

Region-Specific Impact of Repeated Synthetic Cannabinoid Exposure and Withdrawal on Endocannabinoid Signaling, Gliosis, and Inflammatory Markers in the Prefrontal Cortex and Hippocampus

 ,
,  ,
,  , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
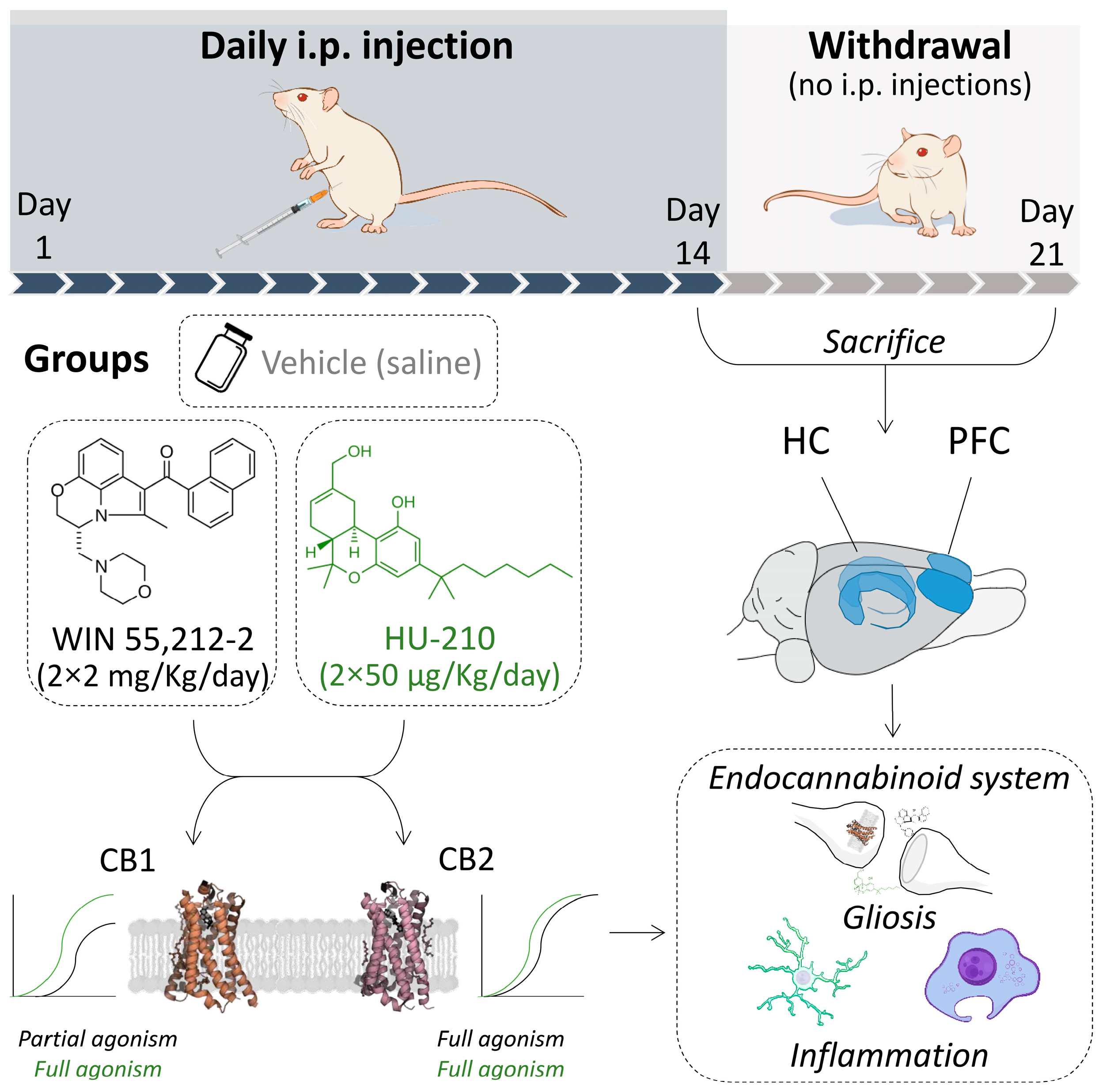
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals
2.3. Drug Administration
2.4. Experimental Design
2.5. Tissue Collection
2.6. RNA Isolation and RT-qPCR Analysis
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
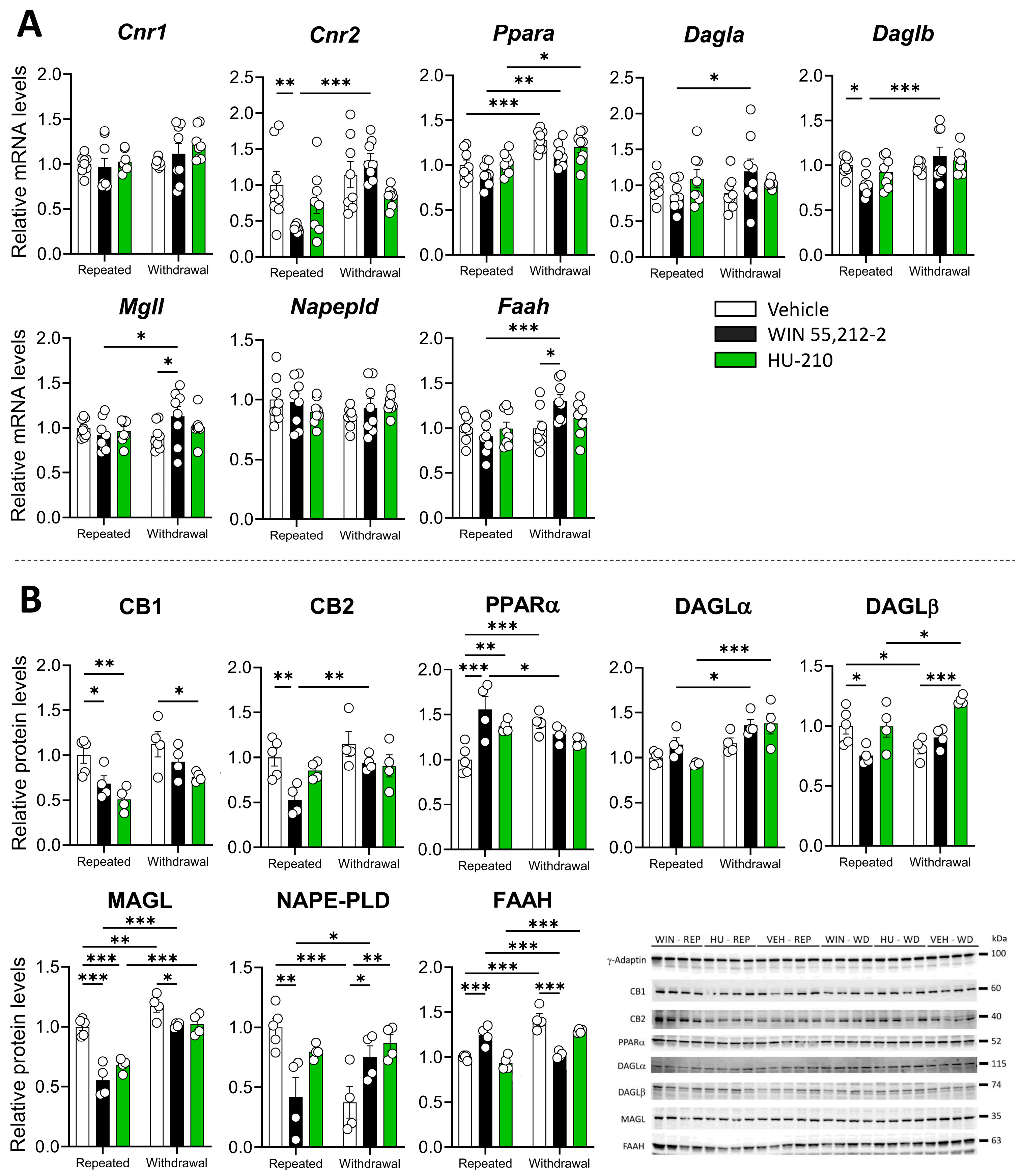
3.1. Repeated Administration and Withdrawal of WIN 55,212-2 and HU-210 Lead to Distinct Effects on the Endocannabinoid System in the Prefrontal Cortex
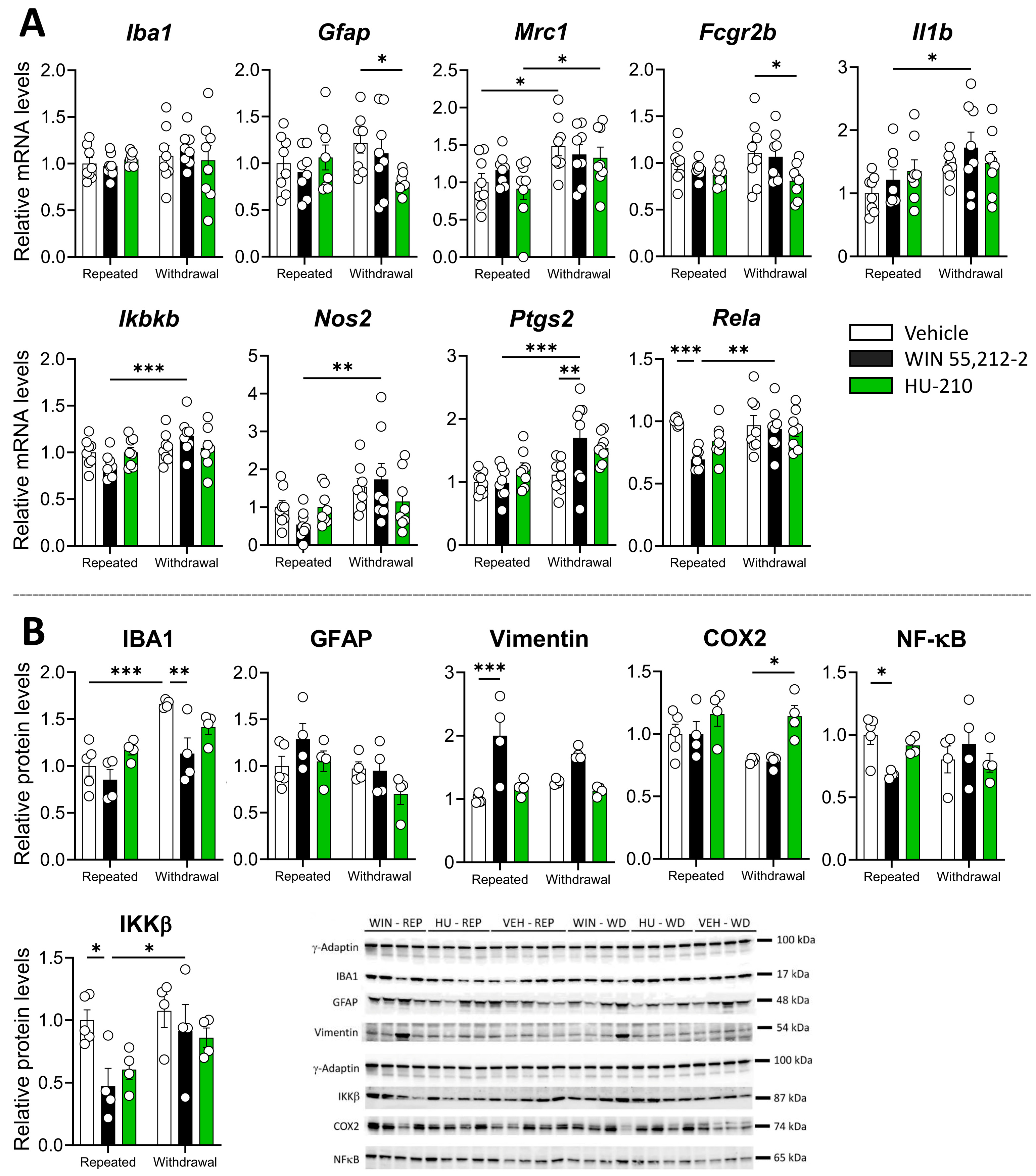
3.2. Repeated Administration and Withdrawal of WIN 55,212-2 and HU-210 Modulate Gliosis and Inflammatory Markers in the Prefrontal Cortex
3.3. Repeated Administration and Withdrawal of WIN 55,212-2 and HU-210 Differentially Modify the Endocannabinoid System in Hippocampus
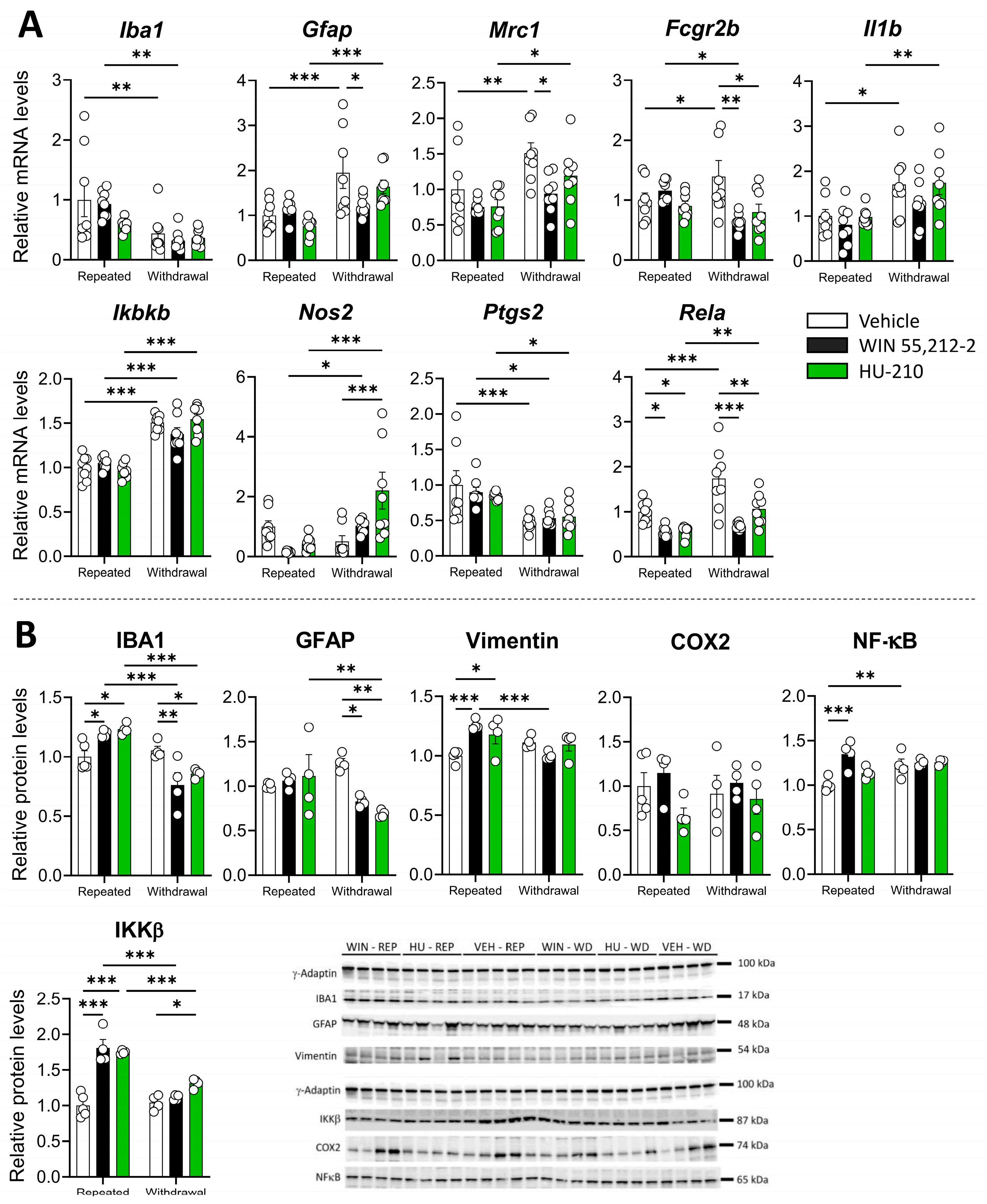
3.4. Repeated Administration of WIN 55,212-2 and HU-210 Increases Markers of Gliosis in the Hippocampus That Are Resolved During Withdrawal
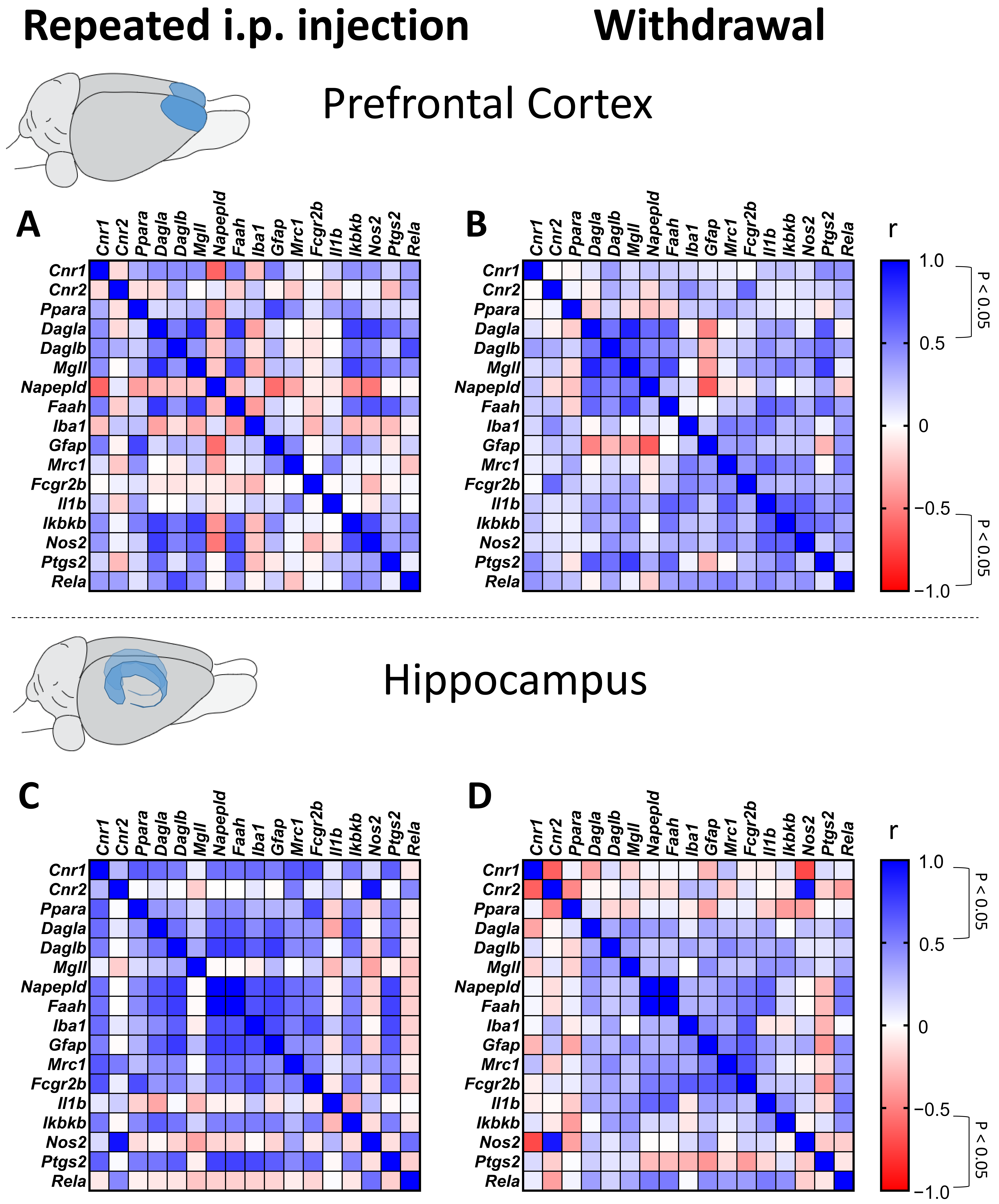
3.5. Relationship Between Gliosis, Inflammation Markers, and Endocannabinoid System During Repeated Administration and Withdrawal of WIN 55,212-2 and HU-210
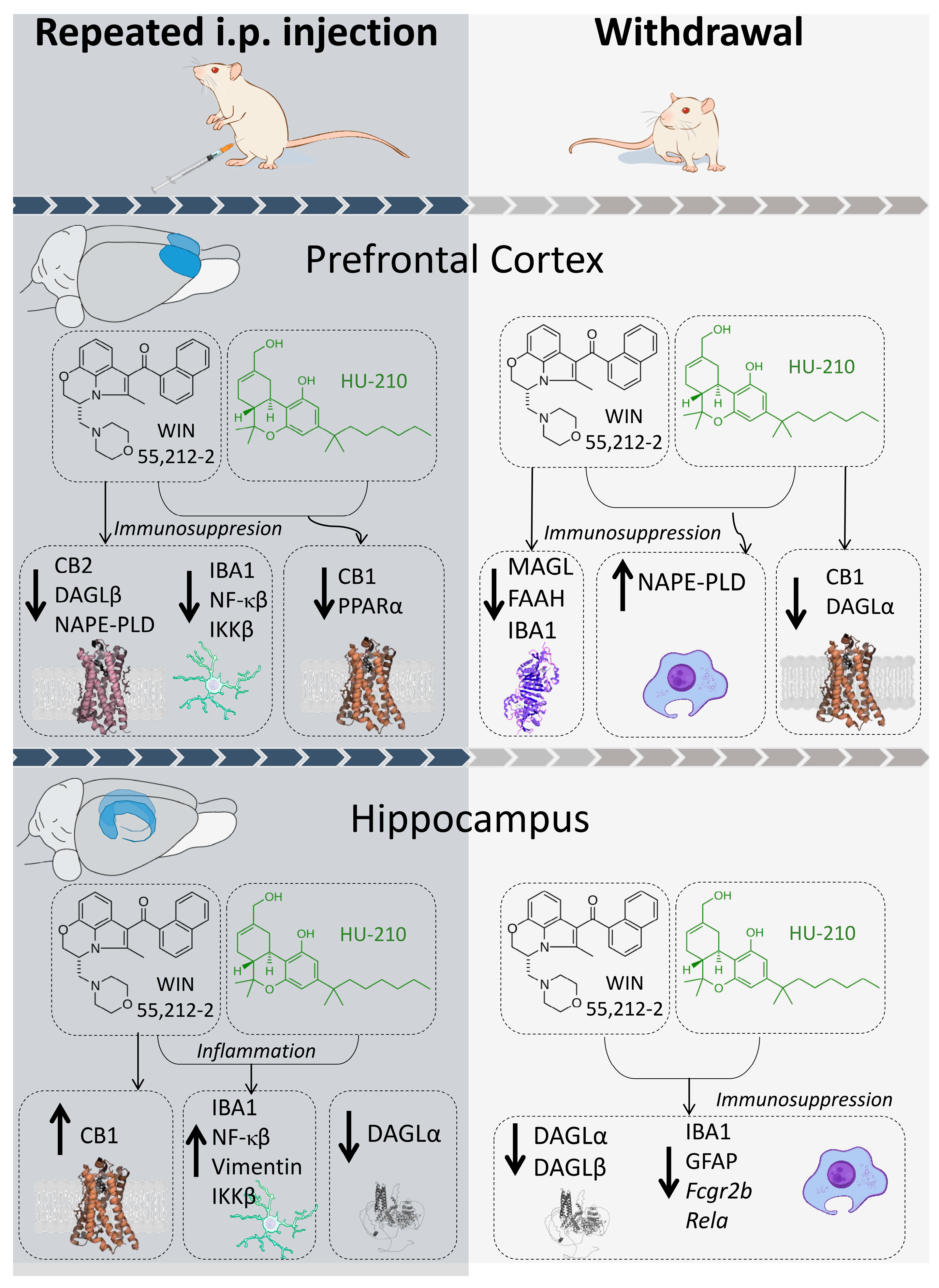
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coelho, M.P.; Duarte, P.; Calado, M.; Almeida, A.J.; Reis, C.P.; Gaspar, M.M. The current role of cannabis and cannabinoids in health: A comprehensive review of their therapeutic potential. Life Sci. 2023, 329, 121838. [Google Scholar] [CrossRef]
- Marshell, R.; Kearney-Ramos, T.; Brents, L.; Hyatt, W.; Tai, S.; Prather, P.; Fantegrossi, W. In vivo effects of synthetic cannabinoids JWH-018 and JWH-073 and phytocannabinoid Δ9-THC in mice: Inhalation versus intraperitoneal injection. Pharmacol. Biochem. Behav. 2014, 124, 40–47. [Google Scholar] [CrossRef] [PubMed]
- El-aal, A.; Hemeda, M.S.; Sayed, H.Y. New designer drugs: An emergent community challenge. Med. Updates 2023, 15, 69–76. [Google Scholar] [CrossRef]
