
Timing of Chromosome DNA Integration throughout the Yeast Cell Cycle

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Media
2.2. Molecular Biology Techniques and Microscopy
2.3. Transformation and Synchronization Procedure
2.4. Measurement of ROS
2.5. p-Value Calculation
Relative Efficiency (Er) Calculation
3. Results
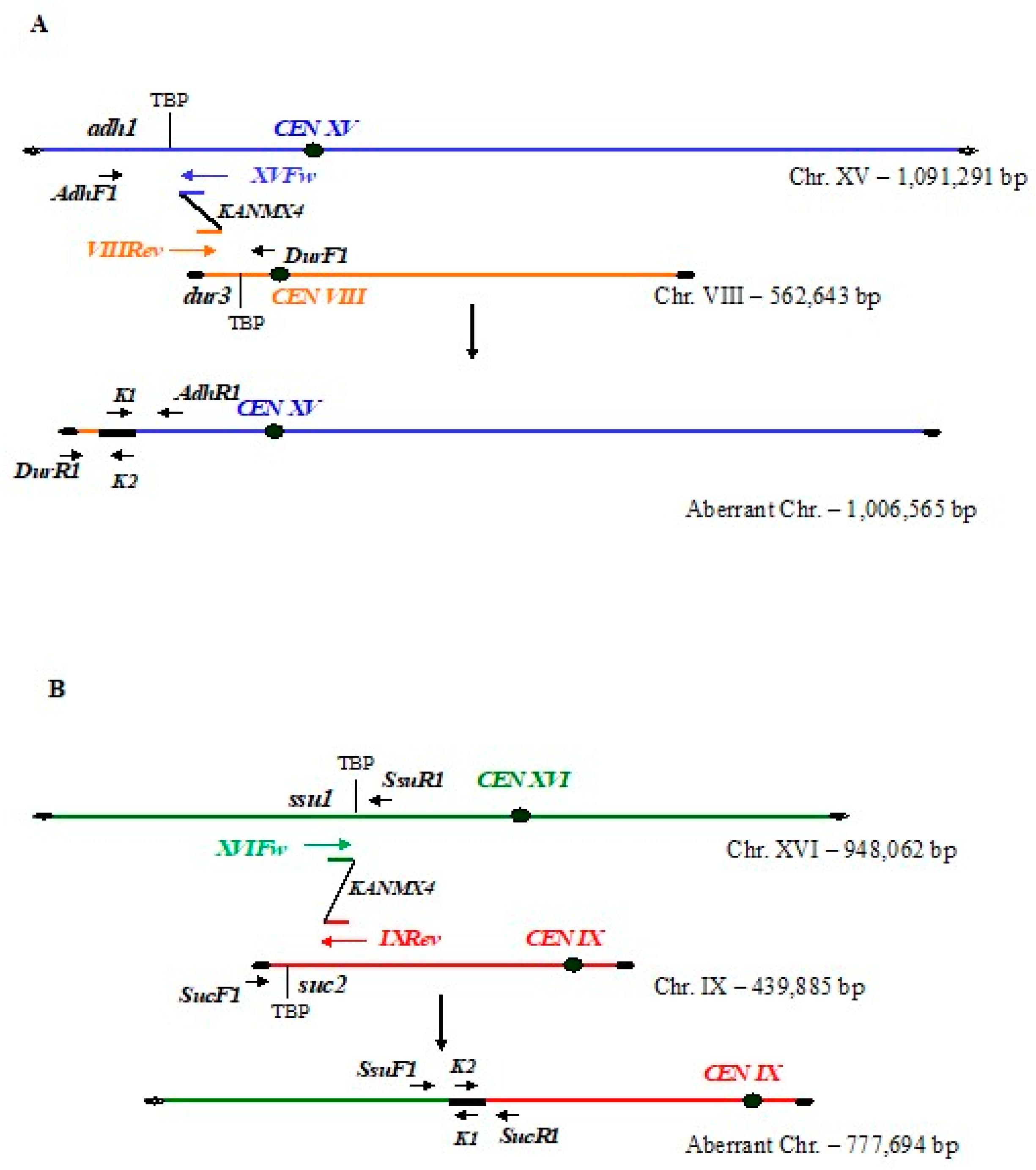
3.1. Integration of Bridge-Induced Translocation and Site-Specific DNA Cassettes
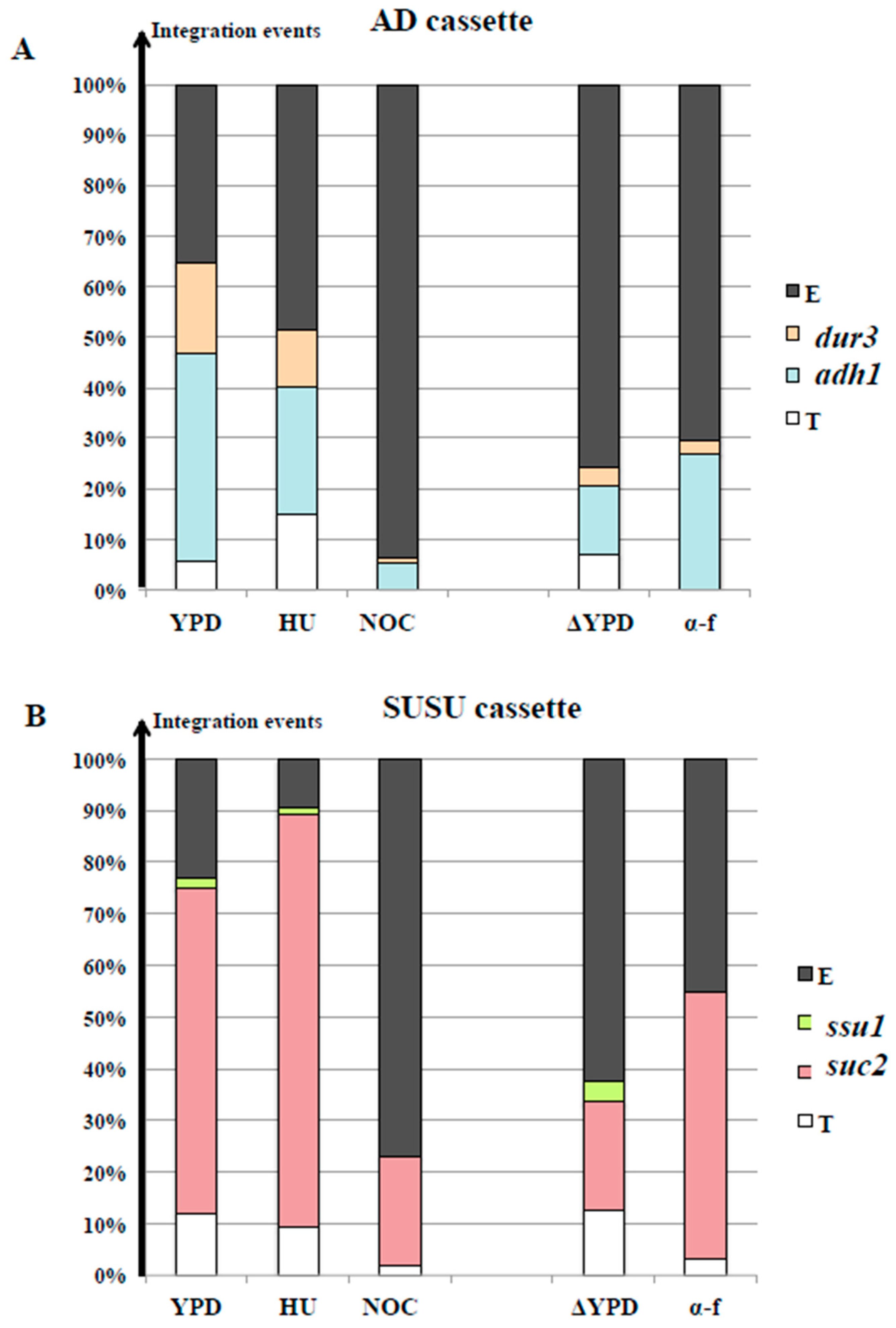
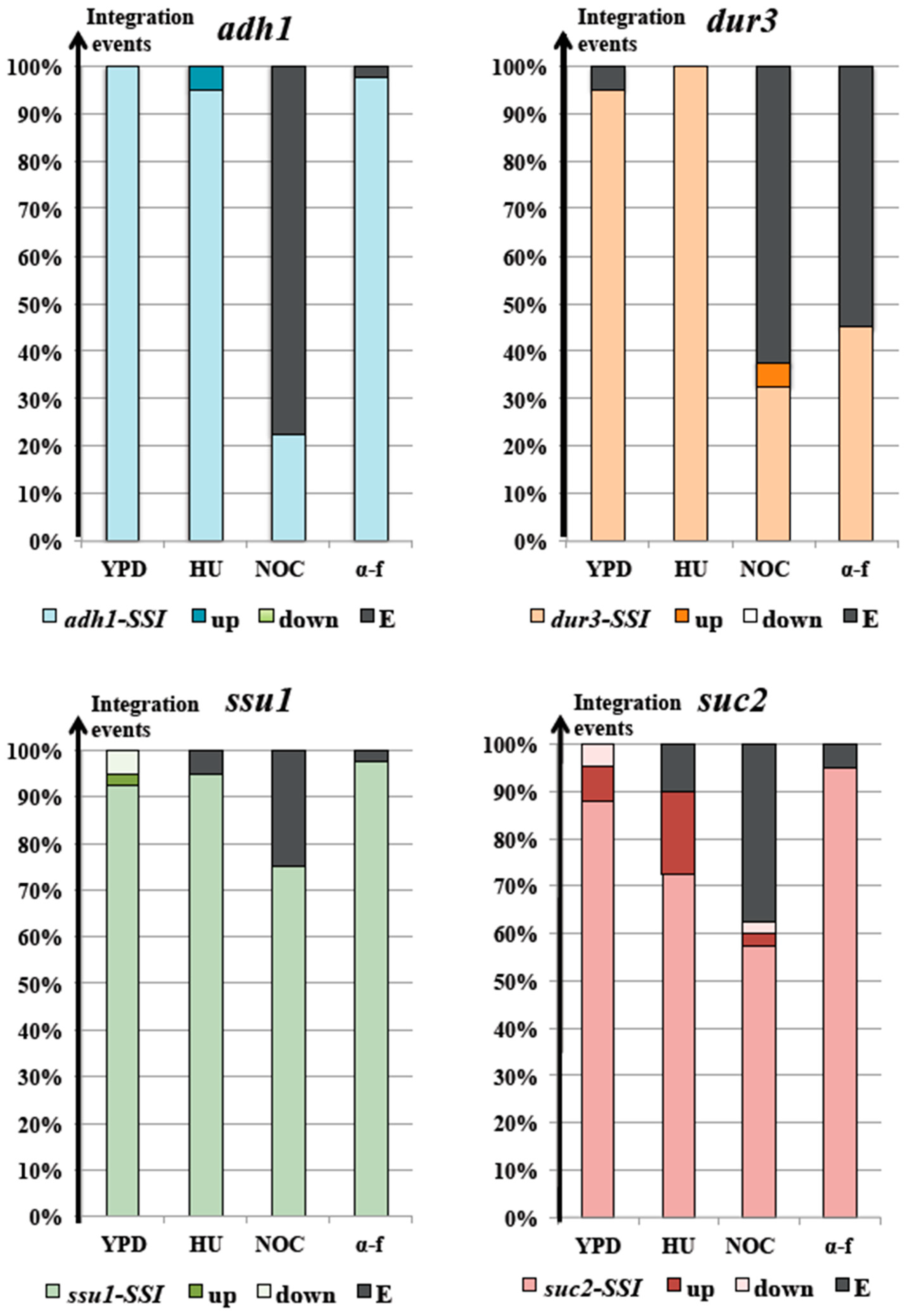
3.2. The Locus and the Cell Cycle Phase During Integration Strongly Affected Transformation Efficiency with Linear DNA Molecules
3.3. The S Phase May Have a Leading Role in Translocation Success
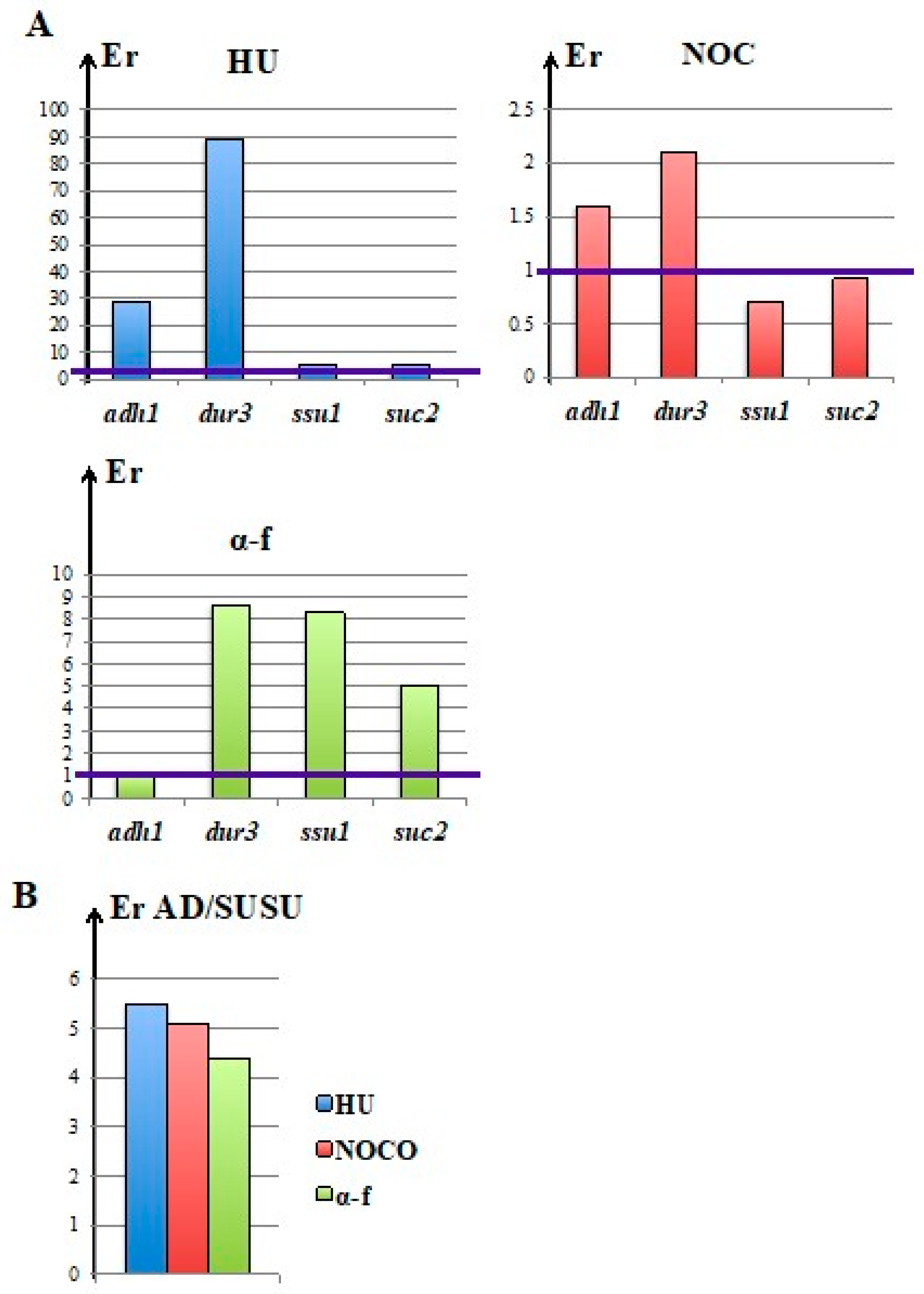
3.4. Nocodazole and α-Factor Strongly Decreased the Occurrence of Homologous Integration and Chromosomal Translocation
