
Multifaceted Therapeutic Potential of Plant-Derived Exosomes: Immunomodulation, Anticancer, Anti-Aging, Anti-Melanogenesis, Detoxification, and Drug Delivery


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
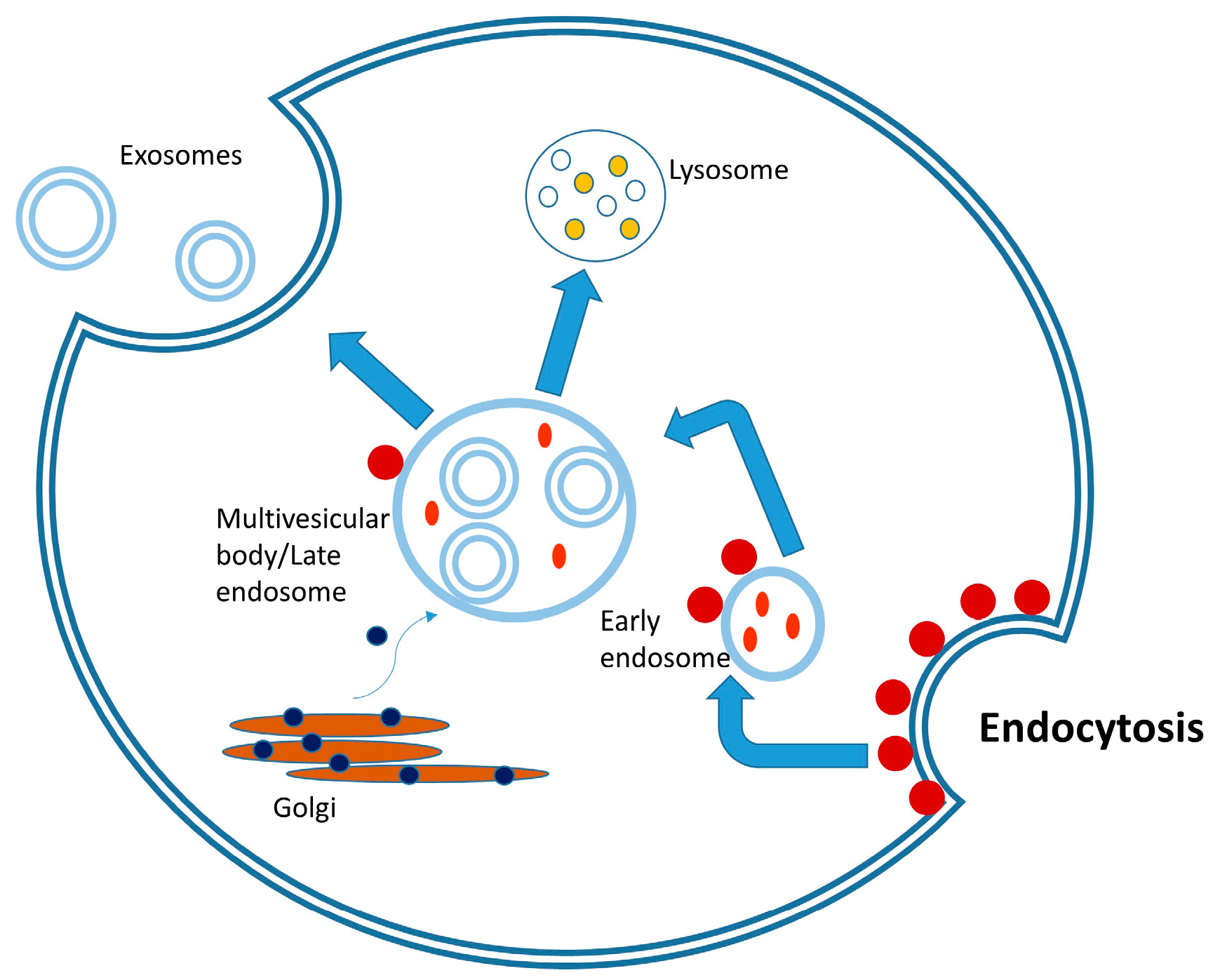
2. Biogenesis of Exosomes
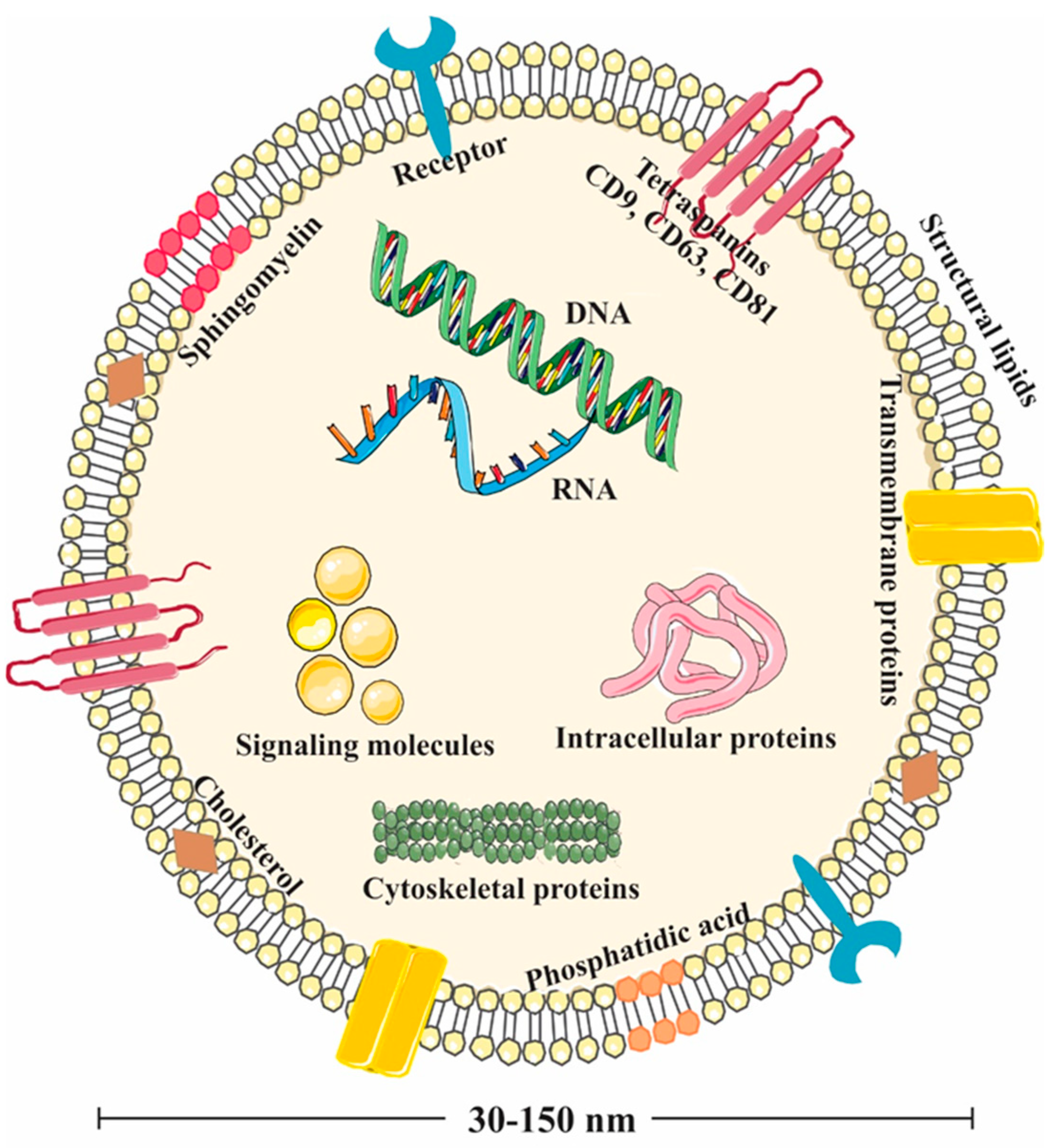
3. EVs Derived from Natural Green Nano-Factories
3.1. Lipids in PDEVs
3.2. Proteins in PDEVs
3.3. Metabolites in PDEVs
3.4. miRNAs in PDEVs
Stability of Plant-Derived miRNAs
3.5. Therapeutic Approaches of PDEVs
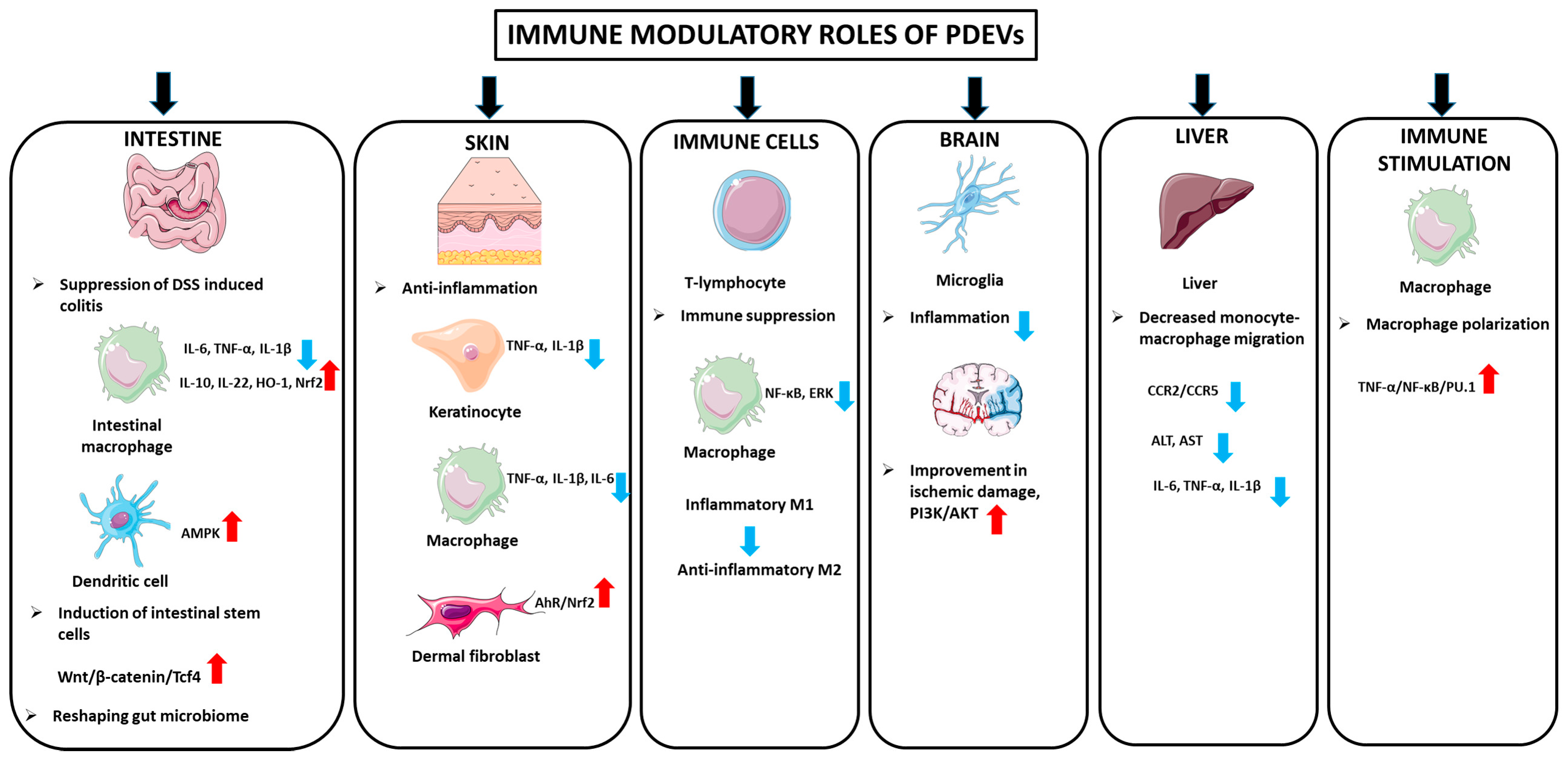
3.5.1. Immunomodulatory Properties
Intestinal Anti-Inflammatory Activity
Anti-Inflammatory Activity in the Skin
Anti-Inflammatory Activity in Immune Cells
Anti-Inflammatory Activity in Neurodegenerative Disorders and Ischemic Brain Diseases
Anti-Inflammatory Activity in Liver
Immune Stimulant Activity
3.5.2. Anticancer Properties
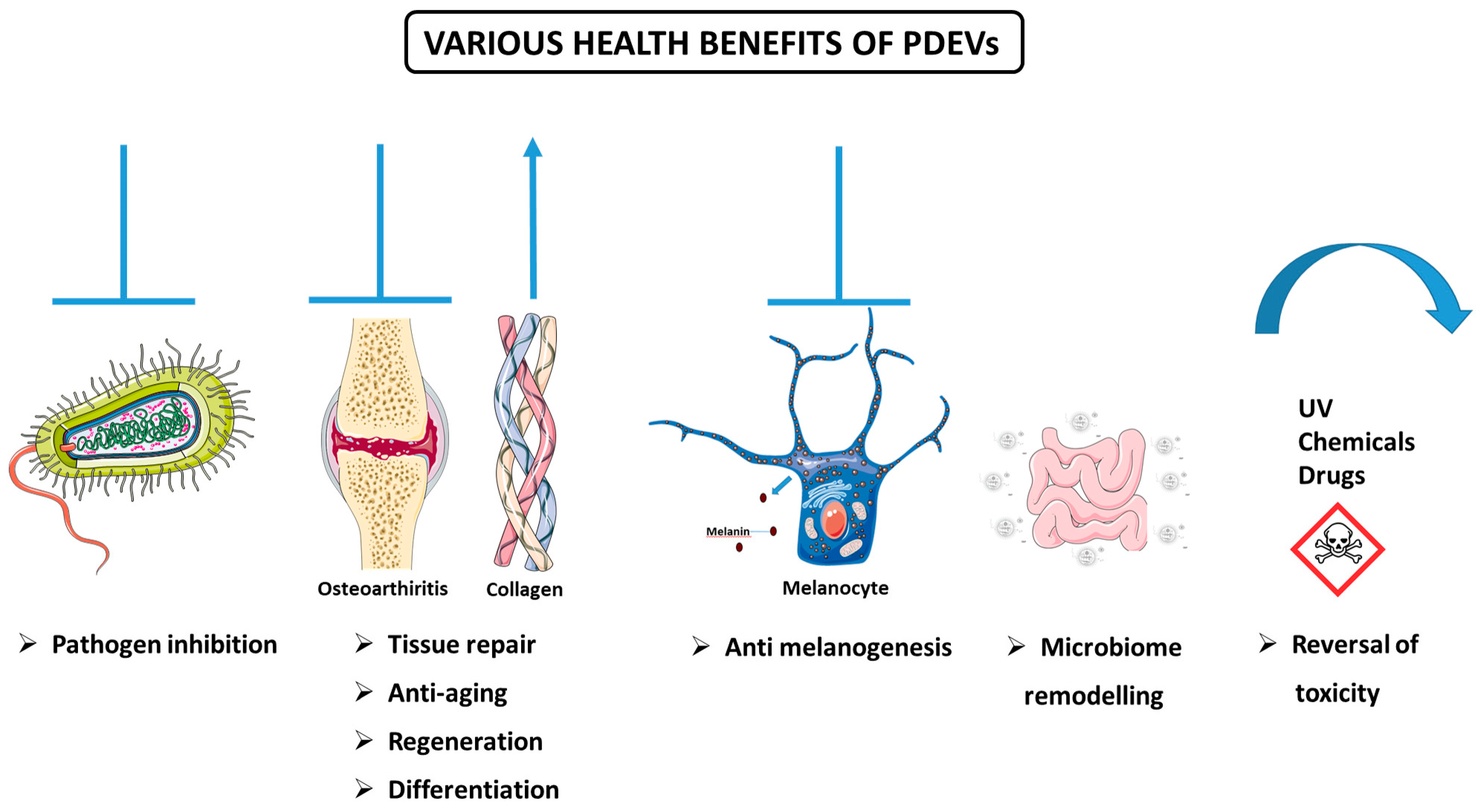
3.5.3. Various Health-Promoting Effects of PDEVs
Defense Against Pathogenic Species
Anti-Arthritic, Regenerative, Proliferative, and Differentiation Activities
Anti-Melanogenesis Activity
Anti-Aging Activity
Remodeling of the Microbiome
Reversal of Toxicity
Other Health Effects
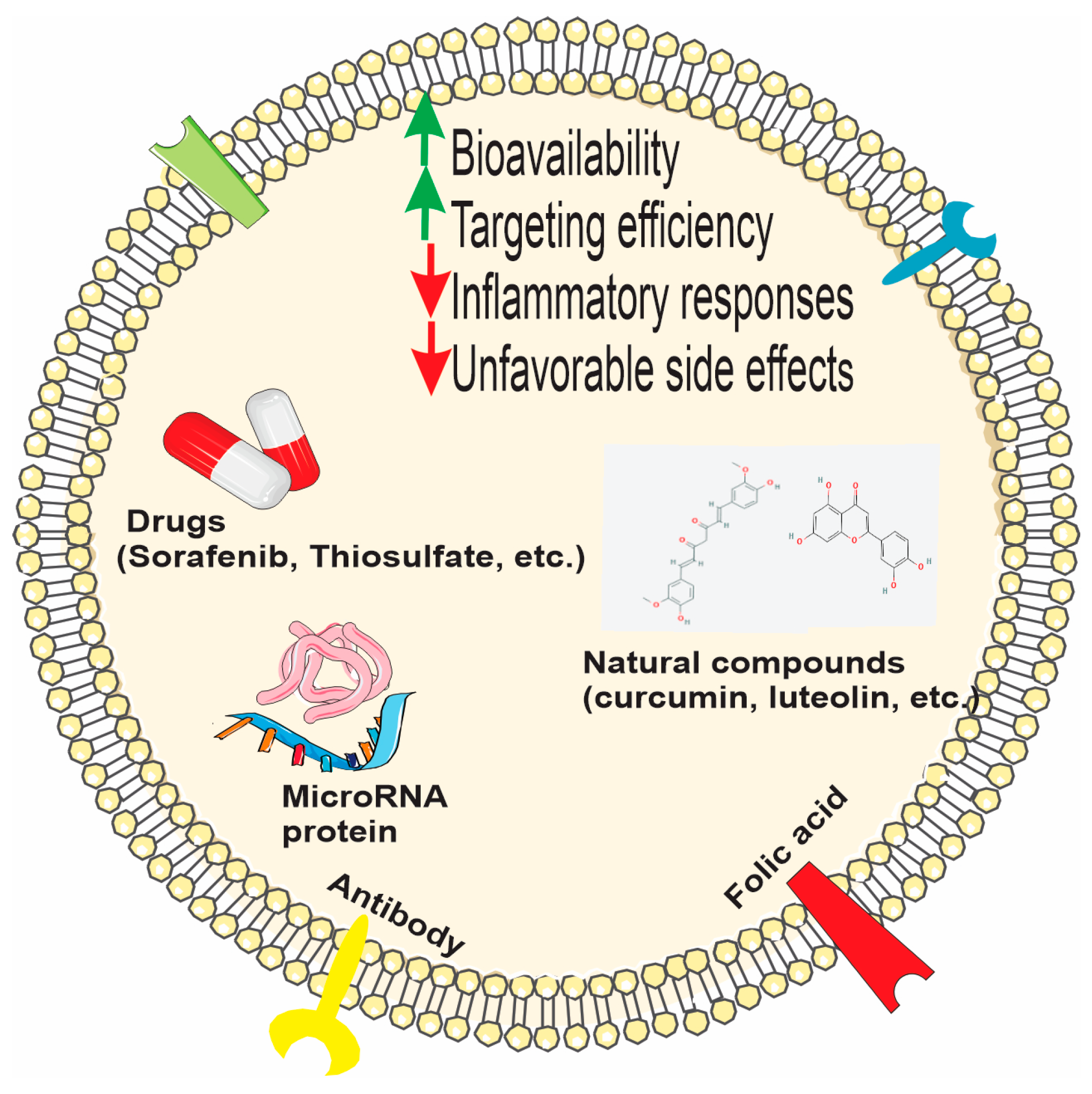
3.5.4. PDEVs and Bio-Macromolecule/Drug Delivery
PDEVs as Carriers of Natural Compounds and Drugs
Bioengineered PDEVs and Hybrid Systems for Targeted Therapies
PDEVs as Carriers of Small RNA
PDEVs as Carriers of Peptides and Proteins
PDEVs as Vaccine Platforms
3.5.5. PDEVs in Clinic
4. Final Remarks and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Design and methodology
Abbreviations
| AMPK | Adenosine monophosphate-activated protein kinase |
| BBB | Blood–brain barrier |
| CAC | Colitis-associated cancer |
| DC | Dendritic cell |
| DSS | Dextran sodium sulfate |
| DOX | Doxorubicin |
| EV | Extracellular vesicle |
| FimA | Fimbriae A |
| GIT | Gastrointestinal tract |
| HO-1 | Heme oxygenase-1 |
| HSP | Heat shock protein |
| IAV | Influenza A virus |
| IBS | Irritable bowel syndrome |
| IEC | Intestinal epithelial cell |
| IL | Interleukin |
| LDLRAP1 | Low-density lipoprotein receptor adaptor protein 1 |
| LPS | Lipopolysaccharide |
| miRNA | microRNA |
| mRNA | messenger RNA |
| mTOR | Mammalian target of rapamycin |
| MV | Membrane vesicles |
| MVB | Multivesicular body |
| NF-κB | Nuclear factor-κB |
| Nrf-2 | Nuclear factor erythroid 2-related factor 2 |
| NS1 | Nonstructural protein 1 |
| PA | Phosphatidic acid |
| PB2 | Polymerase basic protein 2 |
| PCOS | Polycystic ovary syndrome |
| PDEV | Plant-derived extracellular vesicle |
| PI3K | Phosphoinositide 3-kinases |
| ROS | Reactive oxygen species |
| TLR | Toll-like receptor |
| TNBC | Triple-negative breast cancer |
| TNF-α | Tumor necrosis factor alpha |
| TRAIL | TNF-related apoptosis-inducing ligand |
| TYR | Tyrosine |
| GC | Guanine-cytosine |
| ITGB3 | Integrin beta-3 |
| PDGFRB | Platelet-derived growth factor receptor beta |
| Lgr5 | Leucine-rich repeat-containing G protein-coupled receptor 5 |
| TCF4 | Transcription factor 4 |
| ERK | Extracellular signal-regulated kinase |
| PI3K | Phosphatidylinositol 3 kinase |
| AKT | Protein kinase B |
| ALT | Alanine aminotransferase |
| AST | Aspartate aminotransferase |
| CCR2 | CC chemokine receptor type 2 |
| CCR5 | CC chemokine receptor type 5 |
| CCR6 | CC chemokine receptor type 6 |
| ZNF773 | Zinc finger protein 773 |
| KMT2C | Lysine N-methyltransferase 2C |
| ACAN | Aggrecan |
| SOX9 | SRY-Box Transcription Factor 9 |
| COMP | Cartilage oligomeric matrix protein |
| COL | Collagen |
| HUVEC | Human umbilical vein endothelial cells |
| SIRT1 | Sirtuin 1 |
| PGC1α | Peroxisome proliferator-activated receptor-γ coactivator 1-α |
| TRP | Tyrosinase-related protein-1 |
| NLRP3 | Nucleotide-binding domain, leucine-rich-containing family, pyrin domain-containing-3 |
| FLG | Filaggrin |
| AQP3 | Aquaporin 3 |
| MAPK | Mitogen-activated protein kinase |
| LGG | Low-grade glioma |
| Cdc42 | Cell division control protein 42 homolog |
| Trkb | Tropomyosin receptor kinase B |
| GPX4 | Glutathione peroxidase 4 |
| ASBT | Apical sodium-dependent bile acid transporter |
| PPAR-α | Peroxisome proliferator-activated receptor alpha |
| UCP1 | Mitochondrial uncoupling protein 1 |
| PRDM16 | PR domain-containing 16 |
| CPT | Carnitine palmitoyltransferase |
| SREBP1-c | Sterol regulatory element binding protein 1c |
| CD | Cluster of differentiation |
| ACC | Acetyl-CoA carboxylase |
| LXR | Liver X receptor |
| Cebpα | CCAAT enhancer binding protein |
| Th17 | T helper 17 |
| JAK-STAT | Janus kinase signal transduction and activation of transcription |
| NOX4 | NADPH oxidase 4 |
| MLE-12 | Mouse lung type II epithelial cell line |
| MMP | Matrix metalloproteinase |
| Mef2D | Myocyte enhancer factor 2D |
References
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Rome, S. Biological properties of plant-derived extracellular vesicles. Food Funct. 2019, 10, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Ju, S.; Mu, J.; Dokland, T.; Zhuang, X.; Wang, Q.; Jiang, H.; Xiang, X.; Deng, Z.B.; Wang, B.; Zhang, L.; et al. Grape exosome-like nanoparticles induce intestinal stem cells and protect mice from DSS-induced colitis. Mol. Ther. 2013, 21, 1345–1357. [Google Scholar] [CrossRef] [PubMed]
