Abstract
Both sensory neurons and immune cells, albeit at markedly different levels, express the vanilloid (capsaicin) receptor, Transient Receptor Potential, Vanilloid-1 (TRPV1). Activation of TRPV1 channels in sensory afferent nerve fibers induces local effector functions by releasing neuropeptides (most notably, substance P) which, in turn, trigger neurogenic inflammation. There is good evidence that chronic activation or inactivation of this inflammatory pathway can modify tumor growth and metastasis. TRPV1 expression was also demonstrated in a variety of mammalian immune cells, including lymphocytes, dendritic cells, macrophages and neutrophils. Therefore, the effects of TRPV1 agonists and antagonists may vary depending on the prominent cell type(s) activated and/or inhibited. Therefore, a comprehensive understanding of TRPV1 activity on immune cells and nerve endings in distinct locations is necessary to predict the outcome of therapies targeting TRPV1 channels. Here, we review the neuro-immune modulation of cancer growth and metastasis, with focus on the consequences of TRPV1 activation in nerve fibers and immune cells. Lastly, the potential use of TRPV1 modulators in cancer therapy is discussed.
1. Introduction: TRPV1, Capsaicin, and Neurogenic Inflammation
Natural products provide a window of opportunity to discover novel molecular pathways. A prominent example is capsaicin, the pungent ingredient in hot chili peppers. Hot pepper is indeed “hot” because capsaicin and noxious heat activate the very same receptor on sensory neurons [1], now known as Transient Receptor Potential, Vanilloid-1 (TRPV1) [2]. TRPV1 is the founding member of the family of temperature-sensitive TRP channels, collectively referred to as “thermoTRPs” [3]. Combined, thermoTRPs cover a broad range of temperatures, from noxious hot to noxious cold [4]. The central role of TRPV1 in noxious heat sensation was highlighted by the burn injuries that participants of clinical trials with TRPV1 antagonists suffered as adverse effect [5]. The 2021 Nobel Prize in Medicine and Physiology was awarded to David Julius (shared with Ardem Patapoutian), in part for discovering the molecular mechanisms of heat sensation that started with the molecular cloning of TRPV1 in 1997 [1].
Capsaicin represents an area of sensory pharmacology intimately familiar to those of us who enjoy eating hot, spicy food. Capsaicin causes a burning sensation on the tongue that dissipates (“desensitizes”) upon repeated challenge [6,7]. Animals desensitized to capsaicin are also unresponsive to various unrelated chemical and physical stimuli [8]. The therapeutic potential of capsaicin desensitization was recognized early. High-dose capsaicin patches [9] and site-specific injections [10] are already in clinical practice to relieve chronic neuropathic pain. The molecular mechanisms of capsaicin desensitization are, however, largely unknown.
TRPV1 is predominantly expressed on sensory neurons. Indeed, a whole subdivision of primary sensory neurons is named after their unique sensitivity to capsaicin [11,12,13].
Capsaicin-sensitive (TRPV1-expressing) sensory neurons are bipolar cells with cell bodies in sensory (dorsal root and trigeminal) ganglia. A subset of vagal neurons with somata in nodose ganglia also express TRPV1 [14]. Generally speaking, TRPV1-expressing neurons detect harmful information in the periphery and convey this information into the central nervous system (CNS). Importantly, these neurons not only detect noxious stimuli as an afferent function, but they also produce and release pro-inflammatory neuropeptides from their peripheral endings as a local efferent function [15]. Notable neuropeptides include substance P (SP) and calcitonin gene-related peptide (CGRP). These neuropeptides trigger the biochemical cascade known as neurogenic inflammation [16,17]. The role of neurogenic inflammation in various disease states (from migraine to diabetes) is well beyond the scope of this review (interested readers are referred to [17]. Here, it suffices to mention that neurogenic inflammation affects carcinogenesis [18], tumor growth [19], and metastasis formation [20].
Albeit at much lower densities than on capsaicin-sensitive sensory neurons, TRPV1 is expressed in various non-neuronal cells, including immune cells [21]. This implies that TRPV1 agonists (or antagonists) may produce complex and unpredictable effects by simultaneously acting on neuronal and non-neuronal targets. Moreover, neuropeptides released from capsaicin-sensitive nerve endings can also influence the activity of both cancer cells and immune cells [22]. For instance, SP may enhance anti-tumoral activity by triggering dendritic cell migration [23] and augmenting antigen-induced lymphocyte proliferation [24]. Conversely, SP can stimulate the proliferation of cancers that express the SP receptor, NK-1 [25]. Take glioblastoma cells as an example: they express both TRPV1 [26] and NK-1 receptors [27]. On the one hand, capsaicin may induce apoptosis in this tumor by activating TRPV1 [26]. On the other hand, capsaicin may accelerate tumor growth by liberating SP from sensory nerves that, in turn, stimulates trophic NK-1 receptors in cancer cells [26]. Indeed, NK-1 receptor antagonists were proposed to represent a new therapeutic approach in glioblastoma patients [28].
2. Neurogenic Inflammation and Tumor Promotion in the Skin
165 years ago, Robert Virchow in his “Reiztheorie” (inflammation theory) postulated a causative connection between inflammation and cancer based on his observation that white blood cells are found in increased numbers in the vicinity of tumors [29]. In support of this theory, the first experimental proof was provided by Yamagiwa and Ichikawa by painting coal tar on the ear of rabbits [30]. The cancer causing agent in coal tar was later identified as 7,12-dimethylbenz[a]anthracene (DMBA). DMBA as a single carcinogen can induce the formation of epithelial tumors when applied to the back skin of mice [31,32]. The tumor-causing dose of DMBA, however, can be greatly reduced if croton oil were applied subsequent to the application of a dose of DMBA too low to induce tumors by itself [32,33]. In this classic two-stage carcinogenesis experiment, DMBA is thought to initiate the tumor development by mutagenic activity, whereas croton oil is believed to promote the formation of tumors by maintaining chronic inflammation.
Based on this seminal observation, the mouse ear erythema assay was routinely used to detect tumor-promoting compounds in natural product extracts. In fact, a close relationship was postulated between the inflammatory and tumor promoting activities of natural products [34]. The prototypical tumor promoter, 12-O-tetradecanoylphorbol-13-acetate (TPA), is indeed a very potent inflammatory agent in the mouse skin [35]. TPA is the main ingredient in croton oil, the seed oil of the Euphorbiaceae Croton tiglium. Using the mouse ear erythema assay, another extremely irritant compound was discovered in the soap of Euphorbia resinifera Berg [36], named after its source: resiniferatoxin (RTX). Unlike TPA, RTX does not promote the formation of tumors [37]. Although TPA and RTX use completely different molecular pathways to cause inflammation [38], protein kinase-C (PKC) [38,39] and TRPV1 [40], respectively, both pathways participate in neurogenic inflammation [41].
In CD1 mice (a mouse strain particularly sensitive to tumor promotion by TPA), desensitization by RTX of the neurogenic inflammatory response completely prevented the early (3 h) erythema and edema response to TPA, and partially blocked the edema at later times (up to 24 h) [41]. However, RTX desensitization had no measurable effect on TPA-induced epidermal hyperplasia and/or tumor formation [41]. These findings imply that tumor promoters indeed do evoke neurogenic inflammation, but it is the non-neurogenic component of their inflammatory reaction that promote the formation of skin tumors. A similar conclusion was reached in mouse strains with different susceptibilities to phorbol ester tumor promotion [18].
Human keratinocytes express functional TRPV1 [42]. Activation by capsaicin of keratinocyte TRPV1 triggers the release of the pro-inflammatory mediators, interleukin-8 and prostaglandin-E2 [43]. This observation raises the question whether capsaicin can promote skin carcinogenesis via non-neurogenic inflammation. In the back skin of ICR mice, topical capsaicin (a single application of 10 µmol) followed by TPA twice a week resulted in no increase in the number of skin tumors compared to solvent controls [44]. Following initiation by DMBA, RTX did not promote the formation of skin tumors either [37]. Last, no difference was noted in skin tumor development between wild-type or solvent control animals and those whose TRPV1 had been ablated by genetic manipulation (TRPV1 KO mice) [45] or neonatal RTX administration (A. Szallasi and P.M. Blumberg, unpublished observations).
Combined, these observations imply that (1) RTX by itself is not a complete carcinogen and (2) the elimination by RTX of the neurogenic inflammatory response does not modify the response in the two-stage carcinogenesis model.
3. Neuro-Immune Regulation of Cancer
Interaction between the immune system and the CNS is crucial for maintaining homeostasis. Abnormal activation and/or disturbance of this neuro-immune interaction may have serious pathological consequences [46]. Chronic, stress-induced alterations in immune responses may have detrimental consequences for cancer patients [47]. For example, chronic or disproportionate stress may suppress protective immunity, induce or exacerbate chronic inflammation, or enhance immuno-suppressive mechanisms [47,48]. Early psychosocial intervention may significantly reduce cancer mortality [49].
The nervous system plays a crucial role in carcinogenesis, particularly through altering anti-cancer immunity (Figure 1) [50]. Multiple components of neuro-immune interactions exist, including the adrenergic sympathetic nerves, the hypothalamic–pituitary–adrenal axis, and the parasympathetic nerves [51,52,53]. The sympathetic nervous system was shown to promote the formation of solid tumors [54]. Adrenergic sympathetic nerves directly innervate all primary and secondary immune organs [55], and the excessive activation of these nerves can suppress anti-tumoral immunity. By contrast, the activation of the parasympathetic system was shown to exert anti-tumoral and anti-metastatic effects via enhancing anti-tumoral immunity [49,56]. Specifically, activation of parasympathetic nerves can downregulate the expression of programmed cell death protein-1 (PD-1) on CD4+ tumor-infiltrating T-lymphocytes, and improve the expression of interferon-γ, thus enhancing anti-tumor immunity [56].
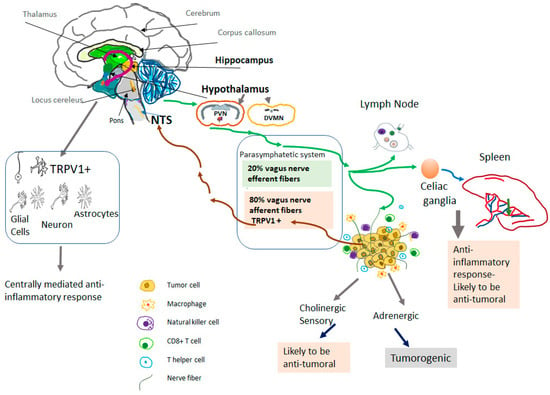
Figure 1.
TRPV1-mediated neuro-immune response to cancer. TRPV1 is found primarily in afferent sensory nerve fibers. TRPV1 is also expressed in the central nervous system, mainly in areas that are responsible for controlling the autonomic output to the visceral organs. Specifically, TRPV1 channels were demonstrated in the paraventricular nucleus of the hypothalamus (PVN) and the dorsal motor nucleus of the vagus (DMVN). Besides the hypothalamus, TRPV1 is expressed in the hippocampus and midbrain and the expression is not only found in neurons but also in glial cells and astrocytes. Direct central activity of TRPV1 agonists might be responsible for a systemic anti-inflammatory response. In addition to the activation of vagal TRPV1+ afferent fibers, it may induce a systemic anti-inflammatory response that might be responsible for the anti-tumoral effects of vagal activity. Vagal afferent nerve fibers may activate intra neuronal nerve fibers. NTS-Nucleus Tractus Solitarius.
The cholinergic parasympathetic system Innervates the majority of visceral organs directly via the vagal nerve or through the celiac ganglia (Figure 1). The parasympathetic efferent fibers terminate in the sympathetic celiac ganglia [57,58]. Approximately 80% of nerve fibers in the vagal nerve are afferent (sensory) fibers that carry sensory information from the visceral organs to the brain [59]. In turn, efferent fibers of the vagal nerve transmit central neuronal response into the visceral tissues as well as innate and adaptive immune cells [60,61]. The vagus nerve mediates neuro-immune interactions in cancer [62], in addition to its fundamental physiological regulatory functions in controlling circulation, temperature, digestion, respiration, and the immune response [63,64,65,66].
