
Eosinophils Contribute to Oral Tolerance via Induction of RORγt-Positive Antigen-Presenting Cells and RORγt-Positive Regulatory T Cells
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
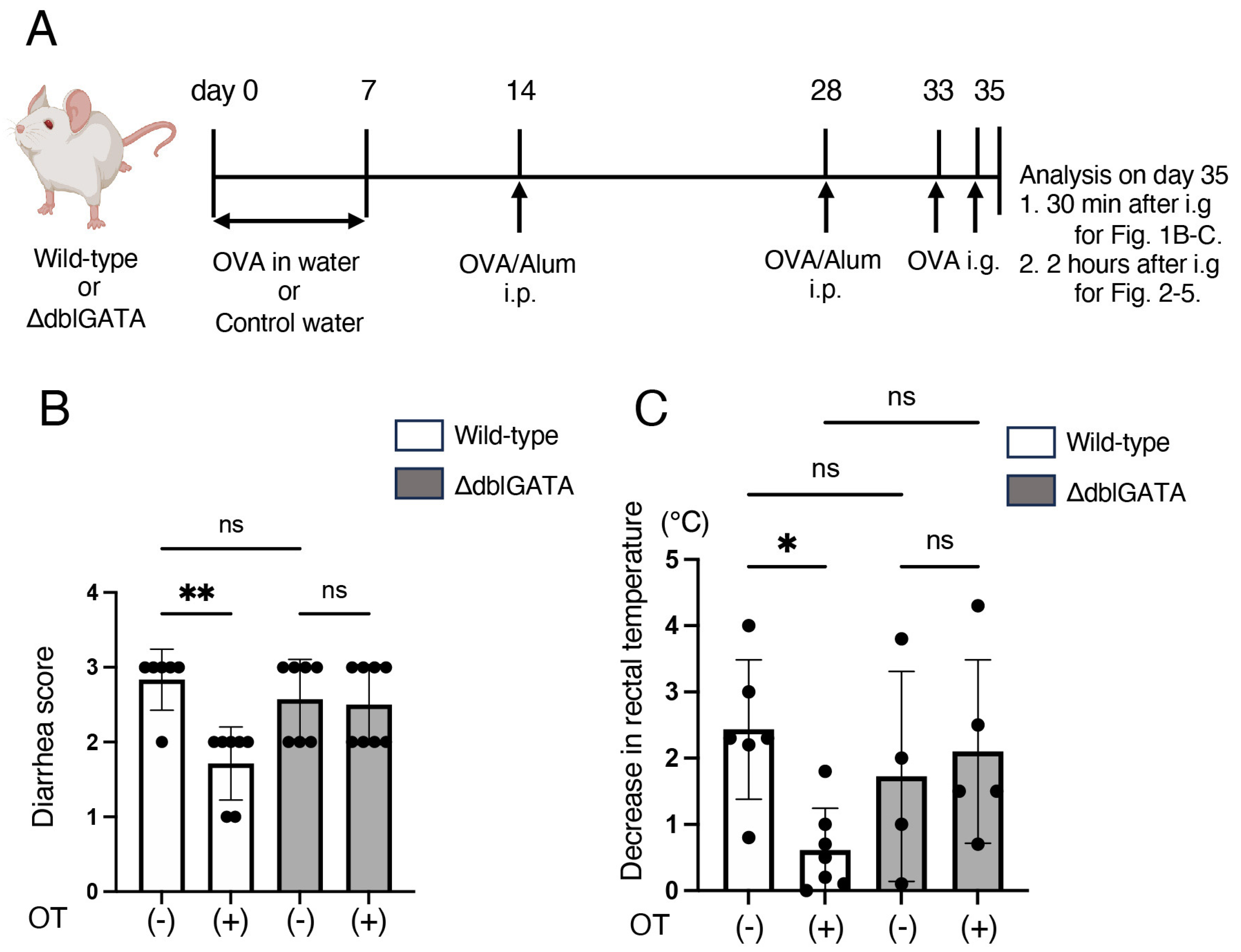
2.2. Induction of Oral Tolerance, Sensitization, and Food Allergy
2.3. Serum IgE Levels
2.4. Isolation of Cells from Small Intestinal Mucosa and Mesenteric Lymph Nodes
2.5. Flow Cytometry
2.6. Histological Analysis
2.7. Microbiota Analysis
2.8. Statistical Analysis
2.9. Data Availability
3. Results
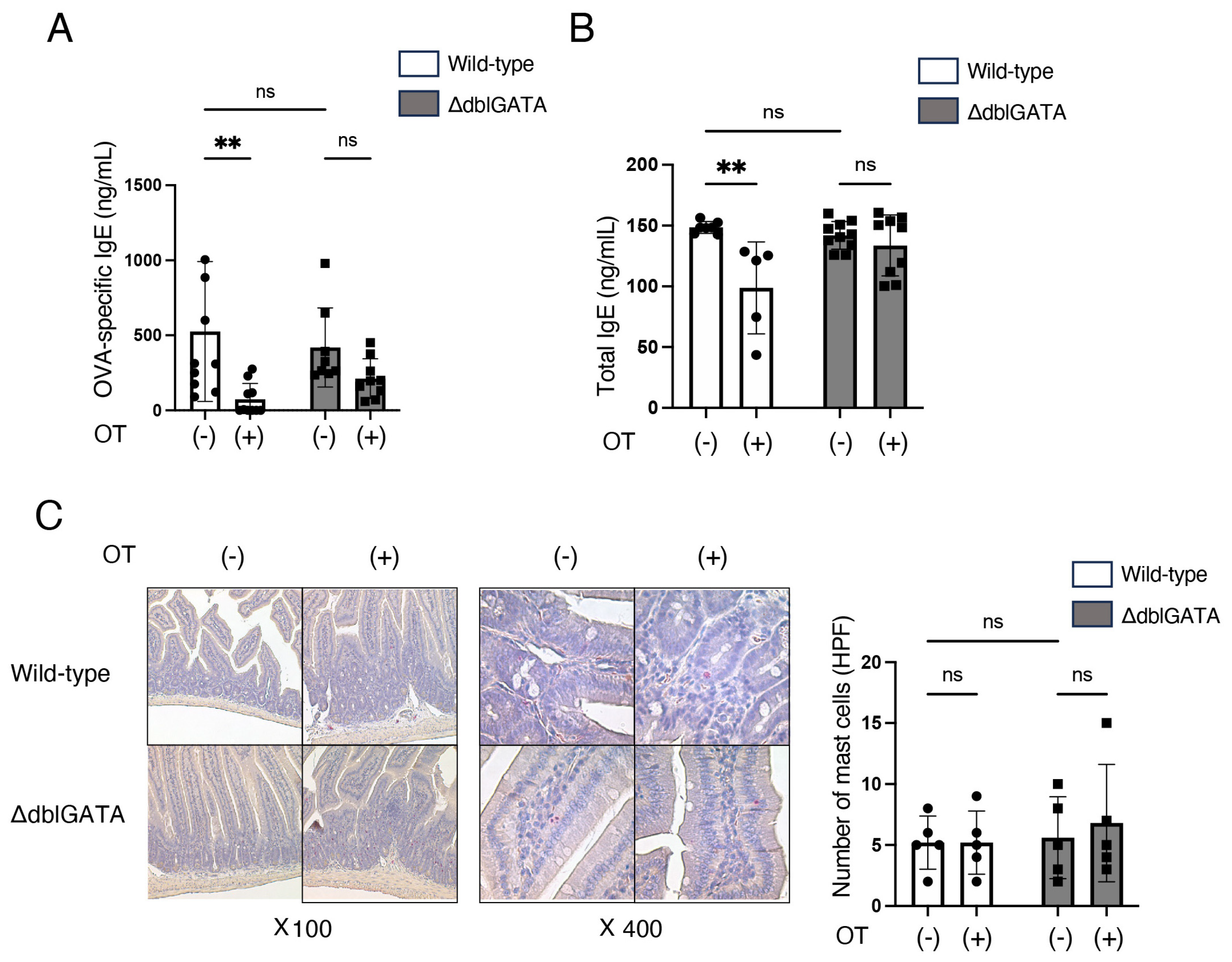
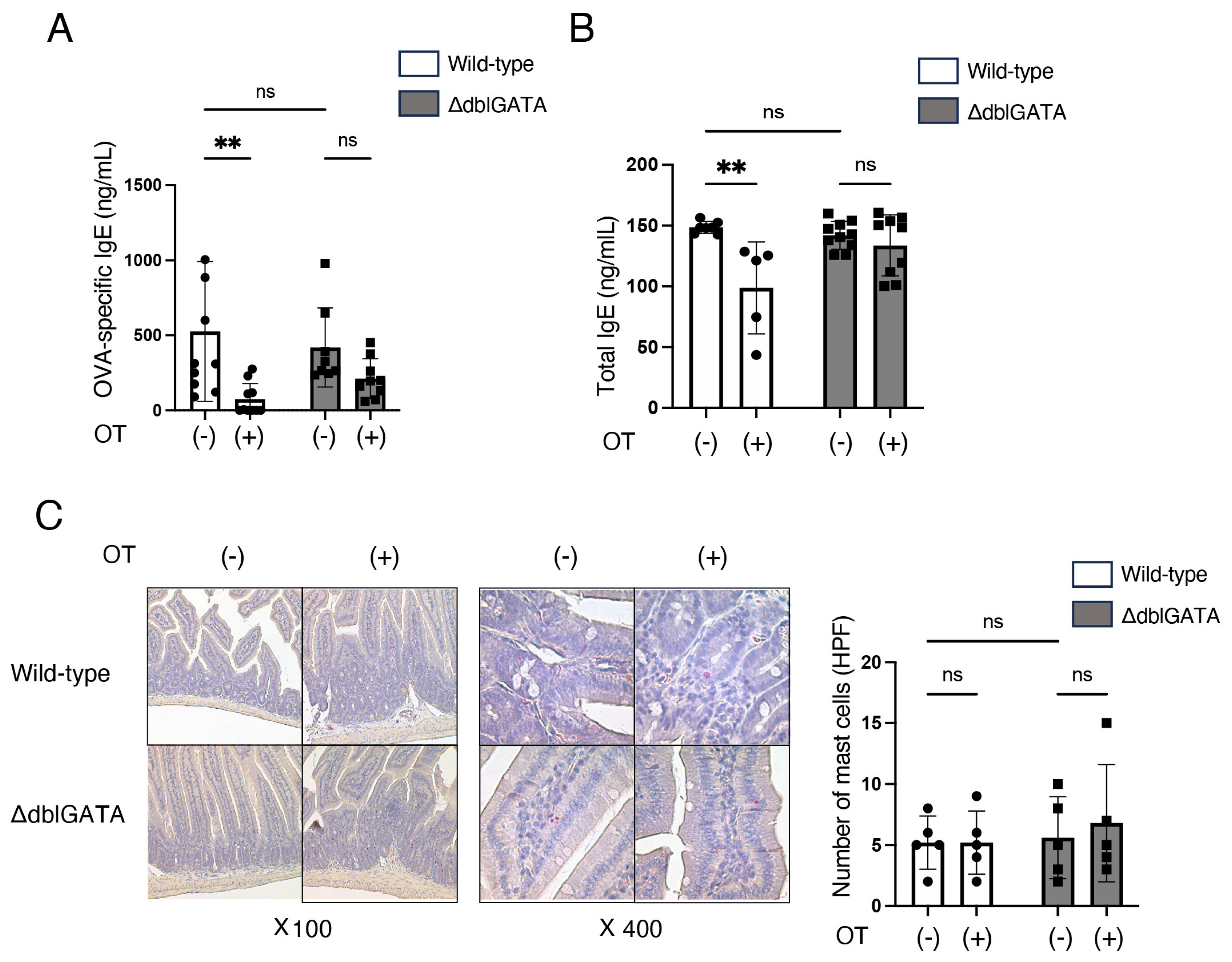
3.1. Eosinophils Are Involved in the Induction of Oral Tolerance to Food Allergy
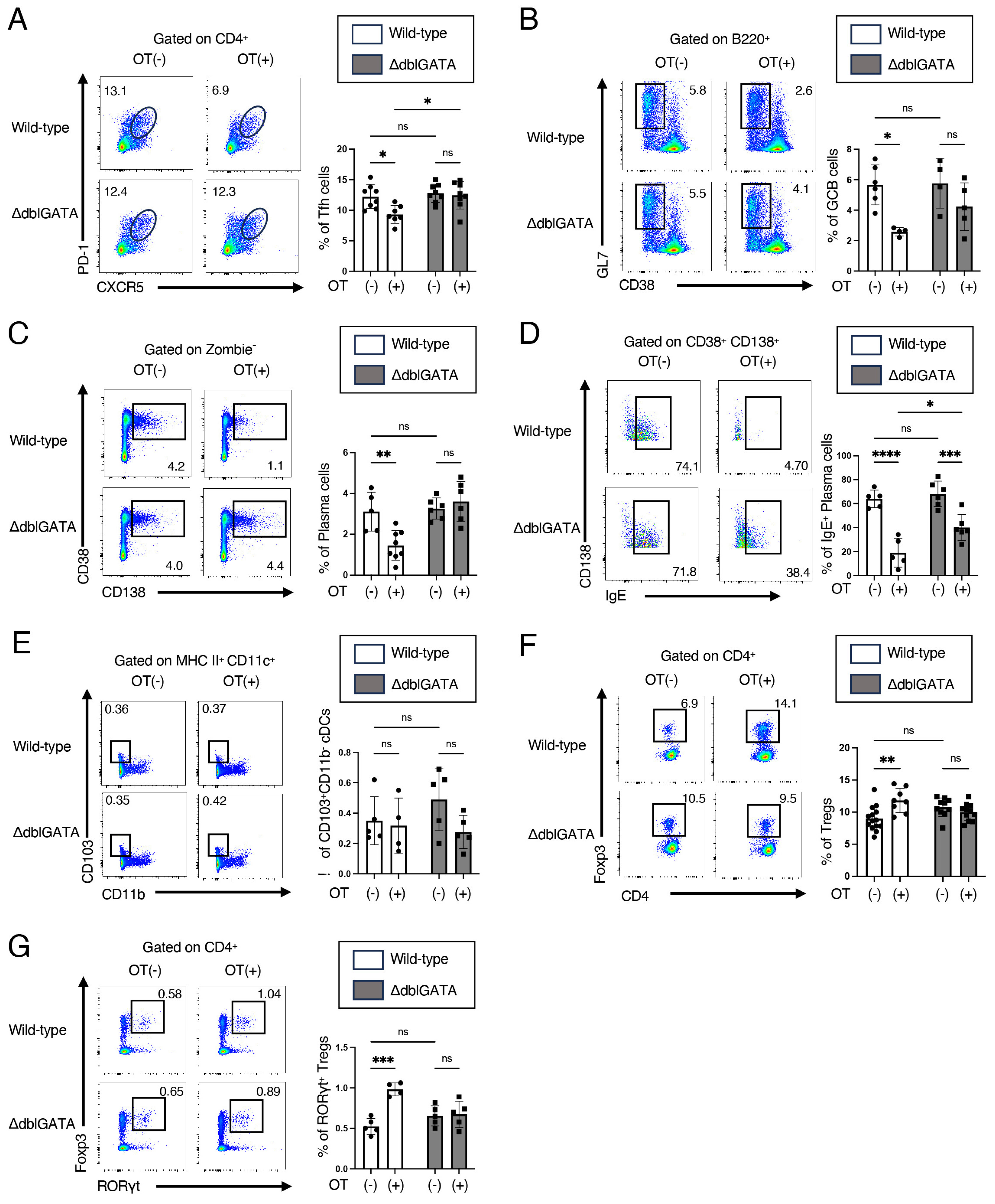
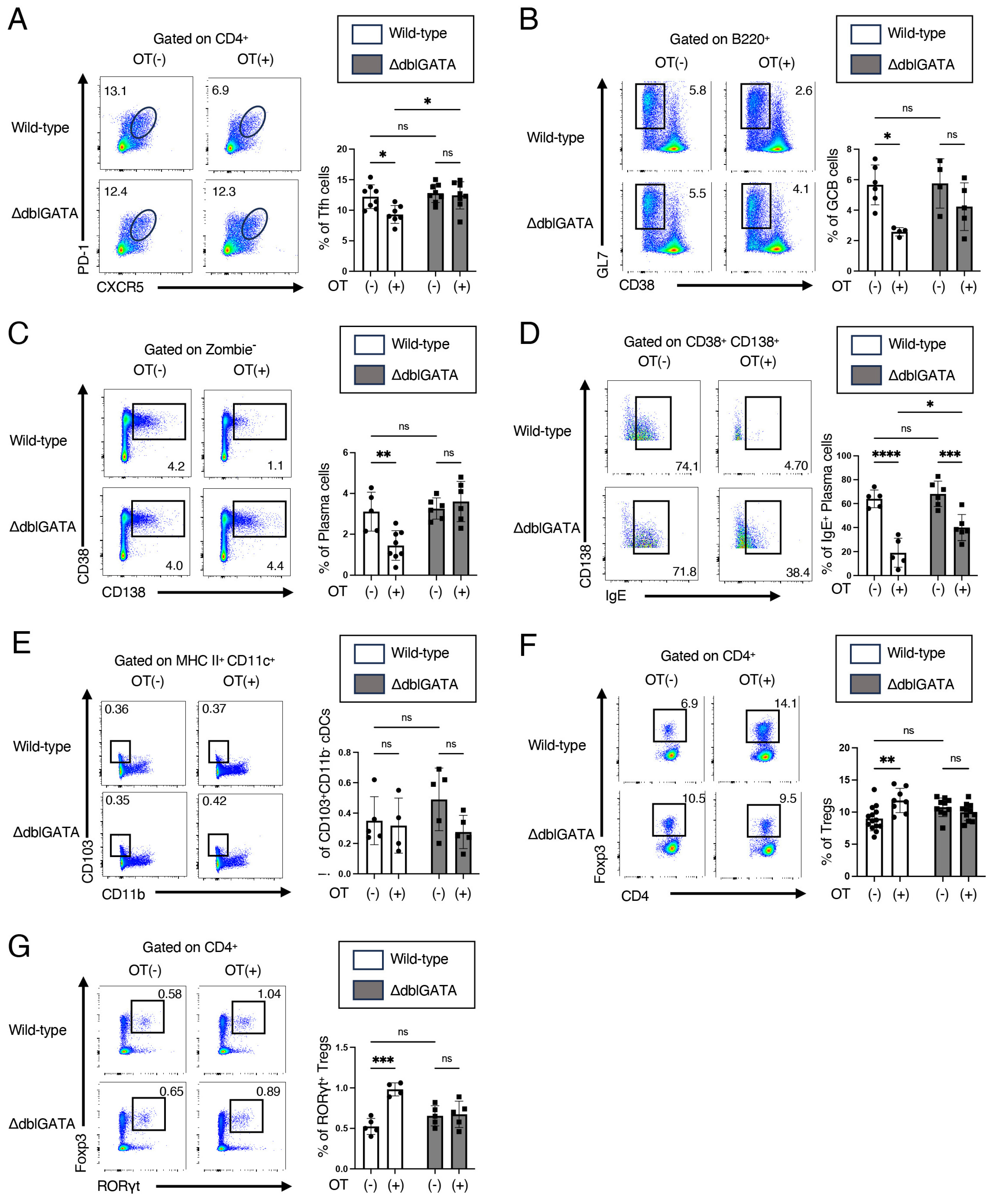
3.2. Eosinophils Contribute to RORγt+ Treg Differentiation and Attenuate the Germinal Center Responses in Mesenteric Lymph Nodes
3.3. Eosinophils Are Crucial for the Differentiation of RORγt+ APCs
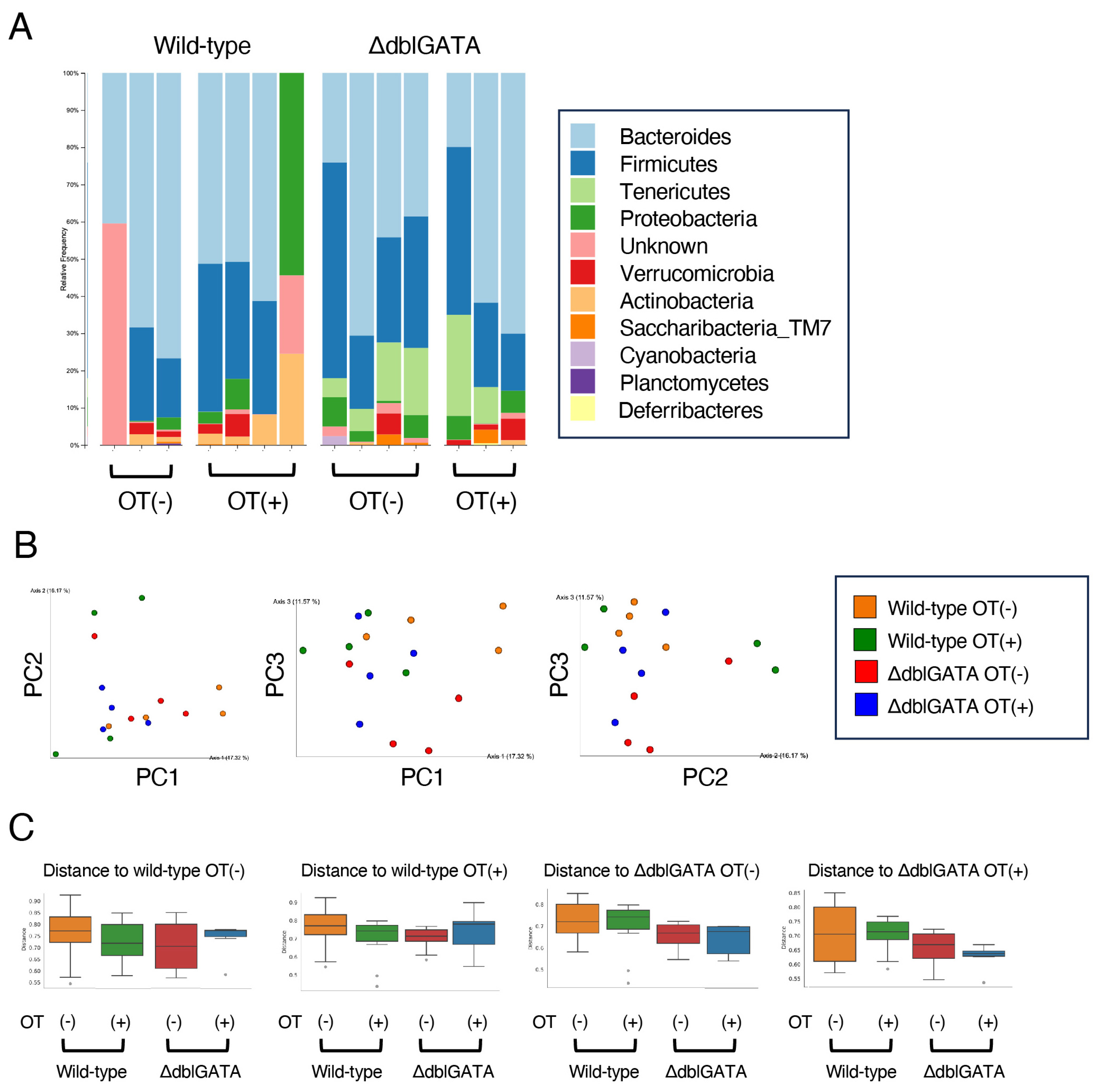
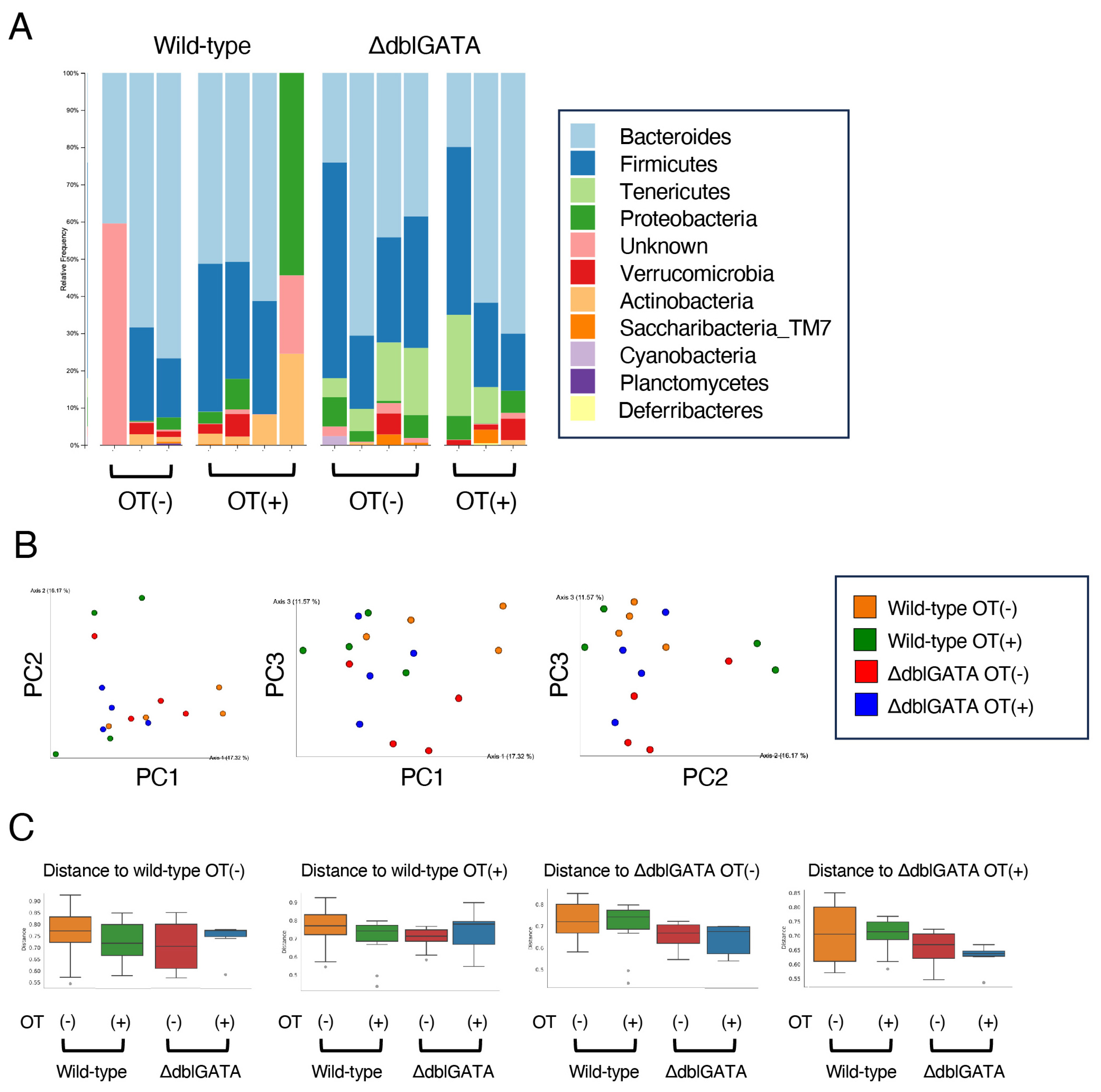
3.4. The Microbiome in the Small Intestine Is Similar between Wild-Type and ΔdblGATA Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rezende, R.M.; Weiner, H.L. History and mechanisms of oral tolerance. Semin. Immunol. 2017, 30, 3–11. [Google Scholar] [CrossRef]
- Rezende, R.M.; Weiner, H.L. Oral tolerance: An updated review. Immunol. Lett. 2022, 245, 29–37. [Google Scholar] [CrossRef]
- Coombes, J.L.; Siddiqui, K.R.; Arancibia-Carcamo, C.V.; Hall, J.; Sun, C.M.; Belkaid, Y.; Powrie, F. A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-beta and retinoic acid-dependent mechanism. J. Exp. Med. 2007, 204, 1757–1764. [Google Scholar] [CrossRef]
