

Activation of Metabotropic Glutamate Receptor (mGlu2) and Muscarinic Receptors (M1, M4, and M5), Alone or in Combination, and Its Impact on the Acquisition and Retention of Learning in the Morris Water Maze, NMDA Expression and cGMP Synthesis


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drug Preparation and Administration
2.3. Morris Water Maze (MWM)
2.4. Synaptosome Preparation and Western Blotting
2.5. cGMP ELISA
2.6. Statistics
3. Results
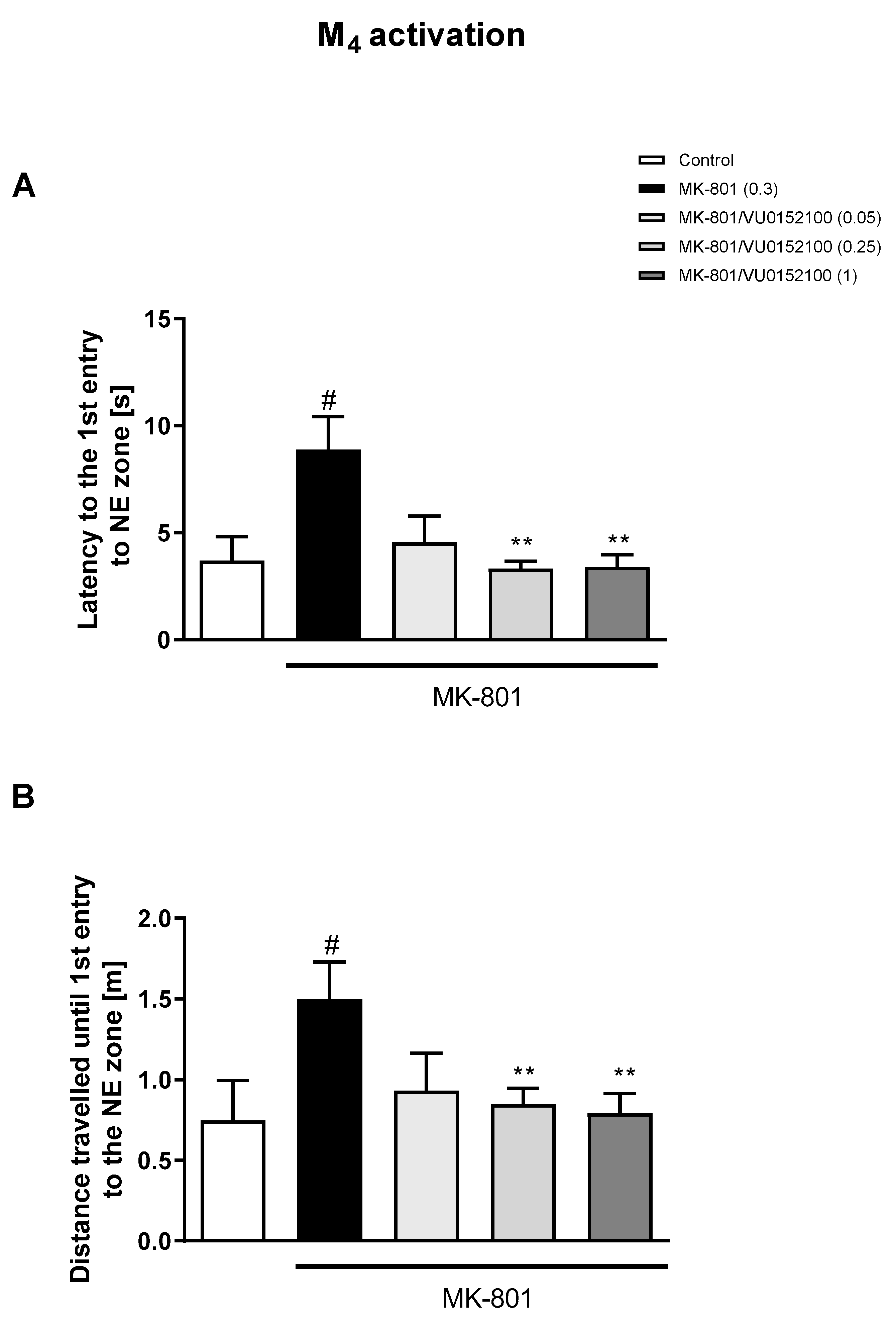
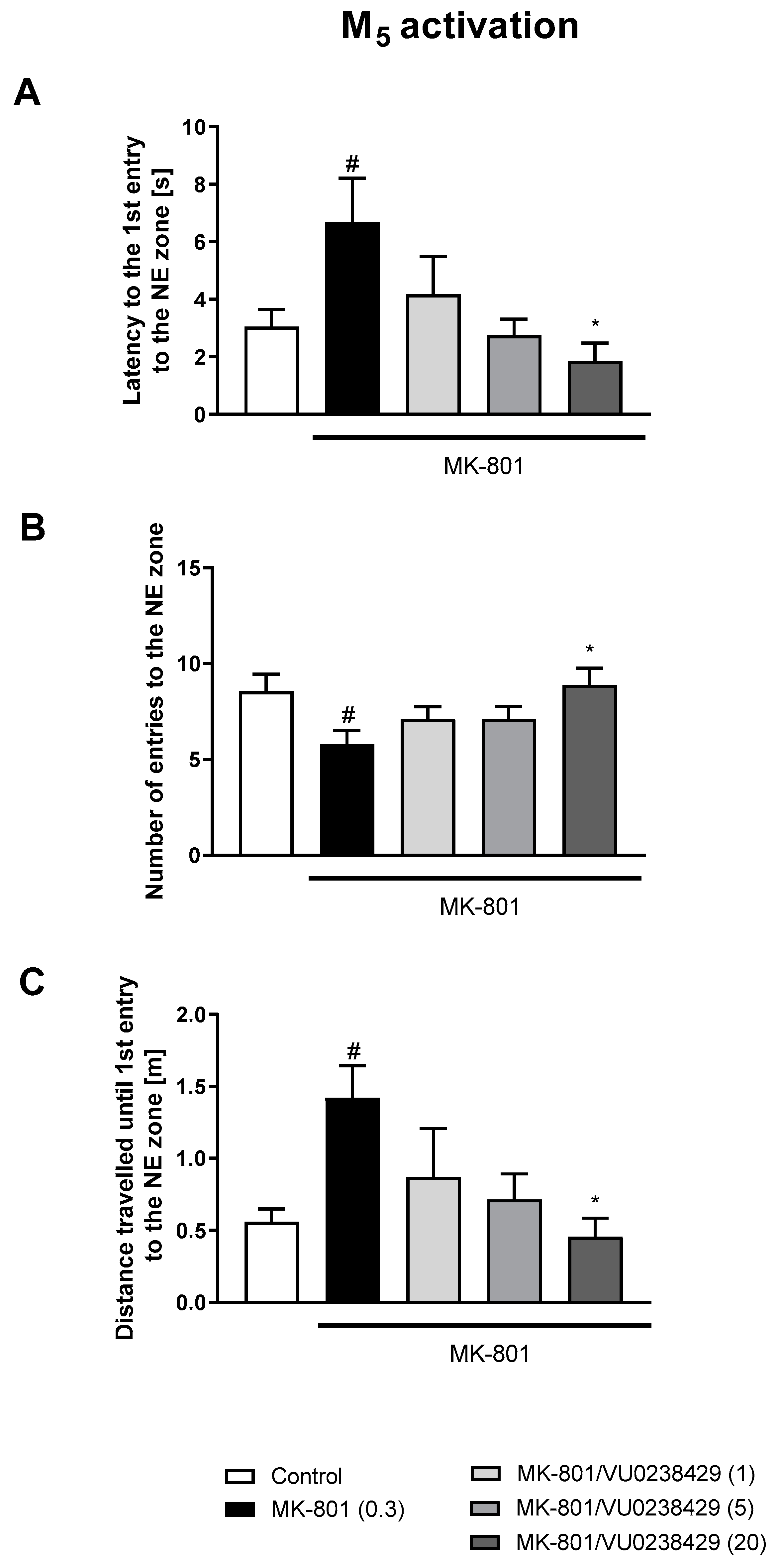
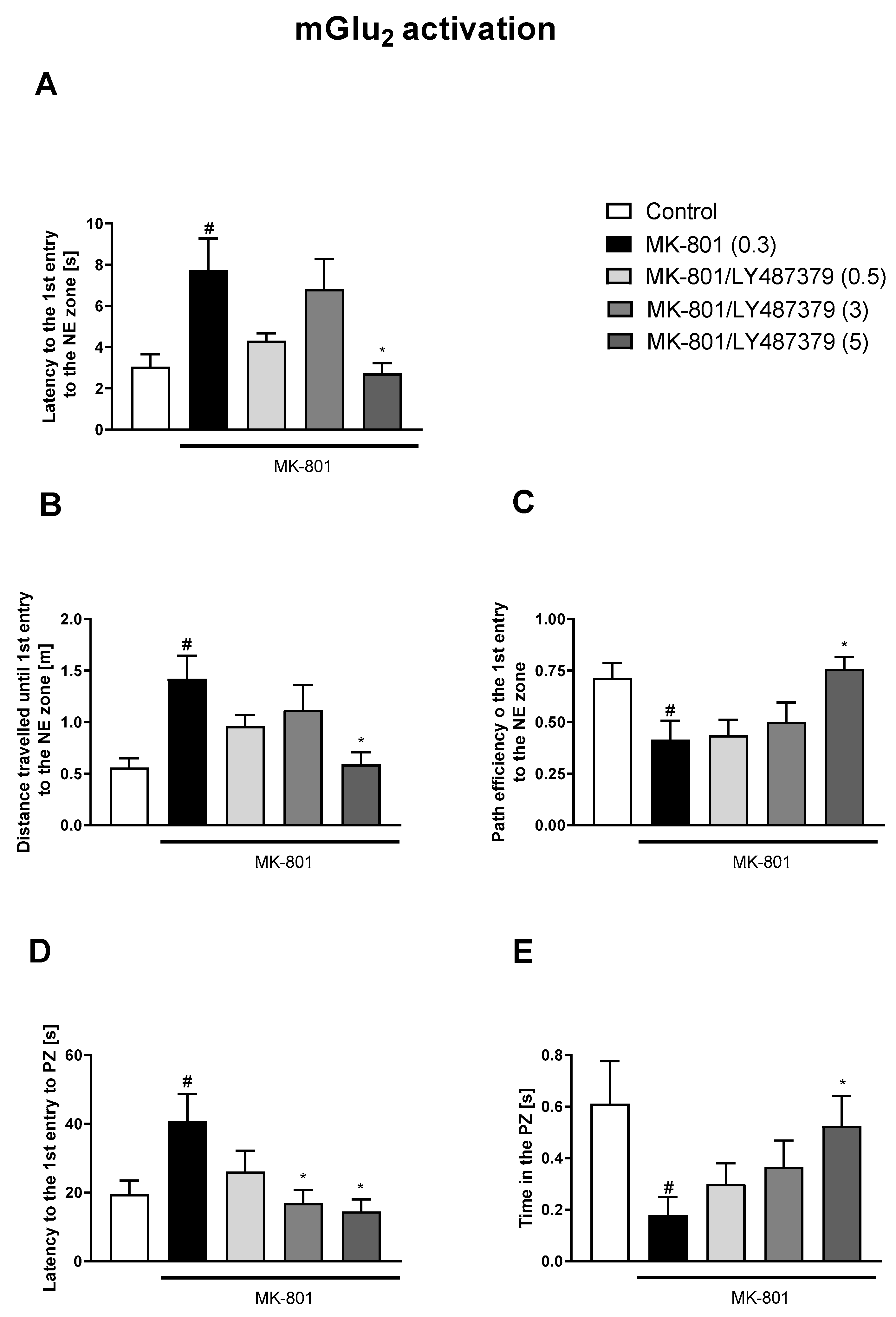
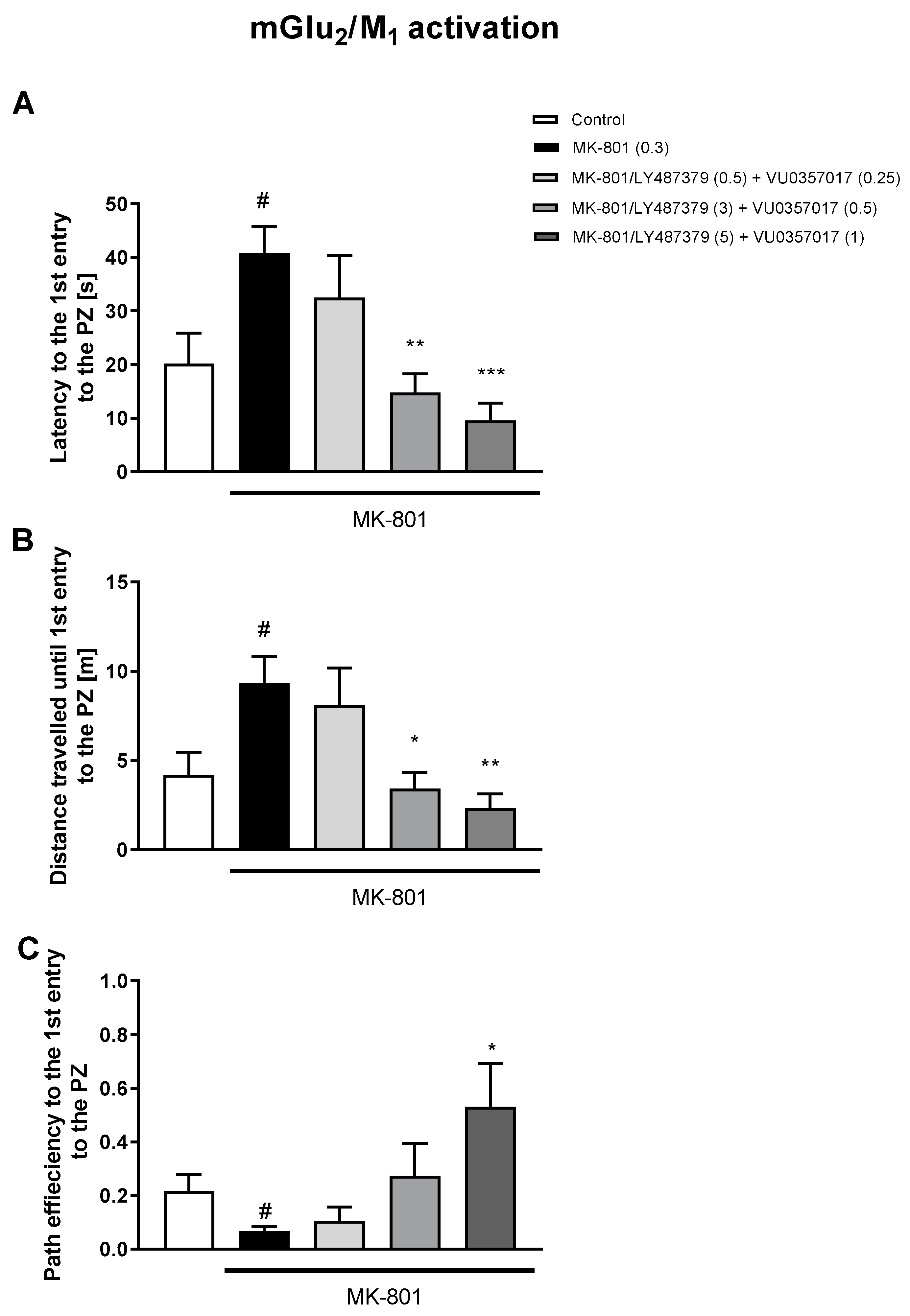
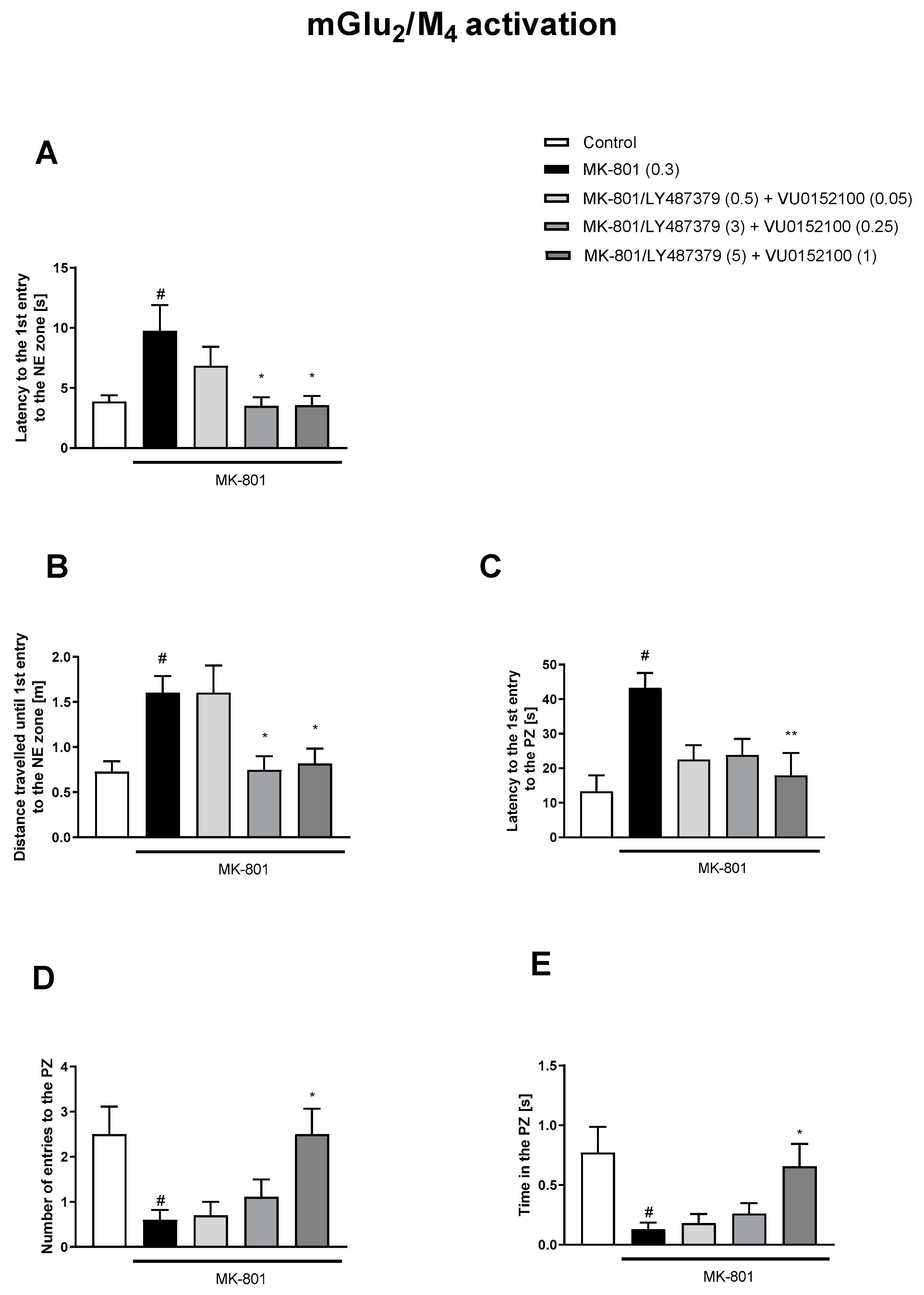
3.1. Morris Water Maze
3.1.1. Acquisition Phase
3.1.2. Retention Trial
3.2. GluN2B Expression
3.3. cGMP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kahn, R.S.; Sommer, I.E.; Murray, R.M.; Meyer-Lindenberg, A.; Weinberger, D.R.; Cannon, T.D.; O’Donovan, M.; Correll, C.U.; Kane, J.M.; van Os, J.; et al. Schizophrenia. Nat. Rev. Dis. Prim. 2015, 1, 15067. [Google Scholar] [CrossRef]
- Bon, C.L.M.; Garthwaite, J. On the role of nitric oxide in hippocampal long-term potentiation. J. Neurosci. 2003, 23, 1941–1948. [Google Scholar] [CrossRef] [PubMed]
- Danysz, W.; Zajaczkowski, W.; Parsons, C.G. Modulation of learning processes by ionotropic glutamate receptor ligands. Behav. Pharmacol. 1995, 6, 455–474. [Google Scholar] [CrossRef]
- Manahan-Vaughan, D.; Ngomba, R.T.; Storto, M.; Kulla, A.; Catania, M.V.; Chiechio, S.; Rampello, L.; Passarelli, F.; Capece, A.; Reymann, K.G.; et al. An increased expression of the mGlu5 receptor protein following LTP induction at the perforant path–dentate gyrus synapse in freely moving rats. Neuropharmacology 