
Immunological Fluid Biomarkers in Frontotemporal Dementia: A Systematic Review
 , and
, and
Abstract
1. Introduction
2. Methods
2.1. Eligibility Criteria
2.2. Search Strategy
2.3. Data Extraction
2.4. Data Synthesis and Analysis
2.5. Enrichment Analysis
2.6. Risk of Bias
3. Results
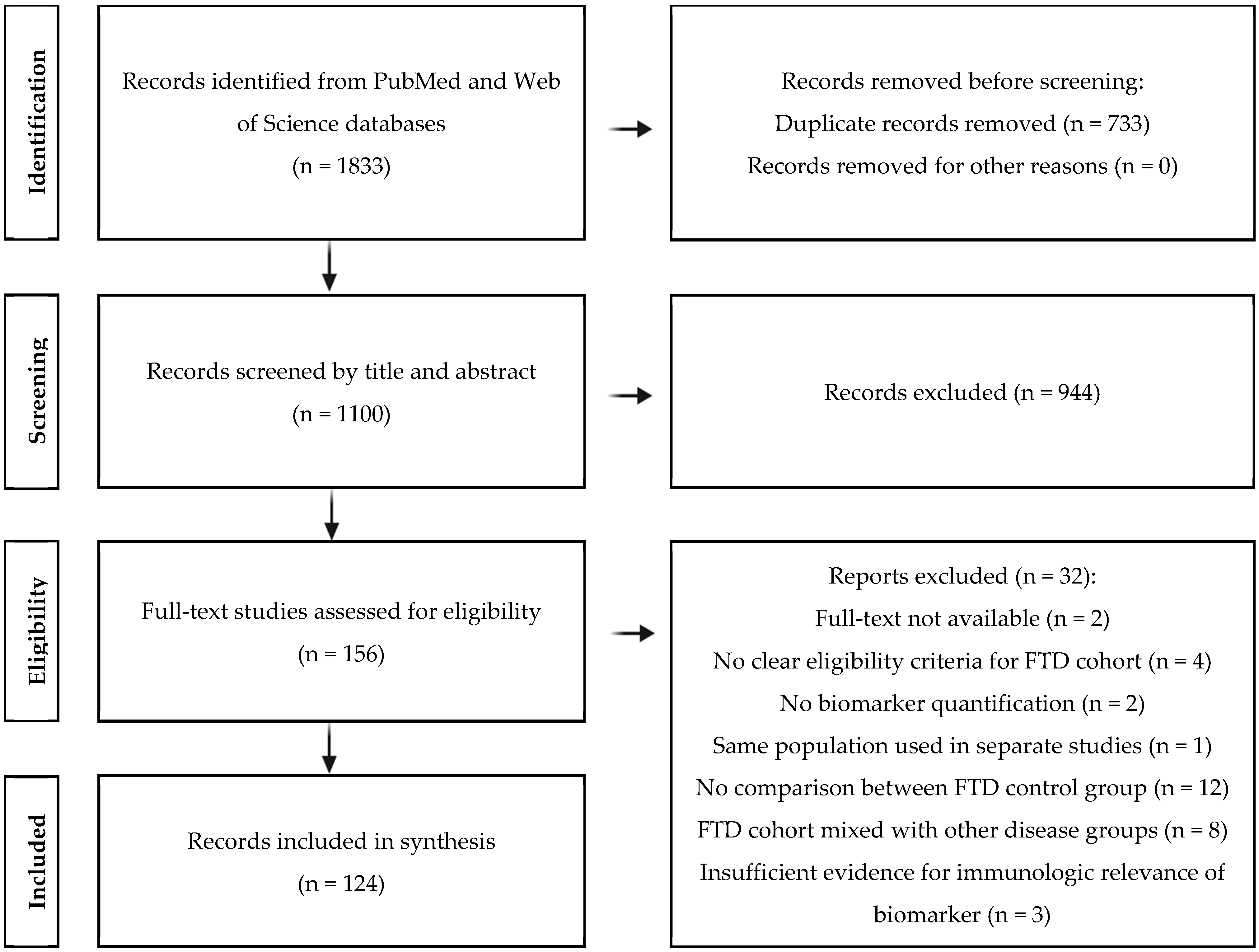
3.1. Results of Study Selection Process
3.2. Analytical Study Characteristics
3.3. Biomarker Results
3.4. Sample Sizes and Demographics
3.5. Diagnostic Criteria for FTD
3.6. Risk of Bias Assessment
3.7. FTD Patients and Controls
3.8. Diagnostic Performance
3.9. Correlation of Immune Markers

{kind=link}
{kind=link}
| Biomarker | Specificity | Sensitivity | AUC | |
|---|---|---|---|---|
| FTD vs. HC | ||||
| CSF | ↑ GFAP | 0.63 [29] | 0.67 [29] | 0.71 [29] |
| ↑ YKL-40 | 0.53 [109], 0.73 [29], 0.83 [31] | 0.54 [31], 0.70 [29], 0.86 [109] | 0.69–0.88 [29,31,33,35,109] | |
| ↑ CHIT1 | 0.63 [29] | 0.73 [29] | 0.69 [29] | |
| ↑ MFG-E8 | NA | NA | 0.55 [33] | |
| Blood | ↑ GFAP | NA | NA | 0.76–0.83 [16,20] |
| ↑ YKL-40 | NA | NA | 0.65 [91] | |
| Infl. miRNA | 0.88 [95] | 0.86 [95] | 0.95 [95] | |
| ↑ HERV-K | 0.67 [90] | 0.92 [90] | 0.87 [90] | |
| FTD vs. AD | ||||
| CSF | ↑ YKL-40 | 0.16 [109] | 0.85 [109] | 0.54–0.60 [33,35,109] |
| Blood | ↓ GFAP | 0.67 [25], 0.73 [100], 0.92 [110] | 0.71 [110], 0.75 [100], 0.82 [25] | 0.65–0.85 [16,18,19,25,27,100,110] |
| FTD vs. ALS | ||||
| CSF | ↓ CXCL12 | NA | NA | 0.57 [72] |
| Blood | Infl. miRNA | 0.75 [95] | 0.62 [95] | 0.78 [95] |
| ↑ HERV-K | NA | NA | 0.66 [90] | |
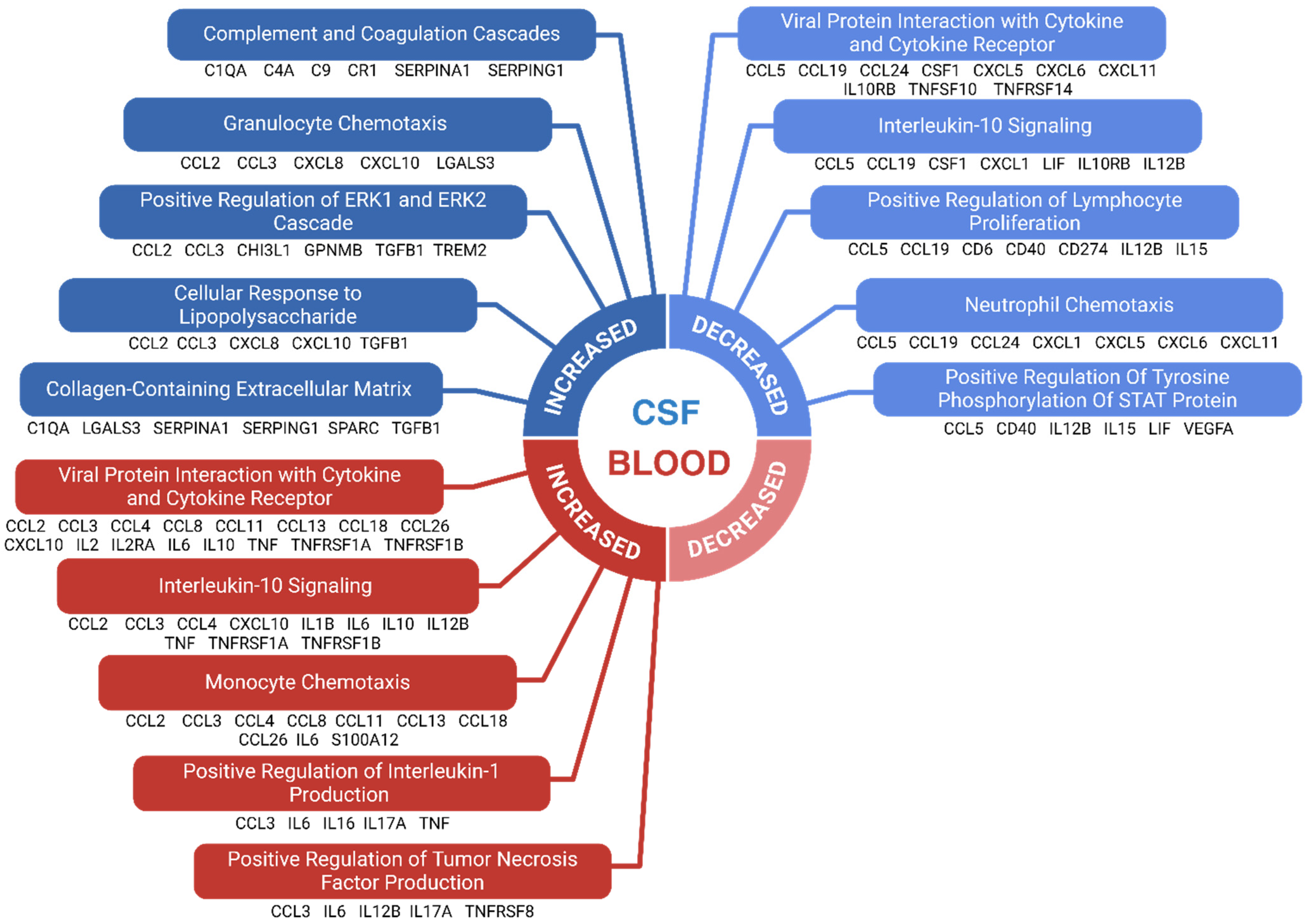
3.10. Results of Enrichment Analysis
4. Discussion
4.1. Methodological Variability Among Studies
4.2. Immunological Biomarkers in FTD vs. Healthy Controls
4.3. Contradictory Immunological Biomarkers
4.4. CSF and Blood Discrepancies
4.5. FTD Versus Alzheimer’s Disease
4.6. FTD Versus ALS
4.7. Immunological Biomarkers Among FTD Subtypes
4.8. Biomarker Correlations and Diagnostic Potential
4.9. Enriched Immunological Pathways in FTD
4.10. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease | lvPPA | Logopenic variant primary progressive aphasia |
| ADA | Adenosine deaminase | MAPT | Microtubule-associated protein tau |
| ALS | Amyotrophic lateral sclerosis | MCI | Mild cognitive impairment |
| ANA | Antinuclear antibody | MCP1/2/4 | Monocyte chemoattractant protein 1/2/4 |
| APN | Adiponectin | MDC | Macrophage-derived chemokine |
| APOE/L1 | Apolipoprotein E/L1 | MFG-E8 | Milk fat globule-EGF factor 8 |
| AUC | Area under the curve | MIF | Macrophage migration inhibitory factor |
| Aβ42 | Amyloid beta 42 | MIG | Monokine induced by gamma interferon |
| BAFF | B-cell-activating factor | MMP-10 | Matrix metalloproteinase 10 |
| bvFTD | Behavioral variant frontotemporal dementia | MMSE | Mini-mental state examination |
| C1q-C9 | Complement components 1q to 9 | MS | Multiple sclerosis |
| C9orf72 | Chromosome 9 open reading frame 72 | Myosin-2 | Myosin II |
| Cathepsin S | Cathepsin S | NfL | Neurofilament Light Chain |
| CCL | Chemokine (C-C motif) ligand | nfvPPA | Nonfluent variant primary progressive aphasia |
| CCR | Chemokine (C-C motif) receptor | NOD2 | Nucleotide-binding oligomerization domain-containing 2 |
| CD | Cluster of Differentiation | NOS | Nitric oxide synthase |
| CFH/CFI | Complement factor H/I | PD | Parkinson’s disease |
| CHI3L1 | Chitinase-3-like protein 1 | PDGF-BB | Platelet-derived growth factor-BB |
| CHI3L2 | Chitinase-3-like protein 2 | PD-L1 | Programmed death-ligand 1 |
| CHIT1 | Chitotriosidase 1 | PGRN | Progranulin |
| CRP | C-reactive protein | PPA | Primary progressive aphasia |
| CSF | Cerebrospinal fluid | PRISMA | Preferred reporting items for systematic reviews and meta-analyses |
| CSF-1 | Colony-stimulating factor 1 | PROS-1 | Protein S |
| CTACK | Cutaneous T-cell-attracting chemokine | QUADAS-2 | Quality Assessment of Diagnostic Accuracy Studies 2 |
| CXC | Chemokine (C-X-C motif) | RANTES | Regulated upon activation, normal T-cell expressed and presumably secreted |
| CXCL | Chemokine (C-X-C motif) ligand | S100A12/A8/B | S100 calcium-binding protein A12/A8/B |
| Cystatin C | Cystatin C | sAPPβ | Soluble amyloid precursor protein beta |
| DKK-3 | Dickkopf-3 | SCF | Stem cell factor |
| DNER | Delta/notch-like EGF repeat-containing | SCGF-BB | Stem cell growth factor-BB |
| EAE | Experimental autoimmune encephalomyelitis | SDF-1α | Stromal cell-derived factor 1 alpha |
| ERK/MAPK | Extracellular signal-regulated kinase/mitogen-activated protein kinase | SERPINA1/G1 | Serpin family A member 1 (alpha-1 antitrypsin) |
| FGF | Fibroblast growth factor 19 | SLPI | Secretory leukocyte protease inhibitor |
| Flt3L | Fms-like tyrosine kinase 3 ligand | SORL1 | Sortilin-related receptor 1 |
| FTD | Frontotemporal dementia | SPARC | Secreted protein acidic and rich in cysteine (osteonectin) |
| FTLD | Frontotemporal lobar degeneration | SPP1 | Secreted phosphoprotein 1 (osteopontin) |
| Gal-3 | Galectin-3 | SST | Somatostatin |
| G-CSF | Granulocyte colony-stimulating factor | svPPA | Semantic variant primary progressive aphasia |
| GFAP | Glial fibrillary acidic protein | TARC | Thymus and activation-regulated chemokine (CCL17) |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor | Tdp-43 | TAR DNA-binding protein 43 |
| GPNMB | Glycoprotein non-metastatic melanoma protein B | TGF-α/β | Transforming growth factor alpha/beta |
| GRN | Granulin | TNFRI/II | Tumor necrosis factor receptor I/II |
| GRO-α | Growth-regulated oncogene alpha | TNFRSF14 | Tumor necrosis factor receptor superfamily member 14 (HVEM) |
| GSN | Gelsolin | TNFRSF8 | Tumor necrosis factor receptor superfamily member 8 (CD30) |
| HERV-K | Human endogenous retrovirus K | TNFRSF9 | Tumor necrosis factor receptor superfamily member 9 (4-1BB) |
| HGF | Hepatocyte growth factor | TNFα | Tumor necrosis factor-alpha |
| ICAM-1 | Intercellular adhesion molecule 1 | TRAIL | TNF-related apoptosis-inducing ligand |
| IFN-α2/γ | Interferon alpha 2/gamma | TREM2 | Triggering receptor expressed on myeloid cells 2 |
| IgG | Immunoglobulin G | TWEAK | TNF-related weak inducer of apoptosis |
| IGHV1-2 | Immunoglobulin heavy variable 1-2 | uPA | Urokinase-type plasminogen activator |
| IL | Interleukin | VCAM | Vascular cell adhesion molecule |
| Irisin | Irisin | VEGF | Vascular endothelial growth factor |
| KYNA | Kynurenic acid | YKL-40 | Chitinase-3-like protein 1 |
| LBP | Lipopolysaccharide-binding protein | β-NGF | Beta nerve growth factor |
| LIF | Leukemia inhibitory factor |
References
- Hogan, D.B.; Jetté, N.; Fiest, K.M.; Roberts, J.I.; Pearson, D.; Smith, E.E.; Roach, P.; Kirk, A.; Pringsheim, T.; Maxwell, C.J. The Prevalence and Incidence of Frontotemporal Dementia: A Systematic Review. Can. J. Neurol. Sci. 2016, 43 (Suppl. S1), S96–S109. [Google Scholar] [CrossRef]
- Greaves, C.V.; Rohrer, J.D. An update on genetic frontotemporal dementia. J. Neurol. 2019, 266, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Antonioni, A.; Raho, E.M.; Lopriore, P.; Pace, A.P.; Latino, R.R.; Assogna, M.; Mancuso, M.; Gragnaniello, D.; Granieri, E.; Pugliatti, M.; et al. Frontotemporal Dementia, Where Do We Stand? A Narrative Review. Int. J. Mol. Sci. 2023, 24, 11732. [Google Scholar] [CrossRef] [PubMed]
- Kansal, K.; Mareddy, M.; Sloane, K.L.; Minc, A.A.; Rabins, P.V.; McGready, J.B.; Onyike, C.U. Survival in Frontotemporal Dementia Phenotypes: A Meta-Analysis. Dement. Geriatr. Cogn. Disord. 2016, 41, 109–122. [Google Scholar] [CrossRef]
- Bright, F.; Werry, E.L.; Dobson-Stone, C.; Piguet, O.; Ittner, L.M.; Halliday, G.M.; Hodges, J.R.; Kiernan, M.C.; Loy, C.T.; Kassiou, M.; et al. Neuroinflammation in frontotemporal dementia. Nat. Rev. Neurol. 2019, 15, 540–555. [Google Scholar] [CrossRef]