- Esterhazy, D.; Loschko, J.; London, M.; Jove, V.; Oliveira, T.Y.; Mucida, D. Classical dendritic cells are required for dietary antigen-mediated induction of peripheral regulatory T cells and tolerance. Nat. Immunol. 2016, 17, 545–555. [Google Scholar] [CrossRef]
- Kim, K.S.; Hong, S.W.; Han, D.; Yi, J.; Jung, J.; Yang, B.G.; Lee, G.Y.; Lee, M.; Surh, C.D. Dietary antigens limit mucosal immunity by inducing regulatory T cells in the small intestine. Science 2016, 351, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Wohlfert, E.A.; Grainger, J.R.; Bouladoux, N.; Konkel, J.E.; Oldenhove, G.; Ribeiro, C.H.; Hall, J.A.; Yagi, R.; Naik, S.; Bhairavabhotla, R.; et al. GATA3 controls Foxp3+ regulatory T cell fate during inflammation in mice. J. Clin. Investig. 2011, 121, 4503–4515. [Google Scholar] [CrossRef] [PubMed]
- Sefik, E.; Geva-Zatorsky, N.; Oh, S.; Konnikova, L.; Zemmour, D.; McGuire, A.M.; Burzyn, D.; Ortiz-Lopez, A.; Lobera, M.; Yang, J.; et al. Individual intestinal symbionts induce a distinct population of RORγ+ regulatory T cells. Science 2015, 349, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Thornton, A.M.; Korty, P.E.; Tran, D.Q.; Wohlfert, E.A.; Murray, P.E.; Belkaid, Y.; Shevach, E.M. Expression of Helios, an Ikaros transcription factor family member, differentiates thymic-derived from peripherally induced Foxp3+ T regulatory cells. J. Immunol. 2010, 184, 3433–3441. [Google Scholar] [CrossRef]
- Schiering, C.; Krausgruber, T.; Chomka, A.; Frohlich, A.; Adelmann, K.; Wohlfert, E.A.; Pott, J.; Griseri, T.; Bollrath, J.; Hegazy, A.N.; et al. The alarmin IL-33 promotes regulatory T-cell function in the intestine. Nature 2014, 513, 564–568. [Google Scholar] [CrossRef]
