Abstract
In humans, a total of 12 galectins have been identified. Their intracellular and extracellular biological functions are explored and discussed in this review. These galectins play important roles in controlling immune responses within the tumour microenvironment (TME) and the infiltration of immune cells, including different subsets of T cells, macrophages, and neutrophils, to fight against cancer cells. However, these infiltrating cells also have repair roles and are hijacked by cancer cells for pro-tumorigenic activities. Upon a better understanding of the immunomodulating functions of galectin-3 and -9, their inhibitors, namely, GB1211 and LYT-200, have been selected as candidates for clinical trials. The use of these galectin inhibitors as combined treatments with current immune checkpoint inhibitors (ICIs) is also undergoing clinical trial investigations. Through their network of binding partners, inhibition of galectin have broad downstream effects acting on CD8+ cytotoxic T cells, regulatory T cells (Tregs), Natural Killer (NK) cells, and macrophages as well as playing pro-inflammatory roles, inhibiting T-cell exhaustion to support the fight against cancer cells. Other galectin members are also included in this review to provide insight into potential candidates for future treatment(s). The pitfalls and limitations of using galectins and their inhibitors are also discussed to cognise their clinical application.
1. Introduction
The function of the immune system in fighting cancer cells has been of long-standing interest in the context of cancer therapies [1], with research dating back to William B. Coley’s study in 1819 [2]. Since the first ICI, Cytotoxic-T-lympocyte-antigen-4 (CTLA-4), also known as ipilimumab (Yervoy), was tested and approved for the treatment of metastatic melanoma in 2015 [3,4], the number of checkpoint inhibitors has increased. In particular, when programmed cell death/ligand-1 (PD-1/PD-L1) immune checkpoint proteins are identified, these checkpoint inhibitors are now the standard regimen for immuno-oncology (I-O) therapy when tackling different solid tumours [5]. The most common PD-1 and PD-L1 inhibitors are pembrolizumab (Keytruda), nivolumab (Opdivo), cemiplimab (Libtayo), atezolizumab (Tecentriq), nivolumab (Bavencio), and durvalumab (Imfinzi) [6]. In addition, ipilimumab (Yervoy) is the most common CTLA-4 inhibitor. However, the response rates (RRs) of this approach vary according to types and lines of treatment [7]. It has been found to have a good-to-moderate response, with over a 50% RR against classic Hodgkin’s lymphoma, melanoma, and first-line combination-treated non-small cell lung cancer (NSCLC). However, some cancers, such as extensive-stage small cell lung cancer (SCLC), hepatocellular carcinoma (HCC), PD-L1+ gastric (gastroesophageal junction type), and cervical cancers, have shown less than a 25% RR.
The current challenges to the efficiency of I-O therapies include the exhaustion of cytotoxic T cells [8] and the need to increase the subpopulation of Tregs and other immune cells during immunosuppression [9]. To combat these challenges, other immunomodulators have been identified, such as T-cell immunoreceptor with immunoglobulin and ITIM domains (TIGIT). TIGIT is found in T cells, NK cells, and tumour cells. The mechanism of immune inactivation may occur through ITIM-dependent negative pathways [10]. Numerous TIGIT antibodies, including BMS-986207, tiragolumab, and vibostolimab, have been developed and tested in clinical trials [11]. The recent results of these trials, such as CITYSCRAPE [12], have generated additional interest in investigating any novel immunosuppressors and their interacting protein mechanisms. The results of the subsequent phase III trials, such as the SKYSCRAPER series (NCT04619707, NCT04513925, NCT046665843, etc.), will need further analysis to enhance this approach to treatment. A list of this series of studies was summarised by Brazel et al. (2023) [13].
Recently, galectins have been identified as immunomodulators [14], joining TIGIT as new potential targets for immunotherapies. The potential candidate reagents and ongoing trial studies are listed in Table 1. In particular, newly developed reagents, such as GB1211 [15], a galectin-3 small-molecule inhibitor, and LYT-200, an anti-galectin-9 humanised antibody, are currently on trial (NCT05240131/GALLANT-1 and NCT04666688, respectively). The safety of GB1211 has also been reported, with limited grade 1 and grade 2 adverse effects in healthy participants (NCT03809052) [16]. The efficiency of these candidates in the current trial studies will further support the use of galectins in cancer treatments in combination with PD-1/PD-L1 and TIGIT. Other galectin inhibitors, such as OTX008 and ProLectin-M, have also been investigated in mouse animal model studies [17,18,19,20]. Other newly identified small-molecule/peptide inhibitors and antibodies also represent new potential agents for treatment and are discussed in Section 3.1 below.

Table 1.
Clinical trial studies (extracted from ClinicalTrials.gov, accessed on 11 May 2023).
2. Human Galectins
There are 12 galectins in humans, as listed in Table 2. The genomic locations of these genes and their protein product structures are shown in Figure 1 and Figure 2, respectively. Galectins can be detected in the cytoplasm and exhibit a secretory form. Protein topology analysis [24] has shown that conventional secretory proteins, which contain an N-terminal signal peptide that starts with a few positively charged amino acids, such as lysine (K) and arginine (I), followed by around 12–16 hydrophobic amino acids [25], are driven into the endoplasmic reticulum (ER). The secretory proteins are then embedded inside lipid bilayer vesicles and transported via budding-off into the Golgi apparatus, after which the vesicles can be further fused with the cell membrane to export the protein outside the cell [26]. However, further analysis of the coding and protein sequence of galectins has resulted in no signal peptide sequence being detected. Their secretory forms are suspected to be produced through the non-canonical secretion pathway [27,28]. Unlike other proteins, their recognition is not based on protein peptides in the form of amino acid chains on the binding partner(s). Galectins contain a carbohydrate-recognition domain (CRD) as a binding motif to recognise the glycosylation sites on other proteins and for binding. The CRD mainly detects glycoproteins and is required for post-translational modification. Their immunological roles have been established and reviewed [29]. Different galectins have been identified that bind with immune cells, such as cytotoxic T cells, dendritic cells, and macrophages, to regulate cancer cell immunosurveillance (also listed in Table 2). The roles of these human galectins are discussed in further detail below.
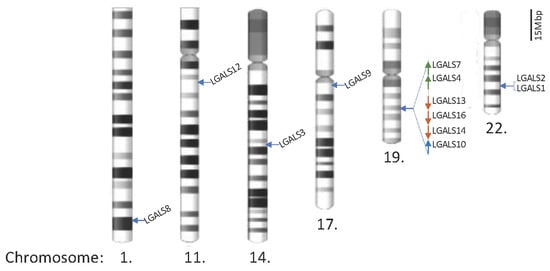
Figure 1.
Locations of galectin genes in the human genome. Twenty galectin genes have been identified and are located on chromosomes 1, 11, 14, 17, 19, and 22. The partial q arms of chromosomes 1 and 11 are only shown to reflect the scale, and the scale bar represents 15 megabase pairs (Mbp) in gene distance.
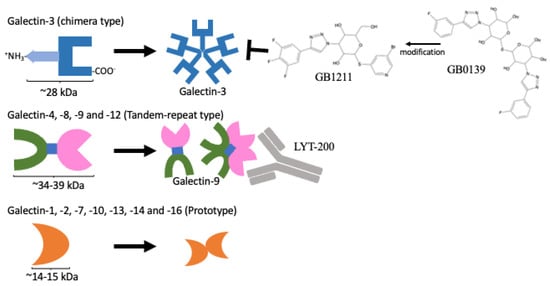
Figure 2.
Protein domain structures of galectins and their inhibitors. Only the galectin-3 protein contains CRD domain and an extra amino domain allowing it to form an oligomer (upper panel). A protein containing two distinct CRDs, galectin-4, 6, 8, 9, and 12 (middle panel). A single CRD protein that can form homo-dimers, galectin-1, 2, 5, 7, 10, 11, 13, 14, and 16 (lower panel). GB1211 and LYT-200 represent a newly developed galectin-3-specific inhibitor [15] and a humanised monoclonal antibody against galectin-9, respectively, and are currently under clinical trial.
2.1. Galectin-3: The Only Chimera-Type Galectin with Oncogenic Functioning
Among all the members of the galectin family, only galectin-3 contains a 12-amino-acid N-terminus with a serine (S) residue at position six for phosphorylation, followed by a 100-amino-acid collagen-like sequence (CLS) domain and a 130-amino-acid CRD at the C-terminus [30] as a single protein unit of about 28 kDa in size. The N-terminus of galectin-3 facilitates the formation of a multi-unit chimera complex, as shown in Figure 2. Post-translational modifications, such as phosphorylation on serine 6, can promote tumour aggressiveness and metastasis, whereas mutating this residue into alanine (A) caused the tumorigenicity of breast cancer cells implanted in nude mice to be abolished [31]. In terms of its intracellular roles, galectin-3 has been found to bind different interacting partners to control tumorigenicity. It can bind with K-Ras [32] to activate downstream signalling pathways, including PI3K/Akt, PLC/PKC, Raf/MEK/ERK, RALGDS/Ral, and TIAM1/Rac, to promote cancer cell survival, migration, invasion, and cell cycle progression through the hijacking of the upstream receptor tyrosine kinases, such as epidermal growth factor receptor (EGFR) and insulin-like growth factor receptors (IGFRs), as well as G-protein coupling receptors (GPCRs) [33]. In addition, it can also interact with the proline-rich region (PRR) of ALG-2-interacting protein X (Alix) for an endosomal-sorting complex functioning [34,35]. Alix-depleted basal-like breast cancer cells have also been shown to induce EGFR activity and EGFR-dependent PD-L1 presentation [36]. In addition to these cytoplastic roles, galectin-3 can also interact with the β-catenin signalling component PCDH24 and spliceosome complex member GEMIN4 to regulate gene expression and alter isoform expression [37,38]. Galectin-3 has also been found to upregulate p21 expression via Sp1 binding to control the cell cycle progression [39].
Interestingly, galectin-3 also plays an extracellular role in controlling cancer growth. In NSCLC, inhibition of galectin-3 increases the oxidative stress in vitro (in A549 and H1792 cell lines) and PDX models [40]. Galectin-3 acts on the integrin αvβ3 extracellularly via K-Ras to promote oncogenesis. In contrast, overexpression of β3 in the null cell line H727 promoted the growth of cancer cells, which were perturbed by shRNA KRAS or galectin-3 knockdown.
In addition to galectin-3 promoting pro-oncogenic roles in cancer cells, it also acts on other immune cells for immunosuppression. In healthy individuals, galectin-3 plays an important role in systemic innate immunity for pathogen recognition as a pattern-recognition receptor (PRR)/danger-associated molecular pattern (DAMP) and is secreted by myeloid cells, including neutrophils, monocytes, macrophages, and dendritic cells (DCs) [41]. According to binding assays of galectin-3 and chemokines, galectin-3 can bind to numerous chemokines, including CCL1, -5, -13, -19, -20, -21, -22, -24, -25, -26, -27, and -28 and CXCL7, -10, -12, -13, -14, -16, and -17 to perturb the migration properties of different immune cells [42]. Galectin-3 also plays a direct role in killing effector T cells through CD71 and integrin-associated protein (IAP) binding [43]. It can also inhibit macrophage cytotoxicity and phagocytosis [44]. Cancer cells secrete galectin-3 within the TME to serve in a general immunosuppressive role.
