
Comprehensive Bio-Screening of Phytochemistry and Biological Capacity of Oregano (Origanum vulgare) and Salvia triloba Extracts against Oral Cariogenic and Food-Origin Pathogenic Bacteria

 ,
,  , ,
, ,  , ,
, ,  ,
,  and
and
Abstract
1. Introduction
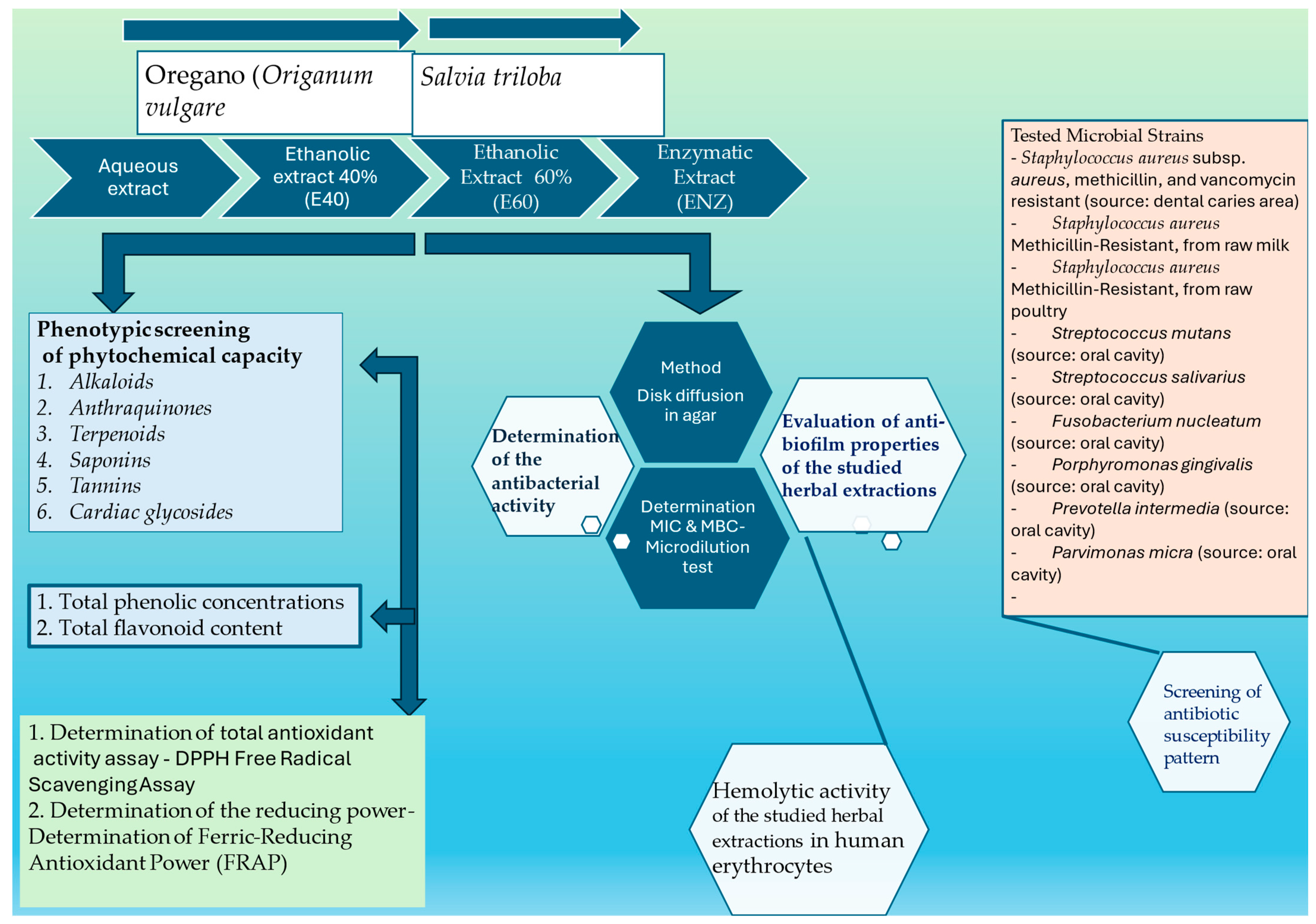
2. Materials and Methods
2.1. Plant Materials
2.1.1. Preparation Methods for Plant Extracts
2.1.2. Aqueous Extract (A)
2.1.3. Ethanolic Extract (E)
2.1.4. Enzymatic Extract (ENZ)
2.2. Screening the Phytochemical Capacity and Antioxidant Activities of Plant Extracts
2.2.1. Phytochemical Identification
- -
- -
- -
- Detection of terpenoids
- -
- Detection of saponins (foam test) [32]
- -
- Detection of tannins (ferric chloride test—Braymer’s test) [33]
- -
2.2.2. Total Phenolic Concentrations [34]
2.2.3. Estimation of Total Flavonoid Content
2.2.4. Total Antioxidant Activity Assay [35] (DPPH Free-Radical Scavenging Assay)
2.2.5. The Reducing Power Assay
2.3. Determination of the Antibacterial Activity
2.3.1. Tested Bacterial Strains
Phenotypic Screening of Antibiotic Susceptibility Pattern of Bacterial Strains
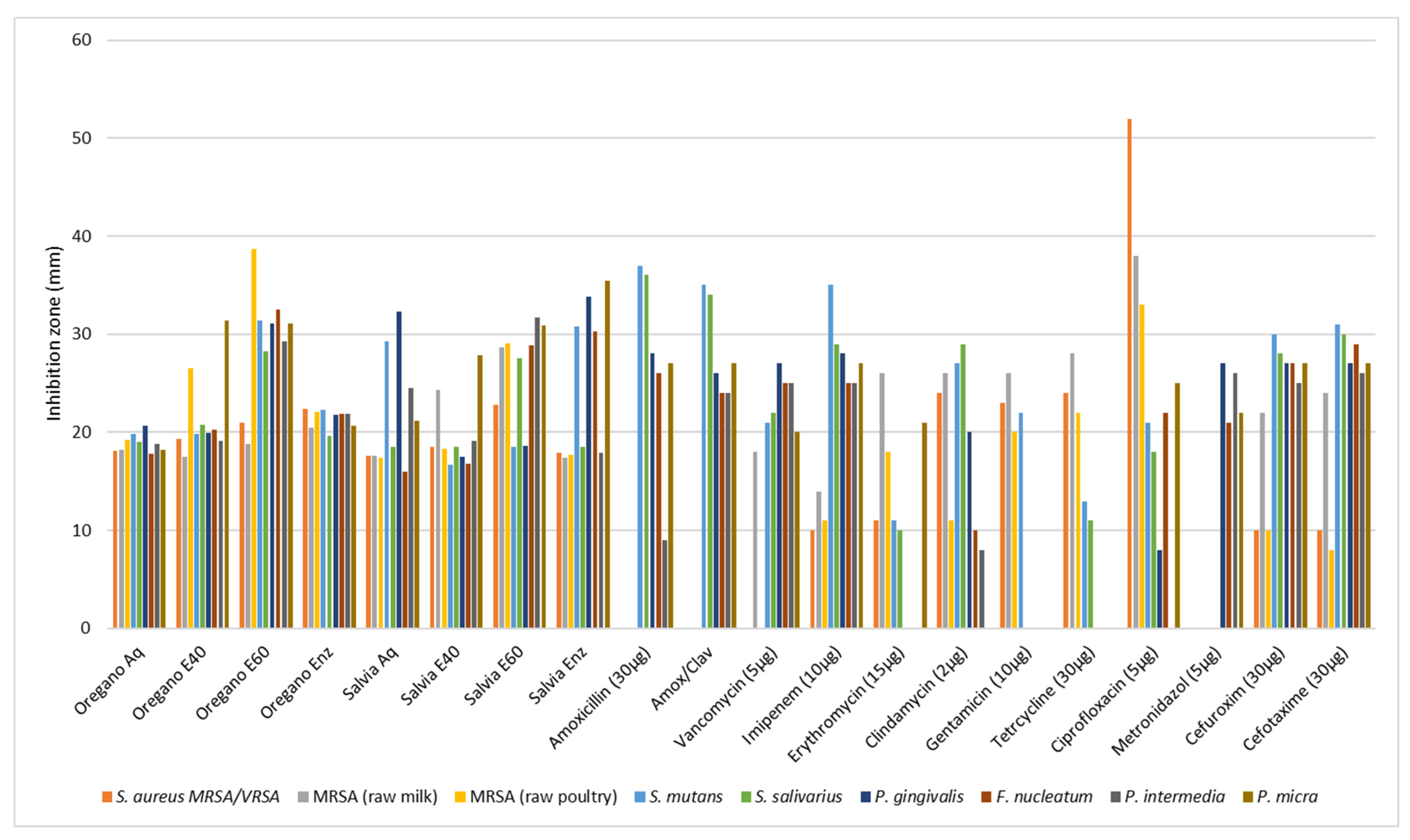
2.3.2. First Assay for Evaluation of the Antibacterial Activity—Diffusion in Agar Test
2.3.3. Second Assay for Evaluation of the Antibacterial Activity—Determination of Minimum Inhibitory Concentration and Minimum Bactericidal Concentration—Microdilution Broth Test
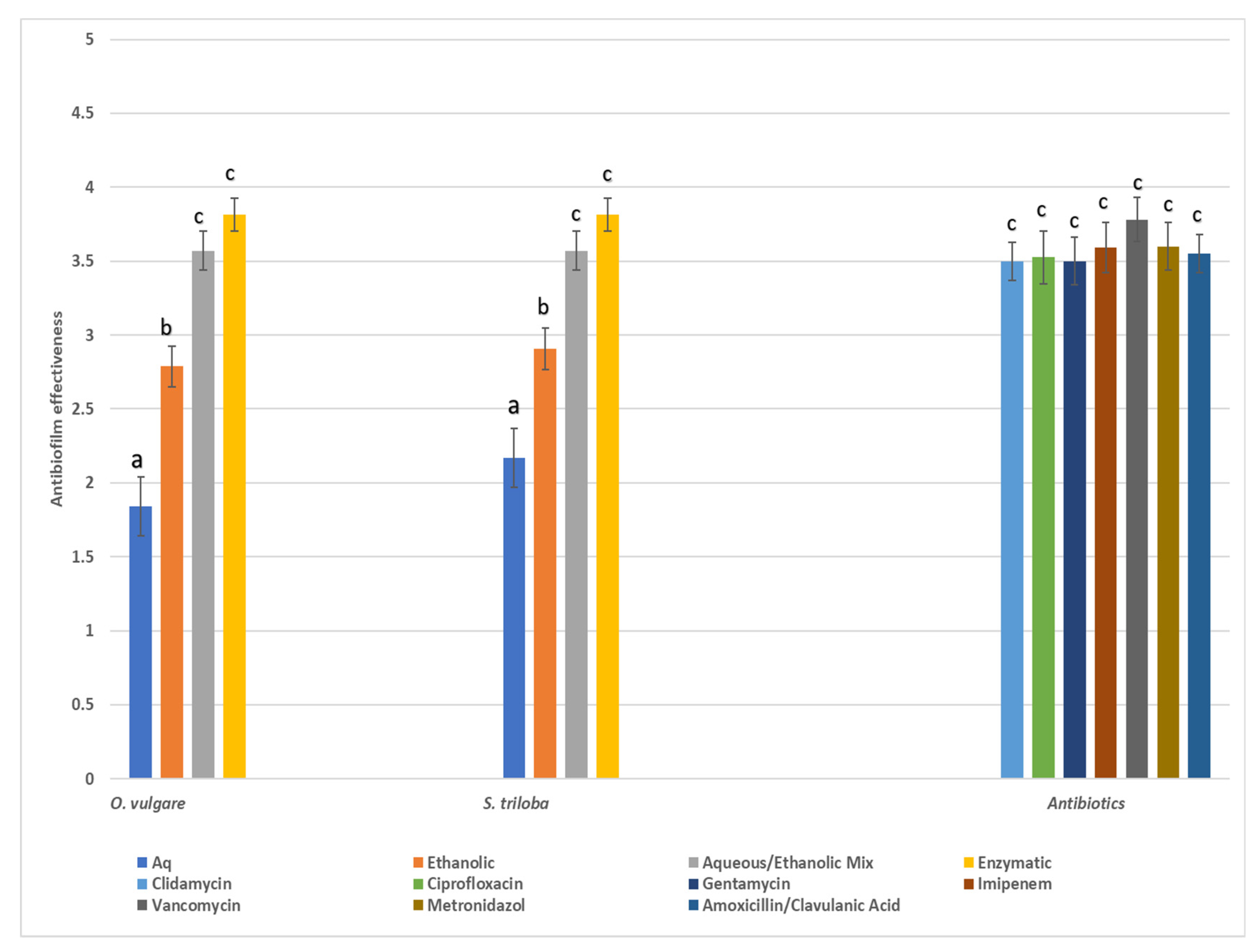
2.4. In Vitro Evaluation of Antibacterial Efficacies—Anti-Biofilm Properties of the Studied Herbal Extractions
2.5. Hemolytic Potential of Tested Plant Extracts in Human Erythrocytes
2.6. Statistical Analysis
3. Results
4. Discussion
4.1. Pattern of Total Phenolics and Flavonoids Content
4.2. Evaluation of Antioxidant Capacity: DPPH Assay and Ferric-Reducing Assay Power (FRAP)
4.3. Antimicrobial and Anti-Biofilm Activity of the Studied Extracts
4.3.1. O. vulgare Performance
4.3.2. S. tribola Performance
4.4. Hemolytic Activity
4.5. General Aspects
5. Conclusions
- This study reports on the multifactorial dependence of phytochemical isolation on factors such as extraction methods, analytical techniques, and plant species’ unique characteristics. Despite variations in extraction methodologies, all types of extracts demonstrated significant antioxidant capacity, highlighting the potential health benefits of these plants.
- Additionally, the extracts exhibited promising antibacterial activity against a diverse range of oral pathogens, including antibiotic-resistant strains, and showed efficacy in inhibiting oral biofilm formation. While this study has limitations, such as its focus on only two plant species and methodological constraints, it provides a foundation for future research to explore the mechanisms underlying the observed bioactivities and validate the efficacy and safety of plant-derived treatments through in vivo models or clinical trials.
- The extracts from both plants showed an exceptional antibacterial action against the two S. aureus strains which originated from poultry meat and raw milk. This finding is essential for the future development of food preservatives based on these extracts.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Antoniadou, M.; Rozos, G.; Vaiou, N.; Zaralis, K.; Ersanli, C.; Alexopoulos, A.; Tzora, A.; Varzakas, T.; Voidarou, C.C. The In Vitro Assessment of Antibacterial and Antioxidant Efficacy in Rosa damascena and Hypericum perforatum Extracts against Pathogenic Strains in the Interplay of Dental Caries, Oral Health, and Food Microbiota. Microorganisms 2023, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Rezabakhsh, A.; Mahmoodpoor, A.; Soleimanpour, H. Historical perspective of aspirin: A journey from discovery to clinical practice Ancient and modern history. J. Cardiovasc. Thorac. Res. 2021, 13, 179–180. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Camele, I.; Mohamed, A.A. A Comprehensive Review on the Biological, Agricultural and Pharmaceutical Properties of Secondary Metabolites Based-Plant Origin. Int. J. Mol. Sci. 2023, 24, 3266. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. Biochemistry and Molecular Biology of Plants. Rockville; Buchanan, B.B., Gruissem, W., Eds.; American Society of Plant Phys Iologists: Rockville, MD, USA, 2000. [Google Scholar]
- Lambers, H.; Chapin, F.S., III; Pons, T.L. Ecological Biochemistry: Allelopathy and Defense Against Herbivores. In Plant Physiological Ecology; Springer: New York, NY, USA, 2008; pp. 445–477. [Google Scholar] [CrossRef]
- Alibi, S.; Crespo, D.; Navas, J. Plant-Derivatives Small Molecules with Antibacterial Activity. Antibiotics 2021, 10, 231. [Google Scholar] [CrossRef] [PubMed]
- Bajguz, A.; Piotrowska-Niczyporuk, A. Biosynthetic Pathways of Hormones in Plants. Metabolites 2023, 13, 884. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Shakeri, A.; Iranshahi, M.; Boozari, M. A Review of the Phytochemistry and Antimicrobial Properties of Origanum vulgare L. and Subspecies. Iran. J. Pharm. Res. 2021, 20, 268–285. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Luo, Y.; Zhao, H. Synthetic biology: Putting synthesis into biology. Wiley Interdiscip. Rev. Syst. Biol. Med. 2011, 3, 7–20. [Google Scholar] [CrossRef]
- Caselli, E.; Fabbri, C.; D’Accolti, M.; Soffritti, I.; Bassi, C.; Mazzacane, S.; Franchi, M. Defining the oral microbiome by whole-genome sequencing and resistome analysis: The complexity of the healthy picture. BMC Microbiol. 2020, 20, 120. [Google Scholar] [CrossRef]
- Pisano, M.; Giordano, F.; Sangiovanni, G.; Capuano, N.; Acerra, A.; D’Ambrosio, F. The Interaction between the Oral Microbiome and Systemic Diseases: A Narrative Review. Microbiol. Res. 2023, 14, 1862–1878. [Google Scholar] [CrossRef]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef]
- Brglez Mojzer, E.; Knez Hrnčič, M.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction Methods, Antioxidative Action, Bioavailability and Anticarcinogenic Effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef] [PubMed]