- Neumann, C.; Blume, J.; Roy, U.; The, P.P.; Vasanthakumar, A.; Beller, A.; Liao, Y.; Heinrich, F.; Arenzana, T.L.; Hackney, J.A.; et al. c-Maf-dependent Treg cell control of intestinal TH17 cells and IgA establishes host-microbiota homeostasis. Nat. Immunol. 2019, 20, 471–481. [Google Scholar] [CrossRef]
- Ohnmacht, C.; Park, J.H.; Cording, S.; Wing, J.B.; Atarashi, K.; Obata, Y.; Gaboriau-Routhiau, V.; Marques, R.; Dulauroy, S.; Fedoseeva, M.; et al. The microbiota regulates type 2 immunity through RORγt+ T cells. Science 2015, 349, 989–993. [Google Scholar] [CrossRef]
- Abdel-Gadir, A.; Stephen-Victor, E.; Gerber, G.K.; Noval-Rivas, M.; Wang, S.; Harb, H.; Wang, L.; Li, N.; Crestani, E.; Spielman, S.; et al. Microbiota therapy acts via a regulatory T cell MyD88/RORγt pathway to suppress food allergy. Nat. Med. 2019, 25, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Kedmi, R.; Najar, T.A.; Mesa, K.R.; Grayson, A.; Kroehling, L.; Hao, Y.; Hao, S.; Pokrovskii, M.; Xu, M.; Talbot, J.; et al. A RORγt+ cell instructs gut microbiota-specific Treg cell differentiation. Nature 2022, 610, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Lyu, M.; Suzuki, H.; Kang, L.; Gaspal, F.; Zhou, W.; Goc, J.; Zhou, L.; Zhou, J.; Zhang, W.; Shen, Z.; et al. ILC3s select microbiota-specific regulatory T cells to establish tolerance in the gut. Nature 2022, 610, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Akagbosu, B.; Tayyebi, Z.; Shibu, G.; Paucar-Iza, Y.A.; Deep, D.; Parisotto, Y.F.; Fisher, L.; Pasolli, H.A.; Thevin, V.; Elmentaite, R.; et