Numerous galectin-3 inhibitors have been identified that tackle the immunomodulating role of immune cells. Belpectin (GR-MD-02) was developed to prevent non-alcoholic steatohepatitis (NASH) cirrhosis [45], which is a risk factor for developing HCC [46]. GB1211 is the first oral bioavailable galectin-3 inhibitor with an affinity to human galectin-3 at Kd = 0.025 μM. It was modified from the inhaled galectin-3 inhibitor GB0139 for the treatment of idiopathic pulmonary fibrosis [15] (Figure 2). Belpectin and GB0139 were developed to inhibit fibrosis via the control of galectin-3 [47]. A stage I clinical trial study saw pembrolizumab combined with belpectin against metastatic melanoma or head and neck squamous cell carcinoma (HNSCC) (NCT02575404, listed in Table 1), and the safety and efficacy were assessed [21]. The results were promising; the combination reactivated effective memory T cells and reduced suppressive monocytic myeloid cells, showing mainly grade I adverse effects (with no grade III and IV). Another galectin-3 inhibitor, GB1211, has been combined with atezolizumab for use against NSCLC (stage I/II) and is undergoing trials. The results and their extension to phases II and III will provide further insight into the applicability of galectin-3 as a combinatorial treatment with standard ICI (PD-1/PD-L1).
Table 2.
Intracellular and extracellular binding partners of 12 human galectins.
Table 2.
Intracellular and extracellular binding partners of 12 human galectins.
| Gene/Protein Name (Chromosome Position [48]) | Intracellular (Cytoplasmic/Nucleus) | Extracellular |
|---|---|---|
| LGALS1/Galectin-1 (Chr. 22q13.1) | Cytoplasmic: GRP78 [49] Gemin4 [37] H-Ras [50] PCDH24 [38] | CC and CXC chemokines [42] CD43 [51,52] CD45 [51,53] NRP1 [54] VEGFR2 [55] |
| LGALS2/Galectin-2 (Chr. 22q13.1) | Binds to surface of CD14(interm.–high) monocyte and promote M1 macrophage differentiation [56] | |
| LGALS3/Galectin-3 (Chr. 14q22.3) | Cytoplasmic: Alix (EGFR trafficking) [34,35,57] Gemin4 [37] K-Ras [32,40] PCDH24 [38] Nucleus: hnRNPA2B1 [58] Sp1 [39] | CC and CXC chemokines [42] CD29 [43] CD43 [43] CD45 [43] CD71 [43] EGFR [59] Interferon-γ [60] Integrin αvβ3 [40] LAG3 [61] MUC1 [62] |
| LGALS4/Galectin-4 (Chr. 19q13.2) | CD3 [63] | |
| LGALS7/Galectin-7 (Chr. 19q13.2) | Cytoplasmic: Bcl-2 [64] | |
| LGALS8/Galectin-8 (Chr. 1q43) | αM (CD11b, neutrophils) [65] CD166 [66] Podoplanin [67,68] | |
| LGALS9/Galectin-9 (Chr. 17q11.2) | Cytoplasmic: Binding to intracellular TIM-3 to modulate mTOR phosphorylation [69] Cytoplasmic–Lysosomes: Interact with Lamp2 to regulate lysosomal functions and autophagy [70] | 4-1BB [71] CD40 [72] CD44 [73] CD206 [74] Dectin-1 (macrophages) [75] DR3 [76] PD-1 [77] PDI [78,79] TCR [80,81] TIM-3 [77,82,83] VISTA [84] |
| LGALS10/Galectin-10/Charcot-Leyden crystal protein CLC (Chr. 19q13.2) | Cytoplasmic–Granules: Eosinophil-derived neurotoxin EDN (RNS2) and eosinophil cationic protein ECP (RNS3) co-localised with CD63. It is required for the maturation of eosinophil during granulogenesis [85] | |
| LGALS12/Galectin-12/GRIP1 (Chr. 11q12.3) | Cytoplasmic–Endosome/Lysosomes: VPS13C in lipid droplets and promotes the polarisation to M1 macrophage via TLR4 pathway [86,87] | |
| LGALS13/Galectin-13/ placental protein 13 (Chr. 19q13.2) | Nucleus: HOXA1 [88] | Binds to T lymphocytes and induces apoptosis [89]; Binds to neutrophils and shifts to immunoregulatory phenotype and promotes high PD-L1 expression [90] |
| LGALS14/Galectin-14 (Chr. 19q13.2) | Binds to T lymphocytes and induces apoptosis [89] c-Rel [91] | |
| LGALS16/Galectin-16 (Chr. 19q13.2) | c-Rel [92] |
Remarks: Colour code is based on the galectin’s structure: chimera type (galectin-3) is highlighted in light blue; tandem-repeat type is highlighted in light green; prototype has not been highlighted.
2.2. Tandem-Repeat Type Galectins
2.2.1. Galectin-9 Acts on the Immunosuppression of Cancers
Galectin-9 was first identified in 1997 in embryonic mouse kidneys as a 36 kDa protein [93] with two distinct CRDs (as shown in Figure 2) and is associated with human Hodgkin’s lymphoma [94]. It has been shown to be overexpressed in different cancers (Table 3). High mobility group box 1 (HMGB1) has been demonstrated to induce the Toll-like receptor 4-mediated pathway to promote galectin-9 expression in cancer cells [95]. However, associations between the overexpression of galectin-9 in tumour tissues (immunohistochemistry (IHC) staining) and clinicopathological/survival parameters have been quite controversial in different studies [96,97]. The galectin-9 that is produced by cancer cells seems to work dynamically with the surrounding cells. In pancreatic adenocarcinoma, the expression of galectin-9 is positively correlated with PD-L1 in tumour tissues [97]. Interestingly, the serum level of galectin-9 is somehow reflective of the staging of colon cancer and NSCLC patients [98]. Galectin-9 may be a good diagnostic and prognostic marker for cancer staging. However, the role of systemic serum galectin-9 is still elusive. In contrast, galectin-9 overexpression in isolated tumour-infiltrating lymphocytes (TILs) at a tumour site represents T-cell exhaustion and impairment in NK cells [99]. Hence, the roles of infiltrating and peripheral leukocytes may be different within the TME and systemic immunity, respectively.
Table 3.
Expression in different cancers [100,101,102,103,104].
The interacting partners of galectin-9 have been identified and are mainly expressed on the cell surface of different immune cells, including NK cells, effector T cells, Tregs, monocytes, macrophages, and neutrophils (Table 2 and Figure 3). Galectin-9 studies have revealed an immunoregulation [105] within the TME. These molecules are displayed as transmembrane proteins that transduce signals from the cell surface to intracellular effectors to control immune cell proliferation, survival, and cytokine secretion [106]. All these molecules contain either an immunoreceptor tyrosine-based activation motif (ITAM) or an inhibitory motif (ITIM) domain that further activates or inhibits downstream activities, respectively. ITAM-containing proteins recruit Src/Lck proteins for phosphorylation to activate downstream processes. In contrast, ITIM-containing proteins recruit phosphoinositide phosphatase (SHIP) proteins to dephosphorylate the downstream components via hydrolysation for the inactivation [107]. Galectin-9 has been identified to bind to PD-1 and TIM-3 in CD8+ cytotoxic T cells [77,82]. In addition, it was also shown to interact with T-cell receptor (TCR) on Treg, which has also been found to suppress cytotoxic T via the galectin-9–TIM-3 axis in HIV contexts [108]. However, in T-cell/tumour-cell co-culture environments, galectin-9 induces T-cell apoptosis, which can be blocked by galectin-9 antibodies [80]. The dominant role of galectin-9 in the TME may still be pro-tumorigenic. TIM-3 is also an ITIM-containing protein that suppresses the proliferation of effector T cells, NK cells, and monocytes, which is similar to the roles of TIGIT, where TIGIT acts mainly on effector-T and NK cells (Figure 3) [109]. Other ITIM molecules, including CD40, VISTA, and 4-1BB, are activated by galectin-9 to suppress the expansion of effector T cells [71,72,73,84]. In addition, galectin-9 acts on VISTA to also suppress human cytotoxic T lymphocyte activity [84].
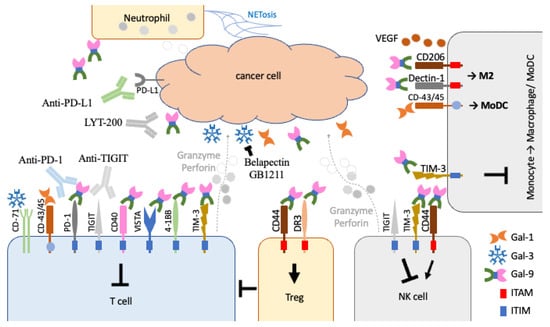
Figure 3.
Effects of galectin-1, -3, and -9 in the control of different types of immune cells within the TME. Galectin-1 (Gal-1) and -3 (Gal-3) have been shown to bind to CD45 and CD-71 to inhibit effector T cells (T cell). In contrast, galectin-9 (Gal-9) binds to TIM-3 and numerous ITIM-containing immunomodulators on effector T cells and NK cells for tumoural immunosuppression. It can also act on Tregs through CD44 and DR3 to inhibit effector T cells. It can also promote M2 macrophage polarisation and VEGF secretion in angiogenesis. The recruited neutrophils may also contribute to galectin-9 secretion within the environment.
Galectin-9 also acts on Treg cell differentiation and stability through CD44 and DR3 to inhibit pro-inflammatory T helper 1 (Th1) and Th17 cells [110,111]. In addition, galectin-9 also acts on Dectin-1 for M2 macrophage polarisation in protective responses and on protein VEGF for angiogenesis as pro-oncogenic roles in the cancer cells [74,75]. In a more complicated situation, galectin-9 acts on both modulators for activation CD44 and inhibition TIM-3 on NK cells in mice [112]. However, in human NK cells, Gal-9+ NK cells only promoted IFNγ expression and not the expression of cytolytic molecules when compared to their control counterpart. The controlling role of galectin-9 in NK cell functioning may be different between humans and mice.
Inhibiting galectin-9 may help to activate adaptive immunity during cancer treatment, regarding the ICIs PD-1/PD-L1 [73,76]. Due to the limited role of galectin-9 intracellularly, instead of small-molecule inhibitors, humanised monoclonal antibodies can also serve as inhibitory reagents. LYT-200 is one of the fully humanised IgG4 monoclonal antibodies against galectin-9. A clinical trial study (NCT04666688) is currently ongoing, assessing the safety, pharmacokinetics, and efficacy of LYT-200 as a monotherapy or combined with tislelizumab (PD-1 inhibitor) or gemcitabine/nab-paclitaxel chemotherapy in patients with metastatic head and neck, colorectal, pancreatic, or urothelial cancers (see Table 1). The results might provide further insight into the role of galectin-9 inhibition in cancer treatments. In addition, a recent study suggests that circulating galectin-9 and PD-L1 levels are independent of their tumoural expression levels. Further detailed analyses of galectin levels should be conducted [113].
In addition to lymphocytes, neutrophils are the dominant leukocytes in the innate immune response through phagocytosis and NETosis [114,115]. An in vitro study demonstrated that galectin-9 could activate neutrophils to enact a killing effect on cancer cells [116], and neutrophils also contribute to the secretion of galectin-9 [117]. However, in clinical trials, neutrophil tumour infiltration and the neutrophil-to-lymphocyte ratio (NLR) have been found to be correlated with poor prognostic markers in patients with different solid cancers [118,119,120].
Recently, mucosal-associated invariant T (MAIT) cells were suggested to play roles in immunotherapy in both blood and solid tumours [121]. Galectin-9 has also been shown to deplete MAIT cells in CLL [122]. It will be interesting to see future research on the regulation of MAIT cells and the damping of immunotherapy by galectin-9.
