
Interaction Between PHF8 and a Segment of KDM2A, Which Is Controlled by the Phosphorylation Status at a Specific Serine in an Intrinsically Disordered Region of KDM2A, Regulates rRNA Transcription and Cell Proliferation in a Breast Cancer Cell Line

 , , , , and
, , , , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. Introduction of Plasmids and siRNAs into Mammalian Cells
2.3. Plasmids
2.4. Western Blotting of Cell Lysates and Immunoprecipitation
2.5. Methods for Tissue Processing and Immunostaining
2.6. Metabolic Labeling Assay of Newly Synthesized RNA Using 5-Ethynyl Uridine (EU)
2.7. Detection of Phosphorylated Peptide by Phos-Tag
2.8. Production of Anti-Phosphorylated Ser 731
2.9. RNA Extraction and Quantitative Reverse Transcription–Polymerase Chain Reaction (qRT-PCR)
2.10. Statistical Analyses
3. Results
3.1. PHF8 Binds to KDM2A and Is Required for Reduction in rRNA Transcription by Mild Starvation
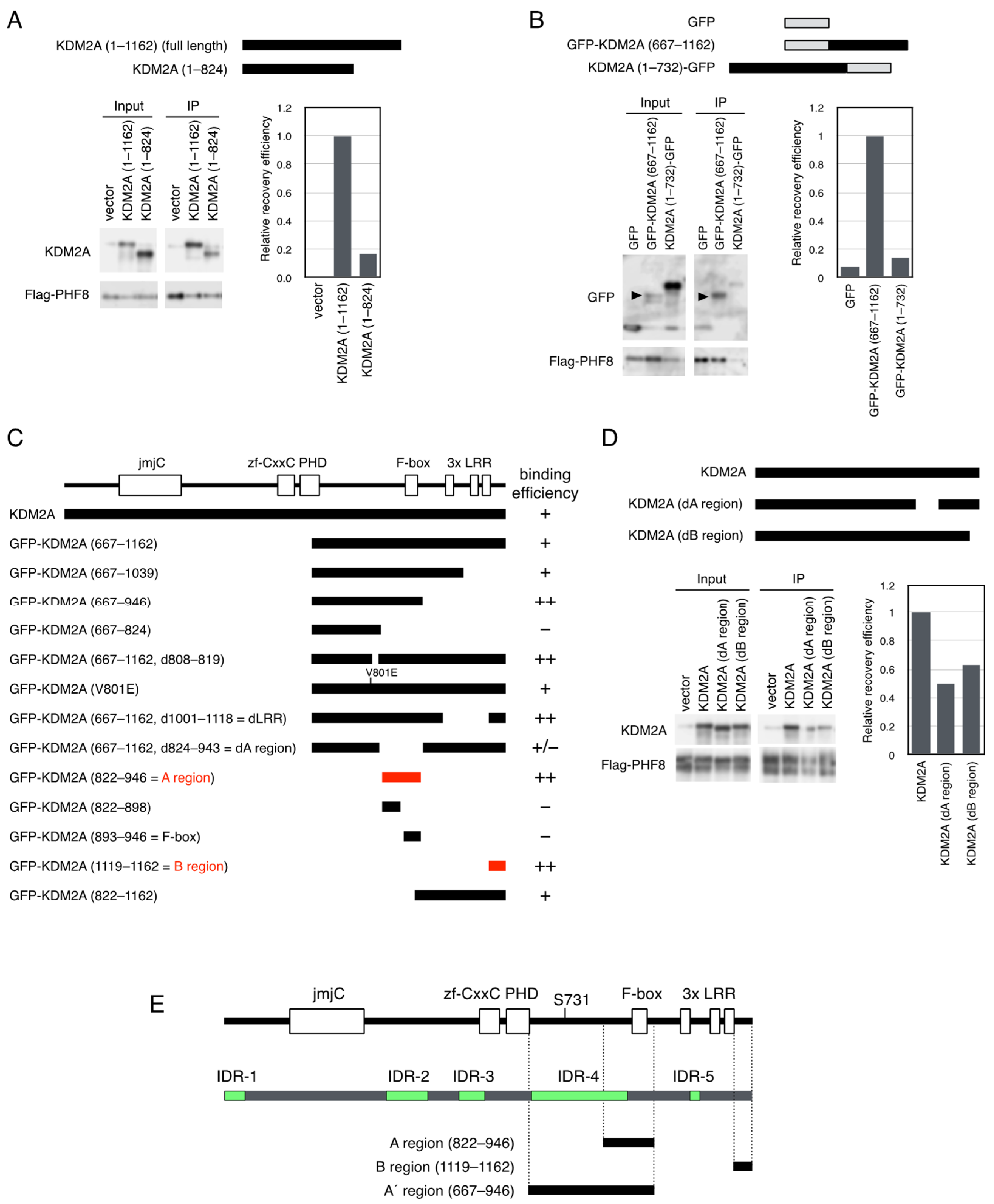
3.2. Identification of KDM2A Regions That Are Involved in Binding to PHF8
3.3. KDM2A with Deleted a Region Reduces rRNA Transcription More Strongly than Wild-Type KDM2A
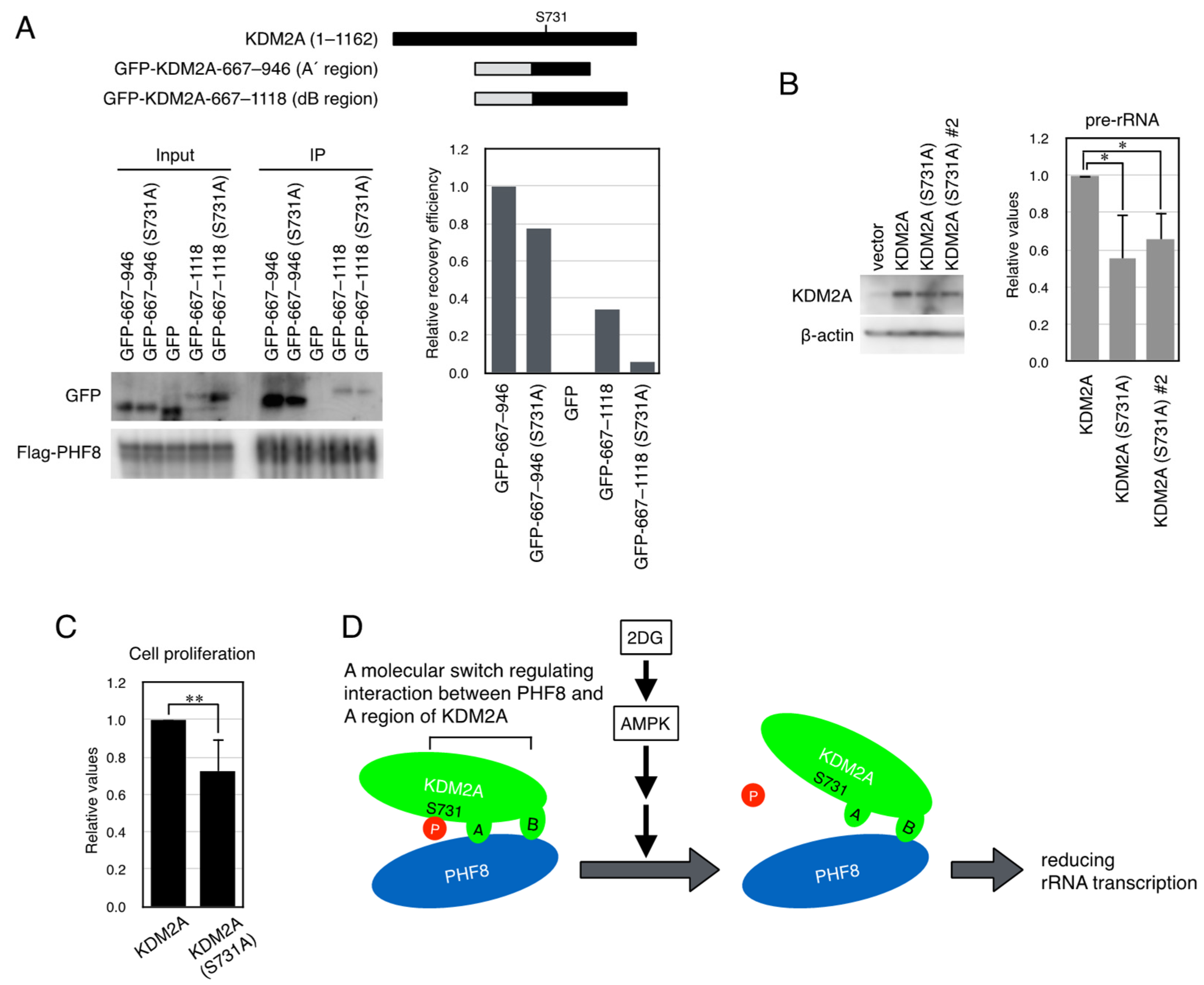
3.4. KDM2A Fragment That Contains the a Region Shows Switchable Interaction with PHF8
3.5. A Dephosphorylation-Mimicked Mutation at Ser731 Reduced the Interaction Between PHF8 and the KDM2A Fragments and rRNA Transcription
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| 2DG | 2-deoxy-D-glucose |
| IDR | intrinsically disordered region |
| KDM2A | lysine-specific demethylase 2A |
| PHF8 | PHD Finger Protein 8 |
| PHD | plant homeodomain |
| KD | knockdown |
| HP1 | heterochromatin protein 1 |
| HEK | human embryonic kidney |
| siRNA | small interfering RNA |
| GFP | green fluorescent protein |
| HRP | horseradish peroxidase |
| EU | 5-ethynyl uridine |
| ECL | electrogenerated chemiluminescence |
| qRT-PCR | quantitative reverse transcription–polymerase chain reaction |
| LRR | leucine-rich repeats |
| AICAR | 5-aminoimidazole-4-carboxamide-1-β-D-ribofuranoside |
| B2M | β2-microglobulin |
| Pol I | RNA polymerase I |
References
- Bywater, M.J.; Pearson, R.B.; McArthur, G.A.; Hannan, R.D. Dysregulation of the basal RNA polymerase transcription apparatus in cancer. Nat. Rev. Cancer 2013, 13, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Chédin, S.; Laferté, A.; Hoang, T.; Lafontaine, D.L.; Riva, M.; Carles, C. Is ribosome synthesis controlled by pol I transcription? Cell Cycle 2007, 6, 11–15. [Google Scholar] [CrossRef]