2003, 44, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, N.; Gerber, U.; Ster, J. Activation of Group II Metabotropic Glutamate Receptors Promotes LTP Induction at Schaffer Collateral-CA1 Pyramidal Cell Synapses by Priming NMDA Receptors. J. Neurosci. 2016, 36, 11521–11531. [Google Scholar] [CrossRef]
- Luo, L.; Chen, W.-H.; Wang, M.; Zhu, D.-M.; She, J.-Q.; Ruan, D.-Y. Modulation of long-term potentiation by individual subtypes of muscarinic acetylcholine receptor in the rat dentate gyrus. Hippocampus 2008, 18, 989–995. [Google Scholar] [CrossRef]
- Shinoe, T.; Matsui, M.; Taketo, M.M.; Manabe, T. Modulation of Synaptic Plasticity by Physiological Activation of M 1 Muscarinic Acetylcholine Receptors in the Mouse Hippocampus. J. Neurosci. 2005, 25, 11194–11200. [Google Scholar] [CrossRef]
- Fernández de Sevilla, D.; Núñez, A.; Buño, W. Muscarinic Receptors, from Synaptic Plasticity to its Role in Network Activity. Neuroscience 2021, 456, 60–70. [Google Scholar] [CrossRef]
- Patil, S.T.; Zhang, L.; Martenyi, F.; Lowe, S.L.; Jackson, K.A.; Andreev, B.V.; Avedisova, A.S.; Bardenstein, L.M.; Gurovich, I.Y.; Morozova, M.A.; et al. Activation of mGlu2/3 receptors as a new approach to treat schizophrenia: A randomized Phase 2 clinical trial. Nat. Med. 2007, 13, 1102–1107. [Google Scholar] [CrossRef]
- Kinon, B.J.; Zhang, L.; Millen, B.A.; Osuntokun, O.O.; Williams, J.E.; Kollack-Walker, S.; Jackson, K.; Kryzhanovskaya, L.; Jarkova, N. A multicenter, inpatient, phase 2, double-blind, placebo-controlled dose-ranging study of LY2140023 monohydrate in patients with DSM-IV schizophrenia. J. Clin. Psychopharmacol. 2011, 31, 349–355. [Google Scholar] [CrossRef]