- Lui, H.; Zhang, J.; Makinson, S.R.; Cahill, M.K.; Kelley, K.W.; Huang, H.Y.; Shang, Y.; Oldham, M.C.; Martens, L.H.; Gao, F.; et al. Progranulin Deficiency Promotes Circuit-Specific Synaptic Pruning by Microglia via Complement Activation. Cell 2016, 165, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Hartnell, I.J.; Blum, D.; Nicoll, J.A.R.; Dorothee, G.; Boche, D. Glial cells and adaptive immunity in frontotemporal dementia with tau pathology. Brain 2021, 144, 724–745. [Google Scholar] [CrossRef]
- van der Ende, E.L.; Heller, C.; Sogorb-Esteve, A.; Swift, I.J.; McFall, D.; Peakman, G.; Bouzigues, A.; Poos, J.M.; Jiskoot, L.C.; Panman, J.L.; et al. Elevated CSF and plasma complement proteins in genetic frontotemporal dementia: Results from the GENFI study. J. Neuroinflamm. 2022, 19, 217. [Google Scholar] [CrossRef]
- Zang, X.; Chen, S.; Zhu, J.; Ma, J.; Zhai, Y. The Emerging Role of Central and Peripheral Immune Systems in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 872134. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA Statement. Open Med. 2009, 3, e123–e130. [Google Scholar]
- Milacic, M.; Beavers, D.; Conley, P.; Gong, C.; Gillespie, M.; Griss, J.; Haw, R.; Jassal, B.; Matthews, L.; May, B.; et al. The Reactome Pathway Knowledgebase 2024. Nucleic Acids Res. 2023, 52, D672–D678. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed]
- Carbon, S.; Dietze, H.; Lewis, S.E.; Mungall, C.J.; Munoz-Torres, M.C.; Basu, S.; Chisholm, R.L.; Dodson, R.J.; Fey, P.; Thomas, P.D.; et al. Expansion of the Gene Ontology knowledgebase and resources. Nucleic Acids Res. 2017, 45, D331–D338. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef] [PubMed]
- Whiting, P.F.; Rutjes, A.W.; Westwood, M.E.; Mallett, S.; Deeks, J.J.; Reitsma, J.B.; Leeflang, M.M.; Sterne, J.A.; Bossuyt, P.M.; QUADAS-2 Group. QUADAS-2: A revised tool for the quality assessment of diagnostic accuracy studies. Ann. Intern. Med. 2011, 155, 529–536. [Google Scholar] [CrossRef]
- Zhu, N.; Santos-Santos, M.; Illán-Gala, I.; Montal, V.; Estellés, T.; Barroeta, I.; Altuna, M.; Arranz, J.; Muñoz, L.; Belbin, O.; et al. Plasma glial fibrillary acidic protein and neurofilament light chain for the diagnostic and prognostic evaluation of frontotemporal dementia. Transl. Neurodegener. 2021, 10, 50. [Google Scholar] [CrossRef]
- Van Der Ende, E.L.; Bron, E.E.; Poos, J.M.; Jiskoot, L.C.; Panman, J.L.; Papma, J.M.; Meeter, L.H.; Dopper, E.G.P.; Wilke, C.; Synofzik, M.; et al. A data-driven disease progression model of fluid biomarkers in genetic frontotemporal dementia. Brain A J. Neurol. 2022, 145, 1805–1817. [Google Scholar] [CrossRef]
- Baiardi, S.; Quadalti, C.; Mammana, A.; Dellavalle, S.; Zenesini, C.; Sambati, L.; Pantieri, R.; Polischi, B.; Romano, L.; Suffritti, M.; et al. Diagnostic value of plasma p-tau181, NfL, and GFAP in a clinical setting cohort of prevalent neurodegenerative dementias. Alzheimer’s Res. Ther. 2022, 14, 153. [Google Scholar] [CrossRef]
- Sarto, J.; Ruiz-García, R.; Guillén, N.; Ramos-Campoy, Ó.; Falgàs, N.; Esteller, D.; Contador, J.; Fernández, G.; González, Y.; Tort-Merino, A.; et al. Diagnostic Performance and Clinical Applicability of Blood-Based Biomarkers in a Prospective Memory Clinic Cohort. Neurology 2023, 100, E860–E873. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Tao, Q.; Lu, P.; Meng, F.; Zhuang, L.; Qiao, S.; Zhang, Y.; Luo, B.; Liu, Y.; et al. Diagnostic value of isolated plasma biomarkers and its combination in neurodegenerative dementias: A multicenter cohort study. Clin. Chim. Acta 2024, 558, 118784. [Google Scholar] [CrossRef]
- Oeckl, P.; Weydt, P.; Steinacker, P.; Anderl-Straub, S.; Nordin, F.; Volk, A.E.; Diehl-Schmid, J.; Andersen, P.M.; Kornhuber, J.; Danek, A.; et al. Different neuroinflammatory profile in amyotrophic lateral sclerosis and frontotemporal dementia is linked to the clinical phase. J. Neurol. Neurosurg. Psychiatry 2019, 90, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Katisko, K.; Cajanus, A.; Huber, N.; Jääskeläinen, O.; Kokkola, T.; Kärkkäinen, V.; Rostalski, H.; Hartikainen, P.; Koivisto, A.M.; Hannonen, S.; et al. GFAP as a biomarker in frontotemporal dementia and primary psychiatric disorders: Diagnostic and prognostic performance. J. Neurol. Neurosurg. Psychiatry 2021, 92, 1305–1312. [Google Scholar] [CrossRef]
- Fenoglio, C.; Serpente, M.; Arcaro, M.; Carandini, T.; Sacchi, L.; Pintus, M.; Rotondo, E.; Borracci, V.; Ghezzi, L.; Bouzigues, A.; et al. Inflammatory plasma profile in genetic symptomatic and presymptomatic Frontotemporal Dementia—A GENFI study. Brain Behav. Immun. 2024, 122, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Saraceno, C.; Cervellati, C.; Trentini, A.; Crescenti, D.; Longobardi, A.; Geviti, A.; Bonfiglio, N.S.; Bellini, S.; Nicsanu, R.; Fostinelli, S.; et al. Serum Beta-Secretase 1 Activity Is a Potential Marker for the Differential Diagnosis between Alzheimer’s Disease and Frontotemporal Dementia: A Pilot Study. Int. J. Mol. Sci. 2024, 25, 8354. [Google Scholar] [CrossRef] [PubMed]
- Oeckl, P.; Anderl-Straub, S.; Von Arnim, C.A.F.; Baldeiras, I.; Diehl-Schmid, J.; Grimmer, T.; Halbgebauer, S.; Kort, A.M.; Lima, M.; Marques, T.M.; et al. Serum GFAP differentiates Alzheimer’s disease from frontotemporal dementia and predicts MCI-to-dementia conversion. J. Neurol. Neurosurg. Psychiatry 2022, 93, 659–667. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.R.; Shen, X.N.; Han, J.; Cui, M.; Tan, L.; Dong, Q.; Zubarev, R.A.; Yu, J.T. Deamidation-related blood biomarkers show promise for early diagnostics of neurodegeneration. Biomark. Res. 2022, 10, 91. [Google Scholar] [CrossRef]
- Oeckl, P.; Halbgebauer, S.; Anderl-Straub, S.; Steinacker, P.; Hussa, A.M.; Neugebauer, H.; Von Arnim, C.A.F.; Diehl-Schmid, J.; Grimmer, T.; Kornhuber, J.; et al. Glial Fibrillary Acidic Protein in Serum is Increased in Alzheimer’s Disease and Correlates with Cognitive Impairment. J. Alzheimer’s Dis. 2019, 67, 481–488. [Google Scholar] [CrossRef]
- Ishiki, A.; Kamada, M.; Kawamura, Y.; Terao, C.; Shimoda, F.; Tomita, N.; Arai, H.; Furukawa, K. Glial fibrillar acidic protein in the cerebrospinal fluid of Alzheimer’s disease, dementia with Lewy bodies, and frontotemporal lobar degeneration. J. Neurochem. 2016, 136, 258–261. [Google Scholar] [CrossRef]
- Abu-Rumeileh, S.; Steinacker, P.; Polischi, B.; Mammana, A.; Bartoletti-Stella, A.; Oeckl, P.; Baiardi, S.; Zenesini, C.; Huss, A.; Cortelli, P.; et al. CSF biomarkers of neuroinflammation in distinct forms and subtypes of neurodegenerative dementia. Alzheimer’s Res. Ther. 2019, 12, 2. [Google Scholar] [CrossRef]
- Woollacott, I.O.C.; Nicholas, J.M.; Heller, C.; Foiani, M.S.; Moore, K.M.; Russell, L.L.; Paterson, R.W.; Keshavan, A.; Schott, J.M.; Warren, J.D.; et al. Cerebrospinal Fluid YKL-40 and Chitotriosidase Levels in Frontotemporal Dementia Vary by Clinical, Genetic and Pathological Subtype. Dement. Geriatr. Cogn. Disord. 2020, 49, 56–76. [Google Scholar] [CrossRef]
- Antonell, A.; Tort-Merino, A.; Ríos, J.; Balasa, M.; Borrego-Écija, S.; Auge, J.M.; Muñoz-García, C.; Bosch, B.; Falgàs, N.; Rami, L.; et al. Synaptic, axonal damage and inflammatory cerebrospinal fluid biomarkers in neurodegenerative dementias. Alzheimer’s Dement. 2020, 16, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Borrego–Écija, S.; Pérez-Millan, A.; Antonell, A.; Fort-Aznar, L.; Kaya-Tilki, E.; León-Halcón, A.; Lladó, A.; Molina-Porcel, L.; Balasa, M.; Juncà-Parella, J.; et al. Galectin-3 is upregulated in frontotemporal dementia patients with subtype specificity. Alzheimer’s Dement. 2024, 20, 1515–1526. [Google Scholar] [CrossRef] [PubMed]