2.2.2. Other Tandem-Repeat Type Galectins: Galectin-4, -8, and -12
Galectin-4, -8, and -12 have been less well studied than galectin-1, -3, and -9. Although there is increasing interest in studying these molecules, we are still far from obtaining a complete picture. Galectin-4 was first identified as a ligand on a human blood group antigen [123]. Its expression has been detected in metastatic prostate cancer cells (Table 3) [124]. A high level of galectin-4 has also been shown to be associated with tissues in the advanced stages of breast and colorectal cancer [125,126]. Interestingly, galectin-8 has been found in malignant tumour tissues, especially in breast cancer [127]. Within the TME, secreted galectin-8 binds to podoplanin on tumour-associated macrophages and lymphatic endothelial cells to promote lymphangiogenesis and lymph node metastasis in breast cancer mouse models [67,68]. In contrast to most galectins, galectin-12 has been shown to promote the M1 macrophage polarisation [86,87] and may, therefore, play an opposite role(s) to other galectins in tumour suppression. The regulatory roles and inhibitory mechanisms of these three galectins in cancer development still need to be further investigated.
2.3. Prototype Galectins
2.3.1. Galectin-1 Is a Prototype Galectin with Diverse Roles in Intra- and Extracellular Processes
Galectin-1 is an important protein in various biological processes, such as cell adhesion, migration, and cell death. Numerous studies have shown that galectin-1 is overexpressed in different tumours, including breast, lung, ovarian, pancreas, and prostate cancers (listed in Table 3). Intracellularly, galectin-1 binds to oncogenic H-Ras protein and facilitates H-Ras-to-membrane anchorage [50] and subsequent interaction with Raf kinase to further activate downstream oncogenic pathways [128]. In addition, galectin-1 also interacts with gem nuclear organelle association protein 4 (Gemin4), as does galectin-3 [37]. Gemin4 is important for microRNA biosynthesis in gene silencing [129]. Interestingly, polymorphism on Gemin4 is associated with poor prognosis in early-stage NSCLC patients [130]. Although the role of microRNAs in cancer cell gene regulation has become widely acknowledged, the mechanisms and roles still need to be elucidated. Another binding partner of galectin-1 is glucose-regulated protein 78 (GRP78) (Table 2). The overexpression of galectin-1 is also correlated with lymph node and distant metastasis in gastric cancer and advanced staging [49]. High expressions of galectin-1 and GRP78 also reflect poor overall survival in gastric cancer patients. In NSCLC cells, GRP78 is responsive to oxidative stress and promotes autophagy and epithelial–mesenchymal transition (EMT) [131]. Additionally, another galectin-1-interacting molecule, protocadherin LKC (PCDH24) [38], is also involved in the β-catenin pathway in EMT [132].
In the extracellular environment, the secretion of galectin-1 occurs through a non-canonical pathway, as with other members of the galectin family. It has been suggested that it is secreted through stress-induced exocytosis or the exosome pathway [27]. Similarly to galectin-3, secreted galectin-1 also binds to numerous chemokines, including CCL1, -5, -13, -20, -21, -22, -24, -25, -26, and -28 and CXCL9, -11, -12, -13, -16, and -17 for modulating the migration properties of different immune cells [42]. In addition, secreted galectin-1 binds to cell surface CD43 and CD45 to induce effector T-cell apoptosis for immunosuppression in the TME [52,133]. On the other hand, binding to CD43 on neutrophils was shown to promote neutrophil recruitment to the tumour site [53]. Galectin-1 can also bind to CD43/45 on monocyte-derived dendritic cells (MoDCs) for activation and migration [51]. MoDCs can further provide tolerogenic effects and suppress effector T cells through the PD-1/PD-L1 pathways [134]. The recruitment of dendritic cells and neutrophils can promote the Th17 polarisation [135] and cancer-cell priming [136] to promote the metastasis [136,137].
Galectin-1 also acts on endothelial cells. It has also been found to bind to vascular endothelial growth factor 2 (VEGF2) to control endothelial cell survival via the Akt pathway and promote angiogenesis around the tumour site [55]. Galectin-1 has also been found to bind to Neurophilin-1 (NRP-1) [54], which serves as the VEGFR coreceptors in endothelial cells for tumour angiogenesis [138].
On the clinical side, galectin-1 has been used as a prognostic marker for the prediction of treatment outcomes, even though the mechanism by which systemic/serum galectin-1 level increases is unknown. For example, in melanoma patients receiving treatment with the monoclonal antibody bevacizumab against VEGFR, a higher level of serum galectin-1 was associated with better overall survival following treatment [139]. On the other hand, patients who showed the overexpression of galectin-1 in their tumour tissue had poor overall survival in the advanced-stage small-cell lung cancer [140]. In colorectal cancer patients, galectin-1 attenuated cytotoxic CD8+ T-cell-killing activity and showed high expression, especially in stroma cells, which led to poor overall survival in colorectal cancer patients [141].
When galectin-1 has been used as a treatment or in clinical trials, it has been less promising than the other candidates (galectin-3 and galectin-9) mentioned above. Most of the studies are still on the bench side. OTX008 is the most commonly used galectin-1 inhibitor [142]. OTX008 has been examined in thyroid cancer cell lines, oral squamous cell carcinoma, hepatocellular orthotopic implantation models, and SCLC patient-derived xenograft-implanted nude mice. The tumour cells/implanted tumours treated with galectin-1 inhibitor OTX008 were significantly suppressed [18,20,140,143]. The limited availability of galectin-1 inhibitors may harm possible future clinical trial studies; further increasing the available specific inhibitors is discussed below and by Marino et al. [14].
2.3.2. Another Prototype, Galectin-2, Has Controversial Roles in Different Cancers
Galectin-1 and galectin-2 share similar protein sequences and structures; however, their recognition and binding to carbohydrate side chains are different [144]. Extracellularly, secreted galectin-2 has been found to bind to CD14 on CD14intermediate/high monocytes to promote M1 macrophage differentiation for a pro-inflammatory effect [56]. Based on our observations, the overexpression of galectin-2 can play a tumour-suppressive role in the H-RasG12V-activated HCC cells [145], where H-Ras also plays an important role in HCC oncogenesis [146,147,148,149]. These findings may suggest that galectin-2 has a tumour-suppressive role. However, in triple-negative breast cancer, galectin-2 was found to act on immunosuppression in vivo to promote tumour growth but not to act on cancer cells [150]. These results suggest that galectin-2 may play dynamic regulatory roles in cancer and immune cell populations in terms of controlling tumour behaviour against different genetic backgrounds, such as H-Ras status. Increased serum galectin-2 was detected in circulating blood samples from breast and colon cancer patients and was correlated with different cytokines: IL-6; GCS-F; GROα/CXCL1; and MCP-1 [126]. It may serve as a prognostic marker. However, the systemic role of galectin-2 is still elusive.
2.3.3. Other Prototype Galectins (-7, -10, -13, -14, and -16) Are Located on Chromosome 19
With the exception of galectin-4, which is classified as a tandem-repeat-type protein structure, as stated above, all the other galectin genes are clustered on chromosome 19 in the q13.2 position, including LGALS4, LGALS7, LGALS10, LGALS13, LGALS14, and LGALS16, and their protein products are classified as prototype galectins (Figure 1 and Figure 2). LGALS13, -16, and -14 are in a sense direction, whereas LGALS4, -7, and -10 are in an anti-sense direction [151]. LGALS7 and LGALS4 are upstream of the LGALS13, -16, and -14 clusters, whereas LGALS10 is downstream of these clusters.
Galectin-13 and -14, which are also known as placental galectins, can induce apoptosis in T lymphocytes. CD95 (Fas) and IL-8 could be induced in non-CD3-/CD28-activated T lymphocytes, particularly in the case of miscarriage [89]. The high expression of these galectins and placental galectin-16 is associated with poor prognosis and poor overall survival in breast and ovarian cancers [102]. These galectins are mainly expressed in the placenta and brain, with limited expression in the breasts, bone marrow, stomach, pancreas, retina, and skin. High percentages of thyroid and lung cancer tissues (>60%) express galectin-13 and galectin-14, respectively [102]. Greater attention should be paid to their immunological roles in cancer treatment. However, these placental galectins seem to act in the opposite direction in thyroid cancer. Intracellularly, galectin-13 can interact with the Hox family protein HOXA1 [88]. The upregulation of HOXA1 is also associated with poor prognosis in breast cancer and HCC patients [152,153]. Its intracellular role(s) should not be neglected. Galectin-14 can promote cell migration and invasion by upregulating MMP9 and N-cadherin in the trophoblasts [154]. Both galectins are also associated with pro-oncogenic roles. Furthermore, galectin-16 is the least-studied member of the family. It has been found to be expressed in different cancer cell lines, and its expression is upregulated by the cAMP activation [155]. Its role in cancer development is still far from understood.
3. From Bench to Bedside
3.1. Availability of Galectin-Specific Inhibitors
Luckily, due to the small size of galectin proteins, their protein structures and binding site properties have been revealed by numerous nuclear magnetic resonance (NMR) studies [156]. An understanding of protein structures and functions can expedite the drug development process. Galectin inhibitors can be developed in different forms, including (1) small-molecule carbohydrates, (2) natural polysaccharides and their derivatives, (3) peptides and peptidomimetics, and (4) humanised monoclonal antibodies [157]. Their structures and binding affinities to galectin-1, galectin-3, and galectin-9 were fully reviewed by Mariño et al. (2023) [14]. Interestingly, further NMR spectroscopic analyses and competitive ELISA binding assays for eluting protein structure and substrate specifics using synthetic hydroxypropyl methacrylamide (HPMA) copolymers with multi-Galβ4GlcNAc (LacNAc), which is specific to the CRD of human galectin-1 and galectin-3 with differential affinity at the sub-nanomolar level, have been explored [158]. This kind of assays might allow the identification of potential small-molecular inhibitors for future applications. In addition, peptide inhibitors and monoclonal antibodies are also useful to target secreted galectins and not interfere with the intracellular roles of galectins to minimise the side effects. New development techniques for synthesising glycosyl-side chain molecules and amino acid polypeptides may provide further new targets for galectin-based treatment in future.
3.2. Applications, Safety/Pitfalls/Limitations, and Ongoing Clinical Trials
At present, all ICIs come in the form of antibodies. This creates a limitation for future treatment potential, especially for brain metastasis, where the antibodies are unable to pass through the blood–brain barrier. In addition, the limited availability of immune cells within the central nervous system also imposes a limitation on the use of ICIs in patients with metastases at sites such as the brain and spine. The development of different small-molecule inhibitors for PD-1/PD-L1 is ongoing [159]. Further understanding of the resident and infiltrating immune cells across to the brain/spine via the blood–brain barrier [160] might make the impossible possible.
T-cell exhaustion is a common phenomenon in treatment using ICIs. Galectin inhibitors are a new approach to improve this situation and the durability of the treatment regimen. Combinations of ICIs and other inhibitors in the form of inhibitory antibodies or TKIs have been the subject of ongoing studies [161]. All the components, including cytotoxic T cells, NK cells, and their inhibitory molecules, within the TME are required to orchestrate the anti-tumorigenic mode of killing tumour cells.
The potential pitfalls of galectin inhibitors in future treatments should be noted in drug development. The first limitation is the unknown factors associated with laboratory settings, such as the differences between mouse models and real humans. The blood components of laboratory mice are different to those of humans. Mice have a high percentage of lymphocytes (around 70% of all leukocytes). In human bodies, the predominant immune cells are neutrophils, which account for over 50% of all leukocytes [162]. This issue is not easily addressed even in humanised mouse models [163]. It is difficult to understand a drug or biological reagent’s effect without human trial studies. In addition, as discussed above, most galectins can alter immune responses. For example, galectin-3 has been shown to play important roles against pathogens [41]. For subjects treated with galectin-3 inhibitors, infection-related adverse effects in subsequent phase 2 or 3 trial studies should be noted as a safety parameter. Treatment using galectin-9 inhibitors also raises similar concerns to galectin-3-inhibitor treatment and can affect different types of immune cells. It may also raise additional concerns regarding autoimmune issues.