- Miron, R.J.; Zhang, Y. Understanding exosomes: Part 1-Characterization, quantification and isolation techniques. Periodontol. 2000 2024, 94, 231–256. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Zhuang, X.; Wang, Q.; Jiang, H.; Deng, Z.B.; Wang, B.; Zhang, L.; Kakar, S.; Jun, Y.; Miller, D.; et al. Interspecies communication between plant and mouse gut host cells through edible plant derived exosome-like nanoparticles. Mol. Nutr. Food Res. 2014, 58, 1561–1573. [Google Scholar] [CrossRef]
- Zhang, M.; Viennois, E.; Prasad, M.; Zhang, Y.; Wang, L.; Zhang, Z.; Han, M.K.; Xiao, B.; Xu, C.; Srinivasan, S. Edible ginger-derived nanoparticles: A novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials 2016, 101, 321–340. [Google Scholar] [CrossRef]
- Saadat, Y.R.; Barar, J. Exosomes as versatile nanoscaled biocompartments in cancer therapy and/or resistance. BioImpacts 2022, 12, 87–88. [Google Scholar] [CrossRef]
- Record, M. Exosome-like nanoparticles from food: Protective nanoshuttles for bioactive cargo. Mol. Ther. 2013, 21, 1294–1296. [Google Scholar] [CrossRef]
- Xiao, Y.; Yuan, Y.; Hu, D.; Wang, H. Exosome-Derived microRNA: Potential Target for Diagnosis and Treatment of Sepsis. J. Immunol. Res. 2024, 2024, 4481452. [Google Scholar] [CrossRef]
- Macia, L.; Nanan, R.; Hosseini-Beheshti, E.; Grau, G.E. Host- and Microbiota-Derived Extracellular Vesicles, Immune Function, and Disease Development. Int. J. Mol. Sci. 2019, 21, 107. [Google Scholar] [CrossRef]
- Karabay, A.Z.; Ozkan, T.; Karadag Gurel, A.; Koc, A.; Hekmatshoar, Y.; Sunguroglu, A.; Aktan, F.; Buyukbingöl, Z. Identification of exosomal microRNAs and related hub genes associated with imatinib resistance in chronic myeloid leukemia. Naunyn. Schmiedebergs Arch. Pharmacol. 2024, 397, 9701–9721. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Deng, Z.-B.; Mu, J.; Zhang, L.; Yan, J.; Miller, D.; Feng, W.; McClain, C.J.; Zhang, H.-G. Ginger-derived nanoparticles protect against alcohol-induced liver damage. J. Extracell. Vesicles 2015, 4, 28713. [Google Scholar] [CrossRef]
- Caponnetto, F.; Manini, I.; Skrap, M.; Palmai-Pallag, T.; Di Loreto, C.; Beltrami, A.P.; Cesselli, D.; Ferrari, E. Size-dependent cellular uptake of exosomes. Nanomedicine 2017, 13, 1011–1020. [Google Scholar] [CrossRef]
- Reiner, A.T.; Somoza, V. Extracellular Vesicles as Vehicles for the Delivery of Food Bioactives. J. Agric. Food Chem. 2019, 67, 2113–2119. [Google Scholar] [CrossRef]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef]
- Maxfield, F.R.; McGraw, T.E. Endocytic recycling. Nat. Rev. Mol. Cell Biol. 2004, 5, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Kowal, J.; Tkach, M.; Théry, C. Biogenesis and secretion of exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef]
- Andreu, Z.; Yáñez-Mó, M. Tetraspanins in extracellular vesicle formation and function. Front. Immunol. 2014, 5, 442. [Google Scholar] [CrossRef]
- Simons, M.; Raposo, G. Exosomes-vesicular carriers for intercellular communication. Curr. Opin. Cell Biol. 2009, 21, 575–581. [Google Scholar] [CrossRef]
- Suga, K.; Matsui, D.; Watanabe, N.; Okamoto, Y.; Umakoshi, H. Insight into the Exosomal Membrane: From Viewpoints of Membrane Fluidity and Polarity. Langmuir 2021, 37, 11195–11202. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Shin, K.J.; Chae, Y.C. Regulation of cargo selection in exosome biogenesis and its biomedical applications in cancer. Exp. Mol. Med. 2024, 56, 877–889. [Google Scholar] [CrossRef]
- Wei, H.; Chen, Q.; Lin, L.; Sha, C.; Li, T.; Liu, Y.; Yin, X.; Xu, Y.; Chen, L.; Gao, W.; et al. Regulation of exosome production and cargo sorting. Int. J. Biol. Sci. 2021, 17, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Skryabin, G.O.; Komelkov, A.V.; Savelyeva, E.E.; Tchevkina, E.M. Lipid Rafts in Exosome Biogenesis. Biochemistry 2020, 85, 177–191. [Google Scholar] [CrossRef]
- Farahani, M.; Rubbi, C.; Liu, L.; Slupsky, J.R.; Kalakonda, N. CLL Exosomes Modulate the Transcriptome and Behaviour of Recipient Stromal Cells and Are Selectively Enriched in miR-202-3p. PLoS ONE 2015, 10, e0141429. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef]
- Pegtel, D.M.; Gould, S.J. Exosomes. Annu. Rev. Biochem. 2019, 88, 487–514. [Google Scholar] [CrossRef]
- Boavida, L.C.; Qin, P.; Broz, M.; Becker, J.D.; McCormick, S. Arabidopsis tetraspanins are confined to discrete expression domains and cell types in reproductive tissues and form homo- and heterodimers when expressed in yeast. Plant Physiol. 2013, 163, 696–712. [Google Scholar] [CrossRef]
- Rutter, B.D.; Innes, R.W. Extracellular Vesicles Isolated from the Leaf Apoplast Carry Stress-Response Proteins. Plant Physiol. 2017, 173, 728–741. [Google Scholar] [CrossRef]
- Wang, J.; Ding, Y.; Wang, J.; Hillmer, S.; Miao, Y.; Lo, S.W.; Wang, X.; Robinson, D.G.; Jiang, L. EXPO, an exocyst-positive organelle distinct from multivesicular endosomes and autophagosomes, mediates cytosol to cell wall exocytosis in Arabidopsis and tobacco cells. Plant Cell 2010, 22, 4009–4030. [Google Scholar] [CrossRef]
- Kim, K.; Yoo, H.J.; Jung, J.H.; Lee, R.; Hyun, J.K.; Park, J.H.; Na, D.; Yeon, J.H. Cytotoxic Effects of Plant Sap-Derived Extracellular Vesicles on Various Tumor Cell Types. J. Funct. Biomater. 2020, 11, 22. [Google Scholar] [CrossRef]
- Yang, M.; Liu, X.; Luo, Q.; Xu, L.; Chen, F. An efficient method to isolate lemon derived extracellular vesicles for gastric cancer therapy. J. Nanobiotechnology 2020, 18, 100. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Ko, H.J.; Kim, K.; Sohn, Y.; Min, S.Y.; Kim, J.A.; Na, D.; Yeon, J.H. Anti-melanogenic effects of extracellular vesicles derived from plant leaves and stems in mouse melanoma cells and human healthy skin. J. Extracell. Vesicles 2020, 9, 1703480. [Google Scholar] [CrossRef]