4. TRPV1-Positive Vagal Afferent Neurons and Carcinogenesis
Vagal visceral afferent nerve endings express TRPV1 [14], a non-selective cation channel allosterically modulated and/or activated by a range of thermal, mechanical and chemical stimuli [67,68]. TRPV1 activation may mediate the immune regulatory role of the vagus nerve. Peripheral activation of vagal afferent fibers carries the information to the nucleus tractus solitarii in the brainstem. In return, activated efferent vagus nerve fibers inhibit the production of peripheral pro-inflammatory cytokines through a nicotinic anti-inflammatory pathway [69,70]. In accord, in a concanavalin-A model of hepatitis, the afferent vagal nerve activity is required for the anti-inflammatory response in the liver of treated animals [71].
In experimental breast cancer, afferent vagus nerve fibers mediate the anti-metastatic effect of vagal activity. Chemical (by systemic capsaicin) or surgical vagotomy results in enhanced metastasis formation [20,72,73,74]. Conversely, activation of the vagus nerve by semapimod (a non-steroidal inhibitor of inflammatory cytokine production) decreases breast cancer metastasis [75]. Moreover, vagal stimulation reduces the levels of pro-inflammatory cytokines (e.g., interleukin-1β and tumor necrosis factor-α), and enhances anti-tumor immunity by activating cytotoxic CD8+ T cells and NK cells [76]. Vagal stimulation also inhibits the accumulation and activity of myeloid-derived suppressor cells in breast cancer [77]. Upon activation, afferent vagal fibers release SP, which, in turn, enhances anti-tumoral immunity by increasing the cytotoxic effect of lymphokine-activated NK cells [78]. SP also increases interleukin-12 secretion and promotes the maturation of dendritic cells [79].
In metastatic breast carcinoma, continuous SP treatment by implantable pumps markedly enhances the therapeutic efficacy of radiotherapy by increasing the anti-tumoral immune response [80]. Hence, stimulation of vagal afferent nerve fibers via TRPV1 activation may inhibit chronic inflammatory responses and enhance anti-tumoral immunity, and thus establish an anti-tumoral environment. In keeping with this hypothesis, olvanil, a non-pungent vanilloid agonist that activates TRPV1 more slowly than capsaicin does [81,82], was shown to suppress metastasis of breast carcinoma without altering the rate of primary tumor growth [83]. Olvanil also decreases the systemic inflammatory response and enhances anti-tumoral immunity at a dose that primarily activates the nerve fibers [83].
5. Systemic Activation of Neuronal TRPV1 in Cancer
TRPV1 is expressed in both sensory nerve fibers and non-neuronal tissues, though expression levels differ markedly between neuronal and non-neuronal cells. Nanomolar concentrations of capsaicin (and related TRPV1 agonists) are sufficient to activate nerve fibers [84]. By contrast, over 30 μM capsaicin concentrations are required for TRPV1 activation in isolated immune cells [85]. Hence, selectivity of TRPV1 agonists at lower concentration is much higher for sensory nerve fibers. For example, the affinity of capsaicin for the human neuronal TRPV1 channel ranges from 7 to 30 nM as determined through electrophysiological studies [86,87]. By contrast, capsaicin stimulates human monocytes via TRPV1 at concentrations exceeding 100 µM [85]. Therefore, at lower doses TRPV1 agonists are likely to affect mainly the peripheral nervous system.
TRPV1 is thought to play an important role in the pathology of autoimmune neuritis [88]. In rats, low doses of orally administered capsaicin ameliorated the symptoms of autoimmune neuritis [89]: given at a dose of 50 μg/day, corresponding to consuming one or two chili peppers per day in humans, dietary capsaicin was already effective [89]. The beneficial effect of capsaicin in autoimmune neuritis was attributed to the release of sensory neuropeptides, such as CGRP and SP, which in turn can inhibit macrophages. A similar protective effect of low capsaicin doses was reported in an autoimmune diabetes model [90]. In keeping with the animal experiments, in a large-scale prospective cohort study, daily consumption of spicy (chili-based) food was inversely associated with all-cause mortality, and specific-cause mortality including cancer [91].
The TRPV1-mediated anti-tumoral effect of capsaicin can be attributed to its anti-inflammatory actions. This assumption is supported by Trpv1 gene knockout studies. In a mouse model of allergic contact dermatitis, exacerbated inflammatory reaction was observed in the TRPV1 null animals [92]. In this model, TRPV1 deficiency is associated with the upregulation of pro-inflammatory cytokines, increased infiltration of macrophages, and simultaneous inflammation [92]. Similarly, more severe inflammatory response occurs in TRPV1-null mice challenged with LPS to induce shock and organ damage (renal and hepatic) [93]. Moreover, compared to wild-type littermates, TRPV1-null mice exhibit more severe LPS-induced sepsis [94], and more often develop colitis-associated cancer [95]. In accord, small-molecule TRPV1 agonists were shown to reduce inflammation in sepsis models [96,97,98].
6. Role of the Central Nervous System (CNS) in the Anti-Inflammatory Effects of TRPV1 Agonists
There is growing evidence that TRPV1 agonists may target CNS nuclei, such as the hypothalamus, to modulate inflammatory responses. TRPV1 channels are expressed in the brain, particularly in areas that are responsible for controlling the autonomic output to the visceral organs (Figure 1). For example, in rat brain, TRPV1 channel expression was demonstrated in the paraventricular nucleus of the hypothalamus and the dorsal motor nucleus of the vagus [99]. Using a viral tracer, TRPV1 receptors were also found in liver-related pre-autonomic neurons [100]. In addition to the hypothalamus, TRPV1 is expressed in the hippocampus and midbrain [101,102]. This restricted TRPV1 expression in the brain is conserved across species (rat, monkey, and human) [101,102,103].
TRPV1 can modulate synaptic transmission via both pre- and postsynaptic mechanisms. Depending on the location, TRPV1 may participate in various (sometimes opposing) functions [104]. Expressed pre-synaptically on afferents to the locus ceruleus, TRPV1 potentiates the release of glutamate and adrenaline/noradrenaline [105]. By contrast, TRPV1 suppresses the excitatory transmission in the dentate gyrus via a postsynaptic mechanism [106,107,108]. Importantly, TRPV1 is expressed in microglia and astrocytes [109,110]. In fact, microglial TRPV1 is thought to mediate some central effects of TRPV1 agonists, such as the enhancement of neuronal glutamatergic transmission [111]. In the substantia nigra, TRPV1 activation in astrocytes produces ciliary neurotrophic factor: this rescues nigral dopaminergic neurons in experimental Parkinson disease [112].
“Endovanilloids” (endogenous TRPV1 agonists generated in the CNS [113]) may exert potent anti-inflammatory activity. In models of endotoxemia and polymicrobial sepsis, the potent endovanilloids, N-arachidonoyl dopamine (NADA) and N-oleoyl dopamine, reduced inflammatory responses [114,115,116]. These endovanilloid effects were mediated by TRPV1 expressed in the CNS and not in the peripheral nervous system [114].
Capsaicin is highly lipophilic: it can pass the blood–brain barrier easily and has a longer half-life in the brain compared to peripheral tissues [117]. Therefore, the anti-tumoral effects of TRPV1 agonists might partly be due to activation of TRPV1 in CNS nuclei, such as the hypothalamus, that regulate vagal activity (Figure 1).
Although the local anti-tumoral effects of TRPV1 agonists are well-documented, the systemic effects of TRPV1 agonists on carcinogenesis and metastasis vary depending on the model. For instance, TRPV1 activation may actually accelerate cancer growth [20,118]. Below, the possible causes of these discrepancies are discussed, focusing on non-neuronal TRPV1 activation, as well as the neuronal components of the tumor microenvironment.
7. Innervation of the Cancer
Once solid tumors were thought to lack innervation [119]. This is now known to not be true. In fact, tumor cells can produce their own innervation, a process termed by Palm and Entschladen as “neo-neurogenesis” [120]. Palm and Entschladen also introduced the concept of “neuro-neoplastic synapsis”, defining the structures that enable interaction between peripheral neurons and tumor cells [119,120].
Further studies documented the presence of a heterogeneous group of nerve fibers within tumor tissue that is often associated with the aggressiveness [121,122]. In breast and prostate cancers, increased nerve densities are associated with more aggressive tumor phenotype and poor patient survival [121,123,124]. Cancer cells can produce growth factors, such as nerve growth factor (NGF), and stimulate the growth of new axons into the tumor [125,126]. The type of nerve fibers might be critical in determining the aggressiveness of the cancer. For example, in a chemically induced breast tumor model, sympathetic nerve activation may accelerate cancer growth and progression, while selective parasympathetic nerve fiber activation may conversely inhibit tumor growth [56]. A strong connection between the autonomic nervous system and a tumor was also demonstrated in a mouse model of breast cancer [124]. Genetic depletion of intra-tumoral sympathetic nerves inhibits breast cancer growth [56,127]. To promote growth and metastasis, cancer cells can manipulate the nervous systems by stimulating the growth of new sympathetic nerve fibers into the tumor tissue, and by trans-differentiating the phenotype of sensory neurons into an adrenergic type [128,129]. Indeed, in patients with recurrent breast cancer, the sympathetic nerve density was higher and the parasympathetic nerve density was lower compared to patients without recurrent breast cancer [56].
Among cancer tissue and the CNS, indirect connections were described in several tumor models, including fibrosarcoma [130] and melanoma [131]. Recent studies also show a direct link between cancer and the CNS. For example, using electrical nerve stimulation to generate evoked potentials within the tumor, a direct neural connection was found between 4T1 breast tumors and the brain via the vagus nerve [124]. The electrical activity in the tumor was eliminated by chemical sympathectomy [124]. In another study, the presence of sympathetic nerves within the periphery of breast tumor of 4T1 model was described [132].
These findings allow two important conclusions to be drawn. First, activation of vagal fibers can stimulate intra-tumoral sympathetic fibers, leading to an unwanted sympathetic activity within the tumor microenvironment. Second, identifying the factors that drive trans-differentiation of the sensory neuronal phenotype to an adrenergic phenotype (that, in turn, can accelerate cancer growth and metastasis [128,129,133]) may open up new therapeutic opportunities in cancer therapy.
Although the formation of new nerve fibers is called ‘neo-neurogenesis” [120], it might be more accurate to call this process “abnormal neo-neurogenesis”, since these new nerve fibers can promote cancer growth [62]. Exploring the type and neurochemistry of these nerve fibers within the tumor microenvironment in relation to tumor grade and aggressiveness may provide novel patient-tailored treatment options. There are few reports focusing on these issues. For example, progenitors that express doublecortin (a neural migration protein) can initiate autonomic adrenergic neurogenesis in prostate cancer [134]. On the other hand, loss of TP53 drives the reprogramming of tumor-innervating sensory nerves into adrenergic neurons, promoting the growth of head-and-neck carcinomas [121,135]. Intra-tumoral growth of sensory nerves might be induced by brain-derived neurotrophic factor (BDNF). BDNF signals through tropomyosin receptor kinase-B (TRKB) receptors on sensory neurons, and recruits these nerves to establish innervation of the ductal tree during mammary gland development [136].
Although indirectly, the level of TRPV1 expression within tumor tissue may estimate the content of sensory neurons. Expression analysis of tumoral TRPV1 has demonstrated an association between TRPV1 expression and immune response. For instance, in cervical carcinoma, CD8+ T-cells were positively correlated, whereas M2-type macrophages were negatively correlated with TRPV1 expression [137]. Furthermore, decreased TRPV1 expression was associated with increased risk of cervical squamous cell carcinoma [137]. In renal cell carcinoma, TRPV1 expression positively correlated with M1-type anti-tumoral macrophages, and negatively correlated with M2-type tumorigenic macrophages [138]. In agreement, low expression of TRPV1 in renal cell carcinoma is associated with poor clinical outcomes [138].
A recent pan-cancer study further cemented the prognostic value of tumoral TRPV1 expression. For example, TRPV1 expression showed a positive correlation with the ratio of immune-stimulatory over immunosuppressive signatures (CD8+ T cell/PD-L1) in five cancer types [139]. Moreover, TRPV1 expression was found to be markedly lower in late-stage (stage III-IV) cancers, and TRPV1 downregulation correlated with worse overall survival [139]. Although these studies did not directly measure the density of TRPV1-positive sensory neurons in cancer tissues, it can be argued that changes in intra-tumoral TRPV1 predominantly reflects changes in TRPV1-positive nerve fibers within the tumor microenvironment.