Although immune-related adverse events (irAEs) are inevitable in I-O treatment, the occurrence of irAEs during treatment reflects better prognosis and overall outcomes [164]. If the irAEs can be better managed and controlled using corticosteroids, this is a good sign of successful treatment. The effective and safe dosages for combined treatments could be evaluated and determined in clinical trial settings and provide guidelines for regimens and any dosage reductions in the case of irAEs. While clinical trials are still ongoing with many unforeseen hurdles to overcome, galectin-3 small-molecule inhibitors, such as belapectin and GB1211, and galectin-9 humanised monoclonal antibodies still provide new opportunities for combination therapies to tackle the current unmet and unresolved issues associated with ICI treatments for many difficult-to-treat cancers.
4. Conclusions
Galectins not only have immunomodulatory roles, but they also contribute to intracellular functions and are expressed in different cancers. Their oncogenic roles can be tackled using small-molecule inhibitors, which can induce cancer cell death in contexts where ordinary ICIs cannot gain access. This is especially important in brain metastasis patients, where only limited immune cells are available for treatment [165]. The ability of antibody delivery and the accessibility of circulating immune cells to tumour sites inside the brain are still largely questionable. Inhibitors, such as those of galectin-3, may provide an alternative option for this kind of patient if the ongoing clinical trials are successful. Combined treatment using galectin inhibitors with TKIs and/or chemotherapy treatment may offer another potential treatment regimen direction. Of course, the related toxicity should be better analysed, and dosage management will be required for different levels of adverse events. Finally, galectins and their inhibitors, as new therapeutic targets, provide more flexibility for future treatment regimens and the addressing of unmet treatment needs.
Author Contributions
Conceptualisation, F.C.F.K., S.Y. and J.C.M.H.; resources, F.C.F.K., S.Y., K.W.L. and S.K.L.; writing—original draft preparation, F.C.F.K.; writing—review and editing, S.Y. and J.C.M.H.; funding acquisition, F.C.F.K. and J.C.M.H. All authors have read and agreed to the published version of the manuscript.
Funding
Publication made possible in part by support from the HKU Libraries Open Access Author Fund (Ref: OAAF2022-23-085) sponsored by the HKU Libraries, The University of Hong Kong.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| CRD | Carbohydrate-recognition domain |
| CTLA-4 | Cytotoxic T lymphocyte-associated antigen-4 |
| EMT | Epithelial–Mesenchymal Transition |
| ERK | Extracellular Signal-regulated kinases |
| HCC | Hepatocellular carcinoma |
| ICIs | Immune checkpoint inhibitors |
| I-O | Immuno-oncology |
| irAEs | Immune-related adverse events |
| ITAM | Immunoreceptor tyrosine-based activation motif |
| ITIM | Immunoreceptor tyrosine-based inhibitory motif |
| MEK | Mitogen-activated protein kinase/ ERK kinases |
| NK cells | Natural Killer Cells |
| NSCLC | Non-small cell lung cancer |
| PD-1/-L1 | Programmed cell death-1/-Ligand 1 |
| PI3K | Phosphoinositide 3-kinases |
| PLC/PKC | Phospholipase C/Protein Kinase C |
| Rac | Ras-related C3 botulinum toxin substrates |
| Raf | Rapidly accelerated fibrosarcoma kinases |
| RALGDS | Ras-like (Ral) guanine nucleotide dissociation stimulator |
| SCLC | Small cell lung cancer |
| TIAM1 | T-lymphoma Invasion and Metastasis 1 |
| TIGIT | T-cell immunoreceptor with immunoglobulin and ITIM domains |
| TILs | Tumour-infiltrating lymphocytes |
| TKIs | Tyrosine kinase inhibitors |
| TME | Tumour microenvironment |
| Tregs | Regulatory T cells |
| VEGF/Rs | Vascular Endothelial Growth Factor/Receptors |
References
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Coley, W.B. II. Contribution to the Knowledge of Sarcoma. Ann. Surg. 1891, 14, 199–220. [Google Scholar] [CrossRef]
- Hodi, F.S.; O’Day, S.J.; McDermott, D.F.; Weber, R.W.; Sosman, J.A.; Haanen, J.B.; Gonzalez, R.; Robert, C.; Schadendorf, D.; Hassel, J.C.; et al. Improved survival with ipilimumab in patients with metastatic melanoma. N. Engl. J. Med. 2010, 363, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef]
- Hargadon, K.M.; Johnson, C.E.; Williams, C.J. Immune checkpoint blockade therapy for cancer: An overview of FDA-approved immune checkpoint inhibitors. Int. Immunopharmacol. 2018, 62, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Khan, F.; Qari, H.A.; Upadhyay, T.K.; Alkhateeb, A.F.; Oves, M. Revolutionization in Cancer Therapeutics via Targeting Major Immune Checkpoints PD-1, PD-L1 and CTLA-4. Pharmaceuticals 2022, 15, 335. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.; Pelosof, L.; Lemery, S.; Gong, Y.; Goldberg, K.B.; Farrell, A.T.; Keegan, P.; Veeraraghavan, J.; Wei, G.; Blumenthal, G.M.; et al. Systematic Review of PD-1/PD-L1 Inhibitors in Oncology: From Personalized Medicine to Public Health. Oncologist 2021, 26, e1786–e1799. [Google Scholar] [CrossRef] [PubMed]
- Chow, A.; Perica, K.; Klebanoff, C.A.; Wolchok, J.D. Clinical implications of T cell exhaustion for cancer immunotherapy. Nat. Rev. Clin. Oncol. 2022, 19, 775–790. [Google Scholar] [CrossRef]
- Togashi, Y.; Shitara, K.; Nishikawa, H. Regulatory T cells in cancer immunosuppression—Implications for anticancer therapy. Nat. Rev. Clin. Oncol. 2019, 16, 356–371. [Google Scholar] [CrossRef]
- Preillon, J.; Cuende, J.; Rabolli, V.; Garnero, L.; Mercier, M.; Wald, N.; Pappalardo, A.; Denies, S.; Jamart, D.; Michaux, A.C.; et al. Restoration of T-cell Effector Function, Depletion of Tregs, and Direct Killing of Tumor Cells: The Multiple Mechanisms of Action of a-TIGIT Antagonist Antibodies. Mol. Cancer Ther. 2021, 20, 121–131. [Google Scholar] [CrossRef]
- Florou, V.; Garrido-Laguna, I. Clinical Development of Anti-TIGIT Antibodies for Immunotherapy of Cancer. Curr. Oncol. Rep. 2022, 24, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.C.; Abreu, D.R.; Hussein, M.; Cobo, M.; Patel, A.J.; Secen, N.; Lee, K.H.; Massuti, B.; Hiret, S.; Yang, J.C.H.; et al. Tiragolumab plus atezolizumab versus placebo plus atezolizumab as a first-line treatment for PD-L1-selected non-small-cell lung cancer (CITYSCAPE): Primary and follow-up analyses of a randomised, double-blind, phase 2 study. Lancet Oncol. 2022, 23, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Brazel, D.; Ou, S.I.; Nagasaka, M. Tiragolumab (Anti-TIGIT) in SCLC: Skyscraper-02, a Towering Inferno. Lung Cancer Targets Ther. 2023, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Marino, K.V.; Cagnoni, A.J.; Croci, D.O.; Rabinovich, G.A. Targeting galectin-driven regulatory circuits in cancer and fibrosis. Nat. Rev. Drug Discov. 2023, 22, 295–316. [Google Scholar] [CrossRef] [PubMed]
- Zetterberg, F.R.; MacKinnon, A.; Brimert, T.; Gravelle, L.; Johnsson, R.E.; Kahl-Knutson, B.; Leffler, H.; Nilsson, U.J.; Pedersen, A.; Peterson, K.; et al. Discovery and Optimization of the First Highly Effective and Orally Available Galectin-3 Inhibitors for Treatment of Fibrotic Disease. J. Med. Chem. 2022, 65, 12626–12638. [Google Scholar] [CrossRef]
- Aslanis, V.; Slack, R.J.; MacKinnon, A.C.; McClinton, C.; Tantawi, S.; Gravelle, L.; Nilsson, U.J.; Leffler, H.; Brooks, A.; Khindri, S.K.; et al. Safety and pharmacokinetics of GB1211, an oral galectin-3 inhibitor: A single- and multiple-dose first-in-human study in healthy participants. Cancer Chemother. Pharmacol. 2023, 91, 267–280. [Google Scholar] [CrossRef]
- Astorgues-Xerri, L.; Riveiro, M.E.; Tijeras-Raballand, A.; Serova, M.; Rabinovich, G.A.; Bieche, I.; Vidaud, M.; de Gramont, A.; Martinet, M.; Cvitkovic, E.; et al. OTX008, a selective small-molecule inhibitor of galectin-1, downregulates cancer cell proliferation, invasion and tumour angiogenesis. Eur. J. Cancer 2014, 50, 2463–2477. [Google Scholar] [CrossRef]
- Greer, P.F.C.; Rich, A.; Coates, D.E. Effects of galectin-1 inhibitor OTX008 on oral squamous cell carcinoma cells in vitro and the role of AP-1 and the MAPK/ERK pathway. Arch. Oral. Biol. 2022, 134, 105335. [Google Scholar] [CrossRef]
- Koonce, N.A.; Griffin, R.J.; Dings, R.P.M. Galectin-1 Inhibitor OTX008 Induces Tumor Vessel Normalization and Tumor Growth Inhibition in Human Head and Neck Squamous Cell Carcinoma Models. Int. J. Mol. Sci. 2017, 18, 2671. [Google Scholar] [CrossRef]
- Leung, Z.; Ko, F.C.F.; Tey, S.K.; Kwong, E.M.L.; Mao, X.; Liu, B.H.M.; Ma, A.P.Y.; Fung, Y.M.E.; Che, C.M.; Wong, D.K.H.; et al. Galectin-1 promotes hepatocellular carcinoma and the combined therapeutic effect of OTX008 galectin-1 inhibitor and sorafenib in tumor cells. J. Exp. Clin. Cancer Res. 2019, 38, 423. [Google Scholar] [CrossRef]
- Curti, B.D.; Koguchi, Y.; Leidner, R.S.; Rolig, A.S.; Sturgill, E.R.; Sun, Z.; Wu, Y.; Rajamanickam, V.; Bernard, B.; Hilgart-Martiszus, I.; et al. Enhancing clinical and immunological effects of anti-PD-1 with belapectin, a galectin-3 inhibitor. J. Immunother. Cancer 2021, 9, e002371. [Google Scholar] [CrossRef] [PubMed]