al. Novel antigen-presenting cell imparts Treg-dependent tolerance to gut microbiota. Nature 2022, 610, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.L.M.; Rothenberg, M.E. Spotlight on eosinophils. J. Allergy Clin. Immunol. 2022, 149, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, A.; Crepaz, D.; Arnold, I.C. Emerging functions of tissue-resident eosinophils. J. Exp. Med. 2023, 220, e20221435. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Cantor, A.B.; Yang, H.; Browne, C.; Wells, R.A.; Fujiwara, Y.; Orkin, S.H. Targeted deletion of a high-affinity GATA-binding site in the GATA-1 promoter leads to selective loss of the eosinophil lineage in vivo. J. Exp. Med. 2022, 195, 1387–1395. [Google Scholar] [CrossRef]
- Nei, Y.; Obata-Ninomiya, K.; Tsutsui, H.; Ishiwata, K.; Miyasaka, M.; Matsumoto, K.; Nakae, S.; Kanuka, H.; Inase, N.; Karasuyama, H. GATA-1 regulates the generation and function of basophils. Proc. Natl. Acad. Sci. USA 2013, 110, 18620–18625. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Tomar, S.; Dharne, M.; Ganesan, V.; Smith, A.; Yang, Y.; Waggoner, L.; Wang, Y.-H.; Hogan, S.P. Deletion of ΔdblGata motif leads to increased predisposition and severity of IgE-mediated food-induced anaphylaxis response. PLoS ONE 2019, 14, e0219375. [Google Scholar] [CrossRef]
- Jung, Y.; Wen, T.; Mingler, M.K.; Caldwell, J.M.; Wang, Y.H.; Chaplin, D.D.; Lee, E.H.; Jang, M.H.; Woo, S.Y.; Seoh, J.Y.; et al. Il-1β in eosinophil-mediated small intestinal homeostasis and IgA production. Mucosal Immunol. 