- Daiß, J.L.; Griesenbeck, J.; Tschochner, H.; Engel, C. Synthesis of the ribosomal RNA precursor in human cells: Mechanisms, factors and regulation. Biol. Chem. 2023, 404, 1003–1023. [Google Scholar] [CrossRef]
- Hori, Y.; Engel, C.; Kobayashi, T. Regulation of ribosomal RNA gene copy number, transcription and nucleolus organization in eukaryotes. Nat. Rev. Mol. Cell Biol. 2023, 24, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Laferté, A.; Favry, E.; Sentenac, A.; Riva, M.; Carles, C.; Chédin, S. The transcriptional activity of RNA polymerase I is a key determinant for the level of all ribosome components. Genes. Dev. 2006, 20, 2030–2040. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.S.; Tsai, P.I.; Chien, C.T. F-box proteins: The key to protein degradation. J. Biomed. Sci. 2006, 13, 181–191. [Google Scholar] [CrossRef]
- Zhou, Y.; Schmitz, K.M.; Mayer, C.; Yuan, X.; Akhtar, A.; Grummt, I. Reversible acetylation of the chromatin remodelling complex NoRC is required for non-coding RNA-dependent silencing. Nat. Cell Biol. 2009, 11, 1010–1016. [Google Scholar] [CrossRef]
- Tanaka, Y.; Okamoto, K.; Teye, K.; Umata, T.; Yamagiwa, N.; Suto, Y.; Zhang, Y.; Tsuneoka, M. JmjC enzyme KDM2A is a regulator of rRNA transcription in response to starvation. EMBO J. 2010, 29, 1510–1522. [Google Scholar] [CrossRef]
- Tanaka, Y.; Umata, T.; Okamoto, K.; Obuse, C.; Tsuneoka, M. CxxC-ZF domain is needed for KDM2A to demethylate histone in rDNA promoter in response to starvation. Cell Struct. Funct. 2014, 39, 79–92. [Google Scholar] [CrossRef]
- Tsukada, Y.; Fang, J.; Erdjument-Bromage, H.; Warren, M.E.; Borchers, C.H.; Tempst, P.; Zhang, Y. Histone demethylation by a family of JmjC domain-containing proteins. Nature 2006, 439, 811–816. [Google Scholar] [CrossRef]
- Gagnon-Kugler, T.; Langlois, F.; Stefanovsky, V.; Lessard, F.; Moss, T. Loss of human ribosomal gene CpG methylation enhances cryptic RNA polymerase II transcription and disrupts ribosomal RNA processing. Mol. Cell 2009, 35, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, K.; Majumder, S.; Datta, J.; Motiwala, T.; Bai, S.; Sharma, S.M.; Frankel, W.; Jacob, S.T. Role of human ribosomal RNA (rRNA) promoter methylation and of methyl-CpG-binding protein MBD2 in the suppression of rRNA gene expression. J. Biol. Chem. 2004, 279, 6783–6793. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.D.; Grummitt, C.G.; Hilcenko, C.; Min, S.Y.; Tonkin, L.M.; Johnson, C.M.; Freund, S.M.; Bycroft, M.; Warren, A.J. Solution structure of the nonmethyl-CpG-binding CXXC domain of the leukaemia-associated MLL histone methyltransferase. EMBO J. 2006, 25, 4503–4512. [Google Scholar] [CrossRef]