- Downing, A.M.; Kinon, B.J.; Millen, B.A.; Zhang, L.; Liu, L.; Morozova, M.A.; Brenner, R.; Rayle, T.J.; Nisenbaum, L.; Zhao, F.; et al. A Double-Blind, Placebo-Controlled Comparator Study of LY2140023 monohydrate in patients with schizophrenia. BMC Psychiatry 2014, 14, 351. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, V.L.; Baygani, S.K.; Kinon, B.J.; Krikke-Workel, J.O. A short-term, multicenter, placebo-controlled, randomized withdrawal study of a metabotropic glutamate 2/3 receptor agonist using an electronic patient-reported outcome device in patients with schizophrenia. J. Clin. Psychopharmacol. 2014, 34, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Kinon, B.J.; Millen, B.A.; Zhang, L.; McKinzie, D.L. Exploratory analysis for a targeted patient population responsive to the metabotropic glutamate 2/3 receptor agonist pomaglumetad methionil in schizophrenia. Biol. Psychiatry 2015, 78, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Nisenbaum, L.K.; Downing, A.M.; Zhao, F.; Millen, B.A.; Munsie, L.; Kinon, B.J.; Adams, D.H.; Gomez, J.C.; Penny, M.A. Serotonin 2A Receptor SNP rs7330461 Association with Treatment Response to Pomaglumetad Methionil in Patients with Schizophrenia. J. Pers. Med. 2016, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Cieślik, P.; Domin, H.; Chocyk, A.; Gruca, P.; Litwa, E.; Płoska, A.; Radulska, A.; Pelikant-Małecka, I.; Brański, P.; Kalinowski, L.; et al. Simultaneous activation of mGlu2 and muscarinic receptors reverses MK-801-induced cognitive decline in rodents. Neuropharmacology 2020, 174, 107866. [Google Scholar] [CrossRef]
