sci. 2019, 20, 3614. [Google Scholar] [CrossRef]
- Bora-Singhal, N.; Mohankumar, D.; Saha, B.; Colin, C.M.; Lee, J.Y.; Martin, M.W.; Zheng, X.; Coppola, D.; Chellappan, S. Novel HDAC11 inhibitors suppress lung adenocarcinoma stem cell self-renewal and overcome drug resistance by suppressing Sox2. Sci. Rep. 2020, 10, 4722. [Google Scholar] [CrossRef]
- Eto, T.; Winkler, I.; Purton, L.E.; Levesque, J.P. Contrasting effects of P-selectin and E-selectin on the differentiation of murine hematopoietic progenitor cells. Exp. Hematol. 2005, 33, 232–242. [Google Scholar] [CrossRef]
- Xu, G.; Yang, Z.; Sun, Y.; Dong, H.; Ma, J. Interaction of microRNAs with sphingosine kinases, sphingosine-1 phosphate, and sphingosine-1 phosphate receptors in cancer. Discov. Oncol. 2021, 12, 33. [Google Scholar] [CrossRef]
- Chuang, C.Y.; Huang, M.C.; Chen, H.F.; Tseng, L.H.; Yu, C.Y.; Stone, L.; Huang, H.; Ho, H.; Kuo, H. Granulosa cell-derived induced pluripotent stem cells exhibit pro-trophoblastic differentiation potential. Stem Cell Res. Ther. 2015, 6, 14. [Google Scholar] [CrossRef]
- Pérez-Hernández, M.; van Opbergen, C.J.M.; Bagwan, N.; Vissing, C.R.; Marrón-Liñares, G.M.; Zhang, M.; Vega, E.T.; Sorrentino, A.; Drici, L.; Sulek, K.; et al. Loss of Nuclear Envelope Integrity and Increased Oxidant Production Cause DNA Damage in Adult Hearts Deficient in PKP2: A Molecular Substrate of ARVC. Circulation 2022, 146, 851–867. [Google Scholar] [CrossRef] [PubMed]
- Gangenahalli, G.U.; Singh, V.K.; Verma, Y.K.; Gupta, P.; Sharma, R.K.; Chandra, R.; Luthra, P.M. Hematopoietic stem cell antigen CD34: Role in adhesion or homing. Stem Cells Dev. 2006, 15, 305–313. [Google Scholar] [CrossRef]
- Nielsen, J.S.; McNagny, K.M. Novel functions of the CD34 family. J. Cell Sci. 2008, 121 Pt 22, 3683–3692. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, Y.; Andreyev, O.; Hoyt, R.F., Jr.; Singh, A.; Hunt, T.; Horvath, K.A. Overexpression of FABP3 inhibits human bone marrow derived mesenchymal stem cell proliferation but enhances their survival in hypoxia. Exp. Cell Res. 2014, 323, 56–65. [Google Scholar] [CrossRef]
- Saito, Y.; Nakamura, K.; Yoshida, M.; Sugiyama, H.; Akagi, S.; Miyoshi, T.; Morita, H.; Ito, H. Enhancement of pacing function by HCN4 overexpression in human pluripotent stem cell-derived cardiomyocytes. Stem Cell Res. Ther. 2022, 13, 141. [Google Scholar] [CrossRef]
- Zhang, L.; Laaniste, L.; Jiang, Y.; Alafuzoff, I.; Uhrbom, L.; Dimberg, A. Pleiotrophin enhances PDGFB-induced gliomagenesis through increased proliferation of neural progenitor cells. Oncotarget 2016, 7, 80382–80390. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Gao, F.; Zhang, L.; Wang, C.; Hu, L.; Fan, Z.; Xia, D. Pleiotropin enhances the osteo/dentinogenic differentiation potential of dental pulp stem cells. Connect. Tissue Res. 2021, 62, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Zhang, S.; Zhou, Y.; Sun, T.; Zhu, Y.; Fei, J.; Li, A.; Miao, Y. CCDC88C, an O-GalNAc glycosylation substrate of GALNT6, drives breast cancer metastasis by promoting c-JUN-mediated CEMIP transcription. Cancer Cell Int. 2024, 24, 237. [Google Scholar] [CrossRef]
- Khwanraj, K.; Dharmasaroja, P. Neuroblastoma Cell Death Induced by eEF1A2 Knockdown Is Possibly Mediated by the Inhibition of Akt and mTOR Phosphorylation. Int. J. Hematol. Oncol. Stem Cell Res. 2021, 15, 221–229. [Google Scholar] [CrossRef]
- Shindo, T.; Tanaka, M.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Kawate, H.; Yamauchi, A.; Sakurai, T. Regulation of cardiovascular development and homeostasis by the adrenomedullin-RAMP system. Peptides 2019, 111, 55–61. [Google Scholar] [CrossRef]
- Henry, S.; Kokity, L.; Pirity, M.K. Polycomb protein RYBP activates transcription factor Plagl1 during in vitro cardiac differentiation of mouse embryonic stem cells. Open Biol. 2023, 13, 220305. [Google Scholar] [CrossRef] [PubMed]
- Sabour, D.; Srinivasan, S.P.; Rohani, S.; Wagh, V.; Gaspar, J.A.; Panek, D.; Ardestani, M.A.; Doss, M.X.; Riet, N.; Abken, H.; et al. STRIP2 Is Indispensable for the Onset of Embryonic Stem Cell Differentiation. Mol. Ther. Methods Clin. Dev. 2017, 5, 116–129. [Google Scholar] [CrossRef]