- Giordano-Kelhoffer, B.; Lorca, C.; March Llanes, J.; Rábano, A.; del Ser, T.; Serra, A.; Gallart-Palau, X. Oral Microbiota, Its Equilibrium and Implications in the Pathophysiology of Human Diseases: A Systematic Review. Biomedicines 2022, 10, 1803. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.L.; Torralba, M.; Leong, P.; Saffery, R.; Bockmann, M.; Kuelbs, C.; Singh, S.; Hughes, T.; Craig, J.M.; Nelson, K.E.; et al. Differential network analysis of oral microbiome metatranscriptomes identifies community scale metabolic restructuring in dental caries. PNAS Nexus 2022, 1, pgac239. [Google Scholar] [CrossRef] [PubMed]
- Varzakas, T.; Antoniadou, M. A Holistic Approach for Ethics and Sustainability in the Food Chain: The Gateway to Oral and Systemic Health. Foods 2024, 13, 1224. [Google Scholar] [CrossRef] [PubMed]
- Cugini, C.; Ramasubbu, N.; Tsiagbe, V.K.; Fine, D.H. Dysbiosis from a Microbial and Host Perspective Relative to Oral Health and Disease. Front. Microbiol. 2021, 12, 617485. [Google Scholar] [CrossRef]
- Kennedy, D.O.; Wightman, E.L. Herbal extracts and phytochemicals: Plant secondary metabolites and the enhancement of human brain function. Adv. Nutr. 2011, 2, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Chaachouay, N.; Zidane, L. Plant-Derived Natural Products: A Source for Drug Discovery and Development. Drugs Drug Candidates 2024, 3, 184–207. [Google Scholar] [CrossRef]
- Idir, F.; Van Ginneken, S.; Coppola, G.A.; Grenier, D.; Steenackers, H.P.; Bendali, F. Origanum vulgare ethanolic extracts as a promising source of compounds with antimicrobial, anti-biofilm, and anti-virulence activity against dental plaque bacteria. Front. Microbiol. 2022, 13, 999839. [Google Scholar] [CrossRef]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare 2023, 11, 1946. [Google Scholar] [CrossRef] [PubMed]
- Guefack, M.F.; Ngangoue, M.O.; Mbaveng, A.T.; Nayim, P.; Kuete, J.R.N.; Ngaffo, C.M.N.; Chi, G.F.; Ngameni, B.; Ngadjui, B.T.; Kuete, V. Antibacterial and antibiotic-potentiation activity of the constituents from aerial part of Donella welwitshii (Sapotaceae) against multidrug resistant phenotypes. BMC Complement. Med. Ther. 2022, 22, 194. [Google Scholar] [CrossRef] [PubMed]
- Fair, R.; Tor, Y. Antibiotics and Bacterial Resistance in the 21st Century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef]
- Aljeldah, M.M. Antimicrobial Resistance and Its Spread Is a Global Threat. Antibiotics 2022, 11, 1082. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Resistance Collaborators. The burden of bacterial antimicrobial resistance in the WHO African region in 2019: A cross-country systematic analysis. Lancet Glob. Health 2024, 12, e201–e216. [Google Scholar] [CrossRef]
- Luepke, K.H.; Suda, K.J.; Boucher, H.; Russo, R.L.; Bonney, M.W.; Hunt, T.D.; Mohr, J.F., 3rd. Past, Present, and Future of Antibacterial Economics: Increasing Bacterial Resistance, Limited Antibiotic Pipeline, and Societal Implications. Pharmacotherapy 2017, 37, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Nasim, N.; Sandeep, I.S.; Mohanty, S. Plant-derived natural products for drug discovery: Current approaches and prospects. Nucleus 2022, 65, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Jadimurthy, R.; Jagadish, S.; Nayak, S.C.; Kumar, S.; Mohan, C.D.; Rangappa, K.S. Phytochemicals as Invaluable Sources of Potent Antimicrobial Agents to Combat Antibiotic Resistance. Life 2023, 13, 948. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.W.; Lin, L.G.; Ye, W.C. Techniques for extraction and isolation of natural products: A comprehensive review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef]
- Cendrowski, A.; Kraśniewska, K.; Przybył, J.L.; Zielińska, A.; Kalisz, S. Antibacterial and Antioxidant Activity of Extracts from Rose Fruits (Rosa rugosa). Molecules 2020, 25, 1365. [Google Scholar] [CrossRef]
- Tiranakwit, T.; Puangpun, W.; Tamprasit, K.; Wichai, N.; Siriamornpun, S.; Srisongkram, T.; Weerapreeyakul, N. Phytochemical Screening on Phenolic, Flavonoid Contents, and Antioxidant Activities of Six Indigenous Plants Used in Traditional Thai Medicine. Int. J. Mol. Sci. 2023, 24, 13425. [Google Scholar] [CrossRef]
- Belew, A.A.; W/Hana, G.G.; Meshesha, D.S.; Akele, M.L. Evaluation of Total Phenolic, Total Flavonoid Content and Antioxidant Activity of Rhus Vulgaris. Res. Sq. 2021, PPR:PPR437116. [Google Scholar] [CrossRef]
- Mabotja, M.B.; Venter, S.L.; Du Plooy, C.P.; Kudanga, T.; Amoo, S.O. Phytochemical Content, Antioxidant, Alpha-Glucosidase Inhibitory and Antibacterial Activities of Spineless Cactus Pear Cultivars. Plants 2021, 10, 1312. [Google Scholar] [CrossRef] [PubMed]
- Saeed, N.; Khan, M.R.; Shabbir, M. Antioxidant activity, total phenolic and total flavonoid contents of whole plant extracts Torilis leptophylla L. BMC Complement. Altern. Med. 2012, 12, 221. [Google Scholar] [CrossRef] [PubMed]
- Al-Nawas, B.; Karbach, J. S3 Leitlinie: Odontogene Infektionen. Online Verfügbar Unter. 2017. Available online: https://wwwawmforg/uploads/tx_szleitlinien/007-006l_S3_Odontogene_Infektionen_2017-12pdf (accessed on 30 March 2024).
- Meinen, A.; Reuss, A.; Willrich, N.; Feig, M.; Noll, I.; Eckmanns, T.; Al-Nawas, B.; Markwart, R. Antimicrobial Resistance and the Spectrum of Pathogens in Dental and Oral-Maxillofacial Infections in Hospitals and Dental Practices in Germany. Front. Microbiol. 2021, 12, 676108. [Google Scholar] [CrossRef] [PubMed]
- Bhat, K.G.; Ingalagi, P.; Patil, S.; Patil, S.; Pattar, G. Antimicrobial susceptibility pattern of oral gram negative anaerobes from Indian subjects. Anaerobe 2021, 70, 102367. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests: Approved Standard—CLSI Document M02-A11; Clinical & Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: Approved Standard, 8th ed.; CLSI Document M07-A7; Clinical & Laboratory Standards Institute: Wayne, PA, USA, 2006. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- The European Committee on Antimicrobial Susceptibility Testing. Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth microdilution. Clin. Microbiol. Infect. 2003, 9, 1–7. [Google Scholar] [CrossRef]
- Kowalska-Krochmal, B.; Dudek-Wicher, R. The Minimum Inhibitory Concentration of Antibiotics: Methods, Interpretation, Clinical Relevance. Pathogens 2021, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Veloso, D.J.; Abrão, F.; Martins, C.H.G.; Bronzato, J.D.; Gomes, B.P.F.A.; Higino, J.S.; Sampaio, F.C. Potential antibacterial and anti-halitosis activity of medicinal plants against oral bacteria. Arch. Oral Biol. 2020, 110, 104585. [Google Scholar] [CrossRef]