- Cieślik, P.; Radulska, A.; Pelikant-Małecka, I.; Płoska, A.; Kalinowski, L.; Wierońska, J.M. Reversal of MK-801-Induced Disruptions in Social Interactions and Working Memory with Simultaneous Administration of LY487379 and VU152100 in Mice. Int. J. Mol. Sci. 2019, 20, 2781. [Google Scholar] [CrossRef] [PubMed]
- Levey, A.I. Immunological localization of m1-m5 muscarinic acetylcholine receptors in peripheral tissues and brain. Life Sci. 1993, 52, 441–448. [Google Scholar] [CrossRef]
- Levey, A.I.; Kitt, C.A.; Simonds, W.F.; Price, D.L.; Brann, M.R. Identification and localization of muscarinic acetylcholine receptor proteins in brain with subtype-specific antibodies. J. Neurosci. Off. J. Soc. Neurosci. 1991, 11, 3218–3226. [Google Scholar] [CrossRef]
- Levey, A.I.; Edmunds, S.M.; Koliatsos, V.; Wiley, R.G.; Heilman, C.J. Expression of m1-m4 muscarinic acetylcholine receptor proteins in rat hippocampus and regulation by cholinergic innervation. J. Neurosci. Off. J. Soc. Neurosci. 1995, 15, 4077–4092. [Google Scholar] [CrossRef] [PubMed]
- Correll, C.U.; Angelov, A.S.; Miller, A.C.; Weiden, P.J.; Brannan, S.K. Safety and tolerability of KarXT (xanomeline–trospium) in a phase 2, randomized, double-blind, placebo-controlled study in patients with schizophrenia. Schizophrenia 2022, 8, 109. [Google Scholar] [CrossRef]
- Sauder, C.; Allen, L.A.; Baker, E.; Miller, A.C.; Paul, S.M.; Brannan, S.K. Effectiveness of KarXT (xanomeline-trospium) for cognitive impairment in schizophrenia: Post hoc analyses from a randomised, double-blind, placebo-controlled phase 2 study. Transl. Psychiatry 2022, 12, 491. [Google Scholar] [CrossRef]