- Cox, S.N.; Porcelli, V.; Romano, S.; Palmieri, L.; Fratantonio, D. Blueberry-derived exosome like nanovesicles carry RNA cargo into HIEC-6 cells and down-regulate LPS-induced inflammatory gene expression: A proof-of-concept study. Arch. Biochem. Biophys. 2025, 764, 110266. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Zhang, M.; Liu, D.; Liang, X.; Chang, Y.; Hu, X.; Gao, W. Solanum lycopersicum derived exosome-like nanovesicles alleviate restenosis after vascular injury through the Keap1/Nrf2 pathway. Food Funct. 2025, 16, 539–553. [Google Scholar] [CrossRef]
- Shi, Y.; Wei, J.; Nie, Y.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; Sun, J. Plant-derived miR166a-3p packaged into exosomes to cross-kingdom inhibit mammary cell proliferation and promote apoptosis by targeting APLNR gene. Int. J. Biol. Macromol. 2025, 286, 138470. [Google Scholar] [CrossRef]
- Kim, J.; Li, S.; Zhang, S.; Wang, J. Plant-derived exosome-like nanoparticles and their therapeutic activities. Asian J. Pharm. Sci. 2021, 17, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Suharta, S.; Barlian, A.; Hidajah, A.C.; Notobroto, H.B.; Ana, I.D.; Indariani, S.; Wungu, T.D.K.; Wijaya, C.H. Plant-derived exosome-like nanoparticles: A concise review on its extraction methods, content, bioactivities, and potential as functional food ingredient. J. Food Sci. 2021, 86, 2838–2850. [Google Scholar] [CrossRef]
- Karamanidou, T.; Tsouknidas, A. Plant-Derived Extracellular Vesicles as Therapeutic Nanocarriers. Int. J. Mol. Sci. 2022, 23, 191. [Google Scholar] [CrossRef]
- Blackwood, R.A.; Smolen, J.E.; Transue, A.; Hessler, R.J.; Harsh, D.M.; Brower, R.C.; French, S. Phospholipase D activity facilitates Ca2+-induced aggregation and fusion of complex liposomes. Am. J. Physiol. 1997, 272, C1279–C1285. [Google Scholar] [CrossRef]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24, 637–652.e638. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhuang, X.; Deng, Z.-B.; Jiang, H.; Mu, J.; Wang, Q.; Xiang, X.; Guo, H.; Zhang, L.; Dryden, G. Targeted drug delivery to intestinal macrophages by bioactive nanovesicles released from grapefruit. Mol. Ther. 2014, 22, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, L.; Deng, J.; Ai, J.; Mo, S.; Ding, D.; Xiao, Y.; Hu, S.; Zhu, D.; Li, Q.; et al. Lipidomic analysis of plant-derived extracellular vesicles for guidance of potential anti-cancer therapy. Bioact. Mater. 2025, 46, 82–96. [Google Scholar] [CrossRef]
- Chen, Q.; Li, Q.; Liang, Y.; Zu, M.; Chen, N.; Canup, B.S.; Luo, L.; Wang, C.; Zeng, L.; Xiao, B. Natural exosome-like nanovesicles from edible tea flowers suppress metastatic breast cancer via ROS generation and microbiota modulation. Acta Pharm. Sin. B 2021, 12, 907–923. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Wang, H.; Shi, W.; Chen, L.; Chen, T.; Chen, G.; Wang, W.; Lan, J.; Huang, Z.; Zhang, J.; et al. Aloe derived nanovesicle as a functional carrier for indocyanine green encapsulation and phototherapy. J. Nanobiotechnol. 2021, 19, 439. [Google Scholar] [CrossRef]
- Berger, E.; Colosetti, P.; Jalabert, A.; Meugnier, E.; Wiklander, O.P.B.; Jouhet, J.; Errazurig-Cerda, E.; Chanon, S.; Gupta, D.; Rautureau, G.J.P.; et al. Use of Nanovesicles from Orange Juice to Reverse Diet-Induced Gut Modifications in Diet-Induced Obese Mice. Mol. Ther. Methods Clin. Dev. 2020, 18, 880–892. [Google Scholar] [CrossRef]
- Wei, Y.; Cai, X.; Wu, Q.; Liao, H.; Liang, S.; Fu, H.; Xiang, Q.; Zhang, S. Extraction, Isolation, and Component Analysis of Turmeric-Derived Exosome-like Nanoparticles. Bioengineering 2023, 10, 1199. [Google Scholar] [CrossRef]
- Yi, Q.; Xu, Z.; Thakur, A.; Zhang, K.; Liang, Q.; Liu, Y.; Yan, Y. Current understanding of plant-derived exosome-like nanoparticles in regulating the inflammatory response and immune system microenvironment. Pharmacol. Res. 2023, 190, 106733. [Google Scholar] [CrossRef]
- Woith, E.; Guerriero, G.; Hausman, J.F.; Renaut, J.; Leclercq, C.C.; Weise, C.; Legay, S.; Weng, A.; Melzig, M.F. Plant Extracellular Vesicles and Nanovesicles: Focus on Secondary Metabolites, Proteins and Lipids with Perspectives on Their Potential and Sources. Int. J. Mol. Sci. 2021, 22, 3719. [Google Scholar] [CrossRef]
- Pocsfalvi, G.; Turiák, L.; Ambrosone, A.; Del Gaudio, P.; Puska, G.; Fiume, I.; Silvestre, T.; Vékey, K. Protein biocargo of citrus fruit-derived vesicles reveals heterogeneous transport and extracellular vesicle populations. J. Plant Physiol. 2018, 229, 111–121. [Google Scholar] [CrossRef]
- De Palma, M.; Ambrosone, A.; Leone, A.; Del Gaudio, P.; Ruocco, M.; Turiák, L.; Bokka, R.; Fiume, I.; Tucci, M.; Pocsfalvi, G. Plant Roots Release Small Extracellular Vesicles with Antifungal Activity. Plants 2020, 9, 1777. [Google Scholar] [CrossRef] [PubMed]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Castelli, G.; Logozzi, M.; Mizzoni, D.; Di Raimo, R.; Cerio, A.; Dolo, V.; Pasquini, L.; Screnci, M.; Ottone, T.; Testa, U.; et al. Ex Vivo Anti-Leukemic Effect of Exosome-like Grapefruit-Derived Nanovesicles from Organic Farming-The Potential Role of Ascorbic Acid. Int. J. Mol. Sci. 2023, 24, 15663. [Google Scholar] [CrossRef] [PubMed]
- Perut, F.; Roncuzzi, L.; Avnet, S.; Massa, A.; Zini, N.; Sabbadini, S.; Giampieri, F.; Mezzetti, B.; Baldini, N. Strawberry-Derived Exosome-Like Nanoparticles Prevent Oxidative Stress in Human Mesenchymal Stromal Cells. Biomolecules 2021, 11, 87. [Google Scholar] [CrossRef]