- Cierpicki, T.; Risner, L.E.; Grembecka, J.; Lukasik, S.M.; Popovic, R.; Omonkowska, M.; Shultis, D.D.; Zeleznik-Le, N.J.; Bushweller, J.H. Structure of the MLL CXXC domain-DNA complex and its functional role in MLL-AF9 leukemia. Nat. Struct. Mol. Biol. 2010, 17, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Blackledge, N.P.; Zhou, J.C.; Tolstorukov, M.Y.; Farcas, A.M.; Park, P.J.; Klose, R.J. CpG islands recruit a histone H3 lysine 36 demethylase. Mol. Cell 2010, 38, 179–190. [Google Scholar] [CrossRef]
- Kawakami, E.; Tokunaga, A.; Ozawa, M.; Sakamoto, R.; Yoshida, N. The histone demethylase Fbxl11/Kdm2a plays an essential role in embryonic development by repressing cell-cycle regulators. Mech. Dev. 2015, 135, 31–42. [Google Scholar] [CrossRef]
- Cai, Z.; Yan, L.J. Protein Oxidative Modifications: Beneficial Roles in Disease and Health. J. Biochem. Pharmacol. Res. 2013, 1, 15–26. [Google Scholar]
- Wagner, K.W.; Alam, H.; Dhar, S.S.; Giri, U.; Li, N.; Wei, Y.; Giri, D.; Cascone, T.; Kim, J.H.; Ye, Y.; et al. KDM2A promotes lung tumorigenesis by epigenetically enhancing ERK1/2 signaling. J. Clin. Investig. 2013, 123, 5231–5246. [Google Scholar] [CrossRef]
- Ou, R.; Zhu, L.; Zhao, L.; Li, W.; Tao, F.; Lu, Y.; He, Q.; Li, J.; Ren, Y.; Xu, Y. HPV16 E7-induced upregulation of KDM2A promotes cervical cancer progression by regulating miR-132-radixin pathway. J. Cell Physiol. 2019, 234, 2659–2671. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, Y.; Yu, L.; Chen, J.; Hou, J.; Cui, L.; Ma, D.; Lu, W. Histone demethylase KDM2A promotes tumor cell growth and migration in gastric cancer. Tumour Biol. 2015, 36, 271–278. [Google Scholar] [CrossRef]
- Liu, L.; Liu, J.; Lin, Q. Histone demethylase KDM2A: Biological functions and clinical values (Review). Exp. Ther. Med. 2021, 22, 723. [Google Scholar] [CrossRef]
- Li, F.; Wang, Y.; Hwang, I.; Jang, J.Y.; Xu, L.; Deng, Z.; Yu, E.Y.; Cai, Y.; Wu, C.; Han, Z.; et al. Histone demethylase KDM2A is a selective vulnerability of cancers relying on alternative telomere maintenance. Nat. Commun. 2023, 14, 1756. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Yano, H.; Ogasawara, S.; Yoshioka, S.; Imamura, H.; Okamoto, K.; Tsuneoka, M. Mild Glucose Starvation Induces KDM2A-Mediated H3K36me2 Demethylation Through AMPK To Reduce rRNA Transcription and Cell Proliferation. Mol. Cell Biol. 2015, 35, 4170–4184. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Tanaka, Y.; Ogasawara, S.; Obuse, C.; Nakayama, J.I.; Yano, H.; Tsuneoka, M. KDM2A-dependent reduction of rRNA transcription on glucose starvation requires HP1 in cells, including triple-negative breast cancer cells. Oncotarget 2019, 10, 4743–4760. [Google Scholar] [CrossRef] [PubMed]
- Trefts, E.; Shaw, R.J. AMPK: Restoring metabolic homeostasis over space and time. Mol. Cell 2021, 81, 3677–3690. [Google Scholar] [CrossRef]
- Steinberg, G.R.; Hardie, D.G. New insights into activation and function of the AMPK. Nat. Rev. Mol. Cell Biol. 2023, 24, 255–272. [Google Scholar] [CrossRef]
- Basu, S.; Kihara, D.; Kurgan, L. Computational prediction of disordered binding regions. Comput. Struct. Biotechnol. J. 2023, 21, 1487–1497. [Google Scholar] [CrossRef]
- Cermakova, K.; Hodges, H.C. Interaction modules that impart specificity to disordered protein. Trends Biochem. Sci. 2023, 48, 477–490. [Google Scholar] [CrossRef]
- Patil, A.; Strom, A.R.; Paulo, J.A.; Collings, C.K.; Ruff, K.M.; Shinn, M.K.; Sankar, A.; Cervantes, K.S.; Wauer, T.; St Laurent, J.D.; et al. A disordered region controls cBAF activity via condensation and partner recruitment. Cell 2023, 186, 4936–4955.e26. [Google Scholar] [CrossRef]
- King, M.R.; Ruff, K.M.; Lin, A.Z.; Pant, A.; Farag, M.; Lalmansingh, J.M.; Wu, T.; Fossat, M.J.; Ouyang, W.; Lew, M.D.; et al. Macromolecular condensation organizes nucleolar sub-phases to set up a pH gradient. Cell 2024, 187, 1889–1906.e24. [Google Scholar] [CrossRef]
- Banerjee, P.R.; Holehouse, A.S.; Kriwacki, R.; Robustelli, P.; Jiang, H.; Sobolevsky, A.I.; Hurley, J.M.; Mendell, J.T. Dissecting the biophysics and biology of intrinsically disordered proteins. Trends Biochem. Sci. 2024, 49, 101–104. [Google Scholar] [CrossRef]
- Holehouse, A.S.; Kragelund, B.B. The molecular basis for cellular function of intrinsically disordered protein regions. Nat. Rev. Mol. Cell Biol. 2023, 25, 187–211. [Google Scholar] [CrossRef]
- Horton, J.R.; Upadhyay, A.K.; Qi, H.H.; Zhang, X.; Shi, Y.; Cheng, X. Enzymatic and structural insights for substrate specificity of a family of jumonji histone lysine demethylases. Nat. Struct. Mol. Biol. 2010, 17, 38–43. [Google Scholar] [CrossRef]