- Shekhar, A.; Potter, W.Z.; Lightfoot, J.; Lienemann, J.; Dubé, S.; Mallinckrodt, C.; Bymaster, F.P.; McKinzie, D.L.; Felder, C.C. Selective muscarinic receptor agonist xanomeline as a novel treatment approach for schizophrenia. Am. J. Psychiatry 2008, 165, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Cieślik, P.; Radulska, A.; Burnat, G.; Kalinowski, L.; Wierońska, J.M. Serotonergic–muscarinic interaction within the prefrontal cortex as a novel target to reverse schizophrenia-related cognitive symptoms. Int. J. Mol. Sci. 2021, 22, 8612. [Google Scholar] [CrossRef] [PubMed]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Maei, H.R.; Zaslavsky, K.; Teixeira, C.M.; Frankland, P.W. What is the Most Sensitive Measure of Water Maze Probe Test Performance? Front. Integr. Neurosci. 2009, 3, 4. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef]
- Sałat, K.; Podkowa, A.; Mogilski, S.; Zaręba, P.; Kulig, K.; Sałat, R.; Malikowska, N.; Filipek, B. The effect of GABA transporter 1 (GAT1) inhibitor, tiagabine, on scopolamine-induced memory impairments in mice. Pharmacol. Rep. 2015, 67, 1155–1162. [Google Scholar] [CrossRef]
- Pochwat, B.; Rafało-Ulińska, A.; Domin, H.; Misztak, P.; Nowak, G.; Szewczyk, B. Involvement of extracellular signal-regulated kinase (ERK) in the short and long-lasting antidepressant-like activity of NMDA receptor antagonists (zinc and Ro 25-6981) in the forced swim test in rats. Neuropharmacology 2017, 125, 333–342. [Google Scholar] [CrossRef]
- Ennaceur, A. One-trial object recognition in rats and mice: Methodological and theoretical issues. Behav. Brain Res. 2010, 215, 244–254. [Google Scholar] [CrossRef]
- Swerdlow, N.R.; Geyer, M.A. Using an animal model of deficient sensorimotor gating to study the pathophysiology and new treatments of schizophrenia. Schizophr. Bull. 1998, 24, 285–301. [Google Scholar] [CrossRef] [PubMed]