- Li, S.; Ye, Z.; Zhao, L.; Yao, Y.; Zhou, Z. Evaluation of Antioxidant Activity and Drug Delivery Potential of Cell-Derived Extracellular Vesicles from Citrus reticulata Blanco cv. ‘Dahongpao’. Antioxidants 2023, 12, 1706. [Google Scholar] [CrossRef]
- Hao, Y.; Yang, Q.; Zhang, H.; Bai, C.; Liu, X.; Gao, Y. Ginger-Derived Extracellular Vesicles: A Natural Solution for Alopecia. Curr. Drug Deliv. 2024. ahead of print. [Google Scholar] [CrossRef]
- Tajik, T.; Baghaei, K.; Moghadam, V.E.; Farrokhi, N.; Salami, S.A. Extracellular vesicles of cannabis with high CBD content induce anticancer signaling in human hepatocellular carcinoma. Biomed. Pharmacother. 2022, 152, 113209. [Google Scholar] [CrossRef]
- Shkryl, Y.; Tsydeneshieva, Z.; Menchinskaya, E.; Rusapetova, T.; Grishchenko, O.; Mironova, A.; Bulgakov, D.; Gorpenchenko, T.; Kazarin, V.; Tchernoded, G.; et al. Exosome-like Nanoparticles, High in Trans-δ-Viniferin Derivatives, Produced from Grape Cell Cultures: Preparation, Characterization, and Anticancer Properties. Biomedicines 2024, 12, 2142. [Google Scholar] [CrossRef]
- Calzoni, E.; Bertoldi, A.; Cesaretti, A.; Alabed, H.B.R.; Cerrotti, G.; Pellegrino, R.M.; Buratta, S.; Urbanelli, L.; Emiliani, C. Aloe Extracellular Vesicles as Carriers of Photoinducible Metabolites Exhibiting Cellular Phototoxicity. Cells 2024, 13, 1845. [Google Scholar] [CrossRef]
- Chen, L.; Liu, R.; Zhu, J.; Wang, L.; Li, H.; Liu, J.; Lu, Z. Widely targeted metabolomics reveal the distribution of primary and secondary metabolites in pomegranate fruit. Food Sci. Nutr. 2024, 12, 6648–6659. [Google Scholar] [CrossRef]
- Iravani, S.; Varma, R.S. Plant-Derived Edible Nanoparticles and miRNAs: Emerging Frontier for Therapeutics and Targeted Drug-Delivery. ACS Sustain. Chem. Eng. 2019, 7, 8055–8069. [Google Scholar] [CrossRef]
- Jiang, M.; Sang, X.; Hong, Z. Beyond nutrients: Food-derived microRNAs provide cross-kingdom regulation. Bioessays 2012, 34, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, B. Plant-derived xenomiRs and cancer: Cross-kingdom gene regulation. Saudi J. Biol. Sci. 2021, 28, 2408–2422. [Google Scholar] [CrossRef]
- Shi, L.; Guo, C.; Fang, M.; Yang, Y.; Yin, F.; Shen, Y. Cross-kingdom regulation of plant microRNAs: Potential application in crop improvement and human disease therapeutics. Front. Plant Sci. 2024, 15, 1512047. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, K.; Yamamoto, Y.; Matsuoka, R.; Ochiya, T. Maintaining good miRNAs in the body keeps the doctor away?: Perspectives on the relationship between food-derived natural products and microRNAs in relation to exosomes/extracellular vesicles. Mol. Nutr. Food Res. 2018, 62, 1700080. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yang, J.; Yang, Y.; Liu, J.; Li, H.; Li, R.; Cao, C.; Shi, L.; Wu, W.; He, K. A Timely Review of Cross-Kingdom Regulation of Plant-Derived MicroRNAs. Front. Genet. 2021, 12, 613197. [Google Scholar] [CrossRef]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Res. 2012, 22, 107–126. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, X.; Liu, J.; Dong, L.; Chen, Q.; Liu, J.; Kong, H.; Zhang, Q.; Qi, X.; Hou, D. Honeysuckle-encoded atypical microRNA2911 directly targets influenza A viruses. Cell Res. 2015, 25, 39–49. [Google Scholar] [CrossRef]
- Yan, G.; Xiao, Q.; Zhao, J.; Chen, H.; Xu, Y.; Tan, M.; Peng, L. Brucea javanica derived exosome-like nanovesicles deliver miRNAs for cancer therapy. J. Control Release 2024, 367, 425–440. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Kai, T.; Zhang, L.; Li, A. Lycium ruthenicum Murray derived exosome-like nanovesicles inhibit Aβ-induced apoptosis in PC12 cells via MAPK and PI3K/AKT signaling pathways. Int. J. Biol. Macromol. 2024, 277, 134309. [Google Scholar] [CrossRef]
- Zhu, M.Z.; Xu, H.M.; Liang, Y.J.; Xu, J.; Yue, N.N.; Zhang, Y.; Tian, C.M.; Yao, J.; Wang, L.S.; Nie, Y.Q.; et al. Edible exosome-like nanoparticles from portulaca oleracea L mitigate DSS-induced colitis via facilitating double-positive CD4(+)CD8(+)T cells expansion. J. Nanobiotechnol. 2023, 21, 309. [Google Scholar] [CrossRef] [PubMed]
- Perez-Bermudez, P.; Blesa, J.; Soriano, J.M.; Marcilla, A. Extracellular vesicles in food: Experimental evidence of their secretion in grape fruits. Eur. J. Pharm. Sci. 2017, 98, 40–50. [Google Scholar] [CrossRef]
- Lian, M.Q.; Chng, W.H.; Liang, J.; Yeo, H.Q.; Lee, C.K.; Belaid, M.; Tollemeto, M.; Wacker, M.G.; Czarny, B.; Pastorin, G. Plant-derived extracellular vesicles: Recent advancements and current challenges on their use for biomedical applications. J. Extracell. Vesicles 2022, 11, 12283. [Google Scholar] [CrossRef]
- Deng, Z.; Rong, Y.; Teng, Y.; Mu, J.; Zhuang, X.; Tseng, M.; Samykutty, A.; Zhang, L.; Yan, J.; Miller, D. Broccoli-derived nanoparticle inhibits mouse colitis by activating dendritic cell AMP-activated protein kinase. Mol. Ther. 2017, 25, 1641–1654. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Liao, L.; Gao, M.; Liu, Q. Garlic-derived exosome-like nanovesicles alleviate dextran sulphate sodium-induced mouse colitis via the TLR4/MyD88/NF-κB pathway and gut microbiota modulation. Food Funct. 2023, 14, 7520–7534. [Google Scholar] [CrossRef]
- Wang, F.; Yuan, M.; Shao, C.; Ji, N.; Zhang, H.; Li, C. Momordica charantia-Derived Extracellular Vesicles Provide Antioxidant Protection in Ulcerative Colitis. Molecules 2023, 28, 6182. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Xu, J.; Huang, C.; Hu, J.X.; Xu, H.M.; Guo, X.; Zhang, Y.; Xu, J.K.; Peng, Y.; Zhang, Y.; et al. Houttuynia cordata-Derived Exosome-Like Nanoparticles Mitigate Colitis in Mice via Inhibition of the NLRP3 Signaling Pathway and Modulation of the Gut Microbiota. Int. J. Nanomed. 2024, 19, 13991–14018. [Google Scholar] [CrossRef]