- Clinical Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial a Susceptibility Tests for Bacteria That Grow Aerobically: Approved Standard, 8th ed.; CLSI document M0-A8; CLSI: Wayne, PA, USA, 2009. [Google Scholar]
- Bagheri, R.; Bohlouli, S.; Maleki Dizaj, S.; Shahi, S.; Memar, M.Y.; Salatin, S. The Antimicrobial and Anti-Biofilm Effects of Hypericum perforatum Oil on Common Pathogens of Periodontitis: An In Vitro Study. Clin. Pract. 2022, 12, 1009–1019. [Google Scholar] [CrossRef]
- Ersanli, C.; Tzora, A.; Skoufos, I.; Fotou, K.; Maloupa, E.; Grigoriadou, K.; Voidarou, C.C.; Zeugolis, D.I. The Assessment of Antimicrobial and Anti-Biofilm Activity of Essential Oils against Staphylococcus aureus Strains. Antibiotics 2023, 12, 384. [Google Scholar] [CrossRef] [PubMed]
- Samreen; Qais, F.A.; Ahmad, I. Anti-quorum sensing and biofilm inhibitory effect of some medicinal plants against gram-negative bacterial pathogens: In vitro and in silico investigations. Heliyon 2022, 8, e11113. [Google Scholar] [CrossRef] [PubMed]
- Adeyemo, R.O.; Famuyide, I.M.; Dzoyem, J.P.; Lyndy Joy, M. Anti-Biofilm, Antibacterial, and Anti-Quorum Sensing Activities of Selected South African Plants Traditionally Used to Treat Diarrhoea. Evid. Based Complement. Altern. Med. 2022, 2022, 1307801. [Google Scholar] [CrossRef] [PubMed]
- Souza-Melo, W.O.; Figueiredo-Júnior, E.C.; Freire, J.C.P.; Costa, B.P.; Lira, A.B.; Freires, I.A.; Cavalcanti, Y.W.; Lopes, W.S.; Tavares, J.F.; Pessôa, H.L.F.; et al. Phytochemistry, antifungal and antioxidant activity, and cytotoxicity of byrsonima gardneriana (A. Juss) extract. Arch. Oral Biol. 2021, 123, 104994. [Google Scholar] [CrossRef] [PubMed]
- Sæbø, I.P.; Bjørås, M.; Franzyk, H.; Helgesen, E.; Booth, J.A. Optimization of the Hemolysis Assay for the Assessment of Cytotoxicity. Int. J. Mol. Sci. 2023, 24, 2914. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Kempinski, C.; Chappell, J. Extraction and Analysis of Terpenes/Terpenoids. Curr. Protoc. Plant Biol. 2016, 1, 345–358. [Google Scholar] [CrossRef]
- Knez Hrnčič, M.; Cör, D.; Simonovska, J.; Knez, Ž.; Kavrakovski, Z.; Rafajlovska, V. Extraction Techniques and Analytical Methods for Characterization of Active Compounds in Origanum Species. Molecules 2020, 25, 4735. [Google Scholar] [CrossRef] [PubMed]
- Skoula, M.; Harborne, J.B. The taxonomy and chemistry of Origanum. In Oregan, the Genera Origanum and Lipia; Kintzios, S., Ed.; Medicinal and Aromatic Plants–Industrial Profiles; Taylor & Francis: London, UK, 2002; Volume 29, pp. 67–108. [Google Scholar]
- Lukas, B.; Schmiderer, C.; Novak, J. Phytochemical diversity of Origanum vulgare L. subsp. vulgare (Lamiaceae) from Austria. Biochem. Syst. Ecol. 2013, 50, 106–113. [Google Scholar] [CrossRef]
- Khan, M.; Khan, S.T.; Khan, N.A.; Mahmood, A.; AlKedhairy, A.A.; Alkhathlan, H.Z. The composition of the essential oil and aqueous distillate of Origanum vulgare L. growing in Saudi Arabia and evaluation of their antibacterial activity. Arab. J. Chem. 2018, 11, 1189–1200. [Google Scholar] [CrossRef]
- Lukas, B.; Schmiderer, C.; Novak, J. Essential oil diversity of European Origanum vulgare L. (Lamiaceae). Phytochemistry 2015, 119, 32–40. [Google Scholar] [CrossRef]
- Timothy, C.; Priya, V.; Gayathri, R.; Gayathri, R. Phytochemical analysis and total phenolic content of Origanum vulgare (oregano). Drug Invent. Today 2018, 10, 1903–1905. [Google Scholar]
- Mróz, M.; Kusznierewicz, B. Phytochemical screening and biological evaluation of Greek sage (Salvia fruticosa Mill.) extracts. Sci. Rep. 2023, 13, 22309. [Google Scholar] [CrossRef] [PubMed]
- Al-Kalaldeh, J.Z.; Abu-Dahab, R.; Aff, F.U. Volatile oil composition and antiproliferative activity of Laurus nobilis, Origanum syriacum, Origanum vulgare, and Salvia triloba against human breast adenocarcinoma cells. Nutr. Res. 2010, 30, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Torun, M.; Dincer, C.; Topuz, A.; Sahin-Nadeem HOzdemir, F. Aqueous extraction kinetics of soluble solids, phenolics and favonoids from sage (Salvia fruticosa Miller) leaves. J. Food Sci. Technol. 2015, 52, 2797–2805. [Google Scholar] [CrossRef] [PubMed]
- Bower, A.M.; Real Hernandez, L.M.; Berhow, M.A.; de Mejia, E.G. Bioactive Compounds from Culinary Herbs Inhibit a Molecular Target for Type 2 Diabetes Management, Dipeptidyl Peptidase IV. J. Agric. Food Chem. 2014, 62, 6147–6158. [Google Scholar] [CrossRef] [PubMed]
- Jafari Khorsand, G.; Morshedloo, M.R.; Mumivand, H.; Emami Bistgani, Z.; Maggi, F.; Khademi, A. Natural diversity in phenolic components and antioxidant properties of oregano (Origanum vulgare L.) accessions, grown under the same conditions. Sci. Rep. 2022, 12, 5813. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Azizi, A.; Janke, S.; Schwarz, M.; Zeller, S.; Honermeier, B. Antioxidant capacity variation in the oregano (Origanum vulgare L.) collection of the German National Genebank. Ind. Crops Prod. 2016, 92, 19–25. [Google Scholar] [CrossRef]
- Özüpek, B.; Pekacar, S.; Orhan, D.D. Evaluation of Phytochemical Contents and Biological Activities of Salvia officinalis and Salvia triloba Grown with Organic Farming FABAD. J. Pharm. Sci. Ank. 2023, 48, 125–138. [Google Scholar] [CrossRef]
- Lantzouraki, D.Z.; Tsiaka, T.; Soteriou, N.; Asimomiti, G.; Spanidi, E.; Natskoulis, P.; Gardikis, K.; Sinanoglou, V.J.; Zoumpoulakis, P. Antioxidant Profiles of Vitis vinifera L. and Salvia triloba L. Leaves Using High-Energy Extraction Methodologies. J. AOAC Int. 2020, 103, 413–421. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, H.; Wang, Z.; Lan, G.; Zhang, L. Comparative studies on antioxidant activities of extracts and fractions from the leaves and stem of Epimedium koreanum Nakai. J. Food Sci. Technol. 2013, 50, 1122–1129. [Google Scholar] [CrossRef]
- Indumathy, R.; Aruna, A. Free radical scavenging activities, total phenolic and flavonoid content of Lepidium sativum (Linn.). Int. J. Pharm. Pharm. Sci. 2013, 5, 634–637. [Google Scholar]
- Kakouri, E.; Trigas, P.; Daferera, D.; Skotti, E.; Tarantilis, P.A.; Kanakis, C. Chemical Characterization and Antioxidant Activity of Nine Hypericum Species from Greece. Antioxidants 2023, 12, 899. [Google Scholar] [CrossRef] [PubMed]