- Wenk, G.L. Assessment of Spatial Memory. Curr. Protoc. Toxicol. 1999, 11.3.1–11.3.18. [Google Scholar] [CrossRef]
- Wenk, G.L. Assessment of Spatial Memory Using the T Maze. Curr. Protoc. Neurosci. 1998, 4, 8.5B.1–8.5A.7. [Google Scholar] [CrossRef]
- Blodgett, H.C.; McCutchan, K.; Mathews, R. Spatial learning in the T-maze: The influence of direction, turn, and food location. J. Exp. Psychol. 1949, 39, 800–809. [Google Scholar] [CrossRef]
- Kavraal, S.; Oncu, S.K.; Bitiktas, S.; Artis, A.S.; Dolu, N.; Gunes, T.; Suer, C. Maternal intake of Omega-3 essential fatty acids improves long term potentiation in the dentate gyrus and Morris water maze performance in rats. Brain Res. 2012, 1482, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Aou, S.; Li, X.-L.; Li, A.-J.; Oomura, Y.; Shiraishi, T.; Sasaki, K.; Imamura, T.; Wayner, M.J. Orexin-A (hypocretin-1) impairs Morris water maze performance and CA1-Schaffer collateral long-term potentiation in rats. Neuroscience 2003, 119, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- McNamara, R.K.; DePape, G.E.; Skelton, R.W. Differential effects of benzodiazepine receptor agonists on hippocampal long-term potentiation and spatial learning in the Morris water maze. Brain Res. 1993, 626, 63–70. [Google Scholar] [CrossRef]
- Sabo, S.L.; Lahr, J.M.; Offer, M.; Weekes, A.; Sceniak, M.P. GRIN2B-related neurodevelopmental disorder: Current understanding of pathophysiological mechanisms. Front. Synaptic. Neurosci. 2023, 14, 1090865. [Google Scholar] [CrossRef] [PubMed]
- Platzer, K.; Lemke, J.R. GRIN2B-Related Neurodevelopmental Disorder. 2018 May 31 [updated 2021 Mar 25]. In GeneReviews® [Internet]; Adam, M.P., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993–2023. [Google Scholar]