- Celeste-Villalvir, A.; Crouch, C.; Heads, A.M.; Witte, L.; Weaver, M.; Schmitz, J.M.; Isbell, F.; Schick, V. Investigating the Experiences and Effects of Synthetic Cannabinoid (kush/K2/Spice) Use Among Individuals Experiencing Homelessness in Houston, Texas. Tex. Public Health J. 2022, 74, 23. [Google Scholar]
- Castaneto, M.S.; Gorelick, D.A.; Desrosiers, N.A.; Hartman, R.L.; Pirard, S.; Huestis, M.A. Synthetic cannabinoids: Epidemiology, pharmacodynamics, and clinical implications. Drug Alcohol Depend. 2014, 144, 12–41. [Google Scholar] [CrossRef]
- Adams, A.J.; Banister, S.D.; Irizarry, L.; Trecki, J.; Schwartz, M.; Gerona, R. “Zombie” outbreak caused by the synthetic cannabinoid AMB-FUBINACA in New York. N. Engl. J. Med. 2017, 376, 235–242. [Google Scholar] [CrossRef]
- European Monitoring Centre for Drugs and Drug Addiction. Synthetic Cannabinoids in Europe—A Review; Publications Office of the European Union: Luxembourg, 2021. [Google Scholar]
- Shevyrin, V.; Melkozerov, V.; Endres, G.W.; Shafran, Y.; Morzherin, Y. On a new cannabinoid classification system: A sight on the illegal market of novel psychoactive substances. Cannabis Cannabinoid Res. 2016, 1, 186–194. [Google Scholar] [CrossRef]
- Howlett, A.C.; Abood, M.E. CB1 and CB2 receptor pharmacology. Adv. Pharmacol. 2017, 80, 169–206. [Google Scholar]
- 1Busquets-Garcia, A.; Bains, J.; Marsicano, G. CB1 receptor signaling in the brain: Extracting specificity from ubiquity. Neuropsychopharmacology 2018, 43, 4–20. [Google Scholar] [CrossRef]
- Li, Y.; Kim, J. Deletion of CB2 cannabinoid receptors reduces synaptic transmission and long-term potentiation in the mouse hippocampus. Hippocampus 2016, 26, 275–281. [Google Scholar] [CrossRef]
- Albayram, O.; Alferink, J.; Pitsch, J.; Piyanova, A.; Neitzert, K.; Poppensieker, K.; Mauer, D.; Michel, K.; Legler, A.; Becker, A. Role of CB1 cannabinoid receptors on GABAergic neurons in brain aging. Proc. Natl. Acad. Sci. USA 2011, 108, 11256–11261. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Stella, N.; Zimmer, A. Endocannabinoid signalling and the deteriorating brain. Nat. Rev. Neurosci. 2015, 16, 30–42. [Google Scholar] [CrossRef]