2015, 8, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Chu, V.T.; Beller, A.; Rausch, S.; Strandmark, J.; Zanker, M.; Arbach, O.; Kruglov, A.; Berek, C. Eosinophils promote generation and maintenance of immunoglobulin-A-expressing plasma cells and contribute to gut immune homeostasis. Immunity 2014, 40, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Brass, A.; Knight, C.G.; Cruickshank, S.M. Gut eosinophils and their impact on the mucus-resident microbiota. Immunology 2019, 158, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.; Jimenez-Saiz, R.; Verschoor, C.P.; Walker, T.D.; Goncharova, S.; Loop-Guevara, A.; Shen, P.; Gordon, M.E.; Barra, N.G.; Bassett, J.D.; et al. Indigenous enteric eosinophils control DCs to initiate a primary Th2 immune response in vivo. J. Exp. Med. 2014, 211, 1657–1671. [Google Scholar] [CrossRef]
- FitzPatrick, R.D.; Kennedy, M.H.E.; Lawrence, K.M.; Gauthier, C.M.; Moeller, B.E.; Robinson, A.N.; Reynolds, L.A. Littermate-controlled experiments reveal eosinophils are not essential for maintaining steady-state IgA and demonstrate the influence of rearing conditions on antibody phenotypes in eosinophil-deficient mice. Front. Immunol. 2020, 11, 557960. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, R.; Lee, E.J.; Jang, M.S.; Jeun, E.J.; Hong, C.P.; Kim, J.H.; Park, A.; Yun, C.H.; Hong, S.W.; Kim, Y.M.; et al. Small intestinal eosinophils regulate Th17 cells by producing IL-1 receptor antagonist. J. Exp. Med. 2016, 213, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Chu, V.T.; Frohlich, A.; Steinhauser, G.; Scheel, T.; Roch, T.; Fillatreau, S.; Lee, J.J.; Lohning, M.; Berek, C. Eosinophils are required for the maintenance of plasma cells in the bone marrow. Nat. Immunol. 2011, 12, 151–159. [Google Scholar] [CrossRef]