- Lee, B.H.; Wu, S.C.; Chien, H.Y.; Shen, T.L.; Hsu, W.H. Tomato-fruit-derived extracellular vesicles inhibit Fusobacterium nucleatum via lipid-mediated mechanism. Food Funct. 2023, 14, 8942–8950. [Google Scholar] [CrossRef]
- You, J.Y.; Kang, S.J.; Rhee, W.J. Isolation of cabbage exosome-like nanovesicles and investigation of their biological activities in human cells. Bioact. Mater. 2021, 6, 4321–4332. [Google Scholar] [CrossRef]
- Urzì, O.; Cafora, M.; Ganji, N.R.; Tinnirello, V.; Gasparro, R.; Raccosta, S.; Manno, M.; Corsale, A.M.; Conigliaro, A.; Pistocchi, A.; et al. Lemon-derived nanovesicles achieve antioxidant and anti-inflammatory effects activating the AhR/Nrf2 signaling pathway. iScience 2023, 26, 107041. [Google Scholar] [CrossRef]
- Ramírez, O.; Pomareda, F.; Olivares, B.; Huang, Y.L.; Zavala, G.; Carrasco-Rojas, J.; Álvarez, S.; Leiva-Sabadini, C.; Hidalgo, V.; Romo, P.; et al. Aloe vera peel-derived nanovesicles display anti-inflammatory properties and prevent myofibroblast differentiation. Phytomedicine 2024, 122, 155108. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, S.; Urzì, O.; Meraviglia, S.; Di Simone, M.; Corsale, A.M.; Rabienezhad Ganji, N.; Palumbo Piccionello, A.; Polito, G.; Lo Presti, E.; Dieli, F.; et al. Anti-inflammatory properties of lemon-derived extracellular vesicles are achieved through the inhibition of ERK/NF-κB signalling pathways. J. Cell Mol. Med. 2022, 26, 4195–4209. [Google Scholar] [CrossRef] [PubMed]
- Taşlı, P.N. Usage of celery root exosome as an immune suppressant; Lipidomic characterization of apium graveolens originated exosomes and its suppressive effect on PMA/ionomycin mediated CD4(+) T lymphocyte activation. J. Food Biochem. 2022, 46, e14393. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ma, X.; Lu, Y.; Zhang, T.; Du, Z.; Xu, J.; You, J.; Chen, N.; Deng, X.; Wu, J. Edible Pueraria lobata-Derived Exosomes Promote M2 Macrophage Polarization. Molecules 2022, 27, 8184. [Google Scholar] [CrossRef]
- Iriawati, I.; Vitasasti, S.; Rahmadian, F.N.A.; Barlian, A. Isolation and characterization of plant-derived exosome-like nanoparticles from Carica papaya L. fruit and their potential as anti-inflammatory agent. PLoS ONE 2024, 19, e0304335. [Google Scholar] [CrossRef]
- Emmanuela, N.; Muhammad, D.R.; Iriawati Wijaya, C.H.; Ratnadewi, Y.M.D.; Takemori, H.; Ana, I.D.; Yuniati, R.; Handayani, W.; Wungu, T.D.K.; Tabata, Y. Isolation of plant-derived exosome-like nanoparticles (PDENs) from Solanum nigrum L. berries and Their Effect on interleukin-6 expression as a potential anti-inflammatory agent. PLoS ONE 2024, 19, e0296259. [Google Scholar] [CrossRef]
- Sánchez-López, C.M.; Manzaneque-López, M.C.; Pérez-Bermúdez, P.; Soler, C.; Marcilla, A. Characterization and bioactivity of extracellular vesicles isolated from pomegranate. Food Funct. 2022, 13, 12870–12882. [Google Scholar] [CrossRef]
- Li, S.; Zhang, R.; Wang, A.; Li, Y.; Zhang, M.; Kim, J.; Zhu, Y.; Wang, Q.; Zhang, Y.; Wei, Y.; et al. Panax notoginseng: Derived exosome-like nanoparticles attenuate ischemia reperfusion injury via altering microglia polarization. J. Nanobiotechnol. 2023, 21, 416. [Google Scholar] [CrossRef]
- Cao, Y.; Zhao, Q.; Liu, F.; Zheng, L.; Lin, X.; Pan, M.; Tan, X.; Sun, G.; Zhao, K. Drug Value of Drynariae Rhizoma Root-Derived Extracellular Vesicles for Neurodegenerative Diseases Based on Proteomics and Bioinformatics. Plant Signal. Behav. 2022, 17, 2129290. [Google Scholar] [CrossRef]
- Zhao, X.; Yin, F.; Fu, L.; Ma, Y.; Ye, L.; Huang, Y.; Fan, W.; Gao, W.; Cai, Y.; Mou, X. Garlic-derived exosome-like nanovesicles as a hepatoprotective agent alleviating acute liver failure by inhibiting CCR2/CCR5 signaling and inflammation. Biomater. Adv. 2023, 154, 213592. [Google Scholar] [CrossRef]
- Ou, X.; Wang, H.; Tie, H.; Liao, J.; Luo, Y.; Huang, W.; Yu, R.; Song, L.; Zhu, J. Novel plant-derived exosome-like nanovesicles from Catharanthus roseus: Preparation, characterization, and immunostimulatory effect via TNF-α/NF-κB/PU.1 axis. J. Nanobiotechnol. 2023, 21, 160. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; Lee, M.; Munir, J. Exosomes in Food: Health Benefits and Clinical Relevance in Diseases. Adv. Nutr. 2020, 11, 687–696. [Google Scholar] [CrossRef]
- Raimondo, S.; Naselli, F.; Fontana, S.; Monteleone, F.; Dico, A.L.; Saieva, L.; Zito, G.; Flugy, A.; Manno, M.; Di Bella, M.A. Citrus limon-derived nanovesicles inhibit cancer cell proliferation and suppress CML xenograft growth by inducing TRAIL-mediated cell death. Oncotarget 2015, 6, 19514. [Google Scholar] [CrossRef]
- Ozkan, I.; Kocak, P.; Yildirim, M.; Unsal, N.; Yilmaz, H.; Telci, D.; Sahin, F. Garlic (Allium sativum)-derived SEVs inhibit cancer cell proliferation and induce caspase mediated apoptosis. Sci. Rep. 2021, 11, 14773. [Google Scholar] [CrossRef]
- Kantarcıoğlu, M.; Yıldırım, G.; Akpınar Oktar, P.; Yanbakan, S.; Özer, Z.B.; Yurtsever Sarıca, D.; Taşdelen, S.; Bayrak, E.; Akın Balı, D.F.; Öztürk, S.; et al. Coffee-Derived Exosome-Like Nanoparticles: Are They the Secret Heroes? Turk. J. Gastroenterol. 2023, 34, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zu, M.; Gong, H.; Ma, Y.; Sun, J.; Ran, S.; Shi, X.; Zhang, J.; Xiao, B. Tea leaf-derived exosome-like nanotherapeutics retard breast tumor growth by pro-apoptosis and microbiota modulation. J. Nanobiotechnol. 2023, 21, 6. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Perini, G.; Minopoli, A.; Augello, A.; De Spirito, M.; Palmieri, V.; Papi, M. Plant-derived extracellular vesicles release combined with systemic DOX exhibits synergistic effects in 3D bioprinted triple-negative breast cancer. Biomed. Pharmacother. 2024, 181, 117637. [Google Scholar] [CrossRef]