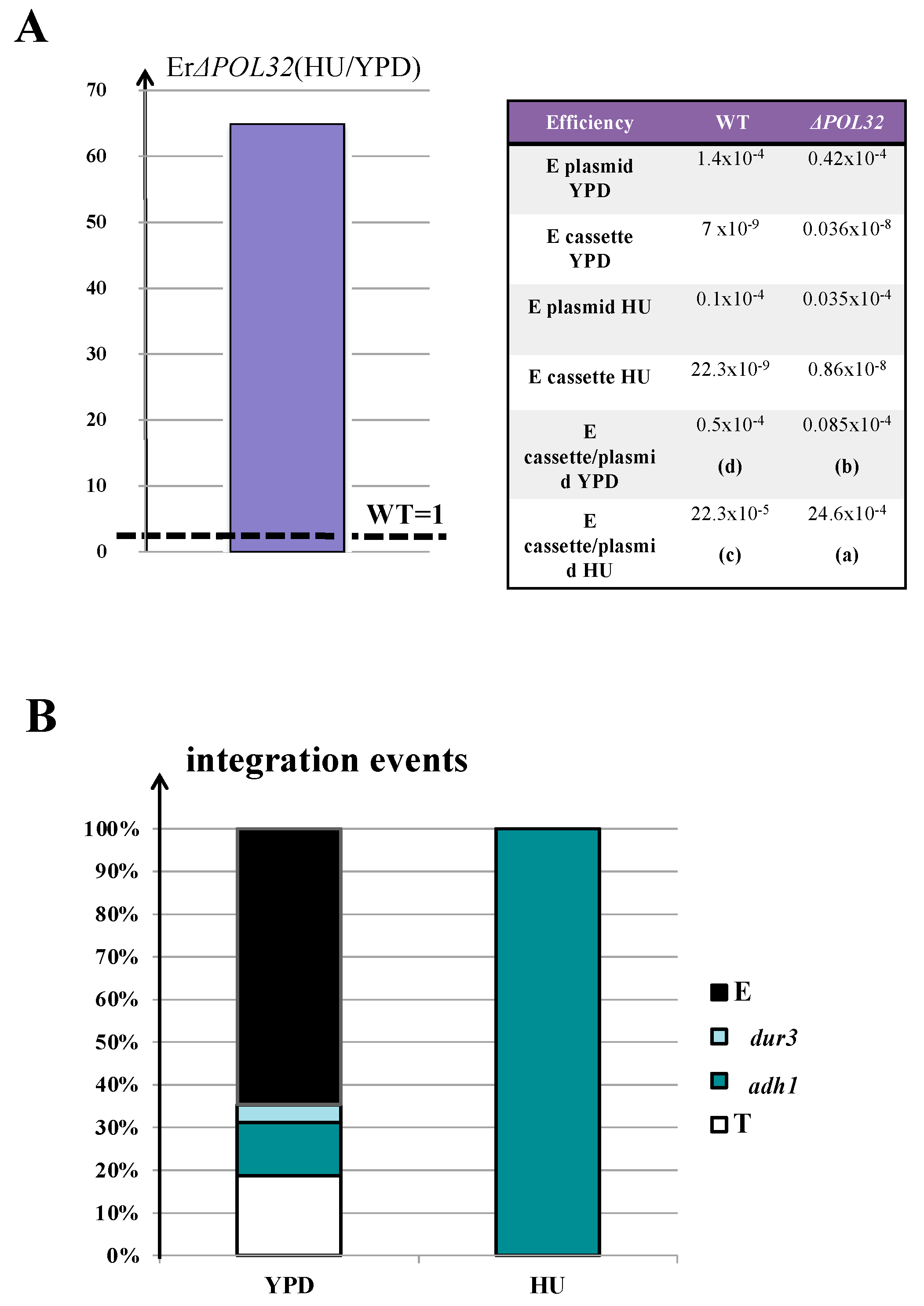
3.5. The Enhancement of AD-Translocation Frequency during Synthesis Is Pol32-Dependent
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tosato, V.; Waghmare, S.K.; Bruschi, C.V. Non-reciprocal chromosomal bridge-induced translocation (BIT) by targeted DNA integration in yeast. Chromosoma 2005, 114, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Sims, J.; Bruschi, C.V.; Bertin, C.; West, N.; Breitenbach, M.; Schroeder, S.; Eisenberg, T. High reactive oxygen species levels are detected at the end of the chronological life span of translocant yeast cells. Mol. Genet. Genom. 2016, 291, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Tosato, V.; Sims, J.; West, N.; Colombin, M.; Bruschi, C.V. Post-translocational adaptation drives evolution through genetic selection and transcriptional shift in S. cerevisiae. Curr. Genet. 2017, 63, 281–292. [Google Scholar] [CrossRef]