- Fallegger, A.; Priola, M.; Artola-Boran, M.; Nunez, N.G.; Wild, S.; Gurtner, A.; Becher, B.; Yousefi, S.; Simon, H.U.; Arnold, I.C.; et al. TGF-β production by eosinophils drives the expansion of peripherally induced neuropilin−RORγt+ regulatory T-cells during bacterial and allergen challenge. Mucosal Immunol. 2022, 15, 504–514. [Google Scholar] [CrossRef]
- Tanaka, S.; Suto, A.; Iwamoto, T.; Kashiwakuma, D.; Kagami, S.; Suzuki, K.; Takatori, H.; Tamachi, T.; Hirose, K.; Onodera, A.; et al. Sox5 and c-Maf cooperatively induce Th17 cell differentiation via RORγt induction as downstream targets of Stat3. J. Exp. Med. 2014, 211, 1857–1874. [Google Scholar] [CrossRef]
- Friend, D.S.; Ghildyal, N.; Austen, K.F.; Gurish, M.F.; Matsumoto, R.; Stevens, R.L. Mast cells that reside at different locations in the jejunum of mice infected with Trichinella spiralis exhibit sequential changes in their granule ultrastructure and chymase phenotype. J. Cell Biol. 1996, 135, 279–290. [Google Scholar] [CrossRef]
- Oettgen, H.C. Mast cells in food allergy: Inducing immediate reactions and shaping long-term immunity. J. Allergy Clin. Immunol. 2023, 151, 21–25. [Google Scholar] [CrossRef]
- Victora, G.D.; Nussenzweig, M.C. Germinal centers. Annu. Rev. Immunol. 2022, 40, 413–442. [Google Scholar] [CrossRef] [PubMed]
- Lochner, M.; Peduto, L.; Cherrier, M.; Sawa, S.; Langa, F.; Varona, R.; Riethmacher, D.; Si-Tahar, M.; Santo, J.P.D.; Eberl, G. In vivo equilibrium of proinflammatory IL-17+ and regulatory IL-10+Foxp3+RORγt+ T cells. J. Exp. Med. 2008, 205, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.