- Myers, S.J.; Yuan, H.; Kang, J.Q.; Tan, F.C.K.; Traynelis, S.F.; Low, C.M. Distinct roles of GRIN2A and GRIN2B variants in neurological conditions. F1000Research 2019, 20, 1940. [Google Scholar] [CrossRef] [PubMed]
- Kantrowitz, J.T.; Javitt, D.C. N-methyl-d-aspartate (NMDA) receptor dysfunction or dysregulation: The final common pathway on the road to schizophrenia? Brain. Res. Bull. 2010, 83, 108–121. [Google Scholar] [CrossRef]
- Gardoni, F.; Mauceri, D.; Malinverno, M.; Polli, F.; Costa, C.; Tozzi, A.; Siliquini, S.; Picconi, B.; Cattabeni, F.; Calabresi, P.; et al. Decreased NR2B subunit synaptic levels cause impaired long-term potentiation but not long-term depression. J. Neurosci. 2009, 29, 669–677. [Google Scholar] [CrossRef]
- Wood, P.L.; Rao, T.S. A review of in vivo modulation of cerebellar cGMP levels by excitatory amino acid receptors: Role of NMDA, quisqualate and kainate subtypes. Prog. Neuropsychopharmacol. Biol. Psychiatry 1991, 15, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Wincott, C.M.; Kim, S.; Titcombe, R.F.; Tukey, D.S.; Girma, H.K.; Pick, J.E.; DeVito, L.M.; Hofmann, F.; Hoeffer, C.; Ziff, E.B. Spatial memory deficits and motor coordination facilitation in cGMP-dependent protein kinase type II-deficient mice. Neurobiol. Learn. Mem. 2013, 99, 32–37. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inactive doses | ||
| LY487379 | ||
| VU0357017 | 0.25 mg/kg | 0.5 mg/kg |
| VU0152100 | 0.05 mg/kg | 0.5 mg/kg |
| VU0238429 | 1 mg/kg | 0.5 mg/kg |
| Intermediate doses | ||
| LY487379 | ||
| VU0357017 | 0.5 mg/kg | 3 mg/kg |
| VU0152100 | 0.25 mg/kg | 3 mg/kg |
| VU0238429 | 5 mg/kg | 3 mg/kg |
| Highest doses | ||