- Tauskela, J.S.; Comas, T.; Hewitt, M.; Aylsworth, A.; Zhao, X.; Martina, M.; Costain, W.J. Effect of synthetic cannabinoids on spontaneous neuronal activity: Evaluation using Ca2+ spiking and multi-electrode arrays. Eur. J. Pharmacol. 2016, 786, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Perdikaris, P.; Tsarouchi, M.; Fanarioti, E.; Natsaridis, E.; Mitsacos, A.; Giompres, P. Long lasting effects of chronic WIN55, 212-2 treatment on mesostriatal dopaminergic and cannabinoid systems in the rat brain. Neuropharmacology 2018, 129, 1–15. [Google Scholar] [CrossRef]
- Andó, R.D.; Bíró, J.; Csölle, C.; Ledent, C.; Sperlágh, B. The inhibitory action of exo-and endocannabinoids on [3H] GABA release are mediated by both CB1 and CB2 receptors in the mouse hippocampus. Neurochem. Int. 2012, 60, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Decara, J.; Pavon, F.J.; Stouffer, D.G.; Edwards, S.; Serrano, A.; Suárez, J.; Parsons, L.H.; Rodríguez de Fonseca, F. Cannabinoid dependence induces sustained changes in GABA release in the globus pallidus without affecting dopamine release in the dorsal striatum: A dual microdialysis probe study. Addict. Biol. 2018, 23, 1251–1261. [Google Scholar] [CrossRef]
- Ferrari, F.; Ottani, A.; Giuliani, D. Inhibitory effects of the cannabinoid agonist HU 210 on rat sexual behaviour. Physiol. Behav. 2000, 69, 547–554. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Decara, J.; Rivera, P.; Arrabal, S.; Vargas, A.; Serrano, A.; Pavón, F.; Dieguez, C.; Nogueiras, R.; Rodríguez de Fonseca, F.; Suárez, J. Cooperative role of the glucagon-like peptide-1 receptor and β3-adrenergic-mediated signalling on fat mass reduction through the downregulation of PKA/AKT/AMPK signalling in the adipose tissue and muscle of rats. Acta Physiol. 2018, 222, e13008. [Google Scholar] [CrossRef]
- De Fonseca, F.R.; Gorriti, M.; Fernandez-Ruiz, J.; Palomo, T.; Ramos, J. Downregulation of rat brain cannabinoid binding sites after chronic Δ9-tetrahydrocannabinol treatment. Pharmacol. Biochem. Behav. 1994, 47, 33–40. [Google Scholar] [CrossRef]