- del Campo, M.; Galimberti, D.; Elias, N.; Boonkamp, L.; Pijnenburg, Y.A.; van Swieten, J.C.; Watts, K.; Paciotti, S.; Beccari, T.; Hu, W.; et al. Novel CSF biomarkers to discriminate FTLD and its pathological subtypes. Ann. Clin. Transl. Neurol. 2018, 5, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Janelidze, S.; Hertze, J.; Zetterberg, H.; Landqvist Waldö, M.; Santillo, A.; Blennow, K.; Hansson, O. Cerebrospinal fluid neurogranin and YKL-40 as biomarkers of Alzheimer’s disease. Ann. Clin. Transl. Neurol. 2015, 3, 12–20. [Google Scholar] [CrossRef]
- Illán-Gala, I.; Alcolea, D.; Montal, V.; Dols-Icardo, O.; Muñoz, L.; De Luna, N.; Turón-Sans, J.; Cortés-Vicente, E.; Belén Sánchez-Saudinós, M.A.; Subirana, A.; et al. CSF sAPPβ, YKL-40, and NfL along the ALS-FTD spectrum. Neurology 2018, 91, E1619–E1628. [Google Scholar] [CrossRef]
- Alcolea, D.; Vilaplana, E.; Suárez-Calvet, M.; Illán-Gala, I.; Blesa, R.; Clarimón, J.; Lladó, A.; Sánchez-Valle, R.; Molinuevo, J.L.; García-Ribas, G.; et al. CSF sAPPβ, YKL-40, and neurofilament light in frontotemporal lobar degeneration. Neurology 2017, 89, 178–188. [Google Scholar] [CrossRef]
- Woollacott, I.O.C.; Swift, I.J.; Sogorb-Esteve, A.; Heller, C.; Knowles, K.; Bouzigues, A.; Russell, L.L.; Peakman, G.; Greaves, C.V.; Convery, R.; et al. CSF glial markers are elevated in a subset of patients with genetic frontotemporal dementia. Ann. Clin. Transl. Neurol. 2022, 9, 1764–1777. [Google Scholar] [CrossRef]
- Ullgren, A.; Öijerstedt, L.; Olofsson, J.; Bergström, S.; Remnestål, J.; van Swieten, J.C.; Jiskoot, L.C.; Seelaar, H.; Borroni, B.; Sanchez-Valle, R.; et al. Altered plasma protein profiles in genetic FTD—A GENFI study. Mol. Neurodegener. 2023, 18, 85. [Google Scholar] [CrossRef]
- Meda, F.; Simrén, J.; Borroni, B.; Cantoni, V.; Archetti, S.; Biasiotto, G.; Andreasson, U.; Blennow, K.; Kvartsberg, H.; Zetterberg, H. Analytical and clinical validation of a blood progranulin ELISA in frontotemporal dementias. Clin. Chem. Lab. Med. 2023, 61, 2195–2204. [Google Scholar] [CrossRef]
- Finch, N.; Baker, M.; Crook, R.; Swanson, K.; Kuntz, K.; Surtees, R.; Bisceglio, G.; Rovelet-Lecrux, A.; Boeve, B.; Petersen, R.C.; et al. Plasma progranulin levels predict progranulin mutation status in frontotemporal dementia patients and asymptomatic family members. Brain 2009, 132, 583–591. [Google Scholar] [CrossRef]
- Gibbons, L.; Rollinson, S.; Thompson, J.C.; Robinson, A.; Davidson, Y.S.; Richardson, A.; Neary, D.; Pickering-Brown, S.M.; Snowden, J.S.; Mann, D.M.A. Plasma levels of progranulin and interleukin-6 in frontotemporal lobar degeneration. Neurobiol. Aging 2015, 36, e1601–e1603. [Google Scholar] [CrossRef]
- Ghidoni, R.; Flocco, R.; Paterlini, A.; Glionna, M.; Caruana, L.; Tonoli, E.; Binetti, G.; Benussi, L. Secretory leukocyte protease inhibitor protein regulates the penetrance of frontotemporal lobar degeneration in progranulin mutation carriers. J. Alzheimer’s Dis. 2014, 38, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Sogorb-Esteve, A.; Swift, I.J.; Woollacott, I.O.C.; Warren, J.D.; Zetterberg, H.; Rohrer, J.D. Differential chemokine alteration in the variants of primary progressive aphasia-a role for neuroinflammation. J. Neuroinflamm. 2021, 18, 224. [Google Scholar] [CrossRef]
- Boström, G.; Freyhult, E.; Virhammar, J.; Alcolea, D.; Tumani, H.; Otto, M.; Brundin, R.M.; Kilander, L.; Löwenmark, M.; Giedraitis, V.; et al. Different Inflammatory Signatures in Alzheimer’s Disease and Frontotemporal Dementia Cerebrospinal Fluid. J. Alzheimer’s Dis. 2021, 81, 629–640. [Google Scholar] [CrossRef]
- Minthon, L.; Edvinsson, L.; Gustafson, L. Somatostatin and neuropeptide Y in cerebrospinal fluid: Correlations with severity of disease and clinical signs in Alzheimer’s disease and frontotemporal dementia. Dement. Geriatr. Cogn. Disord. 1997, 8, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Maetzler, W.; Berg, D.; Synofzik, M.; Brockmann, K.; Godau, J.; Melms, A.; Gasser, T.; Hörnig, S.; Langkamp, M. Autoantibodies against amyloid and glial-derived antigens are increased in serum and cerebrospinal fluid of Lewy body-associated dementias. J. Alzheimer’s Dis. 2011, 26, 171–179. [Google Scholar] [CrossRef]
- Zhou, J.; Wade, S.D.; Graykowski, D.; Xiao, M.F.; Zhao, B.; Giannini, L.A.A.; Hanson, J.E.; van Swieten, J.C.; Sheng, M.; Worley, P.F.; et al. The neuronal pentraxin Nptx2 regulates complement activity and restrains microglia-mediated synapse loss in neurodegeneration. Sci. Transl. Med. 2023, 15, eadf0141. [Google Scholar] [CrossRef]
- Sudre, C.H.; Bocchetta, M.; Heller, C.; Convery, R.; Neason, M.; Moore, K.M.; Cash, D.M.; Thomas, D.L.; Woollacott, I.O.C.; Foiani, M.; et al. White matter hyperintensities in progranulin-associated frontotemporal dementia: A longitudinal GENFI study. NeuroImage Clin. 2019, 24, 102077. [Google Scholar] [CrossRef]
- Roos, P.; von Essen, M.R.; Nielsen, T.T.; Johannsen, P.; Stokholm, J.; Bie, A.S.; Waldemar, G.; Simonsen, A.H.; Heslegrave, A.; Zetterberg, H.; et al. Inflammatory markers of CHMP2B-mediated frontotemporal dementia. J. Neuroimmunol. 2018, 324, 136–142. [Google Scholar] [CrossRef]
- Galimberti, D.; Bonsi, R.; Fenoglio, C.; Serpente, M.; Cioffi, S.M.G.; Fumagalli, G.; Arighi, A.; Ghezzi, L.; Arcaro, M.; Mercurio, M.; et al. Inflammatory molecules in Frontotemporal Dementia: Cerebrospinal fluid signature of progranulin mutation carriers. Brain Behav. Immun. 2015, 49, 182–187. [Google Scholar] [CrossRef]
- Galimberti, D.; Schoonenboom, N.; Scheltens, P.; Fenoglio, C.; Venturelli, E.; Pijnenburg, Y.A.L.; Bresolin, N.; Scarpini, E. Intrathecal chemokine levels in Alzheimer disease and frontotemporal lobar degeneration. Neurology 2006, 66, 146–147. [Google Scholar] [CrossRef]
- Blasko, I.; Lederer, W.; Oberbauer, H.; Walch, T.; Kemmler, G.; Hinterhuber, H.; Marksteiner, J.; Humpel, C. Measurement of thirteen biological markers in CSF of patients with Alzheimer’s disease and other dementias. Dement. Geriatr. Cogn. Disord. 2006, 21, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Magdalinou, N.K.; Paterson, R.W.; Schott, J.M.; Fox, N.C.; Mummery, C.; Blennow, K.; Bhatia, K.; Morris, H.R.; Giunti, P.; Warner, T.T.; et al. A panel of nine cerebrospinal fluid biomarkers may identify patients with atypical parkinsonian syndromes. J. Neurol. Neurosurg. Psychiatry 2015, 86, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Alcolea, D.; Carmona-Iragui, M.; Suárez-Calvet, M.; Sánchez-Saudinós, M.B.; Sala, I.; Antón-Aguirre, S.; Blesa, R.; Clarimón, J.; Fortea, J.; Lleó, A. Relationship between β-Secretase, inflammation and core cerebrospinal fluid biomarkers for Alzheimer’s disease. J. Alzheimer’s Dis. 2014, 42, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Craig-Schapiro, R.; Perrin, R.J.; Roe, C.M.; Xiong, C.; Carter, D.; Cairns, N.J.; Mintun, M.A.; Peskind, E.R.; Li, G.; Galasko, D.R.; et al. YKL-40: A novel prognostic fluid biomarker for preclinical Alzheimer’s disease. Biol. Psychiatry 2010, 68, 903–912. [Google Scholar] [CrossRef]
- Khoonsari, P.E.; Shevchenko, G.; Herman, S.; Remnestal, J.; Giedraitis, V.; Brundin, R.; Gunnarsson, M.D.; Kilander, L.; Zetterberg, H.; Nilsson, P.; et al. Improved Differential Diagnosis of Alzheimer’s Disease by Integrating ELISA and Mass Spectrometry-Based Cerebrospinal Fluid Biomarkers. J. Alzheimer’s Dis. 2019, 67, 639–651. [Google Scholar] [CrossRef]
- Lok, H.C.; Katzeff, J.S.; Hodges, J.R.; Piguet, O.; Fu, Y.H.; Halliday, G.M.; Kim, W.S. Elevated GRO-α and IL-18 in serum and brain implicate the NLRP3 inflammasome in frontotemporal dementia. Sci. Rep. 2023, 13, 8942. [Google Scholar] [CrossRef]