In general, a loss of cholinergic/peptidergic nerve fibers is considered a bad prognostic factor. However, this may not be the case for all types of malignancies. For example, human melanoma samples were found to have increased TRPV1-expressing neuronal innervation [140]. When co-cultured with B16F10 murine melanoma cells, TRPV1-positive nociceptors directly extended neurites toward the cancer cells [140]. The tumor-invading sensory neurons express more CGRP and Trka, the receptor for NGF. In melanoma, increased CGRP levels were associated with CD8+ T-cell exhaustion, inhibiting anti-tumoral immunity [140]. These results imply that TRPV1-sensitive sensory innervation may enhance melanoma aggressiveness. If this hypothesis holds true, TRPV1 receptor blockers may have a therapeutic value in melanoma. For example, TRPV1 antagonist creams may be applied to suspicious melanocytic lesions to prevent the development of tumorigenic melanoma.
An issue that needs to be considered is the heterogeneous nature of sensory nerve endings. Melanoma drives sensory nerve fibers from adjacent skin that are polymodal and are involved in pain sensation. Visceral afferents are, however, different in that their principal role is to maintain homeostasis of the internal environment. Pain sensation associated with visceral afferents is often poorly defined and projects to somatic structures, such as skin and muscle (a well-known example is cardiac pain that often projects to the shoulder). Hence, not only the possible alteration in phenotype but also the type of sensory neuron from which cancer-invading nerve fibers sprout might be critical.
8. TRPV1: Channel Structure, Epigenetic Regulation and Subcellular Expression
Cryo-electron microscopy and X-ray crystallography have provided important insights into TRPV1 channel structure and function [141,142,143]. In contrast to highly selective cation channels, the selectivity filter of TRPV1 is shallow and dynamic, favoring the influx of larger (e.g., Ca2+) or smaller (e.g., Na+) cations [143]. This explains the long-recognized “limited selectivity for Ca2+” of the TRPV1 channel. Furthermore, TRPV1 has fourfold symmetry with different pore profiles for ligand-bound structures and a vanilloid-binding pocket deep within the membrane bilayer [144]. Resiniferatoxin binding to TRPV1 first opens the intracellular gate, followed by selectivity filter dilation and pore loop rearrangement [144]. This resiniferatoxin-induced conformational “wave” is likely to involve additive structural changes in the channel subdomains [144]. Resiniferatoxin binding to each of these four subunits contributes the same activation energy. The potent small-molecule TRPV1 antagonist, SB-366791, binds to this vanilloid site and acts as an allosteric hTRPV1 inhibitor.
TRPV1 can form functional tetramers both with its splice variants [145] and other TRP channels [146]. TRPV1 splice variants may function as negative modulators of the channel activity. TRPV1/TRPA1 hetero-tetramers are intriguing in that they are shown to possess unique activation profiles: for example, TRPV1 agonists of the home-tetrameric channel may function as antagonists of the hetero-tetrameric channel [147].
Epigenetic regulation of TRPV1 gene expression is an emerging area of research. In rats, histone H3 acetylation of the Trpv1 gene promoter region regulated the expression of TRPV1 protein in sensory neurons [148]. In a murine model of diabetic neuropathy, SUMOylation protects the TRPV1 protein from metabolic damage and thus delays the development of neuropathic pain [149]. Phosphorylation by protein kinase C also regulates the activity of the channel protein [150].
The TRPV1 protein is present both in the cell membrane and in subcellular localizations [151]. It was speculated that activated TRPV1 can be recycled into a subcellular organism [152], which may represent a molecular mechanism of desensitization. It is not impossible either that TRPV1 expressed subcellularly (e.g., in mitochondria or the Golgi apparatus) serves distinct biological functions. Indeed, mitochondrial TRPV1 expression was implicated in the proliferation and apoptosis of chronic myeloid leukemia cells [153].
In sensory neurons, TRPV1 most likely functions in the cell membrane to detect heat, protons and other noxious stimuli [154]. In immune cells and cancer, subcellular TRPV1 may be involved in as-yet unsuspected functions. For example, in human breast cancer “non-classical” TRPV1 expression (in the Golgi area) heralds poor prognosis compared to “classical” expression in the cell membrane [155].
9. Capsaicin Actions in Immune Cells: On- or Off-Target?
The wide distribution of TRPV1 suggests a diverse function beyond noxious heat sensation and pain perception. Therefore, capsaicin actions in non-neuronal cells were hardly unexpected. However, it is important to keep in mind that capsaicin actions are not necessarily TRPV1-mediated [8]. In fact, these non-TRPV1 mediated capsaicin actions had long clouded the existence of a specific “capsaicin receptor.” For example, capsaicin was reported to stimulate or inhibit the activity of enzymes [156], influence membrane fluidity [157], or interact at voltage-gated ion channels [158], just to cite a few examples of non-specific capsaicin actions.
To provide unequivocal evidence that a capsaicin response is on-target, (1) one should demonstrate the presence of functional TRPV1, and (2) prove that the capsaicin action is absent when TRPV1 is non-functional (e.g., pharmacological blockade or genetic inactivation). For instance, if a capsaicin action is on-target (that is, mediated by TRPV1), it should be absent in TRPV1-null animals. This was not always the case in the carcinogenesis studies. For example, in a mouse skin carcinogenesis experiment with DMBA, capsaicin equally promoted the formation of tumors in TRPV1 wild-type and knock-out animals [45].
Many studies rely on showing the presence of TRPV1 protein by immunostains. Unfortunately, a number of anti-TRPV1 antibodies turned out to be non-specific, producing a similar staining pattern in tissues obtained from wild-type and TRPV1-null animals (Zs. Helyes, personal communication).
Biological effects noted at very high capsaicin concentrations (for example, 100 µM or higher) are especially suspect. In human NK cells, responses to low concentrations of capsaicin (up to 10 µM) were prevented by the TRPV1 antagonists, capsazepine and SB366971 [159]. When the capsaicin concentration exceeded 50 µM, the effect could no longer be antagonized by TRPV1 blockers [159]. These observations imply that capsaicin influences NK cell activity by a mixture of specific and off-target mechanisms, and at high capsaicin concentrations the non-specific actions predominate.
In conclusion, one should take caution when interpreting capsaicin responses noted at high concentrations. However, even by strict criteria, the existence of functional TRPV1 in immune cells was proven beyond doubt. In CD4+ T-cells, capsaicin was shown to stimulate cytokine production: this response was abolished by both pharmacological inhibition and the genetic knock-out of TRPV1 [85].
10. Role of TRPV1 Expressing Immune Cells in Cancer
Local and systemic immune responses are critical for inhibiting cancer growth and metastasis [62]. As immune editing states, the ability of the immune surveillance to eliminate cancer cells decreases over time, permitting the growth of highly aggressive tumors. For certain tumors, the TRPV1-mediated neuro-immune regulation of the tumor microenvironment enhances anti-tumoral immunity by inhibiting chronic inflammation [20,62,80,160]. Immune cells express TRPV1, albeit at much lower levels than in sensory neurons [161]. The effect of TRPV1 activation in immune cells on tumor growth and metastasis might be different from that of neuronal TRPV1 (Table 1).
TRPV1 is expressed in mouse and human T cells [85,162], macrophages [163,164], dendritic cells [165,166], Jurkat cells [162], and NK cells [159]. TRPV1 plays an important role in thymocyte differentiation. In accord, the numbers of T-cells are markedly reduced in TRPV1-null mice in both the blood and spleen when compared to wild-type animals [167]. TRPV1 channels are believed to play a critical role in T-cell physiology: for example, rises in TRPV1-mediated intracellular Ca2+ concentrations may be required for T-cell activation, proliferation, differentiation, and effector functions [168,169]. In support of this concept, capsaicin increases Ca2+ influx and intracellular Ca2+ concentration in activated CD4+ T-cells, but not in resting T-cells [170,171]. TRPV1 expressed in CD4+ T-cells is involved in the activation and differentiation of T-cells into Th1 effector cells, inducing an inflammatory response in the murine model of colitis [172]. Furthermore, genetic or pharmacological inhibition of TRPV1 in CD4+ T-cells was also associated with a reduction in airway inflammation in a model of allergic asthma [173]. These observations imply that TRPV1 activation in T-cells may indeed enhance inflammatory responses.
Depending on their duration and concentration, TRPV1 agonists may induce T-cell death. For instance, prolonged exposure to high capsaicin concentrations can induce apoptosis in human peripheral T- and Jurkat cells [171,174]. Similarly, in cultured murine Peyer patch cells in vitro, capsaicin can inhibit the cytokine production by T-cells and reduce T-cell viability [175]. Moreover, TRPV1 levels are significantly increased in T-cells isolated from B16F10 tumor-bearing mice compared to control mice [176]; this is also associated with increased basal intracellular Ca2+ levels in T-cells. These findings suggest that TRPV1 agonists may have more profound effects on T-cells in cancer patients, ranging from excessive inflammation to T-cell exhaustion and T-cell death.
The role of TRPV1 in NK cell biology remains unclear. Treatment with a TRPV1 antagonist can increase the population of NK cells, but, at the same time, downregulate the activation of NK/T cells in mice infected with Plasmodium [177]. TRPV1 in human NK cells is functional since 10 µM capsaicin increases intracellular Ca2+ concentrations, while pretreatment with the specific TRPV1 antagonist, SB366791, inhibits this effect [159]. (Parenthetically, the further rise in intracellular Ca2+ in response to capsaicin concentrations exceeding 50 µM is no longer antagonized by SB366791, indicating a non-specific capsaicin action [159].)
The role of TRPV1 in dendritic cells is not clear either. The available evidence implies an indirect effect of TRPV1 activation on dendritic cells through the neuro-immune axis [118]. In humans, TRPV1 expression significantly increases during in vitro differentiation of monocytes to immature dendritic cells [178]. However, it is not known how TRPV1 expression in NK and dendritic cells may change in cancer patients.

Table 1.
TRPV1 expression in immune cells and its postulated role in carcinogenesis.
Table 1.
TRPV1 expression in immune cells and its postulated role in carcinogenesis.
| Cell Type | TRPV1 Functions | Possible Role in Carcinogenesis |
|---|---|---|
| T cells | TRPV1 implicated in TcR-induced Ca2+ influx. Capsaicin increases Ca2+ influx and intracellular Ca2+ concentration in activated CD4+ T-cells, but not in resting T-cells [170,171]. Exposure to prolonged and high concentrations of capsaicin induces apoptosis of human peripheral T- and Jurkat cells [171,174]. | TRPV1 levels are significantly increased in T-cells isolated from B16F10 tumor-bearing mice [176]. Possible role in anti-tumoral immune response and T-cell exhaustion requires further studies. |
| NK cells | Capsaicin increases intracellular Ca2+ concentrations in human NK cells while pretreatment with specific TRPV1 antagonists inhibits this effect [159]. TRPV1 antagonist increases the population of NK cells but also downregulates activation of NK/T cells in mice infected with Plasmodium [177]. | Not known |
| Dendritic cells | TRPV1 expression significantly increases during in vitro differentiation from monocytes to immature dendritic cells [178]. Indirect effect of TRPV1 activation on dendritic cells through neuroimmune axis was suggested [118]. Possible role in antigen presentation. | Not known |
| Mixed leukocyte culture | TRPV1 activation in control mice increases IFN-γ and IL-17, does not alter IL6 release [160]. | TRPV1 activation in tumor-bearing mice (breast carcinoma) decreases IFN-γ and increases IL-6 [160]. |
In the 4T1-related breast carcinoma model, mixed leucocyte cultures of tumor-bearing mice secrete markedly higher levels of IL-6 and TNF-α, and lower levels of IFN-γ [160]. When directly added to in vitro cultured mixed leucocytes, TRPV1 agonists differentially regulate cytokine secretion in tumor-bearing mice compared to controls [160]. Specifically, TRPV1 agonists markedly increase IFN-γ response in control mice, while the opposite effect occurs in tumor-bearing mice [160]. Furthermore, TRPV1 agonists increase IL-17 secretion in control mice, but not in tumor-bearing mice. Lastly, TRPV1 agonists markedly increase IL-6 secretion in tumor-bearing mice, but not in control mice. These observations imply that in the absence of aggressive inflammatory carcinoma, the activation of TRPV1 may enhance immune response against newly formed precancerous lesions by increasing IFN-γ [179] and IL-17 production [180]. On the other hand, the opposite effect may occur in the presence of highly aggressive and inflammatory carcinoma: that is, the activation of TRPV1 in immune cells may create excessive inflammatory conditions [160]. In conclusion, in the presence of inflammatory metastatic carcinoma, doses of TRPV1 agonists high enough to activate TRPV1 in immune cells may have detrimental consequences; this effect may also partly explain the tumor-promoting action of TRPV1 agonists under certain conditions.