- Lisman, J.; Buzsáki, G.; Eichenbaum, H.; Nadel, L.; Ranganath, C.; Redish, A.D. Viewpoints: How the hippocampus contributes to memory, navigation and cognition. Nat. Neurosci. 2017, 20, 1434–1447. [Google Scholar] [CrossRef]
- Scarante, F.F.; Vila-Verde, C.; Detoni, V.L.; Ferreira-Junior, N.C.; Guimarães, F.S.; Campos, A.C. Cannabinoid modulation of the stressed hippocampus. Front. Mol. Neurosci. 2017, 10, 411. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.W. Cannabinoid-based drugs as anti-inflammatory therapeutics. Nat. Rev. Immunol. 2005, 5, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Suárez, J.; Llorente, R.; Romero-Zerbo, S.Y.; Mateos, B.; Bermúdez-Silva, F.J.; de Fonseca, F.R.; Viveros, M.P. Early maternal deprivation induces gender-dependent changes on the expression of hippocampal CB1 and CB2 cannabinoid receptors of neonatal rats. Hippocampus 2009, 19, 623–632. [Google Scholar] [CrossRef]
- Wu, D.F.; Yang, L.Q.; Goschke, A.; Stumm, R.; Brandenburg, L.O.; Liang, Y.J.; Höllt, V.; Koch, T. Role of receptor internalization in the agonist-induced desensitization of cannabinoid type 1 receptors. J. Neurochem. 2008, 104, 1132–1143. [Google Scholar] [CrossRef]
- Sim-Selley, L.J.; Martin, B.R. Effect of Chronic Administration ofR-(+)-[2,3-Dihydro-5-methyl-3-[(morpholinyl) methyl] pyrrolo [1,2,3-de]-1,4-benzoxazinyl]-(1-naphthalenyl) methanone Mesylate (WIN55, 212-2) or Δ9-Tetrahydrocannabinol on Cannabinoid Receptor Adaptation in Mice. J. Pharmacol. Exp. Ther. 2002, 303, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Roig, J.; Agbemenyah, H.; Celarain, N.; Quintana, E.; Ramió-Torrentà, L.; Havemann-Reinecke, U. Dose-dependent effect of cannabinoid WIN-55,212-2 on myelin repair following a demyelinating insult. Sci. Rep. 2020, 10, 590. [Google Scholar] [CrossRef]
- Loría, F.; Petrosino, S.; Mestre, L.; Spagnolo, A.; Correa, F.; Hernangómez, M.; Guaza, C.; Di Marzo, V.; Docagne, F. Study of the regulation of the endocannabinoid system in a virus model of multiple sclerosis reveals a therapeutic effect of palmitoylethanolamide. Eur. J. Neurosci. 2008, 28, 633–641. [Google Scholar] [CrossRef]