- Jan, S.; Rashid, M.; Abd Allah, E.F.; Ahmad, P. Biological Efficacy of Essential Oils and Plant Extracts of Cultivated and Wild E.cotypes of Origanum vulgare L. BioMed Res. Int. 2020, 2020, 8751718. [Google Scholar] [CrossRef] [PubMed]
- Parra, C.; Muñoz, P.; Bustos, L.; Parra, F.; Simirgiotis, M.J.; Escobar, H. UHPLC-DAD Characterization of Origanum vulgare L. from Atacama Desert Andean Region and Antioxidant, Antibacterial and Enzyme Inhibition Activities. Molecules 2021, 26, 2100. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I. Fe3+–Fe2+ Transformation Method: An Important Antioxidant Assay. Methods Mol. Biol. 2015, 1208, 233–246. [Google Scholar] [CrossRef]
- Hickl, J.; Argyropoulou, A.; Sakavitsi, M.E.; Halabalaki, M.; Al-Ahmad, A.; Hellwig, E. Mediterranean herb extracts inhibit microbial growth of representative oral microorganisms and biofilm formation of Streptococcus mutans. PLoS ONE 2018, 13, e0207574. [Google Scholar] [CrossRef]
- Popa, M.; Măruțescu, L.; Oprea, E.; Bleotu, C.; Kamerzan, C.; Chifiriuc, M.C.; Grădișteanu Pircalabioru, G. In Vitro Evaluation of the Antimicrobial and Immunomodulatory Activity of Culinary Herb Essential Oils as Potential Perioceutics. Antibiotics 2020, 9, 428. [Google Scholar] [CrossRef]
- Delamare, A.P.; Moschen-Pistorello, I.; Artico, L.; Atti-Serafini, L.; Echeverrigaray, S. Antibacterial activity of the essential oils of Salvia officinalis L. and Salvia triloba L. cultivated in South Brazil. Food Chem. 2007, 100, 603–608. [Google Scholar] [CrossRef]
- Eltawaty, S.; Busba, A.; Omare, M.-E.; Almagboul, A.; Ahmed, S.O.Y.; Elramli, A.; Elnour, A.; Dafalla, M. Antimicrobial Activity of Libyan salvia fruticosa Mil and multi-Drug Resistant Bacteria. Eur. J. Pharm. Med. Res. 2021, 8, 416–423. [Google Scholar]
- de Oliveira, J.R.; Vilela, P.G.D.F.; Almeida, R.B.A.; de Oliveira, F.E.; Carvalho, C.A.T.; Camargo, S.E.A.; Jorge, A.O.C.; de Oliveira, L.D. Antimicrobial activity of noncytotoxic concentrations of Salvia officinalis extract against bacterial and fungal species from the oral cavity. Gen. Dent. 2019, 67, 22–26. [Google Scholar]
- Amin, K.; Dannenfelser, R. In vitro hemolysis: Guidance for the pharmaceutical scientist. J. Pharm. Sci. 2006, 95, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Brewer, S. Natural Antioxidants: Sources, Compounds, Mechanisms of Action, and Potential Applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Saleh, N.A.M.; El-bary, R.B.E.-d.A.; Mpingirika, E.Z.; Essa, H.L.; El-Sayed, M.M.H.; Sherbetjian, M.S.; Elfandi, H.F.; Wahed, M.A.A.; Arafeh, R.; Amleh, A. Evaluating the Potential Anticancer Properties of Salvia triloba in Human-Osteosarcoma U2OS Cell Line and Ovarian Adenocarcinoma SKOV3 Cell Line. Appl. Sci. 2022, 12, 11545. [Google Scholar] [CrossRef]
- Daniels, G. The molecular genetics of blood group polymorphism. Transpl. Immunol. 2005, 14, 143–153. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Yang, X.; Li, C.; Song, Z. The Oral Microbiota: Community Composition, Influencing Factors, Pathogenesis, and Interventions. Front. Microbiol. 2022, 13, 895537. [Google Scholar] [CrossRef] [PubMed]
- Silbergleit, M.; Vasquez, A.A.; Miller, C.J.; Sun, J.; Kato, I. Oral and intestinal bacterial exotoxins: Potential linked to carcinogenesis. Prog. Mol. Biol. Transl. Sci. 2020, 171, 131–193. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.H.; Li, B.B.; Wang, B.; Zhao, J.; Zhang, X.Y.; Li, T.T.; Li, W.B.; Tang, D.; Qiu, M.J.; Wang, X.C.; et al. The role of Fusobacterium nucleatum in colorectal cancer: From carcinogenesis to clinical management. Chronic Dis. Transl. Med. 2019, 5, 178–187. [Google Scholar] [CrossRef]
- Lemos, J.A.; Palmer, S.R.; Zeng, L.; Wen, Z.T.; Kajfasz, J.K.; Freires, I.A.; Abranches, J.; Brady, L.J. The Biology of Streptococcus mutans. Microbiol. Spectr. 2019, 7, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto-Nakano, M. Role of Streptococcus mutans surface proteins for biofilm formation. Jpn. Dent. Sci. Rev. 2018, 54, 22–29. [Google Scholar] [CrossRef]
- Tanner, A.C.R.; Kressirer, C.A.; Rothmiller, S.; Johansson, I.; Chalmers, N.I. The caries microbiome: Implications for reversing dysbiosis. Adv. Dent. Res 2018, 29, 78–85. [Google Scholar] [CrossRef]
- Löwenmark, T.; Li, X.; Löfgren-Burström, A.; Zingmark, C.; Ling, A.; Kellgren, T.G.; Larsson, P.; Ljuslinder, I.; Wai, S.N.; Edin, S.; et al. Parvimonas micra is associated with tumour immune profiles in molecular subtypes of colorectal cancer. Cancer Immunol. Immunother. 2022, 71, 2565–2575. [Google Scholar] [CrossRef] [PubMed]
- Pignatelli, P.; Nuccio, F.; Piattelli, A.; Curia, M.C. The Role of Fusobacterium nucleatum in Oral and Colorectal Carcinogenesis. Microorganisms 2023, 11, 2358. [Google Scholar] [CrossRef] [PubMed]
- Kerdreux, M.; Edin, S.; Löwenmark, T.; Bronnec, V.; Löfgren-Burström, A.; Zingmark, C.; Ljuslinder, I.; Palmqvist, R.; Ling, A. Porphyromonas gingivalis in Colorectal Cancer and its Association to Patient Prognosis. J. Cancer 2023, 14, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Aladhadh, M. A Review of Modern Methods for the Detection of Foodborne Pathogens. Microorganisms 2023, 11, 1111. [Google Scholar] [CrossRef] [PubMed]
- Shakil, M.H.; Trisha, A.T.; Rahman, M.; Talukdar, S.; Kobun, R.; Huda, N.; Zzaman, W. Nitrites in Cured Meats, Health Risk Issues, Alternatives to Nitrites: A Review. Foods 2022, 11, 3355. [Google Scholar] [CrossRef] [PubMed]
- Redondo-Blanco, S.; Fernández, J.; López-Ibáñez, S.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Plant Phytochemicals in Food Preservation: Antifungal Bioactivity: A Review. J. Food Prot. 2020, 83, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, L.J.; Veronese, N.; Vernuccio, L.; Catanese, G.; Inzerillo, F.; Salemi, G.; Barbagallo, M. Nutrition, Physical Activity, and Other Lifestyle Factors in the Prevention of Cognitive Decline and Dementia. Nutrients 2021, 13, 4080. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.; Chow, Y.L. Exploring the potential of spice-derived phytochemicals as alternative antimicrobial agents. eFood 2023, 5, e126. [Google Scholar] [CrossRef]
- Teshome, S.; Jaleta, A.; Jemal, M.; Abamecha, A.; Gershe, S. Phytochemical screening and evaluation of antimicrobial activity of Pterolobium stellatum root extract. Pharm. Pharmacol. Int. J. 2023, 11, 151–156. Available online: https://medcraveonline.com/PPIJ/PPIJ-11-00413.pdf (accessed on 30 March 2024).