- Bergström, S.; Öijerstedt, L.; Remnestål, J.; Olofsson, J.; Ullgren, A.; Seelaar, H.; van Swieten, J.C.; Synofzik, M.; Sanchez-Valle, R.; Moreno, F.; et al. A panel of CSF proteins separates genetic frontotemporal dementia from presymptomatic mutation carriers: A GENFI study. Mol. Neurodegener. 2021, 16, 79. [Google Scholar] [CrossRef]
- Toft, A.; Sjödin, S.; Simonsen, A.H.; Ejlerskov, P.; Roos, P.; Musaeus, C.S.; Henriksen, E.E.; Nielsen, T.T.; Brinkmalm, A.; Blennow, K.; et al. Endo-lysosomal protein concentrations in CSF from patients with frontotemporal dementia caused by CHMP2B mutation. Alzheimer’s Dement. 2023, 15, e12402. [Google Scholar] [CrossRef]
- Huang, M.; Modeste, E.; Dammer, E.; Merino, P.; Taylor, G.; Duong, D.M.; Deng, Q.; Holler, C.J.; Gearing, M.; Dickson, D.; et al. Network analysis of the progranulin-deficient mouse brain proteome reveals pathogenic mechanisms shared in human frontotemporal dementia caused by GRN mutations. Acta Neuropathol. Commun. 2020, 8, 163. [Google Scholar] [CrossRef]
- Taipa, R.; das Neves, S.P.; Sousa, A.L.; Fernandes, J.; Pinto, C.; Correia, A.P.; Santos, E.; Pinto, P.S.; Carneiro, P.; Costa, P.; et al. Proinflammatory and anti-inflammatory cytokines in the CSF of patients with Alzheimer’s disease and their correlation with cognitive decline. Neurobiol. Aging 2019, 76, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Sogorb-Esteve, A.; Colas, R.A.; Dalli, J.; Rohrer, J.D. Differential Lipid Mediator Involvement in the Different Forms of Genetic Frontotemporal Dementia: Novel Insights into Neuroinflammation. J. Alzheimer’s Dis. 2021, 84, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Abu-Rumeileh, S.; Halbgebauer, S.; Steinacker, P.; Anderl-Straub, S.; Polischi, B.; Ludolph, A.C.; Capellari, S.; Parchi, P.; Otto, M. CSF SerpinA1 in Creutzfeldt–Jakob disease and frontotemporal lobar degeneration. Ann. Clin. Transl. Neurol. 2020, 7, 191–199. [Google Scholar] [CrossRef]
- Sjögren, M.; Folkesson, S.; Blennow, K.; Tarkowski, E. Increased intrathecal inflammatory activity in frontotemporal dementia: Pathophysiological implications. J. Neurol. Neurosurg. Psychiatry 2004, 75, 1107–1111. [Google Scholar] [CrossRef]
- van der Ende, E.L.; Morenas-Rodriguez, E.; McMillan, C.; Grossman, M.; Irwin, D.; Sanchez-Valle, R.; Graff, C.; Vandenberghe, R.; Pijnenburg, Y.A.L.; Laforce, R.; et al. CSF sTREM2 is elevated in a subset in GRN-related frontotemporal dementia. Neurobiol. Aging 2021, 103, e151–e158. [Google Scholar] [CrossRef]
- Heywood, W.E.; Hallqvist, J.; Heslegrave, A.J.; Zetterberg, H.; Fenoglio, C.; Scarpini, E.; Rohrer, J.D.; Galimberti, D.; Mills, K. CSF pro-orexin and amyloid-β38 expression in Alzheimer’s disease and frontotemporal dementia. Neurobiol. Aging 2018, 72, 171–176. [Google Scholar] [CrossRef]
- Busse, S.; Hoffmann, J.; Michler, E.; Hartig, R.; Frodl, T.; Busse, M. Dementia-associated changes of immune cell composition within the cerebrospinal fluid. Brain Behav. Immun. Health 2021, 14, 100218. [Google Scholar] [CrossRef]
- Philips, T.; De Muynck, L.; Thu, H.N.T.; Weynants, B.; Vanacker, P.; Dhondt, J.; Sleegers, K.; Schelhaas, H.J.; Verbeek, M.; Vandenberghe, R.; et al. Microglial upregulation of progranulin as a marker of motor neuron degeneration. J. Neuropathol. Exp. Neurol. 2010, 69, 1191–1200. [Google Scholar] [CrossRef]
- Kankaanpää, J.; Turunen, S.P.; Moilanen, V.; Hörkkö, S.; Remes, A.M. Cerebrospinal fluid antibodies to oxidized LDL are increased in Alzheimer’s disease. Neurobiol. Dis. 2009, 33, 467–472. [Google Scholar] [CrossRef]
- Jesse, S.; Brettschneider, J.; Süssmuth, S.D.; Landwehrmeyer, B.G.; Von Arnim, C.A.F.; Ludolph, A.C.; Tumani, H.; Otto, M. Summary of cerebrospinal fluid routine parameters in neurodegenerative diseases. J. Neurol. 2011, 258, 1034–1041. [Google Scholar] [CrossRef]
- van der Ende, E.L.; Meeter, L.H.; Stingl, C.; van Rooij, J.G.J.; Stoop, M.P.; Nijholt, D.A.T.; Sanchez-Valle, R.; Graff, C.; Öijerstedt, L.; Grossman, M.; et al. Novel CSF biomarkers in genetic frontotemporal dementia identified by proteomics. Ann. Clin. Transl. Neurol. 2019, 6, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Benito, P.; Povedano, M.; Domínguez, R.; Marco, C.; Colomina, M.J.; López-Pérez, Ó.; Santana, I.; Baldeiras, I.; Martínez-Yelámos, S.; Zerr, I.; et al. Increased C-X-C Motif Chemokine Ligand 12 Levels in Cerebrospinal Fluid as a Candidate Biomarker in Sporadic Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2020, 21, 8680. [Google Scholar] [CrossRef]
- Bruggink, K.A.; Kuiperij, H.B.; Gloerich, J.; Otte-Höller, I.; Rozemuller, A.J.M.; Claassen, J.A.H.R.; Küsters, B.; Verbeek, M.M. Dickkopf-related protein 3 is a potential Aβ-associated protein in Alzheimer’s Disease. J. Neurochem. 2015, 134, 1152–1162. [Google Scholar] [CrossRef]
- Janelidze, S.; Hertze, J.; Nägga, K.; Nilsson, K.; Nilsson, C.; Wennström, M.; van Westen, D.; Blennow, K.; Zetterberg, H.; Hansson, O. Increased blood-brain barrier permeability is associated with dementia and diabetes but not amyloid pathology or APOE genotype. Neurobiol. Aging 2017, 51, 104–112. [Google Scholar] [CrossRef] [PubMed]
- González-Sánchez, M.; Jiménez, J.; Narváez, A.; Antequera, D.; Llamas-Velasco, S.; Martín, A.H.S.; Arjona, J.A.M.; Munain, A.L.d.; Bisa, A.L.; Marco, M.P.; et al. Kynurenic Acid Levels are Increased in the CSF of Alzheimer’s Disease Patients. Biomolecules 2020, 10, 571. [Google Scholar] [CrossRef]
- Phan, K.; He, Y.; Pickford, R.; Bhatia, S.; Katzeff, J.S.; Hodges, J.R.; Piguet, O.; Halliday, G.M.; Kim, W.S. Uncovering pathophysiological changes in frontotemporal dementia using serum lipids. Sci. Rep. 2020, 10, 3640. [Google Scholar] [CrossRef]
- Cai, H.; Zhao, T.; Pang, Y.; Fu, X.; Ren, Z.; Quan, S.; Jia, L. Systemic inflammatory markers in ageing, Alzheimer’s disease and other dementias. Brain 2024, 148, 480–492. [Google Scholar] [CrossRef]
- Katzeff, J.S.; Bright, F.; Lo, K.; Kril, J.J.; Connolly, A.; Crossett, B.; Ittner, L.M.; Kassiou, M.; Loy, C.T.; Hodges, J.R.; et al. Altered serum protein levels in frontotemporal dementia and amyotrophic lateral sclerosis indicate calcium and immunity dysregulation. Sci. Rep. 2020, 10, 13741. [Google Scholar] [CrossRef] [PubMed]
- Bellini, S.; Saraceno, C.; Benussi, L.; Squitti, R.; Cimini, S.; Ricci, M.; Canafoglia, L.; Coppola, C.; Puoti, G.; Ferrari, C.; et al. Plasma Small Extracellular Vesicles with Complement Alterations in GRN/C9orf72 and Sporadic Frontotemporal Lobar Degeneration. Cells 2022, 11, 488. [Google Scholar] [CrossRef]
- Sanchez, E.; Wilkinson, T.; Coughlan, G.; Mirza, S.; Baril, A.A.; Ramirez, J.; Binns, M.A.; Black, S.E.; Borrie, M.; Dilliott, A.A.; et al. Association of plasma biomarkers with cognition, cognitive decline, and daily function across and within neurodegenerative diseases: Results from the Ontario Neurodegenerative Disease Research Initiative. Alzheimer’s Dement. 2024, 20, 1753–1770. [Google Scholar] [CrossRef]
- Fraga, V.G.; Ferreira, C.N.; Oliveira, F.R.; Cândido, A.L.; das Graças Carvalho, M.; Reis, F.M.; Caramelli, P.; De Souza, L.C.; Gomes, K.B. Irisin levels are correlated with inflammatory markers in frontotemporal dementia. J. Clin. Neurosci. 2021, 93, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Wen, L.; Jiang, D.; Liu, L.; Nan, H.; Yue, A.; Wang, Y.; Wang, Y.; Qu, M.; Wang, N.; et al. Peripheral inflammation in behavioural variant frontotemporal dementia: Associations with central degeneration and clinical measures. J. Neuroinflamm. 2023, 20, 65. [Google Scholar] [CrossRef]