- Chung, Y.C.; Shin, W.-H.; Baek, J.Y.; Cho, E.J.; Baik, H.H.; Kim, S.R.; Won, S.-Y.; Jin, B.K. CB2 receptor activation prevents glial-derived neurotoxic mediator production, BBB leakage and peripheral immune cell infiltration and rescues dopamine neurons in the MPTP model of Parkinson’s disease. Exp. Mol. Med. 2016, 48, e205. [Google Scholar] [CrossRef]
- Shan, R.; Zhang, Y.; Shi, Y.; Wang, X.; Wang, X.; Ma, G.; Li, Q. Activation of Cannabinoid Type 2 Receptor in Microglia Reduces Neuroinflammation through Inhibiting Aerobic Glycolysis to Relieve Hypertension. Biomolecules 2024, 14, 333. [Google Scholar] [CrossRef]
- Olabiyi, B.F.; Schmoele, A.-C.; Beins, E.C.; Zimmer, A. Pharmacological blockade of cannabinoid receptor 2 signaling does not affect LPS/IFN-γ-induced microglial activation. Sci. Rep. 2023, 13, 11105. [Google Scholar]
- Bie, B.; Wu, J.; Foss, J.F.; Naguib, M. An overview of the cannabinoid type 2 receptor system and its therapeutic potential. Curr. Opin. Anesthesiol. 2018, 31, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, A.; Caissutti, D.; Mattei, V.; Gado, F.; Martellucci, S.; Longo, A.; Recalchi, S.; Manganelli, V.; Riitano, G.; Garofalo, T. Anti-inflammatory activity of a CB2 selective cannabinoid receptor agonist: Signaling and cytokines release in blood mononuclear cells. Molecules 2021, 27, 64. [Google Scholar] [CrossRef] [PubMed]
- Marchalant, Y.; Rosi, S.; Wenk, G.L. Anti-inflammatory property of the cannabinoid agonist WIN-55212-2 in a rodent model of chronic brain inflammation. Neuroscience 2007, 144, 1516–1522. [Google Scholar] [CrossRef]
- Fields, J.A.; Swinton, M.K.; Montilla-Perez, P.; Ricciardelli, E.; Telese, F. The cannabinoid receptor agonist, WIN-55212-2, suppresses the activation of proinflammatory genes induced by interleukin 1 beta in human astrocytes. Cannabis Cannabinoid Res. 2022, 7, 78–92. [Google Scholar] [CrossRef]
- Puffenbarger, R.A.; Boothe, A.C.; Cabral, G.A. Cannabinoids inhibit LPS-inducible cytokine mRNA expression in rat microglial cells. Glia 2000, 29, 58–69. [Google Scholar] [CrossRef]
- Wang, X.; Lin, C.; Jin, S.; Wang, Y.; Peng, Y.; Wang, X. Cannabidiol alleviates neuroinflammation and attenuates neuropathic pain via targeting FKBP5. Brain Behav. Immun. 2023, 111, 365–375. [Google Scholar] [CrossRef] [PubMed]