- Cui, L.; Perini, G.; Augello, A.; Palmieri, V.; De Spirito, M.; Papi, M. Plant-derived extracellular nanovesicles: A promising biomedical approach for effective targeting of triple negative breast cancer cells. Front. Bioeng. Biotechnol. 2024, 12, 1390708. [Google Scholar] [CrossRef]
- Shi, C.; Huang, K.; Soto, J.; Sankaran, R.; Kalia, V.; Onwumere, O.; Young, M.; Einbond, L.; Redenti, S. Piperlongumine inhibits proliferation and oncogenic MYCN expression in chemoresistant metastatic retinoblastoma cells directly and through extracellular vesicles. Biomed. Pharmacother. 2023, 161, 114554. [Google Scholar] [CrossRef]
- Chintapula, U.; Oh, D.; Perez, C.; Davis, S.; Ko, J. Anti-cancer bioactivity of sweet basil leaf derived extracellular vesicles on pancreatic cancer cells. J. Extracell. Biol. 2024, 3, e142. [Google Scholar] [CrossRef]
- Kim, J.; Zhu, Y.; Chen, S.; Wang, D.; Zhang, S.; Xia, J.; Li, S.; Qiu, Q.; Lee, H.; Wang, J. Anti-glioma effect of ginseng-derived exosomes-like nanoparticles by active blood-brain-barrier penetration and tumor microenvironment modulation. J. Nanobiotechnol. 2023, 21, 253. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.M.; Palmquist, J.; Huang, S.D.; Jin, H. Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef]
- Sundaram, K.; Miller, D.P.; Kumar, A.; Teng, Y.; Sayed, M.; Mu, J.; Lei, C.; Sriwastva, M.K.; Zhang, L.; Yan, J.; et al. Plant-Derived Exosomal Nanoparticles Inhibit Pathogenicity of Porphyromonas gingivalis. iScience 2019, 21, 308–327. [Google Scholar] [CrossRef]
- Sarwa, K.K.; Das, P.J.; Mazumder, B. A nanovesicle topical formulation of Bhut Jolokia (hottest capsicum): A potential anti-arthritic medicine. Expert Opin. Drug Deliv. 2014, 11, 661–676. [Google Scholar] [CrossRef]
- Yıldırım, M.; Ünsal, N.; Kabataş, B.; Eren, O.; Şahin, F. Effect of Solanum lycopersicum and Citrus limon-Derived Exosome-Like Vesicles on Chondrogenic Differentiation of Adipose-Derived Stem Cells. Appl. Biochem. Biotechnol. 2024, 196, 203–219. [Google Scholar] [CrossRef]
- Seo, K.; Yoo, J.H.; Kim, J.; Min, S.J.; Heo, D.N.; Kwon, I.K.; Moon, H.J. Ginseng-derived exosome-like nanovesicles extracted by sucrose gradient ultracentrifugation to inhibit osteoclast differentiation. Nanoscale 2023, 15, 5798–5808. [Google Scholar] [CrossRef]
- Hwang, J.H.; Park, Y.S.; Kim, H.S.; Kim, D.H.; Lee, S.H.; Lee, C.H.; Lee, S.H.; Kim, J.E.; Lee, S.; Kim, H.M.; et al. Yam-derived exosome-like nanovesicles stimulate osteoblast formation and prevent osteoporosis in mice. J. Control. Release 2023, 355, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Şahin, F.; Koçak, P.; Güneş, M.Y.; Özkan, İ.; Yıldırım, E.; Kala, E.Y. In Vitro Wound Healing Activity of Wheat-Derived Nanovesicles. Appl. Biochem. Biotechnol. 2018, 188, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Mahdipour, E. Beta vulgaris juice contains biologically active exosome-like nanoparticles. Tissue Cell 2022, 76, 101800. [Google Scholar] [CrossRef]
- Kim, M.; Park, J.H. Isolation of Aloe saponaria-Derived Extracellular Vesicles and Investigation of Their Potential for Chronic Wound Healing. Pharmaceutics 2022, 14, 1905. [Google Scholar] [CrossRef]
- Gupta, R.; Gupta, S.; Gupta, P.; Nüssler, A.K.; Kumar, A. Establishing the Callus-Based Isolation of Extracellular Vesicles from Cissus quadrangularis and Elucidating Their Role in Osteogenic Differentiation. J. Funct. Biomater. 2023, 14, 540. [Google Scholar] [CrossRef] [PubMed]
- Byun, K.A.; Park, Y.; Oh, S.; Batsukh, S.; Son, K.H.; Byun, K. Co-Treatment with Phlorotannin and Extracellular Vesicles from Ecklonia cava Inhibits UV-Induced Melanogenesis. Antioxidants 2024, 13, 408. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Ko, H.; Kim, J.Y.; Kim, H.J.; Kim, H.Y.; Cho, H.E.; Cho, H.D.; Seo, W.S.; Kang, H.C. Improvement in Yield of Extracellular Vesicles Derived from Edelweiss Callus Treated with LED Light and Enhancement of Skin Anti-Aging Indicators. Curr. Issues Mol. Biol. 2023, 45, 10159–10178. [Google Scholar] [CrossRef]
- Ishida, T.; Morisawa, S.; Jobu, K.; Kawada, K.; Yoshioka, S.; Miyamura, M. Atractylodes lancea rhizome derived exosome-like nanoparticles prevent alpha-melanocyte stimulating hormone-induced melanogenesis in B16-F10 melanoma cells. Biochem. Biophys. Rep. 2023, 35, 101530. [Google Scholar] [CrossRef] [PubMed]
- Batsukh, S.; Oh, S.; Lee, J.M.; Joo, J.H.J.; Son, K.H.; Byun, K. Extracellular Vesicles from Ecklonia cava and Phlorotannin Promote Rejuvenation in Aged Skin. Mar. Drugs 2024, 22, 223. [Google Scholar] [CrossRef]
- Han, J.; Wu, T.; Jin, J.; Li, Z.; Cheng, W.; Dai, X.; Yang, K.; Zhang, H.; Zhang, Z.; Zhang, H.; et al. Exosome-like nanovesicles derived from Phellinus linteus inhibit Mical2 expression through cross-kingdom regulation and inhibit ultraviolet-induced skin aging. J. Nanobiotechnol. 2022, 20, 455. [Google Scholar] [CrossRef]
- Spinler, J.K.; Oezguen, N.; Runge, J.K.; Luna, R.A.; Karri, V.; Yang, J.; Hirschi, K.D. Dietary impact of a plant-derived microRNA on the gut microbiome. ExRNA 2020, 2, 11. [Google Scholar] [CrossRef]
- Cui, Z.; Liu, T.; Wen, Y.; Li, W.; Xu, J.; Chen, Y.; Chen, D.; Zhu, Y. Oral administration of cranberry-derived exosomes attenuates murine premature ovarian failure in association with changes in the specific gut microbiota and diminution in ovarian granulosa cell PANoptosis. Food Funct. 2024, 15, 11697–11714. [Google Scholar] [CrossRef]