-H.; Hagemann, S.; Mamareli, P.; Lauer, U.; Hoffmann, U.; Beckstette, M.; Fohse, L.; Prinz, I.; Pezoldt, J.; Suerbaum, S.; et al. Foxp3+ T cells expressing RORγt represent a stable T-cell effector lineage with enhanced suppressive capacity during intestinal inflammation. Mucosal Immunol. 2015, 9, 444–457. [Google Scholar] [CrossRef] [PubMed]
- Abramson, J.; Dobes, J.; Liu, M.; Sonnenberg, G.F. The emerging family of RORγt+ antigen-presenting cells. Nat. Rev. Immunol. 2023, 24, 64–77. [Google Scholar] [CrossRef]
- Hwang, D.; Watanabe-Ishikawa, L.L.; Seyedsadr, M.S.; Mari, E.; Kasimoglu, E.; Sahin, Z.; Boehm, A.; Jang, S.; Rasouli, J.; Vaccaro, C.; et al. GATA1 controls numbers of hematopoietic progenitors and their response to autoimmune neuroinflammation. Blood Adv. 2022, 6, 5980–5994. [Google Scholar] [CrossRef] [PubMed]
- Forster, R.; Davalos-Misslitz, A.C.; Rot, A. CCR7 and its ligands: Balancing immunity and tolerance. Nat. Rev. Immunol. 2008, 8, 362–371. [Google Scholar] [CrossRef]
- Gowthaman, U.; Chen, J.S.; Zhang, B.; Flynn, W.F.; Lu, Y.; Song, W.; Joseph, J.; Gertie, J.A.; Xu, L.; Collet, M.A.; et al. Identification of a T follicular helper cell subset that drives anaphylactic IgE. Science 2019, 365, eaaw6433. [Google Scholar] [CrossRef]
- Melo-Gonzalez, F.; Kammoun, H.; Evren, E.; Dutton, E.E.; Papadopoulou, M.; Bradford, B.M.; Tanes, C.; Fardus-Reid, F.; Swann, J.R.; Bittinger, K.; et al. Antigen-presenting ILC3 regulates T cell-dependent IgA responses to colonic mucosal bacteria. J. Exp. Med. 2019, 216, 728–742. [Google Scholar] [CrossRef]