- Lau, E.S.; Liu, E.; Paniagua, S.M.; Sarma, A.A.; Zampierollo, G.; Lopez, B.; Diez, J.; Wang, T.J.; Ho, J.E. Galectin-3 Inhibition With Modified Citrus Pectin in Hypertension. JACC Basic Transl. Sci. 2021, 6, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Sigamani, A.; Mayo, K.H.; Miller, M.C.; Chen-Walden, H.; Reddy, S.; Platt, D. An Oral Galectin Inhibitor in COVID-19-A Phase II Randomized Controlled Trial. Vaccines 2023, 11, 731. [Google Scholar] [CrossRef]
- von Heijne, G. Membrane-protein topology. Nat. Rev. Mol. Cell Biol. 2006, 7, 909–918. [Google Scholar] [CrossRef]
- Nielsen, H.; Tsirigos, K.D.; Brunak, S.; von Heijne, G. A Brief History of Protein Sorting Prediction. Protein J. 2019, 38, 200–216. [Google Scholar] [CrossRef]
- Zanetti, G.; Pahuja, K.B.; Studer, S.; Shim, S.; Schekman, R. COPII and the regulation of protein sorting in mammals. Nat. Cell Biol. 2011, 14, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Popa, S.J.; Stewart, S.E.; Moreau, K. Unconventional secretion of annexins and galectins. Semin. Cell Dev. Biol. 2018, 83, 42–50. [Google Scholar] [CrossRef]
- Kutzner, T.J.; Higuero, A.M.; Sussmair, M.; Kopitz, J.; Hingar, M.; Diez-Revuelta, N.; Caballero, G.G.; Kaltner, H.; Lindner, I.; Abad-Rodriguez, J.; et al. How presence of a signal peptide affects human galectins-1 and -4: Clues to explain common absence of a leader sequence among adhesion/growth-regulatory galectins. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129449. [Google Scholar] [CrossRef]
- Wan, L.; Hsu, Y.A.; Wei, C.C.; Liu, F.T. Galectins in allergic inflammatory diseases. Mol. Aspects Med. 2021, 79, 100925. [Google Scholar] [CrossRef]
- Clementy, N.; Piver, E.; Bisson, A.; Andre, C.; Bernard, A.; Pierre, B.; Fauchier, L.; Babuty, D. Galectin-3 in Atrial Fibrillation: Mechanisms and Therapeutic Implications. Int. J. Mol. Sci. 2018, 19, 976. [Google Scholar] [CrossRef]
- Mazurek, N.; Sun, Y.J.; Price, J.E.; Ramdas, L.; Schober, W.; Nangia-Makker, P.; Byrd, J.C.; Raz, A.; Bresalier, R.S. Phosphorylation of galectin-3 contributes to malignant transformation of human epithelial cells via modulation of unique sets of genes. Cancer Res. 2005, 65, 10767–10775. [Google Scholar] [CrossRef] [PubMed]
- Mysore, V.P.; Zhou, Z.W.; Ambrogio, C.; Li, L.; Kapp, J.N.; Lu, C.; Wang, Q.; Tucker, M.R.; Okoro, J.J.; Nagy-Davidescu, G.; et al. A structural model of a Ras-Raf signalosome. Nat. Struct. Mol. Biol. 2021, 28, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Guo, Z.; Wang, F.; Fu, L. KRAS mutation: From undruggable to druggable in cancer. Signal Transduct. Target Ther. 2021, 6, 386. [Google Scholar] [CrossRef]
- Liu, W.; Hsu, D.K.; Chen, H.Y.; Yang, R.Y.; Carraway, K.L., 3rd; Isseroff, R.R.; Liu, F.T. Galectin-3 regulates intracellular trafficking of EGFR through Alix and promotes keratinocyte migration. J. Investig. Dermatol. 2012, 132, 2828–2837. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.F.; Tsao, C.H.; Lin, Y.T.; Hsu, D.K.; Chiang, M.L.; Lo, C.H.; Chien, F.C.; Chen, P.; Arthur Chen, Y.M.; Chen, H.Y.; et al. Galectin-3 promotes HIV-1 budding via association with Alix and Gag p6. Glycobiology 2014, 24, 1022–1035. [Google Scholar] [CrossRef]
- Monypenny, J.; Milewicz, H.; Flores-Borja, F.; Weitsman, G.; Cheung, A.; Chowdhury, R.; Burgoyne, T.; Arulappu, A.; Lawler, K.; Barber, P.R.; et al. ALIX Regulates Tumor-Mediated Immunosuppression by Controlling EGFR Activity and PD-L1 Presentation. Cell Rep. 2018, 24, 630–641. [Google Scholar] [CrossRef]
- Park, J.W.; Voss, P.G.; Grabski, S.; Wang, J.L.; Patterson, R.J. Association of galectin-1 and galectin-3 with Gemin4 in complexes containing the SMN protein. Nucleic Acids Res. 2001, 29, 3595–3602. [Google Scholar] [CrossRef]
- Ose, R.; Oharaa, O.; Nagase, T. Galectin-1 and Galectin-3 Mediate Protocadherin-24-Dependent Membrane Localization of beta-catenin in Colon Cancer Cell Line HCT116. Curr. Chem. Genom. 2012, 6, 18–26. [Google Scholar] [CrossRef]
- Jia, W.; Kong, L.; Kidoya, H.; Naito, H.; Muramatsu, F.; Hayashi, Y.; Hsieh, H.Y.; Yamakawa, D.; Hsu, D.K.; Liu, F.T.; et al. Indispensable role of Galectin-3 in promoting quiescence of hematopoietic stem cells. Nat. Commun. 2021, 12, 2118. [Google Scholar] [CrossRef]
- Seguin, L.; Camargo, M.F.; Wettersten, H.I.; Kato, S.; Desgrosellier, J.S.; von Schalscha, T.; Elliott, K.C.; Cosset, E.; Lesperance, J.; Weis, S.M.; et al. Galectin-3, a Druggable Vulnerability for KRAS-Addicted Cancers. Cancer Discov. 2017, 7, 1464–1479. [Google Scholar] [CrossRef]
- Diaz-Alvarez, L.; Ortega, E. The Many Roles of Galectin-3, a Multifaceted Molecule, in Innate Immune Responses against Pathogens. Mediators Inflamm. 2017, 2017, 9247574. [Google Scholar] [CrossRef]
- Eckardt, V.; Miller, M.C.; Blanchet, X.; Duan, R.; Leberzammer, J.; Duchene, J.; Soehnlein, O.; Megens, R.T.; Ludwig, A.K.; Dregni, A.; et al. Chemokines and galectins form heterodimers to modulate inflammation. EMBO Rep. 2020, 21, e47852. [Google Scholar] [CrossRef] [PubMed]
- Stillman, B.N.; Hsu, D.K.; Pang, M.; Brewer, C.F.; Johnson, P.; Liu, F.T.; Baum, L.G. Galectin-3 and galectin-1 bind distinct cell surface glycoprotein receptors to induce T cell death. J. Immunol. 2006, 176, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Huang, Q.; Xiao, W.; Zhao, Y.; Pi, J.; Xu, H.; Zhao, H.; Xu, J.; Evans, C.E.; Jin, H. Advances in Anti-Tumor Treatments Targeting the CD47/SIRPalpha Axis. Front. Immunol. 2020, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Al Attar, A.; Antaramian, A.; Noureddin, M. Review of galectin-3 inhibitors in the treatment of nonalcoholic steatohepatitis. Expert Rev. Clin. Pharmacol. 2021, 14, 457–464. [Google Scholar] [CrossRef]
- Huh, Y.; Cho, Y.J.; Nam, G.E. Recent Epidemiology and Risk Factors of Nonalcoholic Fatty Liver Disease. J. Obes. Metab. Syndr. 2022, 31, 17–27. [Google Scholar] [CrossRef]
- Slack, R.J.; Mills, R.; Mackinnon, A.C. The therapeutic potential of galectin-3 inhibition in fibrotic disease. Int. J. Biochem. Cell Biol. 2021, 130, 105881. [Google Scholar] [CrossRef]
- Ensembl—Galectins. Available online: https://www.ensembl.org/Homo_sapiens/Search/Results?q=galectins;site=ensembl;facet_species=Human (accessed on 2 April 2023).
- Zhang, Q.; Ali, M.; Wang, Y.; Sun, Q.N.; Zhu, X.D.; Tang, D.; Wang, W.; Zhang, C.Y.; Zhou, H.H.; Wang, D.R. Galectin-1 binds GRP78 to promote the proliferation and metastasis of gastric cancer. Int. J. Oncol. 2022, 61, 1–18. [Google Scholar] [CrossRef]
- Paz, A.; Haklai, R.; Elad-Sfadia, G.; Ballan, E.; Kloog, Y. Galectin-1 binds oncogenic H-Ras to mediate Ras membrane anchorage and cell transformation. Oncogene 2001, 20, 7486–7493. [Google Scholar] [CrossRef]
- Fulcher, J.A.; Chang, M.H.; Wang, S.; Almazan, T.; Hashimi, S.T.; Eriksson, A.U.; Wen, X.; Pang, M.; Baum, L.G.; Singh, R.R.; et al. Galectin-1 co-clusters CD43/CD45 on dendritic cells and induces cell activation and migration through Syk and protein kinase C signaling. J. Biol. Chem. 2009, 284, 26860–26870. [Google Scholar] [CrossRef]
- Hernandez, J.D.; Nguyen, J.T.; He, J.; Wang, W.; Ardman, B.; Green, J.M.; Fukuda, M.; Baum, L.G. Galectin-1 binds different CD43 glycoforms to cluster CD43 and regulate T cell death. J. Immunol. 2006, 177, 5328–5336. [Google Scholar] [CrossRef] [PubMed]
- Auvynet, C.; Moreno, S.; Melchy, E.; Coronado-Martinez, I.; Montiel, J.L.; Aguilar-Delfin, I.; Rosenstein, Y. Galectin-1 promotes human neutrophil migration. Glycobiology 2013, 23, 32–42. [Google Scholar] [CrossRef]
- Hsieh, S.H.; Ying, N.W.; Wu, M.H.; Chiang, W.F.; Hsu, C.L.; Wong, T.Y.; Jin, Y.T.; Hong, T.M.; Chen, Y.L. Galectin-1, a novel ligand of neuropilin-1, activates VEGFR-2 signaling and modulates the migration of vascular endothelial cells. Oncogene 2008, 27, 3746–3753. [Google Scholar] [CrossRef] [PubMed]
- Croci, D.O.; Cerliani, J.P.; Dalotto-Moreno, T.; Mendez-Huergo, S.P.; Mascanfroni, I.D.; Dergan-Dylon, S.; Toscano, M.A.; Caramelo, J.J.; Garcia-Vallejo, J.J.; Ouyang, J.; et al. Glycosylation-dependent lectin-receptor interactions preserve angiogenesis in anti-VEGF refractory tumors. Cell 2014, 156, 744–758. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, C.; Vogel, D.Y.; Hollander, M.R.; Baggen, J.M.; Fontijn, R.D.; Nieuwenhuis, S.; Haverkamp, A.; de Vries, M.R.; Quax, P.H.; Garcia-Vallejo, J.J.; et al. Galectin-2 induces a proinflammatory, anti-arteriogenic phenotype in monocytes and macrophages. PLoS ONE 2015, 10, e0124347. [Google Scholar] [CrossRef]
- Chen, H.Y.; Fermin, A.; Vardhana, S.; Weng, I.C.; Lo, K.F.; Chang, E.Y.; Maverakis, E.; Yang, R.Y.; Hsu, D.K.; Dustin, M.L.; et al. Galectin-3 negatively regulates TCR-mediated CD4+ T-cell activation at the immunological synapse. Proc. Natl. Acad. Sci. USA 2009, 106, 14496–14501. [Google Scholar] [CrossRef]
- Fritsch, K.; Mernberger, M.; Nist, A.; Stiewe, T.; Brehm, A.; Jacob, R. Galectin-3 interacts with components of the nuclear ribonucleoprotein complex. BMC Cancer 2016, 16, 502. [Google Scholar] [CrossRef]