- Laumonnier, F.; Holbert, S.; Ronce, N.; Faravelli, F.; Lenzner, S.; Schwartz, C.E.; Lespinasse, J.; Van Esch, H.; Lacombe, D.; Goizet, C.; et al. Mutations in PHF8 are associated with X linked mental retardation and cleft lip/cleft palate. J. Med. Genet. 2005, 42, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Yonezawa, M.; Ye, J.; Jenuwein, T.; Grummt, I. PHF8 activates transcription of rRNA genes through H3K4me3 binding and H3K9me1/2 demethylation. Nat. Struct. Mol. Biol. 2010, 17, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Wang, Y.; Li, X.; Xu, L.; Wang, X.; Sun, T.; Dong, X.; Chen, L.; Mao, H.; Yu, Y.; et al. PHF8 is a histone H3K9me2 demethylase regulating rRNA synthesis. Cell Res. 2010, 20, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, S.; Zhou, Y.; Han, Y.; Li, S.; Xu, Q.; Xu, L.; Zhu, Z.; Deng, Y.; Yu, L.; et al. Phf8 histone demethylase deficiency causes cognitive impairments through the mTOR pathway. Nat. Commun. 2018, 9, 114. [Google Scholar] [CrossRef]
- Koivisto, A.M.; Ala-Mello, S.; Lemmelä, S.; Komu, H.A.; Rautio, J.; Järvelä, I. Screening of mutations in the PHF8 gene and identification of a novel mutation in a Finnish family with XLMR and cleft lip/cleft palate. Clin. Genet. 2007, 72, 145–149. [Google Scholar] [CrossRef]
- Abidi, F.; Miano, M.; Murray, J.; Schwartz, C. A novel mutation in the PHF8 gene is associated with X-linked mental retardation with cleft lip/cleft palate. Clin. Genet. 2007, 72, 19–22. [Google Scholar] [CrossRef]
- Fan, T.; Xie, J.; Huang, G.; Li, L.; Zeng, X.; Tao, Q. PHF8/KDM7B: A Versatile Histone Demethylase and Epigenetic Modifier in Nervous System Disease and Cancers. Epigenomes 2024, 8, 36. [Google Scholar] [CrossRef]
- Wang, Q.; Ma, S.; Song, N.; Li, X.; Liu, L.; Yang, S.; Ding, X.; Shan, L.; Zhou, X.; Su, D.; et al. Stabilization of histone demethylase PHF8 by USP7 promotes breast carcinogenesis. J. Clin. Investig. 2016, 126, 2205–2220. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Borcherding, N.C.; Shao, P.; Maina, P.K.; Zhang, W.; Qi, H.H. Contribution of synergism between PHF8 and HER2 signalling to breast cancer development and drug resistance. eBioMedicine 2020, 51, 102612. [Google Scholar] [CrossRef] [PubMed]
- Shao, P.; Liu, Q.; Maina, P.K.; Cui, J.; Bair, T.B.; Li, T.; Umesalma, S.; Zhang, W.; Qi, H.H. Histone demethylase PHF8 promotes epithelial to mesenchymal transition and breast tumorigenesis. Nucleic Acids Res. 2017, 45, 1687–1702. [Google Scholar] [CrossRef]
- Erdős, G.; Dosztányi, Z. AIUPred: Combining energy estimation with deep learning for the enhanced prediction of protein disorder. Nucleic Acids Res. 2024, 52, W176–W181. [Google Scholar] [CrossRef] [PubMed]