- Wang, W.; Li, Y.; Hao, J.; He, Y.; Dong, X.; Fu, Y.-X.; Guo, X. The interaction between lymphoid tissue inducer-like cells and T cells in the mesenteric lymph node restrains intestinal humoral immunity. Cell Rep. 2020, 32, 107936. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurihara, S.; Suzuki, K.; Yokota, M.; Ito, T.; Hayashi, Y.; Kikuchi, R.; Kageyama, T.; Meguro, K.; Tanaka, S.; Iwata, A.; et al. Eosinophils Contribute to Oral Tolerance via Induction of RORγt-Positive Antigen-Presenting Cells and RORγt-Positive Regulatory T Cells. Biomolecules 2024, 14, 89. https://doi.org/10.3390/biom14010089
Kurihara S, Suzuki K, Yokota M, Ito T, Hayashi Y, Kikuchi R, Kageyama T, Meguro K, Tanaka S, Iwata A, et al. Eosinophils Contribute to Oral Tolerance via Induction of RORγt-Positive Antigen-Presenting Cells and RORγt-Positive Regulatory T Cells. Biomolecules. 2024; 14(1):89. https://doi.org/10.3390/biom14010089
Chicago/Turabian StyleKurihara, Shunjiro, Kotaro Suzuki, Masaya Yokota, Takashi Ito, Yuki Hayashi, Ryo Kikuchi, Takahiro Kageyama, Kazuyuki Meguro, Shigeru Tanaka, Arifumi Iwata, and et al. 2024. "Eosinophils Contribute to Oral Tolerance via Induction of RORγt-Positive Antigen-Presenting Cells and RORγt-Positive Regulatory T Cells" Biomolecules 14, no. 1: 89. https://doi.org/10.3390/biom14010089
APA StyleKurihara, S., Suzuki, K., Yokota, M., Ito, T., Hayashi, Y., Kikuchi, R., Kageyama, T., Meguro, K., Tanaka, S., Iwata, A., Goto, Y., Suto, A., & Nakajima, H. (2024). Eosinophils Contribute to Oral Tolerance via Induction of RORγt-Positive Antigen-Presenting Cells and RORγt-Positive Regulatory T Cells. Biomolecules, 14(1), 89. https://doi.org/10.3390/biom14010089