- Asken, B.M.; Ljubenkov, P.A.; Staffaroni, A.M.; Casaletto, K.B.; Vandevrede, L.; Cobigo, Y.; Rojas-Rodriguez, J.C.; Rankin, K.P.; Kornak, J.; Heuer, H.; et al. Plasma inflammation for predicting phenotypic conversion and clinical progression of autosomal dominant frontotemporal lobar degeneration. J. Neurol. Neurosurg. Psychiatry 2023, 94, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Vieira, É.L.M.; Caramelli, P.; Rocha, N.P.; Freitas Cardoso, M.d.G.; de Miranda, A.S.; Teixeira, A.L.; de Souza, L.C. Tumor necrosis factor superfamily molecules are increased in behavioral variant frontotemporal dementia and correlate with cortical atrophy: An exploratory investigation. J. Neuroimmunol. 2021, 354, 577531. [Google Scholar] [CrossRef]
- Miller, Z.A.; Rankin, K.P.; Graff-Radford, N.R.; Takada, L.T.; Sturm, V.E.; Cleveland, C.M.; Criswell, L.A.; Jaeger, P.A.; Stan, T.; Heggeli, K.A.; et al. TDP-43 frontotemporal lobar degeneration and autoimmune disease. J. Neurol. Neurosurg. Psychiatry 2013, 84, 956–962. [Google Scholar] [CrossRef]
- Olğun, Y.; Aksoy Poyraz, C.; Bozluolçay, M.; Konukoğlu, D.; Poyraz, B.Ç. Plasma Biomarkers in Neurodegenerative Dementias: Unrevealing the Potential of Serum Oxytocin, BDNF, NPTX1, TREM2, TNF-alpha, IL-1 and Prolactin. Curr. Alzheimer Res. 2024, 21, 109–119. [Google Scholar] [CrossRef]
- Cavazzana, I.; Alberici, A.; Bonomi, E.; Ottaviani, R.; Kumar, R.; Archetti, S.; Manes, M.; Cosseddu, M.; Buratti, E.; Padovani, A.; et al. Antinuclear antibodies in Frontotemporal Dementia: The tip’s of autoimmunity iceberg? J. Neuroimmunol. 2018, 325, 61–63. [Google Scholar] [CrossRef]
- Bossolasco, P.; Cancello, R.; Doretti, A.; Morelli, C.; Silani, V.; Cova, L. Adiponectin levels in the serum and cerebrospinal fluid of amyotrophic lateral sclerosis patients: Possible influence on neuroinflammation? J. Neuroinflamm. 2017, 14, 85. [Google Scholar] [CrossRef]
- Ljubenkov, P.A.; Miller, Z.; Mumford, P.; Zhang, J.; Allen, I.E.; Mitic, L.; Staffaroni, A.; Heuer, H.; Rojas, J.C.; Cobigo, Y.; et al. Peripheral Innate Immune Activation Correlates With Disease Severity in GRN Haploinsufficiency. Front. Neurol. 2019, 10, 1004. [Google Scholar] [CrossRef]
- Phan, K.; He, Y.; Fu, Y.H.; Dzamko, N.; Bhatia, S.; Gold, J.; Rowe, D.; Ke, Y.D.; Ittner, L.M.; Hodges, J.R.; et al. Pathological manifestation of human endogenous retrovirus K in frontotemporal dementia. Commun. Med. 2021, 1, 60. [Google Scholar] [CrossRef]
- Villar-Piqué, A.; Schmitz, M.; Hermann, P.; Goebel, S.; Bunck, T.; Varges, D.; Ferrer, I.; Riggert, J.; Llorens, F.; Zerr, I. Plasma YKL-40 in the spectrum of neurodegenerative dementia. J. Neuroinflamm. 2019, 16, 145. [Google Scholar] [CrossRef]
- Dols-Icardo, O.; Suárez-Calvet, M.; Hernández, I.; Amer, G.; Antón-Aguirre, S.; Alcolea, D.; Fortea, J.; Boada, M.; Tárraga, L.; Blesa, R.; et al. Expansion mutation in C9ORF72 does not influence plasma progranulin levels in frontotemporal dementia. Neurobiol. Aging 2012, 33, e1817–e1851. [Google Scholar] [CrossRef]
- Schröder, S.; Heck, J.; Groh, A.; Frieling, H.; Bleich, S.; Kahl, K.G.; Bosch, J.J.; Krichevsky, B.; Schulze-Westhoff, M. White Blood Cell and Platelet Counts Are Not Suitable as Biomarkers in the Differential Diagnostics of Dementia. Brain Sci. 2022, 12, 1424. [Google Scholar] [CrossRef]
- Derkow, K.; Rössling, R.; Schipke, C.; Krüger, C.; Bauer, J.; Fähling, M.; Stroux, A.; Schott, E.; Ruprecht, K.; Peters, O.; et al. Distinct expression of the neurotoxic microRNA family let-7 in the cerebrospinal fluid of patients with Alzheimer’s disease. PLoS ONE 2018, 13, e0200602. [Google Scholar] [CrossRef]
- Sheinerman, K.S.; Toledo, J.B.; Tsivinsky, V.G.; Irwin, D.; Grossman, M.; Weintraub, D.; Hurtig, H.I.; Chen-Plotkin, A.; Wolk, D.A.; McCluskey, L.F.; et al. Circulating brain-enriched microRNAs as novel biomarkers for detection and differentiation of neurodegenerative diseases. Alzheimer’s Res. Ther. 2017, 9, 89. [Google Scholar] [CrossRef]
- Conti, E.; Sala, G.; Diamanti, S.; Casati, M.; Lunetta, C.; Gerardi, F.; Tarlarini, C.; Mosca, L.; Riva, N.; Falzone, Y.; et al. Serum naturally occurring anti-TDP-43 auto-antibodies are increased in amyotrophic lateral sclerosis. Sci. Rep. 2021, 11, 1978. [Google Scholar] [CrossRef]
- Katisko, K.; Kokkonen, N.; Krüger, J.; Hartikainen, P.; Koivisto, A.M.; Helisalmi, S.; Korhonen, V.E.; Kokki, M.; Tuusa, J.; Herukka, S.K.; et al. The Association Between Frontotemporal Lobar Degeneration and Bullous Pemphigoid. J. Alzheimer’s Dis. 2018, 66, 743–750. [Google Scholar] [CrossRef]
- Heikkinen, S.; Huber, N.; Katisko, K.; Kokkola, T.; Hartikainen, P.; Krüger, J.; Leinonen, V.; Korhonen, V.E.; Herukka, S.K.; Remes, A.M.; et al. Serum Cathepsin S Levels Do Not Show Alterations in Different Clinical, Neuropathological, or Genetic Subtypes of Frontotemporal Dementia Patients nor in Comparison to Healthy Control Individuals. J. Alzheimer’s Dis. 2023, 93, 395–401. [Google Scholar] [CrossRef]
- De Luna, N.; Carbayo, Á.; Dols-Icardo, O.; Turon-Sans, J.; Reyes-Leiva, D.; Illan-Gala, I.; Jericó, I.; Pagola-Lorz, I.; Lleixà, C.; Querol, L.; et al. Neuroinflammation-Related Proteins NOD2 and Spp1 Are Abnormally Upregulated in Amyotrophic Lateral Sclerosis. Neurol. Neuroimmunol. Neuroinflamm. 2022, 10, e200072. [Google Scholar] [CrossRef]
- Thijssen, E.H.; Verberk, I.M.W.; Kindermans, J.; Abramian, A.; Vanbrabant, J.; Ball, A.J.; Pijnenburg, Y.; Lemstra, A.W.; van der Flier, W.M.; Stoops, E.; et al. Differential diagnostic performance of a panel of plasma biomarkers for different types of dementia. Alzheimer’s Dement. 2022, 14, e12285. [Google Scholar] [CrossRef]
- Rentzos, M.; Zoga, M.; Paraskevas, G.P.; Kapaki, E.; Rombos, A.; Nikolaou, C.; Tsoutsou, A.; Vassilopoulos, D. IL-15 is elevated in cerebrospinal fluid of patients with Alzheimer’s disease and frontotemporal dementia. J. Geriatr. Psychiatry Neurol. 2006, 19, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Cai, Z.Y.; Sun, X.H.; Shen, D.C.; Yang, X.Z.; Liu, M.S.; Cui, L.Y. Blood-brain barrier dysfunction and myelin basic protein in survival of amyotrophic lateral sclerosis with or without frontotemporal dementia. Neurol. Sci. 2022, 43, 3201–3210. [Google Scholar] [CrossRef]
- Rascovsky, K.; Hodges, J.R.; Knopman, D.; Mendez, M.F.; Kramer, J.H.; Neuhaus, J.; van Swieten, J.C.; Seelaar, H.; Dopper, E.G.; Onyike, C.U.; et al. Sensitivity of revised diagnostic criteria for the behavioural variant of frontotemporal dementia. Brain 2011, 134, 2456–2477. [Google Scholar] [CrossRef]
- Gorno-Tempini, M.L.; Hillis, A.E.; Weintraub, S.; Kertesz, A.; Mendez, M.; Cappa, S.F.; Ogar, J.M.; Rohrer, J.D.; Black, S.; Boeve, B.F.; et al. Classification of primary progressive aphasia and its variants. Neurology 2011, 76, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Cairns, N.J.; Bigio, E.H.; Mackenzie, I.R.A.; Neumann, M.; Lee, V.M.Y.; Hatanpaa, K.J.; White, C.L.; Schneider, J.A.; Grinberg, L.T.; Halliday, G.; et al. Neuropathologic diagnostic and nosologic criteria for frontotemporal lobar degeneration: Consensus of the Consortium for Frontotemporal Lobar Degeneration. Acta Neuropathol. 2007, 114, 5–22. [Google Scholar] [CrossRef]
- McKhann, G.M.; Albert, M.S.; Grossman, M.; Miller, B.; Dickson, D.; Trojanowski, J.Q. Clinical and pathological diagnosis of frontotemporal dementia: Report of the Work Group on Frontotemporal Dementia and Pick’s Disease. Arch. Neurol. 2001, 58, 1803–1809. [Google Scholar] [CrossRef] [PubMed]
- Neary, D.; Snowden, J.S.; Gustafson, L.; Passant, U.; Stuss, D.; Black, S.; Freedman, M.; Kertesz, A.; Robert, P.H.; Albert, M.; et al. Frontotemporal lobar degeneration: A consensus on clinical diagnostic criteria. Neurology 1998, 51, 1546–1554. [Google Scholar] [CrossRef]