- Kappei, D.; Butter, F.; Benda, C.; Scheibe, M.; Draskovic, I.; Stevense, M.; Novo, C.L.; Basquin, C.; Araki, M.; Araki, K.; et al. HOT1 is a mammalian direct telomere repeat-binding protein contributing to telomerase recruitment. EMBO J. 2013, 32, 1681–1701. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Luo, Z.; Jiang, S.; Li, F.; Han, X.; Hu, Y.; Wang, D.; Zhao, Y.; Ma, W.; Liu, D.; et al. The telomere-associated homeobox-containing protein TAH1/HMBOX1 participates in telomere maintenance in ALT cells. J. Cell Sci. 2013, 126 Pt 17, 3982–3989. [Google Scholar] [PubMed]
- Odintsova, T.I.; Muller, E.C.; Ivanov, A.V.; Egorov, T.A.; Bienert, R.; Vladimirov, S.N.; Kostka, S.; Otto, A.; Wittmann-Liebold, B.; Karpova, G.G. Characterization and analysis of posttranslational modifications of the human large cytoplasmic ribosomal subunit proteins by mass spectrometry and Edman sequencing. J. Protein Chem. 2003, 22, 249–258. [Google Scholar] [CrossRef]
- McLachlan, F.; Sires, A.M.; Abbott, C.M. The role of translation elongation factor eEF1 subunits in neurodevelopmental disorders. Hum. Mutat. 2019, 40, 131–141. [Google Scholar] [CrossRef]
- Eisenreich, A.; Bogdanov, V.Y.; Zakrzewicz, A.; Pries, A.; Antoniak, S.; Poller, W.; Schultheiss, H.; Rauch, U. Cdc2-like kinases and DNA topoisomerase I regulate alternative splicing of tissue factor in human endothelial cells. Circ. Res. 2009, 104, 589–599. [Google Scholar] [CrossRef]
- Joshi, P.; Greco, T.M.; Guise, A.J.; Luo, Y.; Yu, F.; Nesvizhskii, A.I.; Cristea, I.M. The functional interactome landscape of the human histone deacetylase family. Mol. Syst. Biol. 2013, 9, 672. [Google Scholar] [CrossRef]
- Kay, M.; Soltani, B.M. LncRNAs in Cardiomyocyte Maturation: New Window for Cardiac Regenerative Medicine. Noncoding RNA 2021, 7, 20. [Google Scholar] [CrossRef]
- Schweiger, V.; Hasimbegovic, E.; Kastner, N.; Spannbauer, A.; Traxler, D.; Gyongyosi, M.; Mester-Tonczar, J. Non-Coding RNAs in Stem Cell Regulation and Cardiac Regeneration: Current Problems and Future Perspectives. Int. J. Mol. Sci. 2021, 22, 9160. [Google Scholar] [CrossRef]
- Shen, H.; Heuzey, E.; Mori, D.N.; Wong, C.K.; Colangelo, C.M.; Chung, L.M.; Bruce, C.; Slizovskiy, I.B.; Booth, C.J.; Kreisel, D.; et al. Haptoglobin enhances cardiac transplant rejection. Circ. Res. 2015, 116, 1670–1679. [Google Scholar] [CrossRef] [PubMed]
- Sid-Otmane, C.; Perrault, L.P.; Ly, H.Q. Mesenchymal stem cell mediates cardiac repair through autocrine, paracrine and endocrine axes. J. Transl. Med. 2020, 18, 336. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, M.; Yu, Y.; Li, C.; Zhang, C. Advances in the study of exosomes derived from mesenchymal stem cells and cardiac cells for the treatment of myocardial infarction. Cell Commun. Signal. 2023, 21, 202. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Xia, Y.; Zhao, H.; Xu, X.; Ma, X.; Tao, L. Stem cell-based therapy in cardiac repair after myocardial infarction: Promise, challenges, and future directions. J. Mol. Cell. Cardiol. 2024, 188, 1–14. [Google Scholar] [CrossRef]
- Deutsch, M.A.; Sturzu, A.; Wu, S.M. At a crossroad: Cell therapy for cardiac repair. Circ. Res. 2013, 112, 884–890. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Weintraub, N.L.; Tang, Y. Identification of Critical Molecular Pathways Induced by HDAC11 Overexpression in Cardiac Mesenchymal Stem Cells. Biomolecules 2025, 15, 662. https://doi.org/10.3390/biom15050662
Zhang C, Weintraub NL, Tang Y. Identification of Critical Molecular Pathways Induced by HDAC11 Overexpression in Cardiac Mesenchymal Stem Cells. Biomolecules. 2025; 15(5):662. https://doi.org/10.3390/biom15050662
Chicago/Turabian StyleZhang, Chongyu, Neal L. Weintraub, and Yaoliang Tang. 2025. "Identification of Critical Molecular Pathways Induced by HDAC11 Overexpression in Cardiac Mesenchymal Stem Cells" Biomolecules 15, no. 5: 662. https://doi.org/10.3390/biom15050662
APA StyleZhang, C., Weintraub, N. L., & Tang, Y. (2025). Identification of Critical Molecular Pathways Induced by HDAC11 Overexpression in Cardiac Mesenchymal Stem Cells. Biomolecules, 15(5), 662. https://doi.org/10.3390/biom15050662