11. Discussion
An estimated quarter of the world’s population consume capsaicin-flavored food on a daily basis, and many more use capsaicin-containing creams and ointments for topical pain relief. Three large epidemiological studies (one from the US [181], one from Europe [182], and another from China [91]) found a distinct health benefit of eating chili pepper, including lower cancer-related death rates. There are no reports of increased skin cancer risk in users of high-dose capsaicin patches either. Therefore, one may conclude that topical (oral mucosa, GI tract, or skin) exposure to TRPV1 agonists, such as capsaicin, is most likely safe.
A separate question is whether one can block carcinogenesis and/or metastasis formation by manipulating TRPV1-expressing nerves and immune cells. The answer is probably tumor-specific. For example, chronic inflammation is a well-known risk factor of pancreatic carcinoma [183]. Sensory neurons that mediate neurogenic inflammation are believed to play an active role in the initiation and progression of this type of cancer [184]. It was speculated that in the pancreas, chronic inflammation could drive carcinogenesis through both K-ras activation and independent of K-ras [185]. If so, silencing TRPV1-expressing nerves using capsaicin may protect against pancreatic cancer. This proposal has recently gained experimental support by the protective action of RTX-desensitization in a melanoma model [140]. By contrast, TRPV1-mediated neurogenic inflammation was reported to protect mice from colonic adenocarcinoma [95]. Similarly, activation of TRPV1-positive sensory nerve fibers was shown to enhance anti-tumoral immunity in a breast cancer model [20]. Therefore, capsaicin desensitization may be a “double-edged sword” in the fight against cancer: it may protect against pancreatic cancer and melanoma and, at the same time, conversely increase the risk for colon and breast cancer.
TRPV1-positive sensory nerves might also play a crucial role in the vicious cycle that drives metastatic bone disease [186]. Cancer-associated bone pain is thought to involve sensitization and continuous activation of TRPV1-expressing nerves by the tumor microenvironment [186] (Figure 2). Indeed, in companion dogs with bone cancer, chemical ablation by intrathecal resiniferatoxin of sensory afferents ameliorated the debilitating cancer pain [187,188]. At present, intrathecal resiniferatoxin as a “molecular scalpel” is undergoing clinical trials for permanent analgesia in women with cervical carcinoma metastatic to the bone. If the hypothesis that capsaicin-sensitive afferents drive metastatic bone disease is true, then resiniferatoxin-treated patients are expected to have a reduced tumor burden in addition to pain relief. On the other hand, sensory afferents may play the opposite role in breast carcinoma where capsaicin desensitization was found to promote the spread of metastatic breast cancer [20,73,189]. Taken together, these findings imply that ablation of sensory nerves with resiniferatoxin should not be used for pain relief in women with metastatic mammary carcinoma. Instead, desensitization by perineural TRPV1 agonist administration of nerve fibers might be a better approach for pain relief. In preclinical pain models, perineural resiniferatoxin was shown to achieve lasting conduction analgesia [190].
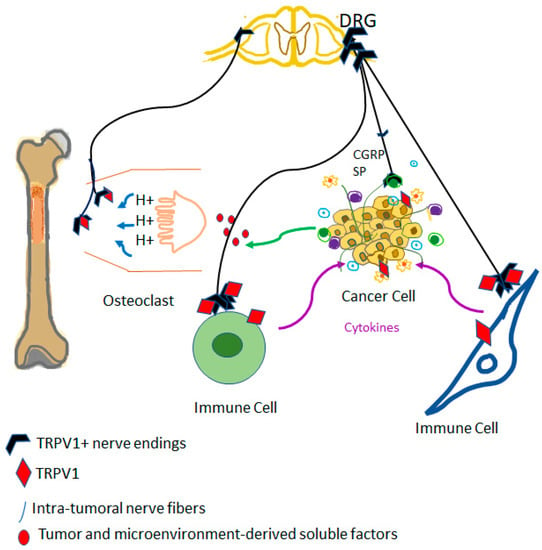
Figure 2.
TRPV1 as a driver of the vicious cycle of metastatic bone disease. Cancer cells metastatic to bone may stimulate osteoclasts that, in turn, create an acidic environment. Protons are well-known activators of TRPV1 on sensory afferents; this is perceived as cancer pain in the central nervous system. Furthermore, TRPV1-expressing efferents are sites of release for pro-inflammatory neuropeptides, such as substance P (SP), and calcitonin gene-related peptide (CGRP): these peptides may promote the growth of metastatic cancer by blocking immune cells and increasing blood supply.
A third question is the effect (if any) of small-molecule TRPV1 antagonists on carcinogenesis. It is safe to assume that these compounds did not display any carcinogenic potential in animal experiments. In clinical trials, most of these antagonists caused a hyperthermic reaction [190,191,192,193]. Furthermore, minor burn injuries were reported as adverse effects [5]. So far, no concern has been raised with regard to carcinogenesis, but the clinical experience with these compounds is very limited.
TRPV1 activation and/or blockade may have markedly different effects on carcinogenesis depending on both the tumor and its microenvironment. Three large epidemiological studies suggest that TRPV1 activation by culinary capsaicin exposure protects against cancer [91,181,182]. In experimental animals, large doses of a TRPV1 agonist might be tumorigenic [194,195], presumably by activating immune cells. The relevance of these observations in humans is, however, unclear since men are unlikely to be exposed to similarly high TRPV1 agonist doses.
In conclusion, there is good evidence to implicate TRPV1-expressing afferents and immune cells in the bi-directional communication between cancer and its microenvironment. The fine details of this interactions are yet to be elucidated. Results obtained with high capsaicin concentrations have to be carefully reevaluated to exclude the possibility of an off-target action. Further studies are warranted to tailor specific TRPV1-based approaches for cancer therapy as some cancer patients may respond to TRPV1 agonists whereas others may benefit from TRPV1 blockade.
Author Contributions
Writing—original draft preparation, N.E. and A.S.; writing—review and editing, N.E. and A.S. All authors have read and agreed to the published version of the manuscript.
Funding
N.E. acknowledges support for her laboratory by Akdeniz University Research Unit (BAP project no: TSA-2018-3007 and TYL-2019-4862) and by TUBITAK (Turkish Research Council).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Montell, C.; Birnbaumer, L.; Flockerzi, V.; Bindels, R.J.; Bruford, E.A.; Caterina, M.J.; Clapham, D.E.; Harteneck, C.; Heller, S.; Julius, D.; et al. A unified nomenclature for the superfamily of TRP cation channels. Mol. Cell 2002, 9, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Patapoutian, A.; Peier, A.M.; Story, G.M.; Viswanath, V. ThermoTRP channels and beyond: Mechanisms of temperature sensation. Nat. Rev. Neurosci. 2003, 4, 529–539. [Google Scholar] [CrossRef] [PubMed]
- García-Ávila, M.; Islas, L.D. What is new about mild temperature sensing? A review of recent findings. Temperature 2019, 6, 132–141. [Google Scholar] [CrossRef]
- Moran, M.M.; Szallasi, A. Targeting nociceptive transient receptor potential channels to treat chronic pain: Current state of the field. Br. J. Pharmacol. 2018, 175, 2185–2203. [Google Scholar] [CrossRef]
- Szolcsanyi, J. A pharmacological approach to elucidation of the role of different nerve fibres and receptor endings in mediation of pain. J. Physiol. 1977, 73, 251–259. [Google Scholar]
- Karrer, T.; Bartoshuk, L. Capsaicin desensitization and recovery on the human tongue. Physiol. Behav. 1991, 49, 757–764. [Google Scholar] [CrossRef]
- Szallasi, A.; Blumberg, P.M. Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol. Rev. 1999, 51, 159–212. [Google Scholar]
- Abrams, R.M.C.; Pedowitz, E.J.; Simpson, D.M. A critical review of the capsaicin 8% patch for the treatment of neuropathic pain associated with diabetic peripheral neuropathy of the feet in adults. Expert Rev. Neurother. 2021, 21, 259–266. [Google Scholar] [CrossRef]
- Campbell, J.N.; Stevens, R.; Hanson, P.; Connolly, J.; Meske, D.S.; Chung, M.K.; Lascelles, B.D.X. Injectable Capsaicin for the Management of Pain Due to Osteoarthritis. Molecules 2021, 26, 778. [Google Scholar] [CrossRef]
- Jancso, G.; Kiraly, E.; Such, G.; Joo, F.; Nagy, A. Neurotoxic effect of capsaicin in mammals. Acta Physiol. Hung. 1987, 69, 295–313. [Google Scholar] [PubMed]
- Maggi, C.A.; Meli, A. The sensory-efferent function of capsaicin-sensitive sensory neurons. Gen. Pharmacol. 1988, 19, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.J.M.; Ciotu, C.I.; Szallasi, A. The Mysteries of Capsaicin-Sensitive Afferents. Front. Physiol. 2020, 11, 554195. [Google Scholar] [CrossRef]
- Ichikawa, H.; Sugimoto, T. The co-expression of VR1 and VRL-1 in the rat vagal sensory ganglia. Brain Res. 2003, 980, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P. Local effector functions of capsaicin-sensitive sensory nerve endings: Involvement of tachykinins, calcitonin gene-related peptide and other neuropeptides. Neuroscience 1988, 24, 739–768. [Google Scholar] [CrossRef]
- Barnes, P.J.; Belvisi, M.G.; Rogers, D.F. Modulation of neurogenic inflammation: Novel approaches to inflammatory disease. Trends Pharmacol. Sci. 1990, 11, 185–189. [Google Scholar] [CrossRef]
- Geppetti, P.; Holzer, P. Neurogenic Inflammation; CRC Press: Boca Raton, FL, USA, 1996. [Google Scholar]
- Szallasi, A.; Blumberg, P.M. Effect of resiniferatoxin pretreatment on the inflammatory response to phorbol-12-myristate-13-acetate in mouse strains with different susceptibilities to phorbol ester tumor promotion. Carcinogenesis 1990, 11, 583–587. [Google Scholar] [CrossRef]
- Bencze, N.; Schvarcz, C.; Kriszta, G.; Danics, L.; Szoke, E.; Balogh, P.; Szallasi, A.; Hamar, P.; Helyes, Z.; Botz, B. Desensitization of Capsaicin-Sensitive Afferents Accelerates Early Tumor Growth via Increased Vascular Leakage in a Murine Model of Triple Negative Breast Cancer. Front. Oncol. 2021, 11, 685297. [Google Scholar] [CrossRef]
- Erin, N. Role of sensory neurons, neuroimmune pathways, and transient receptor potential vanilloid 1 (TRPV1) channels in a murine model of breast cancer metastasis. Cancer Immunol. Immunother. 2020, 69, 307–314. [Google Scholar] [CrossRef]
- Omari, S.A.; Adams, M.J.; Geraghty, D.P. TRPV1 Channels in Immune Cells and Hematological Malignancies. Adv. Pharmacol. 2017, 79, 173–198. [Google Scholar]