- Clinical and neuropathological criteria for frontotemporal dementia. The Lund and Manchester Groups. J. Neurol. Neurosurg. Psychiatry 1994, 57, 416–418. [Google Scholar] [CrossRef]
- Alcolea, D.; Irwin, D.J.; Illán-Gala, I.; Muñoz, L.; Clarimón, J.; McMillan, C.T.; Fortea, J.; Blesa, R.; Lee, E.B.; Trojanowski, J.Q.; et al. Elevated YKL-40 and low sAPPβ:YKL-40 ratio in antemortem cerebrospinal fluid of patients with pathologically confirmed FTLD. J. Neurol. Neurosurg. Psychiatry 2019, 90, 180–186. [Google Scholar] [CrossRef]
- Chouliaras, L.; Thomas, A.; Malpetti, M.; Donaghy, P.; Kane, J.; Mak, E.; Savulich, G.; Prats-Sedano, M.A.; Heslegrave, A.J.; Zetterberg, H.; et al. Differential levels of plasma biomarkers of neurodegeneration in Lewy body dementia, Alzheimer’s disease, frontotemporal dementia and progressive supranuclear palsy. J. Neurol. Neurosurg. Psychiatry 2022, 93, 651–658. [Google Scholar] [CrossRef]
- Gómez-Tortosa, E.; Guerrero-López, R.; Gil-Neciga, E.; Franco, E.; del Ser, T.; Jiménez Escrig, A.; Pérez-Pérez, J.; Sainz, M.J. Plasma progranulin levels in cortical dementia phenotypes with asymmetric perisylvian atrophy. Eur. J. Neurol. 2013, 20, 1319–1324. [Google Scholar] [CrossRef] [PubMed]
- Mohaupt, P.; Kindermans, J.; Vialaret, J.; Anderl-Straub, S.; Werner, L.; Lehmann, S.; Hirtz, C.; Otto, M.; Oeckl, P. Blood-based biomarkers and plasma Aβ assays in the differential diagnosis of Alzheimer’s disease and behavioral-variant frontotemporal dementia. Alzheimers Res. Ther. 2024, 16, 279. [Google Scholar] [CrossRef]
- Hu, W.T.; Chen-Plotkin, A.; Grossman, M.; Arnold, S.E.; Clark, C.M.; Shaw, L.M.; McCluskey, L.; Elman, L.; Hurtig, H.I.; Siderowf, A.; et al. Novel CSF biomarkers for frontotemporal lobar degenerations. Neurology 2010, 75, 2079–2086. [Google Scholar] [CrossRef]
- Galimberti, D.; Venturelli, E.; Villa, C.; Fenoglio, C.; Clerici, F.; Marcone, A.; Benussi, L.; Cortini, F.; Scalabrini, D.; Perini, L.; et al. MCP-1 A-2518G polymorphism: Effect on susceptibility for frontotemporal lobar degeneration and on cerebrospinal fluid MCP-1 levels. J. Alzheimer’s Dis. 2009, 17, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Teunissen, C.E.; Elias, N.; Koel-Simmelink, M.J.A.; Durieux-Lu, S.; Malekzadeh, A.; Pham, T.V.; Piersma, S.R.; Beccari, T.; Meeter, L.H.H.; Dopper, E.G.P.; et al. Novel diagnostic cerebrospinal fluid biomarkers for pathologic subtypes of frontotemporal dementia identified by proteomics. Alzheimer’s Dement. 2016, 2, 86–94. [Google Scholar] [CrossRef]
- Katisko, K.; Solje, E.; Korhonen, P.; Jääskeläinen, O.; Loppi, S.; Hartikainen, P.; Koivisto, A.M.; Kontkanen, A.; Korhonen, V.E.; Helisalmi, S.; et al. Peripheral inflammatory markers and clinical correlations in patients with frontotemporal lobar degeneration with and without the C9orf72 repeat expansion. J. Neurol. 2020, 267, 76–86. [Google Scholar] [CrossRef]
- Galimberti, D.; Venturelli, E.; Fenoglio, C.; Guidi, I.; Villa, C.; Bergamaschini, L.; Cortini, F.; Scalabrini, D.; Baron, P.; Vergani, C.; et al. Intrathecal levels of IL-6, IL-11 and LIF in Alzheimer’s disease and frontotemporal lobar degeneration. J. Neurol. 2008, 255, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Feneberg, E.; Steinacker, P.; Volk, A.E.; Weishaupt, J.H.; Wollmer, M.A.; Boxer, A.; Tumani, H.; Ludolph, A.C.; Otto, M. Progranulin as a candidate biomarker for therapeutic trial in patients with ALS and FTLD. J. Neural Transm. 2016, 123, 289–296. [Google Scholar] [CrossRef]
- Petzold, A.; Jenkins, R.; Watt, H.C.; Green, A.J.E.; Thompson, E.J.; Keir, G.; Fox, N.C.; Rossor, M.N. Cerebrospinal fluid S100B correlates with brain atrophy in Alzheimer’s disease. Neurosci. Lett. 2003, 336, 167–170. [Google Scholar] [CrossRef]
- Alexopoulos, P.; Guo, L.H.; Tsolakidou, A.; Kratzer, M.; Grimmer, T.; Westerteicher, C.; Jiang, M.; Bujo, H.; Diehl-Schmid, J.; Kurz, A.; et al. Interrelations between CSF soluble AβPPβ, amyloid-β 1-42, SORL1, and tau levels in Alzheimer’s disease. J. Alzheimer’s Dis. 2012, 28, 543–552. [Google Scholar] [CrossRef]
- Sjögren, M.; Wallin, A. Pathophysiological aspects of frontotemporal dementia—Emphasis on cytoskeleton proteins and autoimmunity. Mech. Ageing Dev. 2001, 122, 1923–1935. [Google Scholar] [CrossRef] [PubMed]
- Woollacott, I.O.C.; Nicholas, J.M.; Heslegrave, A.; Heller, C.; Foiani, M.S.; Dick, K.M.; Russell, L.L.; Paterson, R.W.; Keshavan, A.; Fox, N.C.; et al. Cerebrospinal fluid soluble TREM2 levels in frontotemporal dementia differ by genetic and pathological subgroup. Alzheimer’s Res. Ther. 2018, 10, 79. [Google Scholar] [CrossRef]
- Piscopo, P.; Rivabene, R.; Galimberti, D.; Crestini, A.; Talarico, G.; Vanacore, N.; Scarpini, E.; Bruno, G.; Confaloni, A. Gender effects on plasma PGRN levels in patients with Alzheimer’s disease: A preliminary study. J. Alzheimer’s Dis. 2013, 35, 313–318. [Google Scholar] [CrossRef]
- Sleegers, K.; Brouwers, N.; Van Damme, P.; Engelborghs, S.; Gijselinck, I.; Van Der Zee, J.; Peeters, K.; Mattheijssens, M.; Cruts, M.; Vandenberghe, R.; et al. Serum biomarker for progranulin-associated frontotemporal lobar degeneration. Ann. Neurol. 2009, 65, 603–609. [Google Scholar] [CrossRef]
- Bossù, P.; Salani, F.; Alberici, A.; Archetti, S.; Bellelli, G.; Galimberti, D.; Scarpini, E.; Spalletta, G.; Caltagirone, C.; Padovani, A.; et al. Loss of function mutations in the progranulin gene are related to pro-inflammatory cytokine dysregulation in frontotemporal lobar degeneration patients. J. Neuroinflamm. 2011, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Triviño, J.J.; von Bernhardi, R. The effect of aged microglia on synaptic impairment and its relevance in neurodegenerative diseases. Neurochem. Int. 2021, 144, 104982. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, M.; Mollenhauer, B.; Bertolotto, A.; Engelborghs, S.; Hampel, H.; Simonsen, A.H.; Kapaki, E.; Kruse, N.; Le Bastard, N.; Lehmann, S.; et al. Recommendations to standardize preanalytical confounding factors in Alzheimer’s and Parkinson’s disease cerebrospinal fluid biomarkers: An update. Biomark. Med. 2012, 6, 419–430. [Google Scholar] [CrossRef]
- Teunissen, C.E.; Petzold, A.; Bennett, J.L.; Berven, F.S.; Brundin, L.; Comabella, M.; Franciotta, D.; Frederiksen, J.L.; Fleming, J.O.; Furlan, R.; et al. A consensus protocol for the standardization of cerebrospinal fluid collection and biobanking. Neurology 2009, 73, 1914–1922. [Google Scholar] [CrossRef]
- Abdelhak, A.; Hottenrott, T.; Morenas-Rodríguez, E.; Suárez-Calvet, M.; Zettl, U.K.; Haass, C.; Meuth, S.G.; Rauer, S.; Otto, M.; Tumani, H.; et al. Glial Activation Markers in CSF and Serum From Patients With Primary Progressive Multiple Sclerosis: Potential of Serum GFAP as Disease Severity Marker? Front. Neurol. 2019, 10, 280. [Google Scholar] [CrossRef]
- Nakao, A. Temporal regulation of cytokines by the circadian clock. J. Immunol. Res. 2014, 2014, 614529. [Google Scholar] [CrossRef]
- Yadav, A.; Saini, V.; Arora, S. MCP-1: Chemoattractant with a role beyond immunity: A review. Clin. Chim. Acta 2010, 411, 1570–1579. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Liao, Y.C.; Wang, Y.F.; Lin, I.F.; Wang, S.J.; Fuh, J.L. Plasma MCP-1 and Cognitive Decline in Patients with Alzheimer’s Disease and Mild Cognitive Impairment: A Two-year Follow-up Study. Sci. Rep. 2018, 8, 1280. [Google Scholar] [CrossRef]
- Sawant, K.V.; Poluri, K.M.; Dutta, A.K.; Sepuru, K.M.; Troshkina, A.; Garofalo, R.P.; Rajarathnam, K. Chemokine CXCL1 mediated neutrophil recruitment: Role of glycosaminoglycan interactions. Sci. Rep. 2016, 6, 33123. [Google Scholar] [CrossRef]