- Nikitin, D.; Tosato, V.; Zavec, A.B.; Bruschi, C.V. Cellular and molecular effects of nonreciprocal chromosome translocation in S. cerevisiae. Proc. Natl. Acad. Sci. USA 2008, 105, 9703–9708. [Google Scholar] [CrossRef] [PubMed]
- Bruschi, C.V.; Comer, A.R.; Howe, G.A. Specificity of DNA uptake during whole cell transformation of S. cerevisiae. Yeast 1987, 3, 131–137. [Google Scholar] [CrossRef]
- Chaustova, L.; Zimkus, A. Study of cell wall permeability properties of synchronous S. cerevisiae cells in different phases of cell cycle. Biologija 2005, 1, 38–40. [Google Scholar]
- Tsakraklides, V.; Brevnova, E.; Stephanopoulos, G.; Shaw, A.J. Improved gene targeting through cell cycle synchronization. PLoS ONE 2015, 10, e0133434. [Google Scholar] [CrossRef]
- Prado, F. Genetic instability is prevented by Mrc1-dependent spatio-temporal separation of replicative and repair activities of homologous recombination. BioEssays 2014, 36, 451–462. [Google Scholar] [CrossRef]
- Tosato, V.; Sidari, S.; Bruschi, C.V. Bridge-induced chromosome translocation in yeast relies upon a Rad/Rdh54 dependent, Pol32-independ pathway. PLoS ONE 2013, 8, e60926. [Google Scholar] [CrossRef]
- Lydeard, J.R.; Jain, S.; Yamaguchi, M.; Haber, J.E. Break-induced replication and telomerase-independent telomere maintenance require Pol32. Nature 2007, 448, 820–823. [Google Scholar] [CrossRef]
- Falbo, K.B.; Alabert, C.; Katou, Y.; Wu, S.; Han, J.; Wehr, T.; Xiao, J.; He, X.; Zhang, Z.; Shi, Y.; et al. Involvement of a chromatin remodeling complex in damage tolerance during DNA replication. Nat. Struct. Mol. Biol. 2009, 16, 1167–1172. [Google Scholar] [CrossRef]
