
As Verified with the Aid of Biotinylated Spermine, the Brain Cannot Take up Polyamines from the Bloodstream Leaving It Solely Dependent on Local Biosynthesis

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Synthesis of B-X-SPM
2.1.2. Characterization of B-X-SPM
2.1.3. Solutions and Drugs
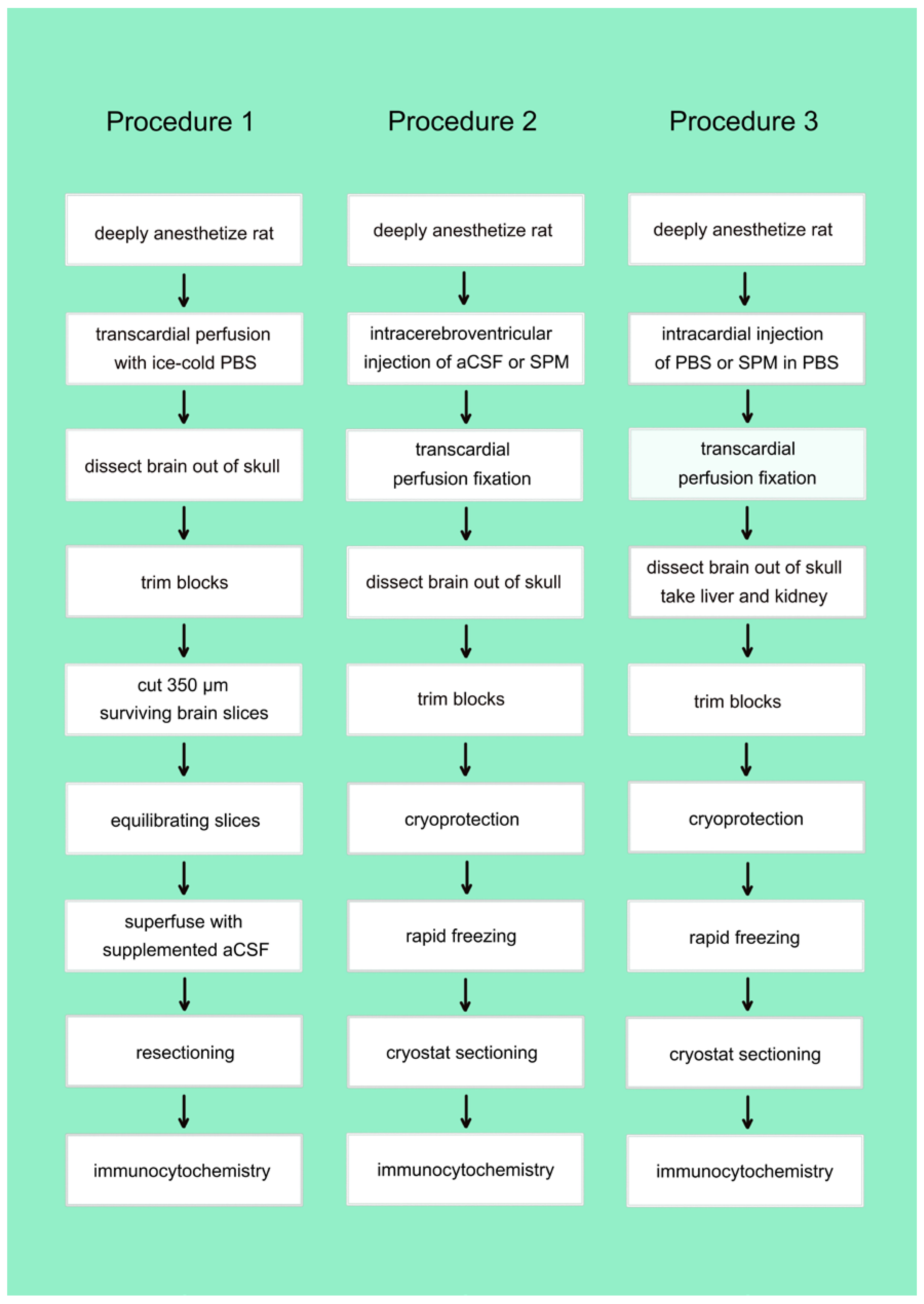
2.1.4. Preparation of Acute Brain Slices
2.1.5. Superfusion of Brain Slices
2.1.6. In Vivo Injections
2.1.7. Immunocytochemistry
2.1.8. Selective Visualization of Biotinylated SPM in Tissues
3. Results
3.1. Synthesis and Characterization of Biotinylated and Extended Spermine (B-X-SPM)
3.2. In Vitro Uptake of Native or Biotinylated SPM in Brain Slices
3.3. Uptake of Native SPM in Acute Brain Slices
3.4. Uptake of Biotinylated SPM (B-X-SPM) in Acute Brain Slices
3.5. In Vivo Uptake of Native or Biotinylated SPM into the Brain from Intraventricular CSF or from the Bloodstream
3.6. Incorporation of B-X-SPM into Astrocytes Is Based on a PA-Specific Uptake System
3.7. In Vivo Uptake of Biotinylated SPM into the Brain from Intraventricular CSF
3.8. In Vivo Uptake of Biotinylated SPM into the Brain from the Bloodstream
4. Discussion
4.1. Technical Considerations
4.2. Native and Biotinylated SPM Are Taken up by Astrocytes and Neurons in Brain Slices
4.3. Uptake of Biotinylated SPM (B-X-SPM) by Brain Slices Is Different in Protoplasmic and Fibrous Astrocytes
4.4. Native or Biotinylated SPM Are Not Taken up into the Brain from the Bloodstream
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rieck, J.; Skatchkov, S.N.; Derst, C.; Eaton, M.J.; Veh, R.W. Unique Chemistry, Intake, and Metabolism of Polyamines in the Central Nervous System (CNS) and Its Body. Biomolecules 2022, 12, 501. [Google Scholar] [CrossRef]
- Matsumoto, M.; Kakizoe, K.; Benno, Y. Comparison of Fecal Microbiota and Polyamine Concentration in Adult Patients with Intractable Atopic Dermatitis and Healthy Adults. Microbiol. Immunol. 2007, 51, 37–46. [Google Scholar] [CrossRef]
- Madeo, F.; Hofer, S.J.; Pendl, T.; Bauer, M.A.; Eisenberg, T.; Carmona-Gutierrez, D.; Kroemer, G. Nutritional Aspects of Spermidine. Annu. Rev. Nutr. 2020, 40, 135–159. [Google Scholar] [CrossRef]
- Kibe, R.; Kurihara, S.; Sakai, Y.; Suzuki, H.; Ooga, T.; Sawaki, E.; Muramatsu, K.; Nakamura, A.; Yamashita, A.; Kitada, Y.; et al. Upregulation of colonic luminal polyamines produced by intestinal microbiota delays senescence in mice. Sci. Rep. 2014, 4, 4548. [Google Scholar] [CrossRef]
- Milovic, V. Polyamines in the gut lumen: Bioavailability and biodistribution. Eur. J. Gastroenterol. Hepatol. 2001, 13, 1021–1025. [Google Scholar] [CrossRef]