- Palma, C. Tachykinins and their receptors in human malignancies. Curr. Drug Targets 2006, 7, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Perner, C.; Flayer, C.H.; Zhu, X.; Aderhold, P.A.; Dewan, Z.N.A.; Voisin, T.; Camire, R.B.; Chow, O.A.; Chiu, I.M.; Sokol, C.L. Substance P Release by Sensory Neurons Triggers Dendritic Cell Migration and Initiates the Type-2 Immune Response to Allergens. Immunity 2020, 53, 1063–1077.e7. [Google Scholar] [CrossRef] [PubMed]
- Payan, D.G.; Brewster, D.R.; Goetzl, E.J. Specific stimulation of human T lymphocytes by substance P. J. Immunol. 1983, 131, 1613–1615. [Google Scholar] [CrossRef] [PubMed]
- Munoz, M.; Covenas, R.; Esteban, F.; Redondo, M. The substance P/NK-1 receptor system: NK-1 receptor antagonists as anti-cancer drugs. J. Biosci. 2015, 40, 441–463. [Google Scholar] [CrossRef]
- Amantini, C.; Mosca, M.; Nabissi, M.; Lucciarini, R.; Caprodossi, S.; Arcella, A.; Giangaspero, F.; Santoni, G. Capsaicin-induced apoptosis of glioma cells is mediated by TRPV1 vanilloid receptor and requires p38 MAPK activation. J. Neurochem. 2007, 102, 977–990. [Google Scholar] [CrossRef]
- Schulz, S.; Stumm, R.; Rocken, C.; Mawrin, C.; Schulz, S. Immunolocalization of full-length NK1 tachykinin receptors in human tumors. J. Histochem. Cytochem. 2006, 54, 1015–1020. [Google Scholar] [CrossRef]
- Munoz, M.; Covenas, R. Glioma and Neurokinin-1 Receptor Antagonists: A New Therapeutic Approach. Anti-Cancer Agents Med. Chem. 2019, 19, 92–100. [Google Scholar] [CrossRef]
- Virchow, R. Reizung und Reizbarkeit. Arch. Pathol. Anat. Physiol. Klin. Med. 1858, 14, 1–63. [Google Scholar] [CrossRef]
- Yamagiwa, K.I.K. Åber die atypische Epithelwucherung. Gann 1914, 8, 11–15. [Google Scholar]
- Berenblum, I.; Shubik, P. The persistence of latent tumour cells induced in the mouse’s skin by a single application of 9:10-dimethyl-1:2-benzanthracene. Br. J. Cancer 1949, 3, 384–386. [Google Scholar] [CrossRef]
- Berenblum, I. The carcinogenic action of 9,10-dimethyl-1,2-benzanthracene on the skin and subcutaneous tissues of the mouse, rabbit, rat and guinea pig. J. Natl. Cancer Inst. 1949, 10, 167–174. [Google Scholar] [PubMed]
- Klein, M. Induction of skin tumors in the mouse with minute doses of 9, 10-dimethyl-1, 2-benzanthracene alone or with croton oil. Cancer Res. 1956, 16, 123–127. [Google Scholar] [PubMed]
- Brune, K.; Kalin, H.; Schmidt, R.; Hecker, E. Inflammatory, tumor initiating and promoting activities of polycyclic aromatic hydrocarbons and diterpene esters in mouse skin as compared with their prostaglandin releasing potency in vitro. Cancer Lett. 1978, 4, 333–342. [Google Scholar] [CrossRef]
- Ohuchi, K.; Watanabe, M.; Takahashi, C.; Hayashi, Y.; Hirasawa, N.; Tsurufuji, S.; Fujiki, H.; Sugimura, T. Analysis of tumor-promoter-induced inflammation in rats: Participation of histamine and prostaglandin E2. Biochim. Biophys. Acta 1987, 925, 156–163. [Google Scholar]
- Hergenhahn, M.; Kusumoto, S.; Hecker, E. On the active principles of the spurge family (Euphorbiaceae). V. Extremely skin-irritant and moderately tumor-promoting diterpene esters from Euphorbia resinifera Berg. J. Cancer Res. Clin. Oncol. 1984, 108, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Hausen, H.Z.; Bornkamm, G.W.; Schmidt, R.; Hecker, E. Tumor initiators and promoters in the induction of Epstein-Barr virus. Proc. Natl. Acad. Sci. USA 1979, 76, 782–785. [Google Scholar] [CrossRef]
- Driedger, P.E.; Blumberg, P.M. Different biological targets for resiniferatoxin and phorbol 12-myristate 13-acetate. Cancer Res. 1980, 40, 1400–1404. [Google Scholar]
- Blumberg, P.M.; Pettit, G.R.; Warren, B.S.; Szallasi, A.; Schuman, L.D.; Sharkey, N.A.; Nakakuma, H.; Dell’Aquila, M.L.; de Vries, D.J. The protein kinase C pathway in tumor promotion. Prog. Clin. Biol. Res. 1989, 298, 201–212. [Google Scholar]
- Szallasi, A.; Blumberg, P.M. Specific binding of resiniferatoxin, an ultrapotent capsaicin analog, by dorsal root ganglion membranes. Brain Res. 1990, 524, 106–111. [Google Scholar] [CrossRef]
- Szallasi, A.; Blumberg, P.M. Neurogenic component of phorbol ester-induced mouse skin inflammation. Cancer Res. 1989, 49, 6052–6057. [Google Scholar]
- Inoue, K.; Koizumi, S.; Fuziwara, S.; Denda, S.; Inoue, K.; Denda, M. Functional vanilloid receptors in cultured normal human epidermal keratinocytes. Biochem. Biophys. Res. Commun. 2002, 291, 124–129. [Google Scholar] [CrossRef]
- Southall, M.D.; Li, T.; Gharibova, L.S.; Pei, Y.; Nicol, G.D.; Travers, J.B. Activation of epidermal vanilloid receptor-1 induces release of proinflammatory mediators in human keratinocytes. J. Pharmacol. Exp. Ther. 2003, 304, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J.; Han, S.S.; Keum, Y.S.; Seo, H.J.; Lee, S.S. Inhibitory effects of curcumin and capsaicin on phorbol ester-induced activation of eukaryotic transcription factors, NF-kappaB and AP-1. Biofactors 2000, 12, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Hwang, M.K.; Bode, A.M.; Byun, S.; Song, N.R.; Lee, H.J.; Lee, K.W.; Dong, Z. Cocarcinogenic effect of capsaicin involves activation of EGFR signaling but not TRPV1. Cancer Res. 2010, 70, 6859–6869. [Google Scholar] [CrossRef] [PubMed]
- Mundt, S.; Greter, M.; Flugel, A.; Becher, B. The CNS Immune Landscape from the Viewpoint of a T Cell. Trends Neurosci. 2019, 42, 667–679. [Google Scholar] [CrossRef]
- Antoni, M.H.; Dhabhar, F.S. The impact of psychosocial stress and stress management on immune responses in patients with cancer. Cancer 2019, 125, 1417–1431. [Google Scholar] [CrossRef]
- Dhabhar, F.S. Enhancing versus suppressive effects of stress on immune function: Implications for immunoprotection and immunopathology. Neuroimmunomodulation 2009, 16, 300–317. [Google Scholar] [CrossRef]
- Oh, P.J.; Shin, S.R.; Ahn, H.S.; Kim, H.J. Meta-analysis of psychosocial interventions on survival time in patients with cancer. Psychol. Health 2016, 31, 396–419. [Google Scholar] [CrossRef]
- Shurin, M.R.; Shurin, G.V.; Zlotnikov, S.B.; Bunimovich, Y.L. The Neuroimmune Axis in the Tumor Microenvironment. J. Immunol. 2020, 204, 280–285. [Google Scholar] [CrossRef]
- Candido, J.; Hagemann, T. Cancer-related inflammation. J. Clin. Immunol. 2013, 33 (Suppl. 1), S79–S84. [Google Scholar] [CrossRef]
- Mueller, S.N. Neural control of immune cell trafficking. J. Exp. Med. 2022, 219, e20211604. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.; Yin, X.T.; Stuart, P.M.; St Leger, A.J. Sensory Nerve Retraction and Sympathetic Nerve Innervation Contribute to Immunopathology of Murine Recurrent Herpes Stromal Keratitis. Investig. Ophthalmol. Vis. Sci. 2022, 63, 4. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Chen, W.; Shen, L.; Chen, Z.; Huang, J. Crosstalk between the peripheral nervous system and breast cancer influences tumor progression. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188828. [Google Scholar] [CrossRef] [PubMed]
- Nance, D.M.; Sanders, V.M. Autonomic innervation and regulation of the immune system (1987–2007). Brain Behav. Immun. 2007, 21, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, A.; Hayama, Y.; Kato, S.; Shimomura, A.; Shimomura, T.; Irie, K.; Kaneko, R.; Yanagawa, Y.; Kobayashi, K.; Ochiya, T. Genetic manipulation of autonomic nerve fiber innervation and activity and its effect on breast cancer progression. Nat. Neurosci. 2019, 22, 1289–1305. [Google Scholar] [CrossRef]
- Andersson, U.; Tracey, K.J. Neural reflexes in inflammation and immunity. J. Exp. Med. 2012, 209, 1057–1068. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Tracey, K.J. Neural circuitry and immunity. Immunol. Res. 2015, 63, 38–57. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.R.; Neuhuber, W.L. Functional and chemical anatomy of the afferent vagal system. Auton. Neurosci. 2000, 85, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Bonaz, B.; Sinniger, V.; Pellissier, S. Vagus Nerve Stimulation at the Interface of Brain-Gut Interactions. Cold Spring Harb. Perspect. Med. 2019, 9, a034199. [Google Scholar] [CrossRef] [PubMed]
- Reardon, C.; Murray, K.; Lomax, A.E. Neuroimmune Communication in Health and Disease. Physiol. Rev. 2018, 98, 2287–2316. [Google Scholar] [CrossRef] [PubMed]
- Erin, N.; Shurin, G.V.; Baraldi, J.H.; Shurin, M.R. Regulation of Carcinogenesis by Sensory Neurons and Neuromediators. Cancers 2022, 14, 2333. [Google Scholar] [CrossRef]
- Sternini, C. Organization of the peripheral nervous system: Autonomic and sensory ganglia. J. Investig. Dermatol. Symp. Proc. 1997, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.S.; Hadaya, J.; Khalsa, S.S.; Yu, C.; Chang, R.; Shivkumar, K. The Vagus Nerve in Cardiovascular Physiology and Pathophysiology: From Evolutionary Insights to Clinical Medicine. In Seminars in Cell and Developmental Biology; Academic Press: Berlin/Heidelberg, Germany, 2023. [Google Scholar]
- Cirillo, G.; Negrete-Diaz, F.; Yucuma, D.; Virtuoso, A.; Korai, S.A.; De Luca, C.; Kaniusas, E.; Papa, M.; Panetsos, F. Vagus Nerve Stimulation: A Personalized Therapeutic Approach for Crohn’s and Other Inflammatory Bowel Diseases. Cells 2022, 11, 4103. [Google Scholar] [CrossRef]
- Ruffoli, R.; Giorgi, F.S.; Pizzanelli, C.; Murri, L.; Paparelli, A.; Fornai, F. The chemical neuroanatomy of vagus nerve stimulation. J. Chem. Neuroanat. 2011, 42, 288–296. [Google Scholar] [CrossRef]
- Fernández-Ballester, G.; Fernández-Carvajal, A.; Ferrer-Montiel, A. Progress in the structural basis of thermoTRP channel polymodal gating. Int. J. Mol. Med. 2023, 24, 743. [Google Scholar] [CrossRef]
- Luu, D.D.; Owens, A.M.; Mebrat, M.D.; Van Horn, W.D. A molecular perspective on identifying TRPV1 thermosensitive regions and disentangling polymodal activation. Temperature 2021, 10, 67–101. [Google Scholar] [CrossRef]
- Ulloa, L. The vagus nerve and the nicotinic anti-inflammatory pathway. Nature Rev. Drug Discov. 2005, 4, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Gallowitsch-Puerta, M.; Pavlov, V.A. Neuro-immune interactions via the cholinergic anti-inflammatory pathway. Life Sci. 2007, 80, 2325–2329. [Google Scholar] [CrossRef]
- Jo, B.G.; Kim, S.H.; Namgung, U. Vagal afferent fibers contribute to the anti-inflammatory reactions by vagus nerve stimulation in concanavalin A model of hepatitis in rats. Mol. Med. 2020, 26, 119. [Google Scholar] [CrossRef] [PubMed]