- Siddiqui, S.A.; Khan, S.; Mehdizadeh, M.; Bahmid, N.A.; Adli, D.N.; Walker, T.R.; Perestrelo, R.; Câmara, J.S. Phytochemicals and bioactive constituents in food packaging—A systematic review. Heliyon 2023, 9, e21196. [Google Scholar] [CrossRef]
- Bouarab Chibane, L.; Degraeve, P.; Ferhout, H.; Bouajila, J.; Oulahal, N. Plant antimicrobial polyphenols as potential natural food preservatives. J. Sci. Food Agric. 2019, 99, 1457–1474. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Singh, K.; Khedkar, R. Phytochemicals. In Functional and Preservative Properties of Phytochemicals; Academic Press: Cambridge, MA, USA, 2020; pp. 341–361. [Google Scholar] [CrossRef]
- Lombrea, A.; Antal, D.; Ardelean, F.; Avram, S.; Pavel, I.Z.; Vlaia, L.; Mut, A.M.; Diaconeasa, Z.; Dehelean, C.A.; Soica, C.; et al. A Recent Insight Regarding the Phytochemistry and Bioactivity of Origanum vulgare L. Essential Oil. Int. J. Mol. Sci. 2020, 21, 9653. [Google Scholar] [CrossRef] [PubMed]
- Prado, L.; Zalba, M.; Amato, D. Sustainability in Oral Health: The Role of Natural Products. Molecules 2020, 25, 2267. [Google Scholar]
- Souza, L.G.A.; Lopes, L.G.C.; Hill, M.P.; Harries, P.; Eggersdorfer, M.; Marušić, K. Sustainable Sourcing of Plant-Based Ingredients for Oral Health Care. Nutrients 2021, 13, 1002. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Source | Characterization Based on Gram Staining | Oxygen Requirements for Growth/ Growth Conditions Used in the Present Study |
|---|---|---|---|
| Streptococcus mutans | Oral cavity | Gram + | Facultatively anaerobic/Growth under anaerobic conditions |
| Streptococcus salivarius | Oral cavity | Gram + | Facultatively anaerobic/Growth under anaerobic conditions |
| Fusobacterium nucleatum | Oral cavity | Gram − | Obligate anaerobic/ Growth under anaerobic conditions |
| Porphyromonas gingivalis | Oral cavity | Gram − | Obligate anaerobic/ Growth under anaerobic conditions |
| Prevotella intermedia | Oral cavity | Gram − | Obligate anaerobic/ Growth under anaerobic conditions |
| Parvimonas micra | Oral cavity | Gram + | Obligate anaerobic/ Growth under anaerobic conditions |
| Staphylococcus aureus, Methicillin, and vancomycin-resistant | Dental caries area | Gram + | Best in the presence of oxygen-rich environment/ Growth under aerobic conditions |
| Staphylococcus aureus, Methicillin-Resistant | Raw milk | Gram + | Best in the presence of oxygen-rich environment/ Growth under aerobic conditions |
| Staphylococcus aureus, Methicillin-Resistant | Poultry meat | Gram + | Best in the presence of oxygen-rich environment/ Growth under aerobic conditions |
| Staphylococcus aureus subsp. aureus | Reference strain ATCC 6538 | ||
| Phytochemical Compounds | Plant Extracts | |||||||
|---|---|---|---|---|---|---|---|---|
| Origanum vulgare | Salvia triloba | |||||||
| A | Ε40 | Ε60 | ΕΝΖ | A | Ε40 | Ε60 | ΕΝΖ | |
| Alkaloids | + | - | - | + | + | - | - | - |
| Anthraquinones | - | - | - | + | + | - | - | - |
| Terpenoids (Salkowski’s test) | + | + | + | + | + | + | + | + |
| Steroids | + | - | - | + | - | - | - | - |
| Saponins | - | + | + | - | + | + | - | - |
| Flavonoids (alkaline reagent test) | + | + | + | + | + | + | + | + |
| Tannins (ferric chloride test) | + | + | + | - | + | + | + | + |
| Glycosides (Keller–Kiliani test) | + | + | + | + | + | + | + | - |
| Plant Extracts | Concentration (μg/mL) | |||||
|---|---|---|---|---|---|---|
| 1 | 10 | 50 | 100 | 300 | 500 | |
| Oregano A | 3.42 ± 0.33 bc | 10.57 ± 0.13 c | 33.34 ± 0.58 b | 38.55 ± 0.21 b | 52.24 ± 0.22 a | 59.32 ± 0.85 a |
| Oregano A* | 5.28 ± 0.1 d | 12.18 ± 0.42 d | 40.86 ± 0.02 e | 45.26 ± 0.53 c | 59.52 ± 0.4 b | 62.92 ± 0.35 b |
| Oregano E40 | 4.12 ± 0.11 cf | 12.19 ± 0.23 d | 50.46 ± 0.34 g | 70.48 ± 0.07 g | 74.5 ± 0.39 e | 77.33 ± 0.11 d |
| Oregano E60 | 12.37 ± 0.28 g | 18.06 ± 0.19 f | 55.24 ± 0.21 h | 76.79 ± 0.22 h | 80.7 ± 0.17 f | 81.36 ± 0.25 e |
| Oregano ENZ | 11.6 ± 0.45 ef | 14.32 ± 0.34 e | 57.99 ± 0.22 i | 78.77 ± 0.17 i | 82.44 ± 0.42 g | 86.43 ± 0.42 g |
| Salvia A | 11.5 ± 0.39 e | 14.12 ± 0.12 e | 39.53 ± 0.45 d | 60.53 ± 0.33 f | 82.21 ± 0.32 g | 84.24 ± 0.29 f |
| Salvia A* | 3.42 ± 0.16 bc | 9.36 ± 0.27 b | 32.38 ± 0.44 b | 48.43 ± 0.34 d | 80.13 ± 0.29 f | 82.38 ± 0.44 e |
| Salvia E40 | 2.63 ± 0.2 b | 10.22 ± 0.19 c | 35.27 ± 0.22 c | 48.36 ± 0.39 d | 72.37 ± 0.26 d | 74.49 ± 0.42 c |
| Salvia E60 | 1.37 ± 0.41 a | 6.33 ± 0.25 a | 22.38 ± 0.41 a | 30.31 ± 0.23 a | 54.97 ± 0.44 b | 60.53 ± 0.41 a |
| Salvia ENZ | 12.71 ± 0.24 g | 17.37 ± 0.42 f | 42.73 ± 0.25 f | 59.25 ± 0.18 e | 85.46 ± 0.21 h | 87.14 ± 0.71 g |
| Plant Extracts | Concentration (μg/mL) | |||||
|---|---|---|---|---|---|---|
| 2.5 | 5 | 10 | 15 | 20 | 25 | |
| Oregano A | 0.15 ± 0 a | 0.21 ± 0 a | 0.31 ± 0 a | 0.57 ± 0 a | 0.77 ± 0 a | 0.85 ± 0 a |
| Oregano A* | 0.21 ± 0 c | 0.40 ± 0 c | 0.55 ± 0 f | 0.81 ± 0 c | 1.01 ± 0 c | 1.21 ± 0 d |
| Oregano E40 | 0.22 ± 0 d | 0.49 ± 0 g | 0.75 ± 0 h | 1.28 ± 0 h | 1.44 ± 0 g | 1.62 ± 0 f |
| Oregano E60 | 0.24 ± 0 c | 0.55 ± 0 h | 0.83 ± 0 i | 1.34 ± 0 i | 1.55 ± 0 h | 1.77 ± 0 g |