- Allred, D.C.; Harvey, J.M.; Berardo, M.; Clark, G.M. Prognostic and predictive factors in breast cancer by immunohistochemical analysis. Mod. Pathol. 1998, 11, 155–168. [Google Scholar]
- Theriault, R.L.; Carlson, R.W.; Allred, C.; Anderson, B.O.; Burstein, H.J.; Edge, S.B.; Farrar, W.B.; Forero, A.; Giordano, S.H.; Goldstein, L.J.; et al. Breast cancer, version 3.2013: Featured updates to the NCCN guidelines. J. Natl. Compr. Cancer Netw. 2013, 11, 753–760, quiz 761. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Sakimura, A.; Park, C.M.; Tomaru, R.; Tanaka, T.; Ozawa, T.; Zhou, Y.; Narita, K.; Kishi, H.; Muraguchi, A.; et al. Feedback control of ErbB2 via ERK-mediated phosphorylation of a conserved threonine in the juxtamembrane domain. Sci. Rep. 2016, 6, 31502. [Google Scholar] [CrossRef]
- Letunic, I.; Copley, R.R.; Schmidt, S.; Ciccarelli, F.D.; Doerks, T.; Schultz, J.; Ponting, C.P.; Bork, P. SMART 4.0: Towards genomic data integration. Nucleic Acids Res. 2004, 32, D142–D144. [Google Scholar] [CrossRef]
- Chen, Z.; Lei, C.; Wang, C.; Li, N.; Srivastava, M.; Tang, M.; Zhang, H.; Choi, J.M.; Jung, S.Y.; Qin, J.; et al. Global phosphoproteomic analysis reveals ARMC10 as an AMPK substrate that regulates mitochondrial dynamics. Nat. Commun. 2019, 10, 104. [Google Scholar] [CrossRef]
- Mayer, C.; Zhao, J.; Yuan, X.; Grummt, I. mTOR-dependent activation of the transcription factor TIF-IA links rRNA synthesis to nutrient availability. Genes Dev. 2004, 18, 423–434. [Google Scholar] [CrossRef]
- Sukumaran, A.; Choi, K.; Dasgupta, B. Insight on Transcriptional Regulation of the Energy Sensing AMPK and Biosynthetic mTOR Pathway Genes. Front. Cell Dev. Biol. 2020, 8, 671. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Manning, B.D. The TSC1-TSC2 complex: A molecular switchboard controlling cell growth. Biochem. J. 2008, 412, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, S.; Bierhoff, H.; Cado, I.; Weber, A.; Tiebe, M.; Grummt, I.; Voit, R. AMP-activated protein kinase adapts rRNA synthesis to cellular energy supply. Proc. Natl. Acad. Sci. USA 2009, 106, 17781–17786. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Baek, A.; Hwang, J.E.; Choi, Y.A.; Jeong, J.; Lee, M.S.; Cho, D.H.; Lim, J.S.; Kim, K.I.; Yang, Y. Adiponectin-activated AMPK stimulates dephosphorylation of AKT through protein phosphatase 2A activation. Cancer Res. 2009, 69, 4018–4026. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stealth siRNA | siRNA Sequence |