- McKenna, M.; McDougall, J.J. Cannabinoid control of neurogenic inflammation. Br. J. Pharmacol. 2020, 177, 4386–4399. [Google Scholar] [CrossRef]
- Lins, B.R.; Anyaegbu, C.C.; Hellewell, S.C.; Papini, M.; McGonigle, T.; De Prato, L.; Shales, M.; Fitzgerald, M. Cannabinoids in traumatic brain injury and related neuropathologies: Preclinical and clinical research on endogenous, plant-derived, and synthetic compounds. J. Neuroinflamm. 2023, 20, 77. [Google Scholar] [CrossRef]
- Robinson, T.E.; Kolb, B. Structural plasticity associated with exposure to drugs of abuse. Neuropharmacology 2004, 47, 33–46. [Google Scholar] [CrossRef]
- Filbey, F.M.; Dunlop, J.; Myers, U.S. Neural effects of positive and negative incentives during marijuana withdrawal. PLoS ONE 2013, 8, e61470. [Google Scholar] [CrossRef]
- Taylor, A.; Nweke, A.; Vincent, V.; Oke, M.; Kulkarni, P.; Ferris, C.F. Chronic exposure to inhaled vaporized cannabis high in Δ9-THC alters brain structure in adult female mice. Front. Neurosci. 2023, 17, 1139309. [Google Scholar] [CrossRef] [PubMed]
- Rivera, P.; Bindila, L.; Pastor, A.; Pérez-Martín, M.; Pavón, F.J.; Serrano, A.; de la Torre, R.; Lutz, B.; Rodríguez de Fonseca, F.; Suárez, J. Pharmacological blockade of the fatty acid amide hydrolase (FAAH) alters neural proliferation, apoptosis and gliosis in the rat hippocampus, hypothalamus and striatum in a negative energy context. Front. Cell. Neurosci. 2015, 9, 98. [Google Scholar] [CrossRef]
- Murphy, N.; Cowley, T.R.; Blau, C.W.; Dempsey, C.N.; Noonan, J.; Gowran, A.; Tanveer, R.; Olango, W.M.; Finn, D.P.; Campbell, V.A. The fatty acid amide hydrolase inhibitor URB597 exerts anti-inflammatory effects in hippocampus of aged rats and restores an age-related deficit in long-term potentiation. J. Neuroinflamm. 2012, 9, 79. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, M.A.; Yadav, S.; Gupta, R.K.; Waggoner, G.R.; Deloach, A.; Calingasan, N.Y.; Beal, M.F.; Kiaei, M. Preferential PPAR-α activation reduces neuroinflammation, and blocks neurodegeneration in vivo. Hum. Mol. Genet. 2016, 25, 317–327. [Google Scholar] [CrossRef]