- Kuo, H.Y.; Hsu, H.T.; Chen, Y.C.; Chang, Y.W.; Liu, F.T.; Wu, C.W. Galectin-3 modulates the EGFR signalling-mediated regulation of Sox2 expression via c-Myc in lung cancer. Glycobiology 2016, 26, 155–165. [Google Scholar] [CrossRef]
- Gordon-Alonso, M.; Hirsch, T.; Wildmann, C.; van der Bruggen, P. Galectin-3 captures interferon-gamma in the tumor matrix reducing chemokine gradient production and T-cell tumor infiltration. Nat. Commun. 2017, 8, 793. [Google Scholar] [CrossRef]
- Kouo, T.; Huang, L.; Pucsek, A.B.; Cao, M.; Solt, S.; Armstrong, T.; Jaffee, E. Galectin-3 Shapes Antitumor Immune Responses by Suppressing CD8+ T Cells via LAG-3 and Inhibiting Expansion of Plasmacytoid Dendritic Cells. Cancer Immunol. Res. 2015, 3, 412–423. [Google Scholar] [CrossRef]
- Piyush, T.; Chacko, A.R.; Sindrewicz, P.; Hilkens, J.; Rhodes, J.M.; Yu, L.G. Interaction of galectin-3 with MUC1 on cell surface promotes EGFR dimerization and activation in human epithelial cancer cells. Cell Death Differ. 2017, 24, 1937–1947. [Google Scholar] [CrossRef] [PubMed]
- Paclik, D.; Danese, S.; Berndt, U.; Wiedenmann, B.; Dignass, A.; Sturm, A. Galectin-4 controls intestinal inflammation by selective regulation of peripheral and mucosal T cell apoptosis and cell cycle. PLoS ONE 2008, 3, e2629. [Google Scholar] [CrossRef] [PubMed]
- Villeneuve, C.; Baricault, L.; Canelle, L.; Barboule, N.; Racca, C.; Monsarrat, B.; Magnaldo, T.; Larminat, F. Mitochondrial proteomic approach reveals galectin-7 as a novel BCL-2 binding protein in human cells. Mol. Biol. Cell 2011, 22, 999–1013. [Google Scholar] [CrossRef]
- Nishi, N.; Shoji, H.; Seki, M.; Itoh, A.; Miyanaka, H.; Yuube, K.; Hirashima, M.; Nakamura, T. Galectin-8 modulates neutrophil function via interaction with integrin alphaM. Glycobiology 2003, 13, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Renard, H.F.; Tyckaert, F.; Lo Giudice, C.; Hirsch, T.; Valades-Cruz, C.A.; Lemaigre, C.; Shafaq-Zadah, M.; Wunder, C.; Wattiez, R.; Johannes, L.; et al. Endophilin-A3 and Galectin-8 control the clathrin-independent endocytosis of CD166. Nat. Commun. 2020, 11, 1457. [Google Scholar] [CrossRef]
- Bieniasz-Krzywiec, P.; Martin-Perez, R.; Ehling, M.; Garcia-Caballero, M.; Pinioti, S.; Pretto, S.; Kroes, R.; Aldeni, C.; Di Matteo, M.; Prenen, H.; et al. Podoplanin-Expressing Macrophages Promote Lymphangiogenesis and Lymphoinvasion in Breast Cancer. Cell. Metab. 2019, 30, 917–936.e910. [Google Scholar] [CrossRef]
- Cueni, L.N.; Detmar, M. Galectin-8 interacts with podoplanin and modulates lymphatic endothelial cell functions. Exp. Cell Res. 2009, 315, 1715–1723. [Google Scholar] [CrossRef]
- Goncalves Silva, I.; Ruegg, L.; Gibbs, B.F.; Bardelli, M.; Fruehwirth, A.; Varani, L.; Berger, S.M.; Fasler-Kan, E.; Sumbayev, V.V. The immune receptor Tim-3 acts as a trafficker in a Tim-3/galectin-9 autocrine loop in human myeloid leukemia cells. Oncoimmunology 2016, 5, e1195535. [Google Scholar] [CrossRef]
- Sudhakar, J.N.; Lu, H.H.; Chiang, H.Y.; Suen, C.S.; Hwang, M.J.; Wu, S.Y.; Shen, C.N.; Chang, Y.M.; Li, F.A.; Liu, F.T.; et al. Lumenal Galectin-9-Lamp2 interaction regulates lysosome and autophagy to prevent pathogenesis in the intestine and pancreas. Nat. Commun. 2020, 11, 4286. [Google Scholar] [CrossRef]
- Madireddi, S.; Eun, S.Y.; Lee, S.W.; Nemcovicova, I.; Mehta, A.K.; Zajonc, D.M.; Nishi, N.; Niki, T.; Hirashima, M.; Croft, M. Galectin-9 controls the therapeutic activity of 4-1BB-targeting antibodies. J. Exp. Med. 2014, 211, 1433–1448. [Google Scholar] [CrossRef]
- Vaitaitis, G.M.; Wagner, D.H., Jr. Galectin-9 controls CD40 signaling through a Tim-3 independent mechanism and redirects the cytokine profile of pathogenic T cells in autoimmunity. PLoS ONE 2012, 7, e38708. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Thalhamer, T.; Franca, R.F.; Xiao, S.; Wang, C.; Hotta, C.; Zhu, C.; Hirashima, M.; Anderson, A.C.; Kuchroo, V.K. Galectin-9-CD44 interaction enhances stability and function of adaptive regulatory T cells. Immunity 2014, 41, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Enninga, E.A.L.; Chatzopoulos, K.; Butterfield, J.T.; Sutor, S.L.; Leontovich, A.A.; Nevala, W.K.; Flotte, T.J.; Markovic, S.N. CD206-positive myeloid cells bind galectin-9 and promote a tumor-supportive microenvironment. J. Pathol. 2018, 245, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Daley, D.; Mani, V.R.; Mohan, N.; Akkad, N.; Ochi, A.; Heindel, D.W.; Lee, K.B.; Zambirinis, C.P.; Pandian, G.S.B.; Savadkar, S.; et al. Dectin 1 activation on macrophages by galectin 9 promotes pancreatic carcinoma and peritumoral immune tolerance. Nat. Med. 2017, 23, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Madireddi, S.; Eun, S.Y.; Mehta, A.K.; Birta, A.; Zajonc, D.M.; Niki, T.; Hirashima, M.; Podack, E.R.; Schreiber, T.H.; Croft, M. Regulatory T Cell-Mediated Suppression of Inflammation Induced by DR3 Signaling Is Dependent on Galectin-9. J. Immunol. 2017, 199, 2721–2728. [Google Scholar] [CrossRef]
- Yang, R.; Sun, L.; Li, C.F.; Wang, Y.H.; Yao, J.; Li, H.; Yan, M.; Chang, W.C.; Hsu, J.M.; Cha, J.H.; et al. Galectin-9 interacts with PD-1 and TIM-3 to regulate T cell death and is a target for cancer immunotherapy. Nat. Commun. 2021, 12, 832. [Google Scholar] [CrossRef]
- Schaefer, K.; Webb, N.E.; Pang, M.; Hernandez-Davies, J.E.; Lee, K.P.; Gonzalez, P.; Douglass, M.V.; Lee, B.; Baum, L.G. Galectin-9 binds to O-glycans on protein disulfide isomerase. Glycobiology 2017, 27, 878–887. [Google Scholar] [CrossRef]
- Bi, S.; Hong, P.W.; Lee, B.; Baum, L.G. Galectin-9 binding to cell surface protein disulfide isomerase regulates the redox environment to enhance T-cell migration and HIV entry. Proc. Natl. Acad. Sci. USA 2011, 108, 10650–10655. [Google Scholar] [CrossRef]
- Yang, R.; Sun, L.; Li, C.F.; Wang, Y.H.; Xia, W.; Liu, B.; Chu, Y.Y.; Bover, L.; Vien, L.; Hung, M.C. Development and characterization of anti-galectin-9 antibodies that protect T cells from galectin-9-induced cell death. J. Biol. Chem. 2022, 298, 101821. [Google Scholar] [CrossRef]
- Colomb, F.; Giron, L.B.; Premeaux, T.A.; Mitchell, B.I.; Niki, T.; Papasavvas, E.; Montaner, L.J.; Ndhlovu, L.C.; Abdel-Mohsen, M. Galectin-9 Mediates HIV Transcription by Inducing TCR-Dependent ERK Signaling. Front. Immunol. 2019, 10, 267. [Google Scholar] [CrossRef]
- Pang, N.; Alimu, X.; Chen, R.; Muhashi, M.; Ma, J.; Chen, G.; Zhao, F.; Wang, L.; Qu, J.; Ding, J. Activated Galectin-9/Tim3 promotes Treg and suppresses Th1 effector function in chronic lymphocytic leukemia. FASEB J. 2021, 35, e21556. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Anderson, A.C.; Schubart, A.; Xiong, H.; Imitola, J.; Khoury, S.J.; Zheng, X.X.; Strom, T.B.; Kuchroo, V.K. The Tim-3 ligand galectin-9 negatively regulates T helper type 1 immunity. Nat. Immunol. 2005, 6, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Yasinska, I.M.; Meyer, N.H.; Schlichtner, S.; Hussain, R.; Siligardi, G.; Casely-Hayford, M.; Fiedler, W.; Wellbrock, J.; Desmet, C.; Calzolai, L.; et al. Ligand-Receptor Interactions of Galectin-9 and VISTA Suppress Human T Lymphocyte Cytotoxic Activity. Front. Immunol. 2020, 11, 580557. [Google Scholar] [CrossRef]
- Grozdanovic, M.M.; Doyle, C.B.; Liu, L.; Maybruck, B.T.; Kwatia, M.A.; Thiyagarajan, N.; Acharya, K.R.; Ackerman, S.J. Charcot-Leyden crystal protein/galectin-10 interacts with cationic ribonucleases and is required for eosinophil granulogenesis. J. Allergy Clin. Immunol. 2020, 146, 377–389.e10. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Lin, H.J.; Huang, C.C.; Chen, Y.C.; Hsu, Y.A.; Lin, C.H.; Lin, H.C.; Chang, C.Y.; Huang, S.H.; Lin, J.M.; et al. Galectin-12 enhances inflammation by promoting M1 polarization of macrophages and reduces insulin sensitivity in adipocytes. Glycobiology 2016, 26, 732–744. [Google Scholar] [CrossRef]
- Hancock-Cerutti, W.; Wu, Z.; Xu, P.; Yadavalli, N.; Leonzino, M.; Tharkeshwar, A.K.; Ferguson, S.M.; Shadel, G.S.; De Camilli, P. ER-lysosome lipid transfer protein VPS13C/PARK23 prevents aberrant mtDNA-dependent STING signaling. J. Cell Biol. 2022, 221, e202106046. [Google Scholar] [CrossRef]
- Yang, T.; Yao, Y.; Wang, X.; Li, Y.; Si, Y.; Li, X.; Ayala, G.J.; Wang, Y.; Mayo, K.H.; Tai, G.; et al. Galectin-13/placental protein 13: Redox-active disulfides as switches for regulating structure, function and cellular distribution. Glycobiology 2020, 30, 120–129. [Google Scholar] [CrossRef]
- Balogh, A.; Toth, E.; Romero, R.; Parej, K.; Csala, D.; Szenasi, N.L.; Hajdu, I.; Juhasz, K.; Kovacs, A.F.; Meiri, H.; et al. Placental Galectins Are Key Players in Regulating the Maternal Adaptive Immune Response. Front. Immunol. 2019, 10, 1240. [Google Scholar] [CrossRef]
- Vokalova, L.; Balogh, A.; Toth, E.; Van Breda, S.V.; Schafer, G.; Hoesli, I.; Lapaire, O.; Hahn, S.; Than, N.G.; Rossi, S.W. Placental Protein 13 (Galectin-13) Polarizes Neutrophils Toward an Immune Regulatory Phenotype. Front. Immunol. 2020, 11, 145. [Google Scholar] [CrossRef]
- Si, Y.; Li, Y.; Yang, T.; Li, X.; Ayala, G.J.; Mayo, K.H.; Tai, G.; Su, J.; Zhou, Y. Structure-function studies of galectin-14, an important effector molecule in embryology. FEBS J. 2021, 288, 1041–1055. [Google Scholar] [CrossRef]