- Yan, Y.; Chen, R.; Wang, X.; Hu, K.; Huang, L.; Lu, M.; Hu, Q. CCL19 and CCR7 Expression, Signaling Pathways, and Adjuvant Functions in Viral Infection and Prevention. Front. Cell Dev. Biol. 2019, 7, 212. [Google Scholar] [CrossRef]
- Boza-Serrano, A.; Vrillon, A.; Minta, K.; Paulus, A.; Camprubí-Ferrer, L.; Garcia, M.; Andreasson, U.; Antonell, A.; Wennström, M.; Gouras, G.; et al. Galectin-3 is elevated in CSF and is associated with Aβ deposits and tau aggregates in brain tissue in Alzheimer’s disease. Acta Neuropathol. 2022, 144, 843–859. [Google Scholar] [CrossRef] [PubMed]
- Hok-A-Hin, Y.S.; Hoozemans, J.J.M.; Hu, W.T.; Wouters, D.; Howell, J.C.; Rábano, A.; van der Flier, W.M.; Pijnenburg, Y.A.L.; Teunissen, C.E.; Del Campo, M. YKL-40 changes are not detected in post-mortem brain of patients with Alzheimer’s disease and frontotemporal lobar degeneration. Alzheimers Res. Ther. 2022, 14, 100. [Google Scholar] [CrossRef]
- Varghese, A.M.; Ghosh, M.; Bhagat, S.K.; Vijayalakshmi, K.; Preethish-Kumar, V.; Vengalil, S.; Chevula, P.C.; Nashi, S.; Polavarapu, K.; Sharma, M.; et al. Chitotriosidase, a biomarker of amyotrophic lateral sclerosis, accentuates neurodegeneration in spinal motor neurons through neuroinflammation. J. Neuroinflamm. 2020, 17, 232. [Google Scholar] [CrossRef]
- Piras, I.; Melis, A.; Ghiani, M.E.; Falchi, A.; Luiselli, D.; Moral, P.; Varesi, L.; Calò, C.M.; Vona, G. Human CHIT1 gene distribution: New data from Mediterranean and European populations. J. Hum. Genet. 2007, 52, 110. [Google Scholar] [CrossRef]
- Warwick, C.A.; Keyes, A.L.; Woodruff, T.M.; Usachev, Y.M. The complement cascade in the regulation of neuroinflammation, nociceptive sensitization, and pain. J. Biol. Chem. 2021, 297, 101085. [Google Scholar] [CrossRef]
- Dejanovic, B.; Huntley, M.A.; De Mazière, A.; Meilandt, W.J.; Wu, T.; Srinivasan, K.; Jiang, Z.; Gandham, V.; Friedman, B.A.; Ngu, H.; et al. Changes in the Synaptic Proteome in Tauopathy and Rescue of Tau-Induced Synapse Loss by C1q Antibodies. Neuron 2018, 100, 1322–1336.e7. [Google Scholar] [CrossRef]
- Nimmo, J.; Byrne, R.A.J.; Daskoulidou, N.; Watkins, L.M.; Carpanini, S.M.; Zelek, W.M.; Morgan, B.P. The complement system in neurodegenerative diseases. Clin. Sci. 2024, 138, 387–412. [Google Scholar] [CrossRef]
- Llorens, F.; Thüne, K.; Tahir, W.; Kanata, E.; Diaz-Lucena, D.; Xanthopoulos, K.; Kovatsi, E.; Pleschka, C.; Garcia-Esparcia, P.; Schmitz, M.; et al. YKL-40 in the brain and cerebrospinal fluid of neurodegenerative dementias. Mol. Neurodegener. 2017, 12, 83. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Benito, P.; Domínguez, R.; Colomina, M.J.; Llorens, F.; Povedano, M.; Ferrer, I. YKL40 in sporadic amyotrophic lateral sclerosis: Cerebrospinal fluid levels as a prognosis marker of disease progression. Aging 2018, 10, 2367–2382. [Google Scholar] [CrossRef]
- Benedet, A.L.; Milà-Alomà, M.; Vrillon, A.; Ashton, N.J.; Pascoal, T.A.; Lussier, F.; Karikari, T.K.; Hourregue, C.; Cognat, E.; Dumurgier, J.; et al. Differences Between Plasma and Cerebrospinal Fluid Glial Fibrillary Acidic Protein Levels Across the Alzheimer Disease Continuum. JAMA Neurol. 2021, 78, 1471–1483. [Google Scholar] [CrossRef]
- Chiam, J.T.; Dobson, R.J.; Kiddle, S.J.; Sattlecker, M. Are blood-based protein biomarkers for Alzheimer’s disease also involved in other brain disorders? A systematic review. J. Alzheimers Dis. 2015, 43, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.B.; Almeida, S.; Lopez-Gonzalez, R. Dysregulated molecular pathways in amyotrophic lateral sclerosis-frontotemporal dementia spectrum disorder. EMBO J. 2017, 36, 2931–2950. [Google Scholar] [CrossRef]
- Guyon, A. CXCL12 chemokine and its receptors as major players in the interactions between immune and nervous systems. Front. Cell. Neurosci. 2014, 8, 65. [Google Scholar] [CrossRef]
- Fröhlich, A.; Pfaff, A.L.; Bubb, V.J.; Quinn, J.P.; Koks, S. Transcriptomic profiling of cerebrospinal fluid identifies ALS pathway enrichment and RNA biomarkers in MND individuals. Exp. Biol. Med. 2023, 248, 2325–2331. [Google Scholar] [CrossRef]
- Thompson, A.G.; Gray, E.; Bampton, A.; Raciborska, D.; Talbot, K.; Turner, M.R. CSF chitinase proteins in amyotrophic lateral sclerosis. J. Neurol. Neurosurg. Psychiatry 2019, 90, 1215–1220. [Google Scholar] [CrossRef]
- Tan, Y.; Zheng, Y.; Xu, D.; Sun, Z.; Yang, H.; Yin, Q. Galectin-3: A key player in microglia-mediated neuroinflammation and Alzheimer’s disease. Cell Biosci. 2021, 11, 78. [Google Scholar] [CrossRef]
- Sparkman, N.L.; Johnson, R.W. Neuroinflammation associated with aging sensitizes the brain to the effects of infection or stress. Neuroimmunomodulation 2008, 15, 323–330. [Google Scholar] [CrossRef]
- Toft, A.; Roos, P.; Jääskeläinen, O.; Musaeus, C.S.; Henriksen, E.E.; Johannsen, P.; Nielsen, T.T.; Herukka, S.K.; Hviid Simonsen, A.; Nielsen, J.E. Serum Neurofilament Light in Patients with Frontotemporal Dementia Caused by CHMP2B Mutation. Dement. Geriatr. Cogn. Disord. 2020, 49, 533–538. [Google Scholar] [CrossRef]
- Khorooshi, R.; Marczynska, J.; Dieu, R.S.; Wais, V.; Hansen, C.R.; Kavan, S.; Thomassen, M.; Burton, M.; Kruse, T.; Webster, G.A.; et al. Innate signaling within the central nervous system recruits protective neutrophils. Acta Neuropathol. Commun. 2020, 8, 2. [Google Scholar] [CrossRef]
- Chitnis, T.; Weiner, H.L. CNS inflammation and neurodegeneration. J. Clin. Investig. 2017, 127, 3577–3587. [Google Scholar] [CrossRef]
- Gubbels Bupp, M.R.; Potluri, T.; Fink, A.L.; Klein, S.L. The Confluence of Sex Hormones and Aging on Immunity. Front. Immunol. 2018, 9, 1269. [Google Scholar] [CrossRef]
- Körtvélyessy, P.; Gukasjan, A.; Sweeney-Reed, C.M.; Heinze, H.J.; Thurner, L.; Bittner, D.M. Progranulin and Amyloid-β Levels: Relationship to Neuropsychology in Frontotemporal and Alzheimer’s Disease. J. Alzheimer’s Dis. 2015, 46, 375–380. [Google Scholar] [CrossRef]
- Longobardi, A.; Nicsanu, R.; Bellini, S.; Squitti, R.; Catania, M.; Tiraboschi, P.; Saraceno, C.; Ferrari, C.; Zanardini, R.; Binetti, G.; et al. Cerebrospinal Fluid EV Concentration and Size Are Altered in Alzheimer’s Disease and Dementia with Lewy Bodies. Cells 2022, 11, 462. [Google Scholar] [CrossRef]
- Comi, C.; Carecchio, M.; Chiocchetti, A.; Nicola, S.; Galimberti, D.; Fenoglio, C.; Cappellano, G.; Monaco, F.; Scarpini, E.; Dianzani, U. Osteopontin is increased in the cerebrospinal fluid of patients with Alzheimer’s disease and its levels correlate with cognitive decline. J. Alzheimer’s Dis. 2010, 19, 1143–1148. [Google Scholar] [CrossRef]
- Rentzos, M.; Paraskevas, G.P.; Kapaki, E.; Nikolaou, C.; Zoga, M.; Rombos, A.; Tsoutsou, A.; Vassilopoulos, D.D. Interleukin-12 is reduced in cerebrospinal fluid of patients with Alzheimer’s disease and frontotemporal dementia. J. Neurol. Sci. 2006, 249, 110–114. [Google Scholar] [CrossRef]
- Çoban, A.; Bilgiç, B.; Lohmann, E.; Küçükali, C.I.; Benbir, G.; Karadeniz, D.; Hanagasi, H.A.; Tüzün, E.; Gürvit, H. Reduced orexin-A levels in frontotemporal dementia: Possible association with sleep disturbance. Am. J. Alzheimer’s Dis. Other Dement. 2013, 28, 606–611. [Google Scholar] [CrossRef]
- Arshad, F.; Varghese, F.; Paplikar, A.; Gangadhar, Y.; Ramakrishnan, S.; Chaudhuri, J.R.; Mahadevan, A.; Alladi, S. Role of Autoantibodies in Neurodegenerative Dementia: An Emerging Association. Dement. Geriatr. Cogn. Disord. 2021, 50, 153–160. [Google Scholar] [CrossRef]
- Pawlowski, M.; Lueg, G.; Gross, C.C.; Johnen, A.; Krämer, J.; Weckesser, M.; Wiendl, H.; Meuth, S.G.; Duning, T. Relevance of raised cerebrospinal fluid monocyte levels in patients with frontotemporal dementia. Neurobiol. Aging 2018, 62, 45–52. [Google Scholar] [CrossRef]