- Skatchkov, S.N.; Antonov, S.M.; Eaton, M.J. Glia and glial polyamines. Role in brain function in health and disease. Biochem. (Mosc.) Suppl. Ser. A Membr. Cell Biol. 2016, 10, 73–98. [Google Scholar] [CrossRef]
- Lajtha, A.; Sershen, H. Substrate specificity of uptake of diamines in mouse brain slices. Arch. Biochem. Biophys. 1974, 165, 539–547. [Google Scholar] [CrossRef]
- Sershen, H.; Lajtha, A. Perinatal changes of transport systems for amino acids in slices of mouse brain. Neurochem. Res. 1976, 1, 417–428. [Google Scholar] [CrossRef]
- Shin, W.-W.; Fong, W.-F.; Pang, S.-F.; Wong, P.C.-L. Limited Blood-Brain Barrier Transport of Polyamines. J. Neurochem. 1985, 44, 1056–1059. [Google Scholar] [CrossRef]
- Hopman, A.H.; Ramaekers, F.C.; Speel, E.J. Rapid synthesis of biotin-, digoxigenin-, trinitrophenyl-, and fluorochrome-labeled tyramides and their application for In situ hybridization using CARD amplification. J. Histochem. Cytochem. 1998, 46, 771–777. [Google Scholar] [CrossRef]
- Madai, V.I.; Poller, W.C.; Peters, D.; Berger, J.; Paliege, K.; Bernard, R.; Veh, R.W.; Laube, G. Synaptic localisation of agmatinase in rat cerebral cortex revealed by virtual pre-embedding. Amino Acids 2012, 43, 1399–1403. [Google Scholar] [CrossRef]
- Somogyi, P.; Takagi, H. A note on the use of picric acid-paraformaldehyde-glutaraldehyde fixative for correlated light and electron microscopic immunocytochemistry. Neuroscience 1982, 7, 1779–1783. [Google Scholar] [CrossRef]
- Bernard, R.; Veh, R.W. Individual neurons in the rat lateral habenular complex project mostly to the dopaminergic ventral tegmental area or to the serotonergic raphe nuclei. J. Comp. Neurol. 2012, 520, 2545–2558. [Google Scholar] [CrossRef]
- Laube, G.; Veh, R.W. Astrocytes, not neurons, show most prominent staining for spermidine/spermine-like immunoreactivity in adult rat brain. Glia 1997, 19, 171–179. [Google Scholar] [CrossRef]
- Malpica-Nieves, C.J.; Rivera, Y.; Rivera-Aponte, D.E.; Phanstiel, O.; Veh, R.W.; Eaton, M.J.; Skatchkov, S.N. Uptake of Biotinylated Spermine in Astrocytes: Effect of Cx43 siRNA, HIV-Tat Protein and Polyamine Transport Inhibitor on Polyamine Uptake. Biomolecules 2021, 11, 1187. [Google Scholar] [CrossRef]
- Krauss, M.; Langnaese, K.; Richter, K.; Brunk, I.; Wieske, M.; Ahnert-Hilger, G.; Veh, R.W.; Laube, G. Spermidine synthase is prominently expressed in the striatal patch compartment and in putative interneurones of the matrix compartment. J. Neurochem. 2006, 97, 174–189. [Google Scholar] [CrossRef]
- Biedermann, B.; Skatchkov, S.N.; Brunk, I.; Bringmann, A.; Pannicke, T.; Bernstein, H.-G.; Faude, F.; Germer, A.; Veh, R.; Reichenbach, A. Spermine/spermidine is expressed by retinal glial (müller) cells and controls distinct K+ channels of their membrane. Glia 1998, 23, 209–220. [Google Scholar] [CrossRef]