| Oregano ENZ | 0.23 ± 0 c | 0.58 ± 0 i | 0.99 ± 0 j | 1.48 ± 0 j | 1.65 ± 0 i | 1.93 ± 0 i |
| Salvia A | 0.24 ± 0 c | 0.37 ± 0 d | 0.44 ± 0 bc | 0.78 ± 0 d | 0.98 ± 0 d | 1.33 ± 0 c |
| Salvia A* | 0.18 ± 0 b | 0.32 ± 0 c | 0.54 ± 0 f | 0.71 ± 0 c | 0.86 ± 0 bc | 1.11 ± 0 c |
| Salvia E40 | 0.19 ± 0 b | 0.31 ± 0 c | 0.48 ± 0 d | 0.68 ± 0 b | 0.85 ± 0 b | 1.09 ± 0 b |
| Salvia E60 | 0.18 ± 0 b | 0.28 ± 0 b | 0.38 ± 0 b | 0.70 ± 0 c | 0.85 ± 0 b | 1.08 ± 0 b |
| Salvia ENZ | 0.26 ± 0 f | 0.38 ± 0 d | 0.51 ± 0 c | 0.88 ± 0 f | 1.31 ± 0 f | 1.76 ± 0 g |
| Reference | ||||||
| Gallic acid (GA) | 0.18 ± 0.0 b | 0.43 ± 0 f | 1.81 ± 0 k | 1.85 ± 0 k | 1.88 ± 0 j | 1.88 ± 0 h |
| Ascorbic acid (AA) | 0.19 ± 0.0 b | 0.42 ± 0 f | 0.68 ± 0 g | 1.12 ± 0 g | 1.65 ± 0 i | 1.88 ± 0 h |
| Bacterial Strain | Antimicrobial Agents | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amoxicillin | Amoxicillin/ Clavulanic Acid | Vancomycin | Imipenem | Erythromycin | Clindamycin | Gentamicin | Tetracycline | Ciprofloxacin | Metronidazole | Cefuroxime | Cefotaxime | |||||||||||||
| mm | MIC mg/mL | mm | MIC mg/mL | mm | MIC mg/mL | mm | MIC mg/mL | mm | MIC mg/mL | mm | MIC mg/mL | mm | MIC mg/mL | mm | MIC mg/mL | mm | MIC mg/mL | mm | MIC mg/mL | mm | MIC mg/mL | mm | MIC mg/mL | |
| Staphylococcus aureus subsp. aureus, methicillin, and vancomycin-resistant | 0 | 0.128 | 0 | 0.0128 | 0 | 0.064 | 10 | 0.032 | 11 | 0.032 | 24 | 0.00025 | 23 | 0.0005 | 24 | 0.001 | 52 | 0.00125 | - * | - | 10 | 0.064 | 10 | 0.064 |
| Methicillin-Resistant S. aureus, from raw milk | 0 | 0.064 | 0 | 0.064 | 18 | 0.002 | 14 | 0.004 | 26 | 0.0005 | 26 | 0.00025 | 26 | 0.0005 | 28 | 0.0005 | 38 | 0.00125 | - | - | 22 | 0.002 | 24 | 0.006 |
| Methicillin-Resistant S. aureus, from raw poultry | 0 | 0.128 | 0 | 0.128 | 0 | 0.032 | 11 | 0.032 | 18 | 0.008 | 11 | 0.008 | 20 | 0.001 | 22 | 0.001 | 33 | 0.00125 | - | - | 10 | 0.032 | 8 | 0.032 |
| Streptococcus mutans | 37 | 0.00025 | 35 | 0.000125 | 21 | 0.0005 | 35 | 0.0005 | 11 | 0.004 | 27 | 0.000125 | 22 | 0.024 | 13 | 0.008 | 21 | 0.0005 | - | - | 30 | 0.00025 | 31 | 0.0005 |
| Streptococcus salivarius | 36 | 0.000125 | 34 | 0.000125 | 22 | 0.0005 | 29 | 0.001 | 10 | 0.016 | 29 | 0.000125 | 0 | 0.096 | 11 | 0.010 | 18 | 0.001 | - | - | 28 | 0.0005 | 30 | 0.0005 |
| Fusobacterium nucleatum | 26 | 0.00025 | 24 | 0.004/0.002 | 25 | 0.00025 | 25 | 0.00025 | 0 | 0.064 | 10 | 0.016 | 0 | 0.128 | 0 | 0.032 | 22 | 0.001 | 21 | 0.001 | 27 | 0.004 | 29 | 0.004 |
| Porphyromonas gingivalis | 28 | 0.00025 | 26 | 0.004/0.002 | 27 | 0.00025 | 28 | 0.00025 | 0 | 0.064 | 20 | 0.001 | 0 | 0.128 | 0 | 0.032 | 8 | 0.008 | 27 | 0.0005 | 27 | 0.002 | 27 | 0.004 |
| Prevotella intermedia | 9 | 0.016 | 24 | 0.004/0.002 | 25 | 0.00025 | 25 | 0.00025 | 0 | 0.064 | 8 | 0.032 | 0 | 0.128 | 0 | 0.032 | 0 | 0.016 | 26 | 0.0005 | 25 | 0.002 | 26 | 0.002 |
| Parvimonas micra | 27 | 0.0005 | 27 | 0.002/0.001 | 20 | 0.0025 | 27 | 0.00015 | 21 | 0.004 | 0 | 0.032 | 0 | 0.256 | 0 | 0.032 | 25 | 0.0005 | 22 | 0.0005 | 27 | 0.00025 | 27 | 0.0005 |
| Pathogen | Disk Content (%) | A (mm) | E40 (mm) | E60 (mm) | Enz (mm) |
|---|---|---|---|---|---|
| S. aureus MRSA/VRSA | 10 | 6 ± 0.1 a | 7.5 ± 0.5 a | 9.7 ± 0.6 ab | 11.5 ± 0.4 d |
| 20 | 13.8 ± 0.8 ef | 10 ± 0.6 ab | 14.7 ± 0.5 b | 15 ± 0.5 e | |
| 50 | 15.7 ± 0.5 ab | 16.2 ± 0.6 b | 18.3 ± 0.7 a | 19.1 ± 1.1 b | |
| 100 | 18.1 ± 0.3 ab | 19.3 ± 0.6 bc | 21 ± 0.1 b | 22.4 ± 0.1 b | |
| MRSA (raw milk) | 10 | 6.5 ± 0.3 a | 8.4 ± 0.3 a | 10.8 ± 0.6 bc | 8.7 ± 0.5 ab |
| 20 | 10.8 ± 0.3 bc | 10.4 ± 0.3 ab | 15.1 ± 0.4 b | 15.3 ± 0.9 e | |
| 50 | 9.7 ± 6.8 a | 14.1 ± 0.1 a | 17.6 ± 0.5 a | 19 ± 0.9 b | |
| 100 | 18.2 ± 0.4 ab | 17.5 ± 0.5 a | 18.8 ± 0.6 a | 20.4 ± 0.4 ab | |
| MRSA (raw poultry) | 10 | 6.9 ± 0.1 a | 9.8 ± 0.6 bc | 12.2 ± 0.2 d | 8 ± 0.5 a |
| 20 | 11.5 ± 0.4 de | 13.2 ± 0.6 de | 18.5 ± 0.3 c | 15.1 ± 0.4 e | |
| 50 | 22.9 ± 5.2 b | 19.5 ± 0.3 d | 27.8 ± 0.9 d | 18.7 ± 0.5 b | |
| 100 | 19.2 ± 0.4 cd | 26.5 ± 0.6 d | 38.7 ± 0.8 f | 22.1 ± 0.5 b | |
| S. mutans | 10 | 9.6 ± 0.7 c | 8.1 ± 0.3 a | 12.1 ± 0.8 c | 9.5 ± 0.4 b |
| 20 | 12.6 ± 0.4 de | 11.2 ± 0.6 ab | 19.3 ± 0.5 de | 12.4 ± 0.2 bc | |
| 50 | 17.1 ± 0.4 ab | 17.3 ± 0.6 cd | 28.6 ± 0.9 d | 18.8 ± 0.3 b | |
| 100 | 19.8 ± 0.5 cd | 19.8 ± 0.4 bc | 31.4 ± 0.6 e | 22.3 ± 2.7 b | |
| S. salivarius | 10 | 6.2 ± 0.2 a | 8.1 ± 0.2 a | 10.7 ± 0.5 b | 7.7 ± 0.6 a |
| 20 | 9.8 ± 0.3 ab | 11.8 ± 0.3 bcd | 18.5 ± 0.7 cd | 11.5 ± 0.5 ab | |
| 50 | 15.2 ± 0.4 ab | 15.9 ± 0.4 b | 25 ± 0.4 c | 17 ± 0.1 b | |
| 100 | 19 ± 0.3 cd | 20.7 ± 1.3 c | 28.2 ± 0.5 c | 19.6 ± 0.5 ab | |
| P. gingivalis | 10 | 8.6 ± 0.5 bc | 10.7 ± 0.5 cd | 12.3 ± 0.4 d | 9.7 ± 0.5 bc |
| 20 | 13.5 ± 0.4 ef | 14.8 ± 0.1 e | 19.5 ± 0.6 de | 14.1 ± 0.7 de | |
| 50 | 18.2 ± 0.5 b | 19.5 ± 0.3 d | 29 ± 0.3 d | 19 ± 0.3 b | |
| 100 | 20.6 ± 0.7 d | 19.9 ± 0.1 bc | 31.1 ± 0.2 e | 21.8 ± 0.4 b | |
| F. nucleatum | 10 | 6.1 ± 0.2 a | 7.9 ± 0.2 a | 13.3 ± 0.2 d | 10.1 ± 0.2 c |
| 20 | 8.9 ± 0.6 a | 14.8 ± 0.2 e | 19.1 ± 0.2 de | 13.8 ± 0.5 cd | |
| 50 | 15.3 ± 0.5 ab | 18.1 ± 0.7 cd | 28.2 ± 0.5 d | 18.7 ± 0.7 b | |