- Si, Y.; Yao, Y.; Jaramillo Ayala, G.; Li, X.; Han, Q.; Zhang, W.; Xu, X.; Tai, G.; Mayo, K.H.; Zhou, Y.; et al. Human galectin-16 has a pseudo ligand binding site and plays a role in regulating c-Rel-mediated lymphocyte activity. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129755. [Google Scholar] [CrossRef]
- Wada, J.; Kanwar, Y.S. Identification and characterization of galectin-9, a novel beta-galactoside-binding mammalian lectin. J. Biol. Chem. 1997, 272, 6078–6086. [Google Scholar] [CrossRef] [PubMed]
- Tureci, O.; Schmitt, H.; Fadle, N.; Pfreundschuh, M.; Sahin, U. Molecular definition of a novel human galectin which is immunogenic in patients with Hodgkin’s disease. J. Biol. Chem. 1997, 272, 6416–6422. [Google Scholar] [CrossRef]
- Teo Hansen Selno, A.; Schlichtner, S.; Yasinska, I.M.; Sakhnevych, S.S.; Fiedler, W.; Wellbrock, J.; Berger, S.M.; Klenova, E.; Gibbs, B.F.; Fasler-Kan, E.; et al. High Mobility Group Box 1 (HMGB1) Induces Toll-Like Receptor 4-Mediated Production of the Immunosuppressive Protein Galectin-9 in Human Cancer Cells. Front. Immunol. 2021, 12, 675731. [Google Scholar] [CrossRef]
- He, Y.; Jia, K.; Dziadziuszko, R.; Zhao, S.; Zhang, X.; Deng, J.; Wang, H.; Hirsch, F.R.; Zhou, C. Galectin-9 in non-small cell lung cancer. Lung Cancer 2019, 136, 80–85. [Google Scholar] [CrossRef]
- Li, E.; Xu, J.; Chen, Q.; Zhang, X.; Xu, X.; Liang, T. Galectin-9 and PD-L1 antibody blockade combination therapy inhibits tumour progression in pancreatic cancer. Immunotherapy 2023, 15, 135–147. [Google Scholar] [CrossRef]
- Blair, B.B.; Funkhouser, A.T.; Goodwin, J.L.; Strigenz, A.M.; Chaballout, B.H.; Martin, J.C.; Arthur, C.M.; Funk, C.R.; Edenfield, W.J.; Blenda, A.V. Increased Circulating Levels of Galectin Proteins in Patients with Breast, Colon, and Lung Cancer. Cancers 2021, 13, 4819. [Google Scholar] [CrossRef] [PubMed]
- Okoye, I.; Xu, L.; Motamedi, M.; Parashar, P.; Walker, J.W.; Elahi, S. Galectin-9 expression defines exhausted T cells and impaired cytotoxic NK cells in patients with virus-associated solid tumors. J. Immunother. Cancer 2020, 8, e001849. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.M.; Emam, A.; Abdelfattah, M.M.; Abdel-Mageed, A.I.; Abdelhafeez, M.A.; Helwa, R. Assessment of galectins -1, -3, -4, -8, and -9 expression in ovarian carcinoma patients with clinical implications. World J. Surg. Oncol. 2022, 20, 276. [Google Scholar] [CrossRef]
- Liu, D.; Zhu, H.; Li, C. Galectins and galectin-mediated autophagy regulation: New insights into targeted cancer therapy. Biomark Res. 2023, 11, 22. [Google Scholar] [CrossRef]
- Fuselier, C.; Dumoulin, A.; Pare, A.; Nehme, R.; Ajarrag, S.; Granger Joly de Boissel, P.; Chatenet, D.; Doucet, N.; St-Pierre, Y. Placental Galectins in Cancer: Why We Should Pay More Attention. Cells 2023, 12, 437. [Google Scholar] [CrossRef]
- Vladoiu, M.C.; Labrie, M.; St-Pierre, Y. Intracellular galectins in cancer cells: Potential new targets for therapy (Review). Int. J. Oncol. 2014, 44, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- The Human Protein Altas—Galectins. Available online: https://www.proteinatlas.org/search/Galectins (accessed on 2 April 2023).
- Kapetanakis, N.I.; Busson, P. Galectins as pivotal components in oncogenesis and immune exclusion in human malignancies. Front. Immunol. 2023, 14, 1145268. [Google Scholar] [CrossRef] [PubMed]
- Wykes, M.N.; Lewin, S.R. Immune checkpoint blockade in infectious diseases. Nat. Rev. Immunol. 2018, 18, 91–104. [Google Scholar] [CrossRef]
- Pauls, S.D.; Marshall, A.J. Regulation of immune cell signaling by SHIP1: A phosphatase, scaffold protein, and potential therapeutic target. Eur. J. Immunol. 2017, 47, 932–945. [Google Scholar] [CrossRef]
- Elahi, S.; Dinges, W.L.; Lejarcegui, N.; Laing, K.J.; Collier, A.C.; Koelle, D.M.; McElrath, M.J.; Horton, H. Protective HIV-specific CD8+ T cells evade Treg cell suppression. Nat. Med. 2011, 17, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C.; Joller, N.; Kuchroo, V.K. Lag-3, Tim-3, and TIGIT: Co-inhibitory Receptors with Specialized Functions in Immune Regulation. Immunity 2016, 44, 989–1004. [Google Scholar] [CrossRef]
- Liu, T.; Soong, L.; Liu, G.; Konig, R.; Chopra, A.K. CD44 expression positively correlates with Foxp3 expression and suppressive function of CD4+ Treg cells. Biol. Direct. 2009, 4, 40. [Google Scholar] [CrossRef]
- Nishikii, H.; Kim, B.S.; Yokoyama, Y.; Chen, Y.; Baker, J.; Pierini, A.; Alvarez, M.; Mavers, M.; Maas-Bauer, K.; Pan, Y.; et al. DR3 signaling modulates the function of Foxp3+ regulatory T cells and the severity of acute graft-versus-host disease. Blood 2016, 128, 2846–2858. [Google Scholar] [CrossRef]
- Rahmati, A.; Bigam, S.; Elahi, S. Galectin-9 promotes natural killer cells activity via interaction with CD44. Front. Immunol. 2023, 14, 1131379. [Google Scholar] [CrossRef]
- Sideras, K.; de Man, R.A.; Harrington, S.M.; Polak, W.G.; Zhou, G.; Schutz, H.M.; Pedroza-Gonzalez, A.; Biermann, K.; Mancham, S.; Hansen, B.E.; et al. Circulating levels of PD-L1 and Galectin-9 are associated with patient survival in surgically treated Hepatocellular Carcinoma independent of their intra-tumoral expression levels. Sci. Rep. 2019, 9, 10677. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, T.; Sperandio, M.; Mocsai, A. Neutrophils as emerging therapeutic targets. Nat. Rev. Drug Discov. 2020, 19, 253–275. [Google Scholar] [CrossRef] [PubMed]
- Robinson, B.S.; Arthur, C.M.; Evavold, B.; Roback, E.; Kamili, N.A.; Stowell, C.S.; Vallecillo-Zuniga, M.L.; Van Ry, P.M.; Dias-Baruffi, M.; Cummings, R.D.; et al. The Sweet-Side of Leukocytes: Galectins as Master Regulators of Neutrophil Function. Front. Immunol. 2019, 10, 1762. [Google Scholar] [CrossRef] [PubMed]
- Ustyanovska Avtenyuk, N.; Choukrani, G.; Ammatuna, E.; Niki, T.; Cendrowicz, E.; Lourens, H.J.; Huls, G.; Wiersma, V.R.; Bremer, E. Galectin-9 Triggers Neutrophil-Mediated Anticancer Immunity. Biomedicines 2021, 10, 66. [Google Scholar] [CrossRef]
- Dunsmore, G.; Rosero, E.P.; Shahbaz, S.; Santer, D.M.; Jovel, J.; Lacy, P.; Houston, S.; Elahi, S. Neutrophils promote T-cell activation through the regulated release of CD44-bound Galectin-9 from the cell surface during HIV infection. PLoS Biol. 2021, 19, e3001387. [Google Scholar] [CrossRef]
- Uribe-Querol, E.; Rosales, C. Neutrophils in Cancer: Two Sides of the Same Coin. J. Immunol. Res. 2015, 2015, 983698. [Google Scholar] [CrossRef]
- Mauracher, L.M.; Hell, L.; Moik, F.; Krall, M.; Englisch, C.; Roiss, J.; Grilz, E.; Hofbauer, T.M.; Brostjan, C.; Knapp, S.; et al. Neutrophils in lung cancer patients: Activation potential and neutrophil extracellular trap formation. Res. Pract. Thromb. Haemost. 2023, 7, 100126. [Google Scholar] [CrossRef]
- Yan, M.; Zheng, M.; Niu, R.; Yang, X.; Tian, S.; Fan, L.; Li, Y.; Zhang, S. Roles of tumor-associated neutrophils in tumor metastasis and its clinical applications. Front. Cell Dev. Biol. 2022, 10, 938289. [Google Scholar] [CrossRef]
- Li, Y.R.; Zhou, K.; Wilson, M.; Kramer, A.; Zhu, Y.; Dawson, N.; Yang, L. Mucosal-associated invariant T cells for cancer immunotherapy. Mol. Ther. 2023, 31, 631–646. [Google Scholar] [CrossRef]
- Bozorgmehr, N.; Hnatiuk, M.; Peters, A.C.; Elahi, S. Depletion of polyfunctional CD26(high)CD8(+) T cells repertoire in chronic lymphocytic leukemia. Exp. Hematol. Oncol. 2023, 12, 13. [Google Scholar] [CrossRef]
- Stowell, S.R.; Arthur, C.M.; Dias-Baruffi, M.; Rodrigues, L.C.; Gourdine, J.P.; Heimburg-Molinaro, J.; Ju, T.; Molinaro, R.J.; Rivera-Marrero, C.; Xia, B.; et al. Innate immune lectins kill bacteria expressing blood group antigen. Nat. Med. 2010, 16, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Tzeng, S.F.; Chao, T.K.; Tsai, C.Y.; Yang, Y.C.; Lee, M.T.; Hwang, J.J.; Chou, Y.C.; Tsai, M.H.; Cha, T.L.; et al. Metastatic Progression of Prostate Cancer Is Mediated by Autonomous Binding of Galectin-4-O-Glycan to Cancer Cells. Cancer Res. 2016, 76, 5756–5767. [Google Scholar] [CrossRef] [PubMed]
- Huflejt, M.E.; Leffler, H. Galectin-4 in normal tissues and cancer. Glycoconj J. 2004, 20, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Duckworth, C.A.; Fu, B.; Pritchard, D.M.; Rhodes, J.M.; Yu, L.G. Circulating galectins -2, -4 and -8 in cancer patients make important contributions to the increased circulation of several cytokines and chemokines that promote angiogenesis and metastasis. Br. J. Cancer 2014, 110, 741–752. [Google Scholar] [CrossRef]
- Danguy, A.; Rorive, S.; Decaestecker, C.; Bronckart, Y.; Kaltner, H.; Hadari, Y.R.; Goren, R.; Zich, Y.; Petein, M.; Salmon, I.; et al. Immunohistochemical profile of galectin-8 expression in benign and malignant tumors of epithelial, mesenchymatous and adipous origins, and of the nervous system. Histol. Histopathol. 2001, 16, 861–868. [Google Scholar]
- Li, Z.L.; Prakash, P.; Buck, M. A “Tug of War” Maintains a Dynamic Protein-Membrane Complex: Molecular Dynamics Simulations of C-Raf RBD-CRD Bound to K-Ras4B at an Anionic Membrane. ACS Cent. Sci. 2018, 4, 298–305. [Google Scholar] [CrossRef]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef]