- Skatchkov, S.N.; Eaton, M.J.; Krušek, J.; Veh, R.W.; Biedermann, B.; Bringmann, A.; Pannicke, T.; Orkand, R.K.; Reichenbach, A. Spatial distribution of spermine/spermidine content and K+-current rectification in frog retinal glial (Müller) cells. Glia 2000, 31, 84–90. [Google Scholar] [CrossRef]
- Kovács, Z.; Skatchkov, S.N.; Veh, R.W.; Szabó, Z.; Németh, K.; Szabó, P.T.; Kardos, J.; Héja, L. Critical Role of Astrocytic Polyamine and GABA Metabolism in Epileptogenesis. Front. Cell. Neurosci. 2022, 15, 787319. [Google Scholar] [CrossRef]
- Bernstein, H.-G.; Müller, M. The cellular localization of the l-ornithine decarboxylase/polyamine system in normal and diseased central nervous systems. Prog. Neurobiol. 1999, 57, 485–505. [Google Scholar] [CrossRef] [PubMed]
- Kilpeläinen, P.; Rybnikova, E.; Hietala, O.; Pelto-Huikko, M. Expression of ODC and its regulatory protein antizyme in the adult rat brain. J. Neurosci. Res. 2000, 62, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Laube, G.; Bernstein, H.-G.; Wolf, G.; Veh, R.W. Differential distribution of spermidine/spermine-like immunoreactivity in neurons of the adult rat brain. J. Comp. Neurol. 2002, 444, 369–386. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.D.R.; Petrova, R.; Eng, L.; Joyner, A.L. Sonic Hedgehog Regulates Discrete Populations of Astrocytes in the Adult Mouse Forebrain. J. Neurosci. 2010, 30, 13597–13608. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weiss, T.; Bernard, R.; Laube, G.; Rieck, J.; Eaton, M.J.; Skatchkov, S.N.; Veh, R.W. As Verified with the Aid of Biotinylated Spermine, the Brain Cannot Take up Polyamines from the Bloodstream Leaving It Solely Dependent on Local Biosynthesis. Biomolecules 2023, 13, 1114. https://doi.org/10.3390/biom13071114
Weiss T, Bernard R, Laube G, Rieck J, Eaton MJ, Skatchkov SN, Veh RW. As Verified with the Aid of Biotinylated Spermine, the Brain Cannot Take up Polyamines from the Bloodstream Leaving It Solely Dependent on Local Biosynthesis. Biomolecules. 2023; 13(7):1114. https://doi.org/10.3390/biom13071114
Chicago/Turabian StyleWeiss, Torsten, René Bernard, Gregor Laube, Julian Rieck, Misty J. Eaton, Serguei N. Skatchkov, and Rüdiger W. Veh. 2023. "As Verified with the Aid of Biotinylated Spermine, the Brain Cannot Take up Polyamines from the Bloodstream Leaving It Solely Dependent on Local Biosynthesis" Biomolecules 13, no. 7: 1114. https://doi.org/10.3390/biom13071114
APA StyleWeiss, T., Bernard, R., Laube, G., Rieck, J., Eaton, M. J., Skatchkov, S. N., & Veh, R. W. (2023). As Verified with the Aid of Biotinylated Spermine, the Brain Cannot Take up Polyamines from the Bloodstream Leaving It Solely Dependent on Local Biosynthesis. Biomolecules, 13(7), 1114. https://doi.org/10.3390/biom13071114