- O’Sullivan, S.E. An update on PPAR activation by cannabinoids. Br. J. Pharmacol. 2016, 173, 1899–1910. [Google Scholar] [CrossRef] [PubMed]
- Antón, M.; Alén, F.; Gomez de Heras, R.; Serrano, A.; Pavón, F.J.; Leza, J.C.; García-Bueno, B.; Rodriguez de Fonseca, F.; Orio, L. Oleoylethanolamide prevents neuroimmune HMGB1/TLR4/NF-kB danger signaling in rat frontal cortex and depressive-like behavior induced by ethanol binge administration. Addict. Biol. 2017, 22, 724–741. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Falkenberg, I.; Martin-Santos, R.; Atakan, Z.; Crippa, J.A.; Giampietro, V.; Brammer, M.; McGuire, P. Cannabinoid modulation of functional connectivity within regions processing attentional salience. Neuropsychopharmacology 2015, 40, 1343–1352. [Google Scholar] [CrossRef]
- Serrano, A.; Vadas, E.; Ferrer, B.; Bilbao, A.; Granado, N.; Suárez, J.; Pavon, F.J.; Moratalla, R.; Rodríguez de Fonseca, F. Genetic deletion of dopamine D1 receptors increases the sensitivity to cannabinoid CB1 receptor antagonist-precipitated withdrawal when compared with wild-type littermates: Studies in female mice repeatedly exposed to the Spice cannabinoid HU-210. Psychopharmacology 2021, 238, 551–557. [Google Scholar] [CrossRef]
- Rodríguez de Fonseca, F.; Cebeira, M.; Ramos, J.A.; Martín, M.; Fernández-Ruiz, J.J. Cannabinoid receptors in rat brain areas: Sexual differences, fluctuations during estrous cycle and changes after gonadectomy and sex steroid replacement. Life Sci. 1994, 54, 159–170. [Google Scholar] [CrossRef]
- Farquhar, C.E.; Breivogel, C.S.; Gamage, T.F.; Gay, E.A.; Thomas, B.F.; Craft, R.M.; Wiley, J.L. Sex, THC, and hormones: Effects on density and sensitivity of CB1cannabinoid receptors in rats. Drug Alcohol Depend. 2019, 194, 20–27. [Google Scholar] [CrossRef]
- Laurikainen, H.; Tuominen, L.; Tikka, M.; Merisaari, H.; Armio, R.L.; Sormunen, E.; Borgan, F.; Veronese, M.; Howes, O.; Haaparanta-Solin, M.; et al. Sex difference in brain CB1 receptor availability in man. Neuroimage 2019, 184, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.D.; Rubin, T.G.; Hunter, R.G.; McEwen, B.S. Hippocampal gene