| 100 | 17.8 ± 0.5 ab | 20.2 ± 0.3 c | 32.5 ± 1.5 ef | 21.9 ± 0.3 b | |
| P. intermedia | 10 | 6.1 ± 0.1 a | 8.6 ± 0.4 ab | 10.6 ± 0.2 b | 8.4 ± 0.7 a |
| 20 | 12.5 ± 0.4 ef | 12.4 ± 0.5 cd | 17.2 ± 0.5 c | 11.2 ± 0.8 a | |
| 50 | 16.8 ± 0.5 ab | 17.1 ± 0.3 bc | 21.9 ± 1.8 b | 18.1 ± 0.2 b | |
| 100 | 18.8 ± 0.3 abc | 19.1 ± 0 bc | 29.3 ± 0.6 b | 21.9 ± 0.7 b | |
| P. micra | 10 | 6.3 ± 0.6 a | 11.7 ± 0.5 d | 12.6 ± 0.6 d | 9.6 ± 0.5 b |
| 20 | 14.8 ± 0.4 f | 18.2 ± 0.4 f | 20 ± 0.2 e | 13.3 ± 0.2 cd | |
| 50 | 16.9 ± 0.4 ab | 24.2 ± 1.1 e | 27.9 ± 0.4 d | 18 ± 0.2 b | |
| 100 | 18.2 ± 0.2 ab | 31.4 ± 0.4 e | 31.1 ± 0.1 e | 20.6 ± 0.4 ab | |
| S. aureus ATCC 12600 (Ref) | 10 | 8.3 ± 0.5 b | 8.1 ± 0.6 a | 9.1 ± 0.1 a | 10.2 ± 0.3 c |
| 20 | 12.8 ± 0.5 de | 12.5 ± 1.3 c | 12.3 ± 0.6 a | 13 ± 0.3 bc | |
| 50 | 16.6 ± 0.4 ab | 17.4 ± 0.6 bc | 18 ± 0.3 a | 17.8 ± 0.4 ab | |
| 100 | 17.6 ± 0.5 a | 19.1 ± 0.2 bc | 18.7 ± 0.5 a | 19.2 ± 0.4 a |
| Pathogen | Disk Content (%) | A (mm) | E40 (mm) | E60 (mm) | Enz (mm) |
|---|---|---|---|---|---|
| S. aureus MRSA/VRSA | 10 | 6.7 ± 0.6 ab | 10 ± 0.7 cd | 10.5 ± 0.5 abc | 6 ± 0 a |
| 20 | 11.3 ± 0.4 bc | 12.6 ± 0.2 cd | 15 ± 0.4 bc | 8.5 ± 0.6 a | |
| 50 | 13.4 ± 0.4 ac | 17.5 ± 0.5 cd | 18.9 ± 0.5 bc | 12.2 ± 0.3 a | |
| 100 | 17.6 ± 0.3 abc | 18.5 ± 0.3 bc | 22.8 ± 2 ab | 17.9 ± 0.4 a | |
| MRSA (raw milk) | 10 | 7.6 ± 0.6 ab | 12.5 ± 0.4 e | 12.6 ± 0.3 d | 7.4 ± 0.5 b |
| 20 | 11.9 ± 0.7 bc | 15 ± 0.2 e | 18.6 ± 0.3 ef | 12 ± 0.4 c | |
| 50 | 14.8 ± 0.5 bc | 18 ± 0.3 d | 23.2 ± 0.6 e | 14.6 ± 0.2 b | |
| 100 | 17.6 ± 0.3 abc | 24.3 ± 0.4 e | 28.7 ± 0.6 c | 17.4 ± 0.4 a | |
| MRSA (raw poultry) | 10 | 6.4 ± 0.4 a | 9.1 ± 0.7 ab | 11.4 ± 0.7 bcd | 7.8 ± 0.5 bc |
| 20 | 10.6 ± 0.1 bc | 10.6 ± 0.2 a | 16.2 ± 0.6 cd | 10.7 ± 0.4 b | |
| 50 | 14.9 ± 0.2 bc | 16.2 ± 0.3 bc | 20.6 ± 0.5 cd | 15.2 ± 0.4 bc | |
| 100 | 17.4 ± 0.4 bc | 18.3 ± 0.6 abc | 29.1 ± 0.8 c | 17.7 ± 0.3 a | |
| S. mutans | 10 | 10.8 ± 0.8 de | 8.6 ± 0.3 ab | 9.8 ± 0.4 a | 12.1 ± 0.5 e |
| 20 | 16.4 ± 0.4 f | 10.7 ± 0.3 ab | 11.9 ± 0.5 a | 17.1 ± 0.2 cd | |
| 50 | 20.7 ± 0.8 g | 15.2 ± 0.3 ab | 16.7 ± 0.5 ab | 21.5 ± 0.7 e | |
| 100 | 29.3 ± 0.9 f | 16.7 ± 0.2 a | 18.5 ± 0.5 a | 30.8 ± 0.6 b | |
| S. salivarius | 10 | 8.4 ± 0.8 bc | 10.8 ± 0.6 de | 11.7 ± 0.4 cd | 9.3 ± 0.3 d |
| 20 | 12.9 ± 0.6 de | 13.1 ± 0.8 d | 17.4 ± 0.5 de | 12.8 ± 0.5 b | |
| 50 | 16.4 ± 0.5 de | 16.3 ± 0.5 bc | 22.2 ± 0.7 de | 17.8 ± 0.3 d | |
| 100 | 18.5 ± 0.6 cd | 18.5 ± 0.5 bc | 27.5 ± 5.5 bc | 18.5 ± 0.4 a | |
| P. gingivalis | 10 | 11.2 ± 0.8 e | 7.6 ± 0.5 a | 10.2 ± 0.2 ab | 12.2 ± 0.6 e |
| 20 | 15.2 ± 0.5 f | 11.9 ± 0.3 c | 14 ± 0.7 b | 18.2 ± 0.3 de | |
| 50 | 20.6 ± 0.5 fg | 14.1 ± 0.3 a | 17 ± 0.2 ab | 28.1 ± 1 g | |
| 100 | 32.3 ± 1.9 g | 17.5 ± 0.3 ab | 18.6 ± 0.4 a | 33.8 ± 1.2 cd | |
| F. nucleatum | 10 | 6.1 ± 0.1 a | 9.8 ± 0.4 bcd | 11.9 ± 0.6 cd | 9.8 ± 0.4 d |
| 20 | 9.3 ± 0.6 a | 10.4 ± 0.4 a | 17.2 ± 0.2 de | 16.7 ± 0.4 c | |
| 50 | 12.4 ± 0.6 a | 13.9 ± 0.2 a | 24.1 ± 1.4 e | 19.6 ± 0.8 d | |
| 100 | 16 ± 0.2 b | 16.8 ± 0.2 a | 28.9 ± 0.9 c | 30.3 ± 0.8 b | |
| P. intermedia | 10 | 9.6 ± 0.2 cde | 8.3 ± 0.6 ab | 11.7 ± 0.5 cd | 9 ± 0.1 cd |
| 20 | 13.7 ± 0.2 e | 12 ± 0.4 c | 19 ± 0.4 f | 12.5 ± 0.7 b | |
| 50 | 19 ± 0.2 ef | 17.3 ± 0.4 cd | 28.4 ± 0.6 f | 16.7 ± 0.4 cd | |
| 100 | 24.5 ± 0.5 e | 19.1 ± 0.8 c | 31.7 ± 0.6 c | 17.9 ± 0.1 a | |
| P. micra | 10 | 9.5 ± 0.6 cd | 10.6 ± 0.6 cd | 11 ± 0.5 abc | 12 ± 0.5 e |
| 20 | 12.4 ± 0.4 de | 15.3 ± 0.5 e | 16.2 ± 0.3 cd | 19.3 ± 0.5 e | |
| 50 | 17.8 ± 0.9 de | 21.6 ± 1.2 e | 20.8 ± 0.5 de | 29.3 ± 0.5 f | |
| 100 | 21.2 ± 0.9 d | 27.8 ± 1.1 f | 30.9 ± 0.3 c | 35.4 ± 1.1 d | |
| S. aureus ATCC 12600 (Ref) | 10 | 9.4 ± 0.5 cd | 10.9 ± 1.1 de | 10.5 ± 0.6 abc | 11.1 ± 0.1 e |
| 20 | 12.9 ± 0.5 de | 14.9 ± 0.1 e | 18.1 ± 0.6 ef | 18 ± 0.2 de | |
| 50 | 17.5 ± 0.6 de | 18.3 ± 1 d | 26.7 ± 0.7 f | 22.2 ± 1 e | |
| 100 | 19.8 ± 0.4 cd | 21 ± 0.4 d | 30.7 ± 0.4 c | 31.8 ± 2 bc |
| Minimum Inhibitory Concentration (mg/mL) | Minimum Bactericidal Concentration (mg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| Pathogen | A | E40 | E60 | Enz | A | E40 | E60 | Enz |
| S. aureus MRSA/VRSA | 6.3 ± 0 b | 3.1 ± 0 b | 1.6 ± 0 b | 1.6 ± 0 b | 12.5 ± 0 b | 6.3 ± 0 c | 3.1 ± 0 b | 1.6 ± 0 a |
| MRSA (raw milk) | 6.3 ± 0 b | 6.3 ± 0 | 3.1 ± 0 c | 3.1 ± 0 c | 12.5 ± 0 b | 12.5 ± 0 | 12.5 ± 0 d | 6.3 ± 0 c |
| MRSA (raw poultry) | 3.1 ± 0 a | 0.8 ± 0 a | 0.4 ± 0 a | 0.8 ± 0 a | 6.3 ± 0 a | 1.6 ± 0 a | 1.6 ± 0 a | 1.6 ± 0 a |
| S. mutans | 3.1 ± 0 b | 3.1 ± 0 b | 1.6 ± 0 b | 3.1 ± 0 c | 6.3 ± 0 a | 12.5 ± 0 d | 3.1 ± 0 b | 6.3 ± 0 c |
| S. salivarius | 6.3 ± 0 b | 3.1 ± 0 b | 1.6 ± 0 b | 3.1 ± 0 c | 12.5 ± 0 b | 6.3 ± 0 c | 6.3 ± 0 c | 6.3 ± 0 c |
| P. gingivalis | 6.3 ± 0 b | 6.3 ± 0 c | 1.6 ± 0 b | 0.8 ± 0 a | 25 ± 0 c | 12.5 ± 0 d | 3.1 ± 0 b | 6.3 ± 0 c |
| F. nucleatum | 6.3 ± 0 b | 3.1 ± 0 b | 0.8 ± 0 a | 3.1 ± 0 c | 25 ± 0 c | 12.5 ± 0 d | 3.1 ± 0 b | 3.1 ± 0 b |
| P. intermedia | 6.3 ± 0 b | 3.1 ± 0 b | 0.8 ± 0 a | 3.1 ± 0 c | 25 ± 0 c | 12.5 ± 0 d | 6.3 ± 0 c | 12.5 ± 0 d |