- Pu, X.; Roth, J.A.; Hildebrandt, M.A.; Ye, Y.; Wei, H.; Minna, J.D.; Lippman, S.M.; Wu, X. MicroRNA-related genetic variants associated with clinical outcomes in early-stage non-small cell lung cancer patients. Cancer Res. 2013, 73, 1867–1875. [Google Scholar] [CrossRef]
- Xia, S.; Duan, W.; Liu, W.; Zhang, X.; Wang, Q. GRP78 in lung cancer. J. Transl. Med. 2021, 19, 118. [Google Scholar] [CrossRef]
- Ose, R.; Yanagawa, T.; Ikeda, S.; Ohara, O.; Koga, H. PCDH24-induced contact inhibition involves downregulation of beta-catenin signaling. Mol. Oncol. 2009, 3, 54–66. [Google Scholar] [CrossRef]
- Earl, L.A.; Bi, S.; Baum, L.G. N- and O-glycans modulate galectin-1 binding, CD45 signaling, and T cell death. J. Biol. Chem. 2010, 285, 2232–2244. [Google Scholar] [CrossRef]
- Iberg, C.A.; Hawiger, D. Natural and Induced Tolerogenic Dendritic Cells. J. Immunol. 2020, 204, 733–744. [Google Scholar] [CrossRef]
- Segura, E.; Touzot, M.; Bohineust, A.; Cappuccio, A.; Chiocchia, G.; Hosmalin, A.; Dalod, M.; Soumelis, V.; Amigorena, S. Human inflammatory dendritic cells induce Th17 cell differentiation. Immunity 2013, 38, 336–348. [Google Scholar] [CrossRef]
- Hedrick, C.C.; Malanchi, I. Neutrophils in cancer: Heterogeneous and multifaceted. Nat. Rev. Immunol. 2022, 22, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Salazar, Y.; Zheng, X.; Brunn, D.; Raifer, H.; Picard, F.; Zhang, Y.; Winter, H.; Guenther, S.; Weigert, A.; Weigmann, B.; et al. Microenvironmental Th9 and Th17 lymphocytes induce metastatic spreading in lung cancer. J. Clin. Investig. 2020, 130, 3560–3575. [Google Scholar] [CrossRef]
- Benwell, C.J.; Johnson, R.T.; Taylor, J.; Price, C.A.; Robinson, S.D. Endothelial VEGFR Coreceptors Neuropilin-1 and Neuropilin-2 Are Essential for Tumor Angiogenesis. Cancer Res. Commun. 2022, 2, 1626–1640. [Google Scholar] [CrossRef] [PubMed]
- Bannoud, N.; Stupirski, J.C.; Cagnoni, A.J.; Hockl, P.F.; Perez Saez, J.M.; Garcia, P.A.; Mahmoud, Y.D.; Gambarte Tudela, J.; Scheidegger, M.A.; Marshall, A.; et al. Circulating galectin-1 delineates response to bevacizumab in melanoma patients and reprograms endothelial cell biology. Proc. Natl. Acad. Sci. USA 2023, 120, e2214350120. [Google Scholar] [CrossRef] [PubMed]
- Corral, J.M.; Puerto-Nevado, L.D.; Cedeno, M.; Rio-Vilarino, A.; Mahillo-Fernandez, I.; Galeano, C.; Banos, N.; Garcia-Foncillas, J.; Domine, M.; Cebrian, A. Galectin-1, a novel promising target for outcome prediction and treatment in SCLC. Biomed. Pharmacother. 2022, 156, 113987. [Google Scholar] [CrossRef]
- Cagnoni, A.J.; Giribaldi, M.L.; Blidner, A.G.; Cutine, A.M.; Gatto, S.G.; Morales, R.M.; Salatino, M.; Abba, M.C.; Croci, D.O.; Marino, K.V.; et al. Galectin-1 fosters an immunosuppressive microenvironment in colorectal cancer by reprogramming CD8(+) regulatory T cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2102950118. [Google Scholar] [CrossRef]
- Zucchetti, M.; Bonezzi, K.; Frapolli, R.; Sala, F.; Borsotti, P.; Zangarini, M.; Cvitkovic, E.; Noel, K.; Ubezio, P.; Giavazzi, R.; et al. Pharmacokinetics and antineoplastic activity of galectin-1-targeting OTX008 in combination with sunitinib. Cancer Chemother. Pharmacol. 2013, 72, 879–887. [Google Scholar] [CrossRef]
- Gheysen, L.; Soumoy, L.; Trelcat, A.; Verset, L.; Journe, F.; Saussez, S. New Treatment Strategy Targeting Galectin-1 against Thyroid Cancer. Cells 2021, 10, 1112. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Arthur, C.M.; Mehta, P.; Slanina, K.A.; Blixt, O.; Leffler, H.; Smith, D.F.; Cummings, R.D. Galectin-1, -2, and -3 exhibit differential recognition of sialylated glycans and blood group antigens. J. Biol. Chem. 2008, 283, 10109–10123. [Google Scholar] [CrossRef]
- Ko, F.C.; Lee, K.W.; Tai, W.C.; Leung, Z.; Yam, J.W. Functional characterization of novel tumour suppressor galectin-2 in hepatocellular carcinoma. Eur. J. Cancer 2016, 61, S62. [Google Scholar]
- Calvisi, D.F.; Ladu, S.; Conner, E.A.; Seo, D.; Hsieh, J.T.; Factor, V.M.; Thorgeirsson, S.S. Inactivation of Ras GTPase-activating proteins promotes unrestrained activity of wild-type Ras in human liver cancer. J. Hepatol. 2011, 54, 311–319. [Google Scholar] [CrossRef]
- Calvisi, D.F.; Ladu, S.; Gorden, A.; Farina, M.; Conner, E.A.; Lee, J.S.; Factor, V.M.; Thorgeirsson, S.S. Ubiquitous activation of Ras and Jak/Stat pathways in human HCC. Gastroenterology 2006, 130, 1117–1128. [Google Scholar] [CrossRef]
- Ko, F.C.; Chan, L.K.; Tung, E.K.; Lowe, S.W.; Ng, I.O.; Yam, J.W. Akt phosphorylation of deleted in liver cancer 1 abrogates its suppression of liver cancer tumorigenesis and metastasis. Gastroenterology 2010, 139, 1397–1407. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Krasnitz, A.; Lucito, R.; Sordella, R.; Vanaelst, L.; Cordon-Cardo, C.; Singer, S.; Kuehnel, F.; Wigler, M.; Powers, S.; et al. DLC1 is a chromosome 8p tumor suppressor whose loss promotes hepatocellular carcinoma. Genes Dev. 2008, 22, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Ji, P.; Gong, Y.; Jin, M.L.; Wu, H.L.; Guo, L.W.; Pei, Y.C.; Chai, W.J.; Jiang, Y.Z.; Liu, Y.; Ma, X.Y.; et al. In vivo multidimensional CRISPR screens identify Lgals2 as an immunotherapy target in triple-negative breast cancer. Sci. Adv. 2022, 8, eabl8247. [Google Scholar] [CrossRef]
- Than, N.G.; Romero, R.; Xu, Y.; Erez, O.; Xu, Z.; Bhatti, G.; Leavitt, R.; Chung, T.H.; El-Azzamy, H.; LaJeunesse, C.; et al. Evolutionary origins of the placental expression of chromosome 19 cluster galectins and their complex dysregulation in preeclampsia. Placenta 2014, 35, 855–865. [Google Scholar] [CrossRef]
- Liu, J.; Liu, J.; Lu, X. HOXA1 upregulation is associated with poor prognosis and tumor progression in breast cancer. Exp. Ther. Med. 2019, 17, 1896–1902. [Google Scholar] [CrossRef]
- Zha, T.Z.; Hu, B.S.; Yu, H.F.; Tan, Y.F.; Zhang, Y.; Zhang, K. Overexpression of HOXA1 correlates with poor prognosis in patients with hepatocellular carcinoma. Tumour Biol. 2012, 33, 2125–2134. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Xu, Y.; Wang, P.; Xu, Y.; Jin, P.; Wu, Z.; Qian, Y.; Bai, L.; Dong, M. Galectin-14 Promotes Trophoblast Migration and Invasion by Upregulating the Expression of MMP-9 and N-Cadherin. Front Cell Dev. Biol. 2021, 9, 645658. [Google Scholar] [CrossRef] [PubMed]
- Kaminker, J.D.; Timoshenko, A.V. Expression, Regulation, and Functions of the Galectin-16 Gene in Human Cells and Tissues. Biomolecules 2021, 11, 1909. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.C.; Nesmelova, I.V.; Daragan, V.A.; Ippel, H.; Michalak, M.; Dregni, A.; Kaltner, H.; Kopitz, J.; Gabius, H.J.; Mayo, K.H. Pro4 prolyl peptide bond isomerization in human galectin-7 modulates the monomer-dimer equilibrum to affect function. Biochem. J. 2020, 477, 3147–3165. [Google Scholar] [CrossRef] [PubMed]
- Laderach, D.J.; Compagno, D. Inhibition of galectins in cancer: Biological challenges for their clinical application. Front. Immunol. 2022, 13, 1104625. [Google Scholar] [CrossRef] [PubMed]
- Bertuzzi, S.; Gimeno, A.; Martinez-Castillo, A.; Lete, M.G.; Delgado, S.; Airoldi, C.; Rodrigues Tavares, M.; Blahova, M.; Chytil, P.; Kren, V.; et al. Cross-Linking Effects Dictate the Preference of Galectins to Bind LacNAc-Decorated HPMA Copolymers. Int. J. Mol. Sci. 2021, 22, 6000. [Google Scholar] [CrossRef]
- Wang, T.; Cai, S.; Cheng, Y.; Zhang, W.; Wang, M.; Sun, H.; Guo, B.; Li, Z.; Xiao, Y.; Jiang, S. Discovery of Small-Molecule Inhibitors of the PD-1/PD-L1 Axis That Promote PD-L1 Internalization and Degradation. J. Med. Chem. 2022, 65, 3879–3893. [Google Scholar] [CrossRef]
- Croese, T.; Castellani, G.; Schwartz, M. Immune cell compartmentalization for brain surveillance and protection. Nat. Immunol. 2021, 22, 1083–1092. [Google Scholar] [CrossRef]
- Vafaei, S.; Zekiy, A.O.; Khanamir, R.A.; Zaman, B.A.; Ghayourvahdat, A.; Azimizonuzi, H.; Zamani, M. Combination therapy with immune checkpoint inhibitors (ICIs); a new frontier. Cancer Cell Int. 2022, 22, 2. [Google Scholar] [CrossRef]
- Eruslanov, E.B.; Singhal, S.; Albelda, S.M. Mouse versus Human Neutrophils in Cancer: A Major Knowledge Gap. Trends Cancer 2017, 3, 149–160. [Google Scholar] [CrossRef]
- Douam, F.; Ploss, A. A humanized “new-trophil” mouse to study early inflammatory processes. Proc. Natl. Acad. Sci. USA 2022, 119, e2216699119. [Google Scholar] [CrossRef] [PubMed]
- Socinski, M.A.; Jotte, R.M.; Cappuzzo, F.; Nishio, M.; Mok, T.S.K.; Reck, M.; Finley, G.G.; Kaul, M.D.; Yu, W.; Paranthaman, N.; et al. Association of Immune-Related Adverse Events With Efficacy of Atezolizumab in Patients With Non-Small Cell Lung Cancer: Pooled Analyses of the Phase 3 IMpower130, IMpower132, and IMpower150 Randomized Clinical Trials. JAMA Oncol. 2023, 9, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, B.; Vajkoczy, P.; Weller, R.O. The movers and shapers in immune privilege of the CNS. Nat. Immunol. 2017, 18, 123–131. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).