expression changes underlying stress sensitization and recovery. Mol. Psychiatry 2014, 19, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Cai, D.; Yang, X.; Shang, Y.; Li, X.; Jia, Y.; Yin, C.; Zou, H.; Xu, Y.; Sun, Q. Stress response simulated by continuous injection of ACTH attenuates lipopolysaccharide-induced inflammation in porcine adrenal gland. Front. Vet. Sci. 2020, 7, 315. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vadas, E.; López-Gambero, A.J.; Vargas, A.; Rodríguez-Pozo, M.; Rivera, P.; Decara, J.; Serrano, A.; Martín-de-las-Heras, S.; Rodríguez de Fonseca, F.; Suárez, J. Region-Specific Impact of Repeated Synthetic Cannabinoid Exposure and Withdrawal on Endocannabinoid Signaling, Gliosis, and Inflammatory Markers in the Prefrontal Cortex and Hippocampus. Biomolecules 2025, 15, 417. https://doi.org/10.3390/biom15030417
Vadas E, López-Gambero AJ, Vargas A, Rodríguez-Pozo M, Rivera P, Decara J, Serrano A, Martín-de-las-Heras S, Rodríguez de Fonseca F, Suárez J. Region-Specific Impact of Repeated Synthetic Cannabinoid Exposure and Withdrawal on Endocannabinoid Signaling, Gliosis, and Inflammatory Markers in the Prefrontal Cortex and Hippocampus. Biomolecules. 2025; 15(3):417. https://doi.org/10.3390/biom15030417
Chicago/Turabian StyleVadas, Evelin, Antonio J. López-Gambero, Antonio Vargas, Miguel Rodríguez-Pozo, Patricia Rivera, Juan Decara, Antonia Serrano, Stella Martín-de-las-Heras, Fernando Rodríguez de Fonseca, and Juan Suárez. 2025. "Region-Specific Impact of Repeated Synthetic Cannabinoid Exposure and Withdrawal on Endocannabinoid Signaling, Gliosis, and Inflammatory Markers in the Prefrontal Cortex and Hippocampus" Biomolecules 15, no. 3: 417. https://doi.org/10.3390/biom15030417
APA StyleVadas, E., López-Gambero, A. J., Vargas, A., Rodríguez-Pozo, M., Rivera, P., Decara, J., Serrano, A., Martín-de-las-Heras, S., Rodríguez de Fonseca, F., & Suárez, J. (2025). Region-Specific Impact of Repeated Synthetic Cannabinoid Exposure and Withdrawal on Endocannabinoid Signaling, Gliosis, and Inflammatory Markers in the Prefrontal Cortex and Hippocampus. Biomolecules, 15(3), 417. https://doi.org/10.3390/biom15030417