- Jääskeläinen, O.; Solje, E.; Hall, A.; Katisko, K.; Korhonen, V.; Tiainen, M.; Kangas, A.J.; Helisalmi, S.; Pikkarainen, M.; Koivisto, A.; et al. Low Serum High-Density Lipoprotein Cholesterol Levels Associate with the C9orf72 Repeat Expansion in Frontotemporal Lobar Degeneration Patients. J. Alzheimer’s Dis. 2019, 72, 127–137. [Google Scholar] [CrossRef]
- Blauwendraat, C.; Wilke, C.; Simón-Sánchez, J.; Jansen, I.E.; Reifschneider, A.; Capell, A.; Haass, C.; Castillo-Lizardo, M.; Biskup, S.; Maetzler, W.; et al. The wide genetic landscape of clinical frontotemporal dementia: Systematic combined sequencing of 121 consecutive subjects. Genet. Med. 2018, 20, 240–249. [Google Scholar] [CrossRef]
| Reported Change in CSF Levels | FTD Versus Healthy controls CSF-Based Immune Markers |
|---|---|
| ↑↑ | C1q [8,17,47] C3b [8,17] CHIT1 [21,29,30,37] GFAP [18,21,26,27,28,29,48] MCP1/CCL2 [43,49,50,51,52] YKL-40 [21,29,30,31,32,33,34,35,36,37,53,54,55,56] |
| ↑ | β-NGF [44,57] C4 [58] C9 [59] CCL3 [43,49,51,52] Gal-3 [32] GPNMB [60] IL-8 [45,50,51,61] Leukotrienes [62] MMP-10 [44] NfL/YKL-40 ratio [35] SERPINA1 [63] SERPING1 [56] SPARC [56] TGF-β [52,64] TREM2 [37,65,66] Thromboxane [62] CD56+ [67] CCR7- CD45RO+ CD4+ [67] CCR7-CD45RA+ CD8+ [67] |
| ↓↓ | CCL19 [43,44] CXCL1 [43,44] CXCL6 [43,44] SST [44,45] |
| ↓ | ADA [44] Aβ42/YKL-40 ratio [34] CCL24 [44] CD5 [44] CD6 [44] CD8A [44] CD40 [44] CD244 [44] CDCP1 [44] CSF-1 [44] CXCL5 [43] CXCL11 [43,44] Cystatin C [56] DNER [44] FGF-5 [44] FGF-19 [44] Flt3L [44] HGF [44,52] IL-10RB [44] IL-12B [44] IL-15 [49,50,61] LIF [44] MCP-2 [44] PD-L1 [44] PGRN [68] RANTES [50] sAPPβ/YKL-40 ratio [35] SCF [44] TGF-α [44] TNFRSF9 [44] TNFRSF14 [44] TRAIL [44] TWEAK [44] uPA [44] VEGF [44,49,50] CD45RA- CD45RO+ CD4+ [67] CCR7+ CD45RO+ CD4+ [67] CCR7+ CD45RO+ CD8+ [67] CCR7- CD45RO+ CD8+ [67] |
| →→ | CCL4 [43,49,50] CCL11 [43,49] CXCL10 [43,49,51] CCL11 [43,50] FGF-2 [50,57,61] G-CSF [50,61] GM-CSF [50,61] IFN-γ [49,50,61] IL-1RA [50,61] IL-2 [50,61] IL-4 [50,61] IL-5 [49,50,61] IL-6 [50,61] IL-7 [49,50,61] IL-9 [50,61] IL-10 [49,50,61] IL-12 [49,50,61] IL-13 [50,61] IL-17 [50,61] Leukocytes [69,70] |
| → | C8G [71] CCL8 [43] CCL13 [49] CCL17 [49] CCL22 [49] CCL23 [43] CCL25 [43] CCL26 [49] CCL28 [43] CFB [47] CXCL9 [43] CXCL12 [72] DKK-3 [73] GFAP CSF/serum ratio [27] ICAM-1 [74] IL-1β [50] IL-16 [49] KYNA [75] MFG-E8 [32] PDGF-BB [50] Prostaglandins [62] VCAM [74] VEGF [50] CD14+ CD3- [67] CD14+ CD3+ [67] CD56+ CD3+ [67] HLA-DR CD14+ [67] CD3+ CD14- [67] CD4+ [67] CD8+ [67] CD19+ [67] HLA-DR CD3+ [67] CD69+ CD3+ [67] CD45RA- CD45RO- CD4+ [67] CCR7+ CD45RA+ CD8+ [67] CD45RA+ CD45RO- CD4+ [67] CD45RA+ CD45RO+ CD4+ [67] CD45RA- CD45RO- CD8+ [67] CD45RA+ CD45RO- CD8+ [67] CD45RA- CD45RO+ CD8+ [67] CD45RA+ CD45RO+ CD8+ [67] CCR7+ CD45RA+ CD4+ [67] CCR7- CD45RA+ CD4+ [67] |
| Contradictory | CX3CL1 [43,44] TNFα [49,50,52,64] |
| Reported change in blood levels | Blood-based immune markers |
| ↑↑ | C3 [8,76,77,78,79] CXCL10 [23,43,50,51,57] GFAP [16,17,18,19,20,21,22,23,24,25,27,80] IL-6 [49,57,76,77,81] IL-17A [23,57,82] IL-1β [23,38,50,57,77,82] MCP1/CCL2 [23,43,51,57] TNFα [23,50,57,77,81,82,83,84,85,86] |
| ↑ | ANA [87] APOL1 [78] APN [88] BAFF [82] C2 [8] C3 in EVs/plasma [79] CD163 [89] CCL4 [23,43,49,50] CCL8 [43] CCL11 [23,50,57] CCL13 [43,49] CCL18 [89] CCL26 [23] HERV-K [90] IL-2 [49,50,57,61,82] IL-2RA [57] IL-6 [23,50,82] IL-7 [23] IL-10 [23,49,50,57,77,82] IL-12p70 [23,49,57,77,82] IL-15 [23,49,50,57] IL-16 [23,49,57] MCP-4 [23] Myosin-2 [78] S100A8 [78] S100A12 [38] SCGF-BB [57] SLPI [42] TNFα [23] TNFRI [84] TNFRII [84] TNFRSF8/CD30 [82] TWEAK [82] YKL-40 [16,83,91] |
| ↓↓ | PGRN [38,39,40,41,42,50,92] |
| ↓ | C1s [78] C7 [49,50,57,78] CCL19 [43] CCL20 [43] GSN [78] IL-1α [23,49,57] IL-5 [23] IGHV1-2 [78] PROS-1 [78] |
| →→ | C1q [8,79] C4 [8,79] CCL11 [43,49] CCL23 [43,49] CRP [77,89,93] FGF-2 [50,57,61] G-CSF [50,57] GM-CSF [23,50,57] IFN-γ [23,49,50,57,77,82] IL-1RA [50,57] IL-4 [23,50,57,77,82] IL-5 [49,50,57,77] IL-8 [23,49,51,57,77] IL-9 [50,57] IL-12 [23,49,50] IL-13 [23,49,50,57,61] IL-17 [49,50,57] IL-18 [49,57,89] miRNA [94,95] VEGF [23,49,50] TNFβ [23,57] |
| → | Anti-β2GPIs [87] Anti-cardiolipin [87] Anti-TDP43 [96] BACE-1 [24] Basophils [93] BP180 NC16A [97] BP230 [97] C4B [8] C5 [8] Cathepsin S [98] CCL7 [43] CCL17 [49] CCL22 [49] CCL25 [43] CCL26 [49] CCL28 [43] CD14 [89] CFB [8] CFI [8] CFH [8,77] CTACK [57] CX3CL1 [43] CXCL1 [43] CXCL5 [43] CXCL6 [43] CXCL9 [43] CXCL11 [43] Gal-3 [32] GRO-α [57] HGF [57] IFN-α2 [57] IL-12p40 [57] IL-1 [86] IL-2 [23] IL-3 [57] IL-22 [77] Irisin [81] LBP [89] LIF [57] MDC [23] MIF [57] MIG [57] NOD2 [99] PDGF-BB [50,57] RANTES [50,57] SCF [57] SDF-1α [57] SPP1 [99] TARC [23] TRAIL [57] TREM2 [86] TWEAK [84] |
| Contradictory | CCL3 [23,43,49,50,57] |
| Age | Sex | NFL | Cortical Atrophy | MMSE | |
|---|---|---|---|---|---|
| Positive Correlations | |||||
| CSF | CXCL9 * [43] IL-15 * [49,50,101] MCP1/CCL2 ** [50,61,114] TREM2 * [49] TGF-β * [64] YKL-40 * [115] | C1q (presymp- tomatic) *** [8] C3b (presymp- tomatic) *** [8] CHIT1 *** [29] YKL-40 *** [29] | C1q (presymp- tomatic) ** [8] C3b (presymp- tomatic) ** [8] | ||
| Blood | CCL13 * [49] GFAP *** [18,25] IL-1β * [38] IL-6 *** [41,51,83] IL-8 * [49] SLPI *** [42] TNFα *** [83] YKL-40 *** [83,91] | GFAP (females) *** [25] | Complement Factor D (presymptomatic) *** [8] GFAP *** [16,22] | C5 * [8] Complement Factor D *** [8] GFAP * [22] IL-17A * [82] LBP * [89] TNFα * [82,83] TNFRII ** [84] | |
| Negative correlations | |||||
| CSF | CCL28 * (females) [43] | CHIT1 (presymptomatic GRN-FTD) ** [37] | |||
| Blood | GFAP ** [16,25] MCP1/CCL2 * [116] | ||||
| No correlation | |||||
| CSF | GFAP [18] IL-11 [117] IL-15 [101] CXCL10 [50] PGRN [118] RANTES [50] S100B [119] SORL1 [120] SST [45] TNF [50] | CCL2 [43], CCL3 [43] CCL4 [43] CCL8 [43] CCL11 [43], CCL19 [43] CC23 [43], CCL25 [43] CCL28 [43], CX3CL1 [43] CXCL1 [43], CXCL5 [43] CXCL6 [43], CXCL9 [43] CXCL10 [43], CXCL11 [43], GFAP [38] IgM and IgG gangliosides [121] IL-6 [117], IL-8 [43] IL-11 [117], IL-15 [101] SST [45] | TNFα [64] TGF-β [64] IL-1β [64] S100B [119] Monocytes [122] | CHIT1 [37] GFAP [28] IL-15 [101] IgM and IgG gangliosides [121] S100B [119] TREM2 [37,65] YKL-40 [37] | |
| Blood | Cathepsin S [98] PGRN [38,41,42,118,123,124] | Cathepsin S [98] GFAP [18] | C9 [59] Gal-3 [32] | BAFF [82], IFN-y [82] IL-1b [82], IL-2 [82] IL-4 [82], IL-6 [82,83] (GRN-FTD) IL-10 [82] IL-12p70 [82] sCD30 [82] TWEAK [82] YKL-40 [82] | IL-6 [125] IL-18 [125] TNF [84] TNFRI [84] TNFRII [84] TWEAK [84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erichsen, P.A.; Henriksen, E.E.; Nielsen, J.E.; Ejlerskov, P.; Simonsen, A.H.; Toft, A. Immunological Fluid Biomarkers in Frontotemporal Dementia: A Systematic Review. Biomolecules 2025, 15, 473. https://doi.org/10.3390/biom15040473
Erichsen PA, Henriksen EE, Nielsen JE, Ejlerskov P, Simonsen AH, Toft A. Immunological Fluid Biomarkers in Frontotemporal Dementia: A Systematic Review. Biomolecules. 2025; 15(4):473. https://doi.org/10.3390/biom15040473
Chicago/Turabian StyleErichsen, Philip Ahle, Emil Elbæk Henriksen, Jørgen Erik Nielsen, Patrick Ejlerskov, Anja Hviid Simonsen, and Anders Toft. 2025. "Immunological Fluid Biomarkers in Frontotemporal Dementia: A Systematic Review" Biomolecules 15, no. 4: 473. https://doi.org/10.3390/biom15040473
APA StyleErichsen, P. A., Henriksen, E. E., Nielsen, J. E., Ejlerskov, P., Simonsen, A. H., & Toft, A. (2025). Immunological Fluid Biomarkers in Frontotemporal Dementia: A Systematic Review. Biomolecules, 15(4), 473. https://doi.org/10.3390/biom15040473