- Erin, N.; Akdas Barkan, G.; Harms, J.F.; Clawson, G.A. Vagotomy enhances experimental metastases of 4THMpc breast cancer cells and alters substance P level. Regul. Pept. 2008, 151, 35–42. [Google Scholar] [CrossRef]
- Erin, N.; Boyer, P.J.; Bonneau, R.H.; Clawson, G.A.; Welch, D.R. Capsaicin-mediated denervation of sensory neurons promotes mammary tumor metastasis to lung and heart. Anticancer Res. 2004, 24, 1003–1009. [Google Scholar] [PubMed]
- Erin, N.; Barkan, G.A.; Clawson, G.A. Vagus nerve regulates breast cancer metastasis to the adrenal gland. Anticancer Res. 2013, 33, 3675–3682. [Google Scholar]
- Erin, N.; Duymus, O.; Ozturk, S.; Demir, N. Activation of vagus nerve by semapimod alters substance P levels and decreases breast cancer metastasis. Regul. Pept. 2012, 179, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yu, Q.; Zhang, X.; Wang, X.; Su, Y.; He, W.; Li, J.; Wan, H.; Jing, X. Electroacupuncture regulates inflammatory cytokines by activating the vagus nerve to enhance antitumor immunity in mice with breast tumors. Life Sci. 2021, 272, 119259. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yi, M.; Niu, M.; Mei, Q.; Wu, K. Myeloid-derived suppressor cells: An emerging target for anticancer immunotherapy. Mol. Cancer 2022, 21, 184. [Google Scholar] [CrossRef] [PubMed]
- Croitoru, K.; Ernst, P.B.; Bienenstock, J.; Padol, I.; Stanisz, A.M. Selective modulation of the natural killer activity of murine intestinal intraepithelial leucocytes by the neuropeptide substance P. Immunology 1990, 71, 196–201. [Google Scholar]
- Janelsins, B.M.; Sumpter, T.L.; Tkacheva, O.A.; Rojas-Canales, D.M.; Erdos, G.; Mathers, A.R.; Shufesky, W.J.; Storkus, W.J.; Falo, L.D.; Jr Morelli, A.E.; et al. Neurokinin-1 receptor agonists bias therapeutic dendritic cells to induce type 1 immunity by licensing host dendritic cells to produce IL-12. Blood 2013, 121, 2923–2933. [Google Scholar] [CrossRef]
- Erin, N.; Korcum, A.F.; Tanriover, G.; Kale, S.; Demir, N.; Koksoy, S. Activation of neuroimmune pathways increases therapeutic effects of radiotherapy on poorly differentiated breast carcinoma. Brain Behav. Immun. 2015, 48, 174–185. [Google Scholar] [CrossRef]
- Chu, C.J.; Huang, S.M.; De Petrocellis, L.; Bisogno, T.; Ewing, S.A.; Miller, J.D.; Zipkin, R.E.; Daddario, N.; Appendino, G.; Di Marzo, V.; et al. N-oleoyldopamine, a novel endogenous capsaicin-like lipid that produces hyperalgesia. J. Biol. Chem. 2003, 278, 13633–13639. [Google Scholar] [CrossRef]
- Liu, L.; Lo, Y.; Chen, I.; Simon, S.A. The responses of rat trigeminal ganglion neurons to capsaicin and two nonpungent vanilloid receptor agonists, olvanil and glyceryl nonamide. J. Neurosci. 1997, 17, 4101–4111. [Google Scholar] [CrossRef]
- Erin, N.; Akman, M.; Aliyev, E.; Tanriover, G.; Korcum, A.F. Olvanil activates sensory nerve fibers, increases T cell response and decreases metastasis of breast carcinoma. Life Sci. 2022, 291, 120305. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.P.; Franco-Cereceda, A.; Lundberg, J.M. Different ioan channel mechanisms between low concentrations of capsaicin and high concentrations of capsaicin and nicotine regarding peptide release from pulmonary afferents. Acta Physiol. Scand. 1992, 146, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Assas, B.M.; Abdulaal, W.H.; Wakid, M.H.; Zakai, H.A.; Miyan, J.; Pennock, J.L. The use of flow cytometry to examine calcium signalling by TRPV1 in mixed cell populations. Anal. Biochem. 2017, 527, 13–19. [Google Scholar] [CrossRef]
- Liu, L.; SA, S. A rapid capsaicin-activated current in rat trigeminal ganglion neurons. Proc. Natl. Acad. Sci. USA 1994, 91, 738–741. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.; Lamotte, R.H.; Klusch, A.; Kniffki, K.D. Multiple capsaicin-evoked currents in isolated rat sensory neurons. Neuroscience 1996, 75, 495–505. [Google Scholar] [CrossRef]
- Grüter, T.; Blusch, A.; Motte, J.; Sgodzai, M.; Bachir, H.; Klimas, R.; Ambrosius, B.; Gold, R.; Ellrichmann, G.; Pitarokoili, K. Immunomodulatory and anti-oxidative effect of the direct TRPV1 receptor agonist capsaicin on Schwann cells. J. Neuroinflamm. 2020, 17, 145. [Google Scholar] [CrossRef]
- Motte, J.; Ambrosius, B.; Gruter, T.; Bachir, H.; Sgodzai, M.; Pedreiturria, X.; Pitarokoili, K.; Gold, R. Capsaicin-enriched diet ameliorates autoimmune neuritis in rats. J. Neuroinflamm. 2018, 15, 122. [Google Scholar] [CrossRef]
- Nevius, E.; Srivastava, P.K.; Basu, S. Oral ingestion of Capsaicin, the pungent component of chili pepper, enhances a discreet population of macrophages and confers protection from autoimmune diabetes. Mucosal. Immunol. 2012, 5, 76–86. [Google Scholar] [CrossRef]
- Lv, J.; Qi, L.; Yu, C.; Yang, L.; Guo, Y.; Chen, Y.; Bian, Z.; Sun, D.; Du, J.; Ge, P.; et al. Consumption of spicy foods and total and cause specific mortality: Population based cohort study. BMJ 2015, 351, h3942. [Google Scholar] [CrossRef]
- Feng, J.; Yang, P.; Mack, M.R.; Dryn, D.; Luo, J.; Gong, X.; Liu, S.; Oetjen, L.K.; Zholos, A.V.; Mei, Z.; et al. Sensory TRP channels contribute differentially to skin inflammation and persistent itch. Nat. Commun. 2017, 8, 980. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, D.H. TRPV1 ablation aggravates inflammatory responses and organ damage during endotoxic shock. Clin. Vaccine Immunol. 2013, 20, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Liang, L.; Smillie, S.J.; Kaiser, F.; Purcell, R.; Rivett, D.W.; Alam, S.; Howat, S.; Collins, H.; Thompson, S.J.; et al. TRPV1 deletion enhances local inflammation and accelerates the onset of systemic inflammatory response syndrome. J. Immunol. 2012, 188, 5741–5751. [Google Scholar] [CrossRef] [PubMed]
- Vinuesa, A.G.; Sancho, R.; Garcia-Limones, C.; Behrens, A.; ten Dijke, P.; Calzado, M.A.; Munoz, E. Vanilloid receptor-1 regulates neurogenic inflammation in colon and protects mice from colon cancer. Cancer Res. 2012, 72, 1705–1716. [Google Scholar] [CrossRef] [PubMed]
- Bryant, P.; Shumate, M.; Yumet, G.; Lang, C.H.; Vary, T.C.; Cooney, R.N. Capsaicin-sensitive nerves regulate the metabolic response to abdominal sepsis. J. Surg. Res. 2003, 112, 152–161. [Google Scholar] [CrossRef]
- Kobayashi, M.; Watanabe, K.; Yokoyama, S.; Matsumoto, C.; Hirata, M.; Tominari, T.; Inada, M.; Miyaura, C. Capsaicin, a TRPV1 Ligand, Suppresses Bone Resorption by Inhibiting the Prostaglandin E Production of Osteoblasts, and Attenuates the Inflammatory Bone Loss Induced by Lipopolysaccharide. ISRN Pharmacol. 2012, 2012, 439860. [Google Scholar] [CrossRef]
- Clark, N.; Keeble, J.; Fernandes, E.S.; Starr, A.; Liang, L.; Sugden, D.; de Winter, P.; Brain, S.D. The transient receptor potential vanilloid 1 (TRPV1) receptor protects against the onset of sepsis after endotoxin. FASEB J. 2007, 21, 3747–3755. [Google Scholar] [CrossRef]
- Szallasi, A.; Di Marzo, V. New perspectives on enigmatic vanilloid receptors. Trends Neurosci. 2000, 23, 491–507. [Google Scholar] [CrossRef]
- Zsombok, A.; Gao, H.; Miyata, K.; Issa, A.; Derbenev, A.V. Immunohistochemical localization of transient receptor potential vanilloid type 1 and insulin receptor substrate 2 and their co-localization with liver-related neurons in the hypothalamus and brainstem. Brain Res. 2011, 1398, 30–39. [Google Scholar] [CrossRef]
- Mezey, E.; Toth, Z.E.; Cortright, D.N.; Arzubi, M.K.; Krause, J.E.; Elde, R.; Guo, A.; Blumberg, P.M.; Szallasi, A. Distribution of mRNA for vanilloid receptor subtype 1 (VR1), and VR1-like immunoreactivity, in the central nervous system of the rat and human. Proc. Natl. Acad. Sci. USA 2000, 97, 3655–3660. [Google Scholar] [CrossRef]
- Tóth, A.; Boczán, J.; Kedei, N.; Lizanecz, E.; Bagi, Z.; Papp, Z.; Édes, I.; Csiba, L.; Blumberg, P.M. Expression and distribution of vanilloid receptor 1 (TRPV1) in the adult rat brain. Brain Res. Mol. Brain Res. 2005, 135, 162–168. [Google Scholar] [CrossRef]
- Steenland, H.W.; Ko, S.W.; Wu, L.J.; Zhuo, M. Hot receptors in the brain. Mol. Pain 2006, 2, 34. [Google Scholar] [CrossRef] [PubMed]
- Storozhuk, M.V.; Moroz, O.F.; Zholos, A.V. Multifunctional TRPV1 Ion Channels in Physiology and Pathology with Focus on the Brain, Vasculature, and Some Visceral Systems. BioMed Res. Int. 2019, 2019, 5806321. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, S.; Vaughan, C.W.; Christie, M.J.; Connor, M. Capsaicin activation of glutamatergic synaptic transmission in the rat locus coeruleus in vitro. J. Physiol. 2002, 543, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Chavez, A.E.; Chiu, C.Q.; Castillo, P.E. TRPV1 activation by endogenous anandamide triggers postsynaptic long-term depression in dentate gyrus. Nat. Neurosci. 2010, 13, 1511–1518. [Google Scholar] [CrossRef]
- Cui, Y.; Perez, S.; Venance, L. Endocannabinoid-LTP Mediated by CB1 and TRPV1 Receptors Encodes for Limited Occurrences of Coincident Activity in Neocortex. Front. Cell. Neurosci. 2018, 12, 182. [Google Scholar] [CrossRef]
- Anstotz, M.; Lee, S.K.; Maccaferri, G. Expression of TRPV1 channels by Cajal-Retzius cells and layer-specific modulation of synaptic transmission by capsaicin in the mouse hippocampus. J. Physiol. 2018, 596, 3739–3758. [Google Scholar] [CrossRef]
- Kim, S.R.; Kim, S.U.; Oh, U.; Jin, B.K. Transient receptor potential vanilloid subtype 1 mediates microglial cell death in vivo and in vitro via Ca2+-mediated mitochondrial damage and cytochrome c release. J. Immunol. 2006, 177, 4322–4329. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.L.; Kong, W.L.; Zeng, M.L.; Shao, L.; Jiang, G.T.; Cheng, J.J.; Kong, S.; He, X.H.; Liu, W.H.; et al. TRPV1 translocated to astrocytic membrane to promote migration and inflammatory infiltration thus promotes epilepsy after hypoxic ischemia in immature brain. J. Neuroinflamm. 2019, 16, 214. [Google Scholar] [CrossRef]
- Marrone, M.C.; Morabito, A.; Giustizieri, M.; Chiurchiu, V.; Leuti, A.; Mattioli, M.; Marinelli, S.; Riganti, L.; Lombardi, M.; Murana, E.; et al. TRPV1 channels are critical brain inflammation detectors and neuropathic pain biomarkers in mice. Nat. Commun. 2017, 8, 15292. [Google Scholar] [CrossRef]