- Timms, K.; Holder, B.; Day, A.; McLaughlin, J.; Forbes, K.A.; Westwood, M. Watermelon-Derived Extracellular Vesicles Influence Human Ex Vivo Placental Cell Behavior by Altering Intestinal Secretions. Mol. Nutr. Food Res. 2022, 66, e2200013. [Google Scholar] [CrossRef]
- Hong, R.; Luo, L.; Wang, L.; Hu, Z.L.; Yin, Q.R.; Li, M.; Gu, B.; Wang, B.; Zhuang, T.; Zhang, X.Y.; et al. Lepidium meyenii Walp (Maca)-derived extracellular vesicles ameliorate depression by promoting 5-HT synthesis via the modulation of gut-brain axis. Imeta 2023, 2, e116. [Google Scholar] [CrossRef]
- Zhang, W.; Song, Q.; Bi, X.; Cui, W.; Fang, C.; Gao, J.; Li, J.; Wang, X.; Qu, K.; Qin, X.; et al. Preparation of Pueraria lobata Root-Derived Exosome-Like Nanovesicles and Evaluation of Their Effects on Mitigating Alcoholic Intoxication and Promoting Alcohol Metabolism in Mice. Int. J. Nanomed. 2024, 19, 4907–4921. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Pan, J. Beta vulgaris-derived exosome-like nanovesicles alleviate chronic doxorubicin-induced cardiotoxicity by inhibiting ferroptosis. J. Biochem. Mol. Toxicol. 2024, 38, e23540. [Google Scholar] [CrossRef]
- Yang, S.; Guo, J.; Chen, D.; Sun, Z.; Pu, L.; Sun, G.; Yang, M.; Peng, Y. The Cardioprotective Effect of Ginseng Derived Exosomes via Inhibition of Oxidative Stress and Apoptosis. ACS Appl. Bio Mater. 2025, 8, 814–824. [Google Scholar] [CrossRef]
- Cui, W.W.; Ye, C.; Wang, K.X.; Yang, X.; Zhu, P.Y.; Hu, K.; Lan, T.; Huang, L.Y.; Wang, W.; Gu, B.; et al. Momordica. charantia-Derived Extracellular Vesicles-Like Nanovesicles Protect Cardiomyocytes Against Radiation Injury via Attenuating DNA Damage and Mitochondria Dysfunction. Front. Cardiovasc. Med. 2022, 9, 864188. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.J.; Won, J.P.; Lee, H.G.; Seo, H.G. Green Onion-Derived Exosome-like Nanoparticles Prevent Ferroptotic Cell Death Triggered by Glutamate: Implication for GPX4 Expression. Nutrients 2024, 16, 3257. [Google Scholar] [CrossRef]
- Vestuto, V.; Conte, M.; Vietri, M.; Mensitieri, F.; Santoro, V.; Di Muro, A.; Alfieri, M.; Moros, M.; Miranda, M.R.; Amante, C.; et al. Multiomic Profiling and Neuroprotective Bioactivity of Salvia Hairy Root-Derived Extracellular Vesicles in a Cellular Model of Parkinson’s Disease. Int. J. Nanomed. 2024, 19, 9373–9393. [Google Scholar] [CrossRef]
- Usui, S.; Zhu, Q.; Komori, H.; Iwamoto, Y.; Nishiuchi, T.; Shirasaka, Y.; Tamai, I. Apple-derived extracellular vesicles modulate the expression of human intestinal bile acid transporter ASBT/SLC10A2 via downregulation of transcription factor RARα. Drug Metab. Pharmacokinet. 2023, 52, 100512. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Song, Q.; Shaw, P.C.; Wu, Y.; Zuo, Z.; Yu, R. Tangerine Peel-Derived Exosome-Like Nanovesicles Alleviate Hepatic Steatosis Induced by Type 2 Diabetes: Evidenced by Regulating Lipid Metabolism and Intestinal Microflora. Int. J. Nanomed. 2024, 19, 10023–10043. [Google Scholar] [CrossRef]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Agrawal, A.K.; Gupta, R. Exosomes for the Enhanced Tissue Bioavailability and Efficacy of Curcumin. AAPS J. 2017, 19, 1691–1702. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, M.; Merlin, D. Advances in plant-derived edible nanoparticle-based lipid nano-drug delivery systems as therapeutic nanomedicines. J. Mater. Chem. B 2018, 6, 1312–1321. [Google Scholar] [CrossRef]
- Sarvarian, P.; Samadi, P.; Gholipour, E.; Shams Asenjan, K.; Hojjat-Farsangi, M.; Motavalli, R.; Motavalli Khiavi, F.; Yousefi, M. Application of Emerging Plant-Derived Nanoparticles as a Novel Approach for Nano-Drug Delivery Systems. Immunol. Investig. 2021, 51, 1039–1059. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, S.; Giavaresi, G.; Lorico, A.; Alessandro, R. Extracellular Vesicles as Biological Shuttles for Targeted Therapies. Int. J. Mol. Sci. 2019, 20, 1848. [Google Scholar] [CrossRef] [PubMed]
- Kameli, N.; Dragojlovic-Kerkache, A.; Savelkoul, P.; Stassen, F.R. Plant-Derived Extracellular Vesicles: Current Findings, Challenges, and Future Applications. Membranes 2021, 11, 411. [Google Scholar] [CrossRef] [PubMed]
- Mammadova, R.; Maggio, S.; Fiume, I.; Bokka, R.; Moubarak, M.; Gellén, G.; Schlosser, G.; Adamo, G.; Bongiovanni, A.; Trepiccione, F.; et al. Protein Biocargo and Anti-Inflammatory Effect of Tomato Fruit-Derived Nanovesicles Separated by Density Gradient Ultracentrifugation and Loaded with Curcumin. Pharmaceutics 2023, 15, 333. [Google Scholar] [CrossRef]
- Wang, Q.; Zhuang, X.; Mu, J.; Deng, Z.-B.; Jiang, H.; Zhang, L.; Xiang, X.; Wang, B.; Yan, J.; Miller, D. Delivery of therapeutic agents by nanoparticles made of grapefruit-derived lipids. Nat. Commun. 2013, 4, 1867. [Google Scholar] [CrossRef]
- Jiang, D.; Li, Z.; Liu, H.; Liu, H.; Xia, X.; Xiang, X. Plant exosome-like nanovesicles derived from sesame leaves as carriers for luteolin delivery: Molecular docking, stability and bioactivity. Food Chem. 2024, 438, 137963. [Google Scholar] [CrossRef]
- Feng, W.; Teng, Y.; Zhong, Q.; Zhang, Y.; Zhang, J.; Zhao, P.; Chen, G.; Wang, C.; Liang, X.J.; Ou, C. Biomimetic Grapefruit-Derived Extracellular Vesicles for Safe and Targeted Delivery of Sodium Thiosulfate against Vascular Calcification. ACS Nano 2023, 17, 24773–24789. [Google Scholar] [CrossRef]
- Fang, Z.; Song, M.; Lai, K.; Cui, M.; Yin, M.; Liu, K. Kiwi-derived extracellular vesicles for oral delivery of sorafenib. Eur. J. Pharm. Sci. 2023, 191, 106604. [Google Scholar] [CrossRef]