| LY487379 | ||
| VU0357017 | 1 mg/kg | 5 mg/kg |
| VU0152100 | 1 mg/kg | 5 mg/kg |
| VU0238429 | 20 mg/kg | 5 mg/kg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wierońska, J.M.; Cieślik, P.; Burnat, G.; Kalinowski, L. Activation of Metabotropic Glutamate Receptor (mGlu2) and Muscarinic Receptors (M1, M4, and M5), Alone or in Combination, and Its Impact on the Acquisition and Retention of Learning in the Morris Water Maze, NMDA Expression and cGMP Synthesis. Biomolecules 2023, 13, 1064. https://doi.org/10.3390/biom13071064
Wierońska JM, Cieślik P, Burnat G, Kalinowski L. Activation of Metabotropic Glutamate Receptor (mGlu2) and Muscarinic Receptors (M1, M4, and M5), Alone or in Combination, and Its Impact on the Acquisition and Retention of Learning in the Morris Water Maze, NMDA Expression and cGMP Synthesis. Biomolecules. 2023; 13(7):1064. https://doi.org/10.3390/biom13071064
Chicago/Turabian StyleWierońska, Joanna M., Paulina Cieślik, Grzegorz Burnat, and Leszek Kalinowski. 2023. "Activation of Metabotropic Glutamate Receptor (mGlu2) and Muscarinic Receptors (M1, M4, and M5), Alone or in Combination, and Its Impact on the Acquisition and Retention of Learning in the Morris Water Maze, NMDA Expression and cGMP Synthesis" Biomolecules 13, no. 7: 1064. https://doi.org/10.3390/biom13071064
APA StyleWierońska, J. M., Cieślik, P., Burnat, G., & Kalinowski, L. (2023). Activation of Metabotropic Glutamate Receptor (mGlu2) and Muscarinic Receptors (M1, M4, and M5), Alone or in Combination, and Its Impact on the Acquisition and Retention of Learning in the Morris Water Maze, NMDA Expression and cGMP Synthesis. Biomolecules, 13(7), 1064. https://doi.org/10.3390/biom13071064