- Nam, J.H.; Park, E.S.; Won, S.Y.; Lee, Y.A.; Kim, K.I.; Jeong, J.Y.; Baek, J.Y.; Cho, E.J.; Jin, M.; Chung, Y.C.; et al. TRPV1 on astrocytes rescues nigral dopamine neurons in Parkinson’s disease via CNTF. Brain 2015, 138, 3610–3622. [Google Scholar] [CrossRef]
- Di Marzo, V.; Blumberg, P.M.; Szallasi, A. Endovanilloid signaling in pain. Curr. Opin. Neurobiol. 2002, 12, 372–379. [Google Scholar] [CrossRef]
- Joffre, J.; Wong, E.; Lawton, S.; Lloyd, E.; Nguyen, N.; Xu, F.; Sempio, C.; Kobzik, L.; Zlatanova, I.; Schumacher, M.; et al. N-Oleoyl dopamine induces IL-10 via central nervous system TRPV1 and improves endotoxemia and sepsis outcomes. J. Neuroinflamm. 2022, 19, 118. [Google Scholar] [CrossRef]
- Zou, S.; Kumar, U. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar] [CrossRef] [PubMed]
- Lawton, S.K.; Xu, F.; Tran, A.; Wong, E.; Prakash, A.; Schumacher, M.; Hellman, J.; Wilhelmsen, K. N-Arachidonoyl Dopamine Modulates Acute Systemic Inflammation via Nonhematopoietic TRPV1. J. Immunol. 2017, 199, 1465–1475. [Google Scholar] [CrossRef]
- Rollyson, W.D.; Stover, C.A.; Brown, K.C.; Perry, H.E.; Stevenson, C.D.; McNees, C.A.; Ball, J.G.; Valentovic, M.A.; Dasgupta, P. Bioavailability of capsaicin and its implications for drug delivery. J. Control. Release 2014, 196, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Bujak, J.K.; Kosmala, D.; Szopa, I.M.; Majchrzak, K.; Bednarczyk, P. Inflammation, Cancer and Immunity-Implication of TRPV1 Channel. Front. Oncol. 2019, 9, 1087. [Google Scholar] [CrossRef] [PubMed]
- Mravec, B. Neurobiology of cancer: Definition, historical overview, and clinical implications. Cancer Med. 2022, 11, 903–921. [Google Scholar] [CrossRef]
- Palm, D.; Entschladen, F. Neoneurogenesis and the neuro-neoplastic synapse. Neuronal Act. Tumor Tissue 2007, 39, 91–98. [Google Scholar]
- Magnon, C.; Hall, S.J.; Lin, J.; Xue, X.; Gerber, L.; Freedland, S.J.; Frenette, P.S. Autonomic nerve development contributes to prostate cancer progression. Science 2013, 341, 1236361. [Google Scholar] [CrossRef]
- Ferdoushi, A.; Griffin, N.; Marsland, M.; Xu, X.; Faulkner, S.; Gao, F.; Liu, H.; King, S.J.; Denham, J.W.; van Helden, D.F.; et al. Tumor innervation and clinical outcome in pancreatic cancer. Sci. Rep. 2021, 11, 7390. [Google Scholar] [CrossRef]
- Huang, D.; Su, S.; Cui, X.; Shen, X.; Zeng, Y.; Wu, W.; Chen, J.; Chen, F.; He, C.; Liu, J.; et al. Nerve fibers in breast cancer tissues indicate aggressive tumor progression. Medicine 2014, 93, e172. [Google Scholar] [CrossRef]
- McCallum, G.A.; Shiralkar, J.; Suciu, D.; Covarrubias, G.; Yu, J.S.; Karathanasis, E.; Durand, D.M. Chronic neural activity recorded within breast tumors. Sci. Rep. 2020, 10, 14824. [Google Scholar] [CrossRef]
- Allen, J.K.; Armaiz-Pena, G.N.; Nagaraja, A.S.; Sadaoui, N.C.; Ortiz, T.; Dood, R.; Ozcan, M.; Herder, D.M.; Haemmerle, M.; Gharpure, K.M.; et al. Sustained Adrenergic Signaling Promotes Intratumoral Innervation through BDNF Induction. Cancer Res. 2018, 78, 3233–3242. [Google Scholar] [CrossRef]
- Renz, B.W.; Takahashi, R.; Tanaka, T.; Macchini, M.; Hayakawa, Y.; Dantes, Z.; Maurer, H.C.; Chen, X.; Jiang, Z.; Westphalen, C.B.; et al. Beta2 Adrenergic-Neurotrophin Feedforward Loop Promotes Pancreatic Cancer. Cancer Cell 2018, 33, 75–90e77. [Google Scholar] [CrossRef] [PubMed]
- Kappos, E.A.; Engels, P.E.; Tremp, M.; Sieber, P.K.; von Felten, S.; Madduri, S.; Meyer Zu Schwabedissen, M.; Fischmann, A.; Schaefer, D.J.; Kalbermatten, D.F. Denervation leads to volume regression in breast cancer. J. Plast. Reconstr. Aesthet. Surg. 2018, 71, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.W.; Nagaraja, A.S.; Lutgendorf, S.K.; Green, P.A.; Sood, A.K. Sympathetic nervous system regulation of the tumour microenvironment. Nat. Rev. Cancer 2015, 15, 563–572. [Google Scholar] [CrossRef]
- Tibensky, M.; Mravec, B. Role of the parasympathetic nervous system in cancer initiation and progression. Clin. Transl. Oncol. 2021, 23, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Lackovicova, L.; Gaykema, R.P.; Banovska, L.; Kiss, A.; Goehler, L.E.; Mravec, B. The time-course of hindbrain neuronal activity varies according to location during either intraperitoneal or subcutaneous tumor growth in rats: Single Fos and dual Fos/dopamine beta-hydroxylase immunohistochemistry. J. Neuroimmunol. 2013, 260, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Horvathova, L.; Tillinger, A.; Padova, A.; Bizik, J.; Mravec, B. Changes in gene expression in brain structures related to visceral sensation, autonomic functions, food intake, and cognition in melanoma-bearing mice. Eur. J. Neurosci. 2020, 51, 2376–2393. [Google Scholar] [CrossRef] [PubMed]
- Szpunar, M.J.; Belcher, E.K.; Dawes, R.P.; Madden, K.S. Sympathetic innervation, norepinephrine content, and norepinephrine turnover in orthotopic and spontaneous models of breast cancer. Brain Behav. Immun. 2016, 53, 223–233. [Google Scholar] [CrossRef]
- Zahalka, A.H.; Frenette, P.S. Nerves in cancer. Nat. Rev. Cancer 2020, 20, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Mauffrey, P.; Tchitchek, N.; Barroca, V.; Bemelmans, A.P.; Firlej, V.; Allory, Y.; Romeo, P.H.; Magnon, C. Progenitors from the central nervous system drive neurogenesis in cancer. Nature 2019, 569, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Amit, M.; Takahashi, H.; Dragomir, M.P.; Lindemann, A.; Gleber-Netto, F.O.; Pickering, C.R.; Anfossi, S.; Osman, A.A.; Cai, Y.; Wang, R.; et al. Loss of p53 drives neuron reprogramming in head and neck cancer. Nature 2020, 578, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rutlin, M.; Huang, S.; Barrick, C.A.; Wang, F.; Jones, K.R.; Tessarollo, L.; Ginty, D.D. Sexually dimorphic BDNF signaling directs sensory innervation of the mammary gland. Science 2012, 338, 1357–1360. [Google Scholar] [CrossRef]
- Wang, Z.; Dong, J.; Tian, W.; Qiao, S.; Wang, H. Role of TRPV1 ion channel in cervical squamous cell carcinoma genesis. Front. Mol. Biosci. 2022, 9, 980262. [Google Scholar] [CrossRef]
- Zheng, L.; Dou, X.; Song, H.; Gao, R.; Tang, X. TRPV1 acts as a Tumor Suppressor and is associated with Immune Cell Infiltration in Clear Cell Renal Cell Carcinoma: Evidence from integrated analysis. J. Cancer 2020, 11, 5678–5688. [Google Scholar] [CrossRef]
- Nie, R.; Liu, Q.; Wang, X. TRPV1 Is a Potential Tumor Suppressor for Its Negative Association with Tumor Proliferation and Positive Association with Antitumor Immune Responses in Pan-Cancer. J. Oncol. 2022, 2022, 6964550. [Google Scholar] [CrossRef]
- Balood, M.; Ahmadi, M.; Eichwald, T.; Ahmadi, A.; Majdoubi, A.; Roversi, K.; Roversi, K.; Lucido, C.T.; Restaino, A.C.; Huang, S.; et al. Nociceptor neurons affect cancer immunosurveillance. Nature 2022, 611, 405–412. [Google Scholar] [CrossRef]
- Kwon, D.H.; Zhang, F.R.; Fedor, J.G.; Suo, Y.; Lee, S.-Y. Vanilloid-dependent TRPV1 opening trajectory from cryoEM ensemble analysis. Nat. Commun. 2022, 13, 2874. [Google Scholar] [CrossRef]
- Fujimura, S.; Kazuhiro, K.; Kuramochi, M.; Sekiguchi, H.; Ikezaki, K.; Mio, M.; Hengphasatporn, K.; Shigeta, Y.; Kubo, T.; Sasaki, Y.C. Agonist and antagonist-diverted twisting motions of a single TRPV1 channel. J. Phys. Chem. B 2020, 124, 11617–11624. [Google Scholar] [CrossRef]
- Zhang, K.; Julius, D.; Cheng, Y. Structural snapshots of TRPV1 revel mechanism of polymodal functionality. Cell 2021, 184, 5138–5150. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Nguyen, P.T.; Vu, S.; Yarov-Yarovoy, V.; Zheng, J. Opening of capsaicin receptor TRPV1 is stabilized equally by its four subunits. J. Biol. Chem. 2023, 15, 104828. [Google Scholar] [CrossRef]
- Wang, C.; Hu, H.-Z.; Colton, C.K.; Wood, J.D.; Zhu, M.X. An alternative splicing product of the murine trpv1 gene dominant negatively modulates the activity of the TRPV1 channel. J. Biol. Chem. 2004, 279, 37423–37430. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sanz, N.; Fernández-Carjaval, A.; Moranelle-Palao, C.; Planells-Cases, R.; Fajardo-Sánchez, E.; Fernández-Ballester, G.; Ferrer-Montiel, A. Identification of a tetramerization domain in the C terminus of the vanilloid receptor. J. Neurosci. 2004, 24, 5307–5314. [Google Scholar] [CrossRef]
- Sadovsky, L.R.; Sreekhrisna, K.T.; Schinaman, R.; Gorka, K.; Mantri, Y.; Haught, J.C.; Huggins, T.G.; Isfort, R.J. Unique responses are observed in transient receptor potential ankyrin 1 and vanilloid 1 (TRPA1 and TRPV1) co-expressing cells. Cells 2014, 3, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Zheng, G.; Wiley, J.W. Epigenetic regulation of genes that modulate chronic stress-induced visceral pain in the peripheral nervous system. Gastroenterology 2015, 148, 148–157. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, Y.; Tian, Q.; Deng, Q.; Wang, Y.; Zhou, T.; Liu, Q.; Mei, K.; Wang, Y.; Liu, H.; et al. TRPV1 SUMOylation regulates nociceptive signaling in models of inflammatory pain. Nat. Commun. 2018, 9, 1529. [Google Scholar] [CrossRef]
- Premkumar, L.S.; Ahern, G.P. Induction of vanilloid receptor channel activity by protein kinase C. Nature 2000, 408, 985–990. [Google Scholar] [CrossRef]
- Hellwig, N.; Albrech, N.; Harteneck, C.; Schultz, G.; Schaefer, M. Homo- and heteromeric assembly of TRPV channel subunits. J. Cell. Sci. 2005, 118, 917–928. [Google Scholar] [CrossRef]
- Planells-Cases, R.; Valente, P.; Ferrer-Montiel, A.; Feng, Q.; Szallasi, A. Complex regulation of TRPV1 and related thermoTRPs: Implications for therapeutic intervention. Adv. Exp. Med. Biol. 2011, 704, 491–515. [Google Scholar]
- Maggi, F.; Morelli, M.B.; Aguzzi, C.; Zeppa, L.; Nabissi, M.; Polidori, C.; Santoni, G.; Amantini, C. Calcium influx, oxidative stress, and apoptosis induced by TRPV1 in chronic myeloid leukemia cells: Synergic effects with imatinib. Front. Mol. Biosci. 2023, 10, 1129202. [Google Scholar] [CrossRef] [PubMed]
- Patapoutian, A.; Tate, S.; Wood, C.J. Transient receptor potential chanels: Targeting pain at the source. Nat. Rev. Drug Discov. 2009, 8, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.; Córdova, C.; Marchant, I.; Zúniga, R.; Ochova, P.; Ramirez-Barrantes, R.; González-Arriagada, W.A.; Rodriguez, B.; Olivero, P. Intracellular aggregated TRPV1 is associated with lowe survival in breast cancer patients. Breast Cancer 2018, 10, 161–168. [Google Scholar]