| P. micra | 12.5 ± 0 c | 0.8 ± 0 a | 0.8 ± 0 a | 1.6 ± 0 b | 25 ± 0 c | 3.1 ± 0 b | 6.3 ± 0 c | 6.3 ± 0 c |
| S. aureus ATCC 12600 (Ref) | 6.3 ± 0 b | 3.1 ± 0 b | 3.1 ± 0 c | 1.6 ± 0 b | 12.5 ± 0 b | 12.5 ± 0 d | 12.5 ± 0 d | 6.3 ± 0 c |
| Minimum Inhibitory Concentration (mg/mL) | Minimum Bactericidal Concentration (mg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| Pathogen | A | E40 | E60 | Enz | A | E40 | E60 | Enz |
| S. aureus MRSA/VRSA | 12.5 ± 0 e | 3.1 ± 0 b | 1.6 ± 0 b | 6.3 ± 0 d | 50 ± 0 d | 12.5 ± 0 c | 6.3 ± 0 d | 25 ± 0 f |
| MRSA (raw milk) | 6.3 ± 0 d | 3.1 ± 0 b | 1.6 ± 0 b | 6.3 ± 0 d | 12.7 ± 0.3 b | 12.5 ± 0 c | 3.1 ± 0 c | 25 ± 0 f |
| MRSA (raw poultry) | 12.5 ± 0 e | 6.3 ± 0 c | 0.8 ± 0 a | 3.1 ± 0 c | 50 ± 0 d | 12.5 ± 0 c | 3.1 ± 0 c | 12.5 ± 0 e |
| S. mutans | 0.8 ± 0 a | 6.3 ± 0 c | 3.1 ± 0 c | 0.8 ± 0 a | 6.3 ± 0 a | 25 ± 0 d | 12.5 ± 0 d | 3.1 ± 0 c |
| S. salivarius | 3.1 ± 0 c | 3.1 ± 0 b | 0.8 ± 0 a | 6.3 ± 0 d | 12.7 ± 0.3 b | 6.3 ± 0 b | 1.6 ± 0 b | 12.5 ± 0 |
| P. gingivalis | 0.8 ± 0 a | 6.3 ± 0 c | 6.3 ± 0 d | 0.8 ± 0 a | 6.3 ± 0 a | 25 ± 0 d | 12.5 ± 0 d | 1.6 ± 0 b |
| F. nucleatum | 12.5 ± 0 | 6.3 ± 0 c | 1.6 ± 0 b | 1.6 ± 0 b | 50 ± 0 d | 12.5 ± 0 c | 3.1 ± 0 c | 1.6 ± 0 b |
| P. intermedia | 1.6 ± 0 b | 3.1 ± 0 b | 0.8 ± 0 a | 3.1 ± 0 c | 12.7 ± 0.3 b | 12.5 ± 0 c | 0.8 ± 0 a | 6.3 ± 0 d |
| P. micra | 1.6 ± 0 b | 1.6 ± 0 a | 0.8 ± 0 a | 0.4 ± 0 a | 6.3 ± 0 a | 3.1 ± 0 a | 0.8 ± 0 a | 1.6 ± 0 b |
| S. aureus ATCC 12600 (Ref) | 6.3 ± 0 d | 1.6 ± 0 a | 0.8 ± 0 a | 0.7 ± 0.2 a | 25 ± 0 c | 3.1 ± 0 a | 3.1 ± 0 c | 0.8 ± 0 a |
| Concentration of Each Plant Extract (μg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 12.5 | 25 | 50 | 100 | 150 | 200 | 250 | 500 | 1000 | |
| Oregano A | 4.35 ± 1 1de | 4.21 ± 0.6 1bc | 4.46 ± 0.7 1cde | 5.45 ± 0.31bc | 7.57 ± 0.32c | 8.45 ± 0.1 23c | 8.64 ± 0.1 23bcd | 9.37 ± 0.2 34ab | 10.1 ± 0.5 4ab |
| Oregano A * | 2.74 ± 0.5 1bc | 3.81 ± 0.7 1bc | 4.54 ± 0.6 12de | 6.04 ± 0.72bcd | 9.04 ± 0.73d | 10.43 ± 0.5 34c | 11.34 ± 0.7 4cde | 19.9 ± 0.9 5c | 21.62 ± 0.4 5cd |
| Oregano E40 | 3.55 ± 0.1 1cd | 4.1 ± 0.9 12bc | 4.17 ± 0.9 12cd | 7.03 ± 1.412cd | 9.07 ± 0.712d | 14.42 ± 3.3 23d | 21.51 ± 1.6 3f | 34.39 ± 2.6 4d | 54.92 ± 9.7 5e |
| Oregano E60 | 4.94 ± 0.6 1de | 6.07 ± 0.4 1d | 6.62 ± 0.5 1f | 9.04 ± 1.1 1a | 10.68 ± 0.3 1e | 22.14 ± 1.2 2ce | 29.09 ± 1 3g | 31.76 ± 0.5 3d | 82.58 ± 5.9 4f |
| Oregano ENZ | 0.55 ± 0.4 1a | 1.43 ± 0.5 1a | 1.94 ± 0.1 1ab | 4.5 ± 0.1 12b | 7.24 ± 0.2 2c | 8.01 ± 0.5 2 | 14.38 ± 4.1 3e | 29.42 ± 0.2 4d | 31.1 ± 0.5 4 |
| Salvia A | 0.88 ± 0.2 1ab | 1.32 ± 0.1 12a | 1.9 ± 0.1 23ab | 2.49 ± 0.2 3a | 2.89 ± 0.3 4a | 4.06 ± 0.4 5ab | 4.76 ± 0.6 5ab | 6.29 ± 0.3 6a | 7.46 ± 0.3 7a |
| Salvia A* | 1.39 ± 0.4 1a | 1.61 ± 0.3 1a | 3.04 ± 0.7 2bc | 4.46 ± 0.3 3b | 5.67 ± 0.3 4b | 7.1 ± 0.5 5bc | 8.23 ± 0.5 6abc | 9.04 ± 0.1 67a | 9.59 ± 0.1 7ab |
| Salvia E40 | 2.6 ± 0.3 1bc | 3.33 ± 0.1 12b | 3.81 ± 0.2 12cd | 4.9 ± 0.4 12b | 5.74 ± 0.4 123b | 7.24 ± 0.2 123bc | 8.6 ± 0.2 23bcd | 6.41 ± 5.6 3a | 10.76 ± 0.2 3ab |
| Salvia E60 | 3.77 ± 0.3 1cde | 4.87 ± 0.4 12cd | 5.63 ± 0.2 2ef | 7.32 ± 0.1 34de | 8.12 ± 0.8 4cd | 10.06 ± 1 5c | 12.99 ± 0.9 6e | 14.89 ± 0.4 7bc | 19.72 ± 0.8 8bc |
| Salvia ENZ | 0.18 ± 0.3 1a | 0.8 ± 0.1 12a | 0.91 ± 0.1 12a | 1.24 ± 0.1 2a | 2.56 ± 0.5 3a | 3.11 ± 0.2 34a | 3.92 ± 0.3 4a | 5.89 ± 0.6 5a | 7.98 ± 0.7 6a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antoniadou, M.; Rozos, G.; Vaou, N.; Zaralis, K.; Ersanli, C.; Alexopoulos, A.; Dadamogia, A.; Varzakas, T.; Tzora, A.; Voidarou, C. Comprehensive Bio-Screening of Phytochemistry and Biological Capacity of Oregano (Origanum vulgare) and Salvia triloba Extracts against Oral Cariogenic and Food-Origin Pathogenic Bacteria. Biomolecules 2024, 14, 619. https://doi.org/10.3390/biom14060619
Antoniadou M, Rozos G, Vaou N, Zaralis K, Ersanli C, Alexopoulos A, Dadamogia A, Varzakas T, Tzora A, Voidarou C. Comprehensive Bio-Screening of Phytochemistry and Biological Capacity of Oregano (Origanum vulgare) and Salvia triloba Extracts against Oral Cariogenic and Food-Origin Pathogenic Bacteria. Biomolecules. 2024; 14(6):619. https://doi.org/10.3390/biom14060619
Chicago/Turabian StyleAntoniadou, Maria, Georgios Rozos, Natalia Vaou, Konstantinos Zaralis, Caglar Ersanli, Athanasios Alexopoulos, Aikaterini Dadamogia, Theodoros Varzakas, Athina Tzora, and Chrysoula (Chrysa) Voidarou. 2024. "Comprehensive Bio-Screening of Phytochemistry and Biological Capacity of Oregano (Origanum vulgare) and Salvia triloba Extracts against Oral Cariogenic and Food-Origin Pathogenic Bacteria" Biomolecules 14, no. 6: 619. https://doi.org/10.3390/biom14060619
APA StyleAntoniadou, M., Rozos, G., Vaou, N., Zaralis, K., Ersanli, C., Alexopoulos, A., Dadamogia, A., Varzakas, T., Tzora, A., & Voidarou, C. (2024). Comprehensive Bio-Screening of Phytochemistry and Biological Capacity of Oregano (Origanum vulgare) and Salvia triloba Extracts against Oral Cariogenic and Food-Origin Pathogenic Bacteria. Biomolecules, 14(6), 619. https://doi.org/10.3390/biom14060619