|---|---|
| siKDM2A | 5′-GAACCCGAAGAAGAAAGGAUUCGUU-3′ |
| siPHF8#1 | 5′-CAACAAAUGCCAAUCUGACUCUCUU-3′ |
| siPHF8#2 | 5′-GAGCUCCGGAGUAGGACUUUUGACA-3′ |
| control siRNA | Stealth RNAi Negative Control Medium GC Duplex, Thermo Fisher |
| Antigen | Antibody |
|---|---|
| KDM2A | Rabbit anti-KDM2A polyclonal antibody (Proteintech, 24311-1-AP) Rabbit anti-KDM2A monoclonal antibody (Abcam, ab191387) Anti-FbxL11 (KDM2A) antibody (Abcam, ab99242) |
| PHF8 | Rabbit anti-PHF8 monoclonal antibody (Cell Signaling Technology, PHF8 (E6K3Y)) |
| Flag | Rabbit polyclonal anti-Flag antibody (Sigma, F7425) Mouse monoclonal anti-Flag antibody (M2, Sigma, F1804) |
| GFP | Rabbit polyclonal anti-GFP antibody (Abcam, ab290) Mouse monoclonal anti-GFP antibody (Santa Cruz, ab290) |
| β-actin | Mouse monoclonal anti-β-actin antibody (Sigma, AC-15) |
| Rabbit IgG | Goat-rabbit IgG-horseradish peroxidase (HRP) (Abcam, ab6721) Goat anti-rabbit IgG-HRP (Santa Cruz, sc-2054, human and mouse IgG adsorbed) Goat anti-rabbit IgG(H&L)-Cy3 conjugated (Zymed, #81-6115) |
| Mouse IgG | Goat anti-mouse IgG-HRP (Santa Cruz, sc-2005) |
| Gene | Primer Sequence |
|---|---|
| pre-rRNA | Forward: 5′-GCTGACACGCTGTCCTCTG-3′ Reverse: 5′-TCGGACGCGCGAGAGAAC-3′ |
| KDM2A | Forward: 5′-TCCCCACACACATTTTGACATC-3′ Reverse: 5′-GGGGTGGCTTGAGAGATCCT-3′ |
| PHF8 | Forward: 5′-AGCCCTACGTTCGTCAGAGA-3′ Reverse: 5′-CAACCCATCCTTCTTCAGGA-3′ |
| Polr2a | Forward: 5′-ATCTCTCCTGCCATGACACC-3′ Reverse: 5′-AGACCAGGCAGGGGAGTAAC-3′ |
| β-actin | Forward: 5′-CGTCTTCCCCTCCATCGT-3′ Reverse: 5′-GAAGGTGTGGTGCCAGATTT-3 |
| B2M | Forward: 5′-CTCGCGCTACTCTCTCTTTCT-3′ Reverse: 5′-TGTCGGATTGATGAAACCCAG-3′ |
| PHF8 score | Papillotubular | Solid tubular | Scirrhpous | Mucinous | Miceopapillary | |||||||||||||||||||||
| 4 | 5 | 6 | 7 | 8 | 4 | 5 | 6 | 7 | 8 | 4 | 5 | 6 | 7 | 8 | 4 | 5 | 6 | 7 | 8 | 4 | 5 | 6 | 7 | 8 | ||
| Number | 2 | 2 | 2 | 8 | 3 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 4 | 5 | 3 | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | |
| HER2 | 0 | 0 | 0 | 0 | 6 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 4 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 2 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | |
| 3 | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | |
| ER | − | 0 | 1 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| + | 2 | 1 | 1 | 6 | 3 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 4 | 4 | 3 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | |
| PgR | − | 0 | 2 | 2 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| + | 2 | 0 | 0 | 6 | 3 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 3 | 4 | 3 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | |
| Triple Negative | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Species | Total AA | JmjC Domain | A Region | IDR-4 | IDR-4 (AA 700-808) |
|---|---|---|---|---|---|
| Mouse | AA 1161 (97.3%) | 168/169 (99.4%) | 118/125 (94.4%) | 208/217 (95.8%) | 108/109 (99%) |
| Chicken | AA 1168 (86%) | 156/169 (92%) | 95/125 (76%) | 171/217 (78.8%) | 100/108 (91.7%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okamoto, K.; Mihara, Y.; Ogasawara, S.; Murakami, T.; Ohmori, S.; Mori, T.; Umata, T.; Kawasaki, Y.; Hirano, K.; Yano, H.; et al. Interaction Between PHF8 and a Segment of KDM2A, Which Is Controlled by the Phosphorylation Status at a Specific Serine in an Intrinsically Disordered Region of KDM2A, Regulates rRNA Transcription and Cell Proliferation in a Breast Cancer Cell Line. Biomolecules 2025, 15, 661. https://doi.org/10.3390/biom15050661
Okamoto K, Mihara Y, Ogasawara S, Murakami T, Ohmori S, Mori T, Umata T, Kawasaki Y, Hirano K, Yano H, et al. Interaction Between PHF8 and a Segment of KDM2A, Which Is Controlled by the Phosphorylation Status at a Specific Serine in an Intrinsically Disordered Region of KDM2A, Regulates rRNA Transcription and Cell Proliferation in a Breast Cancer Cell Line. Biomolecules. 2025; 15(5):661. https://doi.org/10.3390/biom15050661
Chicago/Turabian StyleOkamoto, Kengo, Yutaro Mihara, Sachiko Ogasawara, Takashi Murakami, Sinya Ohmori, Tetsuya Mori, Toshiyuki Umata, Yuki Kawasaki, Kazuya Hirano, Hirohisa Yano, and et al. 2025. "Interaction Between PHF8 and a Segment of KDM2A, Which Is Controlled by the Phosphorylation Status at a Specific Serine in an Intrinsically Disordered Region of KDM2A, Regulates rRNA Transcription and Cell Proliferation in a Breast Cancer Cell Line" Biomolecules 15, no. 5: 661. https://doi.org/10.3390/biom15050661
APA StyleOkamoto, K., Mihara, Y., Ogasawara, S., Murakami, T., Ohmori, S., Mori, T., Umata, T., Kawasaki, Y., Hirano, K., Yano, H., & Tsuneoka, M. (2025). Interaction Between PHF8 and a Segment of KDM2A, Which Is Controlled by the Phosphorylation Status at a Specific Serine in an Intrinsically Disordered Region of KDM2A, Regulates rRNA Transcription and Cell Proliferation in a Breast Cancer Cell Line. Biomolecules, 15(5), 661. https://doi.org/10.3390/biom15050661