- Surh, Y.J.; Lee, S.S. Capsaicin, a double-edged sword: Toxicity, metabolism, and chemopreventive potential. Life Sci. 1995, 56, 1845–1855. [Google Scholar] [CrossRef]
- Torrecillas, A.; Schneider, A.; Fernández-Martínez, A.M.; Ausili, A.; de Godos, A.M.; Corbalán-Garcia, S.; Gómez-Fernández, J.C. Capsaicin fluidifies membrane and localizes itself near the lipid-water interface. ACS Chem. Neurosci. 2015, 6, 1741–1750. [Google Scholar] [CrossRef]
- Bleakman, D.; Brorson, J.R.; Miller, R.J. The effect of capsaicin on voltage-gated calcium currents and calcium sigbnals in cultures dorsal root ganglion cells. Br. J. Pharmacol. 1990, 101, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kwon, H.J.; Kim, G.E.; Cho, M.H.; Yoon, S.Y.; Davies, A.J.; Oh, S.B.; Lee, H.; Cho, Y.K.; Joo, C.H.; et al. Attenuation of natural killer cell functions by capsaicin through a direct and TRPV1-independent mechanism. Carcinogenesis 2014, 35, 1652–1660. [Google Scholar] [CrossRef] [PubMed]
- Erin, N.; Akman, M. Effects of in-vitro modulation of TRPV1 activity on immune response of mice bearing metastatic breast carcinoma: Enhanced inflammatory response may hinder therapeutic potentials of TRPV1 agonists. Life Sci. 2021, 287, 120115. [Google Scholar] [CrossRef]
- Marek-Jozefowicz, L.; Nedoszytko, B.; Grochocka, M.; Zmijewski, M.A.; Czajkowski, R.; Cubala, W.L.; Slominski, A.T. Molecular mechanisms of neurogenic inflammation of the skin. Int. J. Mol. Sci. 2023, 24, 5001. [Google Scholar] [CrossRef]
- Majhi, R.K.; Sahoo, S.S.; Yadav, M.; Pratheek, B.M.; Chattopadhyay, S.; Goswami, C. Functional expression of TRPV channels in T cells and their implications in immune regulation. FEBS J. 2015, 282, 2661–2681. [Google Scholar] [CrossRef]
- Kunde, D.A.; Yingchoncharoen, J.; Jurkovic, S.; Geraghty, D.P. TRPV1 mediates capsaicin-stimulated metabolic activity but not cell death or inhibition of interleukin-1beta release in human THP-1 monocytes. Toxicol. Appl. Pharmacol. 2018, 360, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Sanjai Kumar, P.; Nayak, T.K.; Mahish, C.; Sahoo, S.S.; Radhakrishnan, A.; De, S.; Datey, A.; Sahu, R.P.; Goswami, C.; Chattopadhyay, S.; et al. Inhibition of transient receptor potential vanilloid 1 (TRPV1) channel regulates chikungunya virus infection in macrophages. Arch. Virol. 2021, 166, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Saunders, C.I.; Kunde, D.A.; Crawford, A.; Geraghty, D.P. Expression of transient receptor potential vanilloid 1 (TRPV1) and 2 (TRPV2) in human peripheral blood. Mol. Immunol. 2007, 44, 1429–1435. [Google Scholar] [CrossRef]
- Basu, S.; Srivastava, P. Immunological role of neuronal vanilloid receptor 1 expressed on dendritic cells. Proc. Natl. Acad. Sci. USA 2005, 102, 5120–5125. [Google Scholar] [CrossRef] [PubMed]
- Amantini, C.; Farfariello, V.; Cardinali, C.; Morelli, M.B.; Marinelli, O.; Nabissi, M.; Santoni, M.; Bonfili, L.; Cecarini, V.; Eleuteri, A.M.; et al. The TRPV1 ion channel regulates thymocyte differentiation by modulating autophagy and proteasome activity. Oncotarget 2017, 8, 90766–90780. [Google Scholar] [CrossRef]
- Hogan, P.G.; Lewis, R.S.; Rao, A. Molecular basis of calcium signaling in lymphocytes: STIM and ORAI. Annu. Rev. Immunol. 2010, 28, 491–533. [Google Scholar] [CrossRef]
- Inada, H.; Iida, T.; Tominaga, M. Different expression patterns of TRP genes in murine B and T lymphocytes. Biochem. Biophys. Res. Commun. 2006, 350, 762–767. [Google Scholar] [CrossRef]
- Zhang, F.; Challapalli, S.C.; Smith, P.J. Cannabinoid CB(1) receptor activation stimulates neurite outgrowth and inhibits capsaicin-induced Ca2+ influx in an in vitro model of diabetic neuropathy. Neuropharmacology 2009, 57, 88–96. [Google Scholar] [CrossRef]
- Macho, A.; Calzado, M.A.; Munoz-Blanco, J.; Gomez-Diaz, C.; Gajate, C.; Mollinedo, F.; Navas, P.; Munoz, E. Selective induction of apoptosis by capsaicin in transformed cells: The role of reactive oxygen species and calcium. Cell Death Differ. 1999, 6, 155–165. [Google Scholar] [CrossRef]
- Bertin, S.; Aoki-Nonaka, Y.; Lee, J.; de Jong, P.R.; Kim, P.; Han, T.; Yu, T.; To, K.; Takahashi, N.; Boland, B.S.; et al. The TRPA1 ion channel is expressed in CD4+ T cells and restrains T-cell-mediated colitis through inhibition of TRPV1. Gut 2017, 66, 1584–1596. [Google Scholar] [CrossRef]
- Baker, K.; Raemdonck, K.; Dekkak, B.; Snelgrove, R.J.; Ford, J.; Shala, F.; Belvisi, M.G.; Birrell, M.A. Role of the ion channel, transient receptor potential cation channel subfamily V member 1 (TRPV1), in allergic asthma. Respir. Res. 2016, 17, 67. [Google Scholar] [CrossRef]
- Amantini, C.; Mosca, M.; Lucciarini, R.; Perfumi, M.; Morrone, S.; Piccoli, M.; Santoni, G. Distinct thymocyte subsets express the vanilloid receptor VR1 that mediates capsaicin-induced apoptotic cell death. Cell Death Differ. 2004, 11, 1342–1356. [Google Scholar]
- Takano, F.; Yamaguchi, M.; Takada, S.; Shoda, S.; Yahagi, N.; Takahashi, T.; Ohta, T. Capsicum ethanol extracts and capsaicin enhance interleukin-2 and interferon-gamma production in cultured murine Peyer’s patch cells ex vivo. Life Sci. 2007, 80, 1553–1563. [Google Scholar] [CrossRef]
- Kumar, P.S.; Mukherjee, T.; Khamaru, S.; Radhakrishnan, A.; Nanda-Kishore, D.J.; Chawla, S.; Sahoo, S.S.; Chattopadhyay, S. Elevation of TRPV1 expression on T-cells during experimental immunosuppression. J. Biosci. 2022, 47, 42. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Brito, C.X.; Teixeira, S.A.; Barboza, R.; dos Reis, A.S.; Azevedo-Santos, A.P.; Muscara, M.; Costa, S.K.; Marinho, C.R.; Brain, S.D.; et al. TRPV1 antagonism by capsazepine modulates innate immune response in mice infected with Plasmodium berghei ANKA. Mediat. Inflamm. 2014, 2014, 506450. [Google Scholar] [CrossRef] [PubMed]
- Tóth, B.I.; Benkó, S.; Szőllősi, A.G.; Kovács, L.; Rajnavölgyi, E.; Bíró, T. Transient receptor potential vanilloid-1 signaling inhibits differentiation and activation of human dendritic cells. FEBS Lett. 2009, 583, 1619–1624. [Google Scholar] [CrossRef] [PubMed]
- Koebel, C.M.; Vermi, W.; Swann, J.B.; Zerafa, N.; Rodig, S.J.; Old, L.J.; Smyth, M.J.; Schreiber, R.D. Adaptive immunity maintains occult cancer in an equilibrium state. Nature 2007, 450, 903–907. [Google Scholar] [CrossRef]
- Zou, W.; Restifo, N.P. T(H)17 cells in tumour immunity and immunotherapy. Nat. Rev. Immunol. 2010, 10, 248–256. [Google Scholar] [CrossRef]
- Chopan, M.; Littenberg, B. The Association of Hot Red Chili Pepper Consumption and Mortality: A Large Population-Based Cohort Study. PLoS ONE 2017, 12, e0169876. [Google Scholar] [CrossRef]
- Bonaccio, M.; Di Castelnuovo, A.; Costanzo, S.; Ruggiero, E.; De Curtis, A.; Persichillo, M.; Tabolacci, C.; Facchiano, F.; Cerletti, C.; Donati, M.B.; et al. Chili Pepper Consumption and Mortality in Italian Adults. J. Am. Coll. Cardiol. 2019, 74, 3139–3149. [Google Scholar] [CrossRef]
- Ryan, D.P.; Hong, T.S.; Bardeesy, N. Pancreatic adenocarcinoma. N. Engl. J. Med. 2014, 371, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Stopczynski, R.E.; Normolle, D.P.; Hartman, D.J.; Ying, H.; DeBerry, J.J.; Bielefeldt, K.; Rhim, A.D.; DePinho, R.A.; Albers, K.M.; Davis, B.M. Neuroplastic changes occur early in the development of pancreatic ductal adenocarcinoma. Cancer Res. 2014, 74, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Swidnicka-Siergiejko, A.K.; Gomez-Chou, S.B.; Cruz-Monserrate, Z.; Deng, D.; Liu, Y.; Huang, H.; Ji, B.; Azizian, N.; Daniluk, J.; Lu, W.; et al. Chronic inflammation initiates multiple forms of K-Ras-independent mouse pancreatic cancer in the absence of TP53. Oncogene 2017, 36, 3149–3158. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, T.; Hiasa, M.; Okui, T.; Hata, K. Sensory nerves: A driver of the vicious cycle in bone metastasis? J. Bone Oncol. 2021, 30, 100387. [Google Scholar] [CrossRef]
- Brown, D.C.; Iadarola, M.J.; Perkowski, S.Z.; Erin, H.; Shofer, F.; Laszlo, K.J.; Olah, Z.; Mannes, A.J. Physiologic and antinociceptive effects of intrathecal resiniferatoxin in a canine bone cancer model. Anesthesiology 2005, 103, 1052–1059. [Google Scholar] [CrossRef]
- Brown, D.C.; Agnello, K.; Iadarola, M.J. Intrathecal resiniferatoxin in a dog model: Efficacy in bone cancer pain. Pain 2015, 156, 1018–1024. [Google Scholar] [CrossRef]
- Erin, N.; Zhao, W.; Bylander, J.; Chase, G.; Clawson, G. Capsaicin-induced inactivation of sensory neurons promotes a more aggressive gene expression phenotype in breast cancer cells. Breast Cancer Res. Treat. 2006, 99, 351–364. [Google Scholar] [CrossRef]
- Kissin, I. Vanilloid-induced conduction analgesia: Selective, dose-dependent, long-lasting, with a low level of potential neurotoxicity. Anesth. Analg. 2008, 107, 271–281. [Google Scholar] [CrossRef]
- Szallasi, A.; Cortright, D.N.; Blum, C.A.; Eid, S.R. The vanilloid receptor TRPV1: 10 years from channel cloning to antagonist proof-of-concept. Nat. Rev. Drug Discov 2007, 6, 357–372. [Google Scholar] [CrossRef]
- Gavva, N.R.; Treanor, J.J.; Garami, A.; Fang, L.; Surapaneni, S.; Akrami, A.; Alvarez, F.; Bak, A.; Darling, M.; Gore, A.; et al. Pharmacological blockade of the vanilloid receptor TRPV1 elicits marked hyperthermia in humans. Pain 2008, 136, 202–210. [Google Scholar] [CrossRef]
- Garami, A.; Shimansky, Y.P.; Rumbus, Z.; Vizin, R.C.L.; Farkas, N.; Hegyi, J.; Szakacs, Z.; Solymar, M.; Csenkey, A.; Chiche, D.A.; et al. Hyperthermia induced by transient receptor potential vanilloid-1 (TRPV1) antagonists in human clinical trials: Insights from mathematical modeling and meta-analysis. Pharmacol. Ther. 2020, 208, 107474. [Google Scholar] [CrossRef] [PubMed]
- Toth, B.; Rogan, E.; Walker, B. Tumorigenicity and mutagenicity studies with capsaicin of hot peppers. Anticancer Res. 1984, 4, 117–119. [Google Scholar] [PubMed]
- Bode, A.M.; Dong, Z. The two faces of capsaicin. Cancer Res. 2011, 71, 2809–2814. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).