- Wang, Q.; Ren, Y.; Mu, J.; Egilmez, N.K.; Zhuang, X.; Deng, Z.; Zhang, L.; Yan, J.; Miller, D.; Zhang, H.-G. Grapefruit-derived nanovectors use an activated leukocyte trafficking pathway to deliver therapeutic agents to inflammatory tumor sites. Cancer Res. 2015, 75, 2520–2529. [Google Scholar] [CrossRef]
- Huang, R.; Jia, B.; Su, D.; Li, M.; Xu, Z.; He, C.; Huang, Y.; Fan, H.; Chen, H.; Cheng, F. Plant exosomes fused with engineered mesenchymal stem cell-derived nanovesicles for synergistic therapy of autoimmune skin disorders. J. Extracell. Vesicles 2023, 12, e12361. [Google Scholar] [CrossRef]
- Zhang, M.; Xiao, B.; Wang, H.; Han, M.K.; Zhang, Z.; Viennois, E.; Xu, C.; Merlin, D. Edible ginger-derived nano-lipids loaded with doxorubicin as a novel drug-delivery approach for colon cancer therapy. Mol. Ther. 2016, 24, 1783–1796. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Zhang, Y.; Gong, Y.; Li, G.; Pan, J.; Dou, D.; Ma, K.; Cui, C.; Liu, Y.; Zhu, X. Antibody functionalized curcuma-derived extracellular vesicles loaded with doxorubicin overcome therapy-induced senescence and enhance chemotherapy. J. Control Release 2025, 379, 377–389. [Google Scholar] [CrossRef]
- López de Las Hazas, M.C.; Tomé-Carneiro, J.; Del Pozo-Acebo, L.; Del Saz-Lara, A.; Chapado, L.A.; Balaguer, L.; Rojo, E.; Espín, J.C.; Crespo, C.; Moreno, D.A.; et al. Therapeutic potential of plant-derived extracellular vesicles as nanocarriers for exogenous miRNAs. Pharmacol. Res. 2023, 198, 106999. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Teng, Y.; Samykutty, A.; Mu, J.; Deng, Z.; Zhang, L.; Cao, P.; Rong, Y.; Yan, J.; Miller, D. Grapefruit-derived nanovectors delivering therapeutic miR17 through an intranasal route inhibit brain tumor progression. Mol. Ther. 2016, 24, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Liu, K.; Wang, F.; Fei, X.; Niu, C.; Li, T.; Liu, L. Neutrophil membrane-engineered Panax ginseng root-derived exosomes loaded miRNA 182-5p targets NOX4/Drp-1/NLRP3 signal pathway to alleviate acute lung injury in sepsis: Experimental studies. Int. J. Surg. 2024, 110, 72–86. [Google Scholar] [CrossRef]
- Rabienezhad Ganji, N.; Urzì, O.; Tinnirello, V.; Costanzo, E.; Polito, G.; Palumbo Piccionello, A.; Manno, M.; Raccosta, S.; Gallo, A.; Lo Pinto, M.; et al. Proof-of-Concept Study on the Use of Tangerine-Derived Nanovesicles as siRNA Delivery Vehicles toward Colorectal Cancer Cell Line SW480. Int. J. Mol. Sci. 2023, 25, 546. [Google Scholar] [CrossRef]
- Cui, C.; Du, M.; Zhao, Y.; Tang, J.; Liu, M.; Min, G.; Chen, R.; Zhang, Q.; Sun, Z.; Weng, H. Functional Ginger-Derived Extracellular Vesicles-Coated ZIF-8 Containing TNF-α siRNA for Ulcerative Colitis Therapy by Modulating Gut Microbiota. ACS Appl. Mater. Interfaces 2024, 16, 53460–53473. [Google Scholar] [CrossRef]
- Liu, Y.; Qi, H.; Zong, J.; Li, M.; Yang, Y.; Li, X.; Li, T.; Cho, J.Y.; Yu, T. Oral Piwi-Interacting RNA Delivery Mediated by Green Tea-Derived Exosome-Like Nanovesicles for the Treatment of Aortic Dissection. Adv. Healthc. Mater. 2024, 13, e2401466. [Google Scholar] [CrossRef]
- Kilasoniya, A.; Garaeva, L.; Shtam, T.; Spitsyna, A.; Putevich, E.; Moreno-Chamba, B.; Salazar-Bermeo, J.; Komarova, E.; Malek, A.; Valero, M.; et al. Potential of Plant Exosome Vesicles from Grapefruit (Citrus × paradisi) and Tomato (Solanum lycopersicum) Juices as Functional Ingredients and Targeted Drug Delivery Vehicles. Antioxidants 2023, 12, 943. [Google Scholar] [CrossRef]
- Hou, J.; Wei, W.; Geng, Z.; Zhang, Z.; Yang, H.; Zhang, X.; Li, L.; Gao, Q. Developing Plant Exosomes as an Advanced Delivery System for Cosmetic Peptide. ACS Appl. Bio Mater. 2024, 7, 3050–3060. [Google Scholar] [CrossRef]
- Pomatto, M.A.C.; Gai, C.; Negro, F.; Massari, L.; Deregibus, M.C.; De Rosa, F.G.; Camussi, G. Oral Delivery of mRNA Vaccine by Plant-Derived Extracellular Vesicle Carriers. Cells 2023, 12, 1826. [Google Scholar] [CrossRef] [PubMed]
- Pomatto, M.A.C.; Gai, C.; Negro, F.; Massari, L.; Deregibus, M.C.; Grange, C.; De Rosa, F.G.; Camussi, G. Plant-Derived Extracellular Vesicles as a Delivery Platform for RNA-Based Vaccine: Feasibility Study of an Oral and Intranasal SARS-CoV-2 Vaccine. Pharmaceutics 2023, 15, 974. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karabay, A.Z.; Barar, J.; Hekmatshoar, Y.; Rahbar Saadat, Y. Multifaceted Therapeutic Potential of Plant-Derived Exosomes: Immunomodulation, Anticancer, Anti-Aging, Anti-Melanogenesis, Detoxification, and Drug Delivery. Biomolecules 2025, 15, 394. https://doi.org/10.3390/biom15030394
Karabay AZ, Barar J, Hekmatshoar Y, Rahbar Saadat Y. Multifaceted Therapeutic Potential of Plant-Derived Exosomes: Immunomodulation, Anticancer, Anti-Aging, Anti-Melanogenesis, Detoxification, and Drug Delivery. Biomolecules. 2025; 15(3):394. https://doi.org/10.3390/biom15030394
Chicago/Turabian StyleKarabay, Arzu Zeynep, Jaleh Barar, Yalda Hekmatshoar, and Yalda Rahbar Saadat. 2025. "Multifaceted Therapeutic Potential of Plant-Derived Exosomes: Immunomodulation, Anticancer, Anti-Aging, Anti-Melanogenesis, Detoxification, and Drug Delivery" Biomolecules 15, no. 3: 394. https://doi.org/10.3390/biom15030394
APA StyleKarabay, A. Z., Barar, J., Hekmatshoar, Y., & Rahbar Saadat, Y. (2025). Multifaceted Therapeutic Potential of Plant-Derived Exosomes: Immunomodulation, Anticancer, Anti-Aging, Anti-Melanogenesis, Detoxification, and Drug Delivery. Biomolecules, 15(3), 394. https://doi.org/10.3390/biom15030394