- Bruschi, C.V.; Howe, G.A. High frequency FLP-independent homologous DNA recombination of 2μ plasmid in the yeast S. cerevisiae. Curr. Genet. 1988, 14, 191–199. [Google Scholar] [CrossRef]
- Lee, P.S.; Greenwell, P.W.; Dominska, M.; Gawel, M.; Hamilton, M.; Petes, T.D. A fine-structure map of spontaneous mitotic crossovers in the yeast S. cerevisiae. PLoS Genet. 2009, 5, e1000410. [Google Scholar] [CrossRef]
- Storici, F.; Coglievina, M.; Bruschi, C.V. A 2-micron DNA-based marker recycling system for multiple gene disruption in the yeast Saccharomyces cerevisiae. Yeast 1999, 15, 271–283. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Analysis of genomic DNA by Southern hybridization. In Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Wach, A.; Brachat, A.; Pöhlmann, R.; Philippsen, P. New heterologous modules for classical or PCR-based gene disruptions in Saccharomyces cerevisiae. Yeast 1994, 10, 1793–1808. [Google Scholar] [CrossRef] [PubMed]
- Rossi, B.; Noel, P.; Bruschi, C.V. Different aneuploidies arise from the same bridge-induced chromosomal translocation event in Saccharomyces cerevisiae. Genetics 2010, 186, 775–790. [Google Scholar] [CrossRef][Green Version]
- Lieberman, H.B. Cell Cycle Checkpoint Control Protocols. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2004. [Google Scholar]
- Rose, M.D.; Novick, P.; Thomas, J.H.; Botstein, D.; Fink, G.R. A Saccharomyces cerevisiae genomic plasmid bank based on a centromere-containing shuttle vector. Gene 1987, 60, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, K.A.; Michelson, R.J.; Putnam, C.W.; Weinert, T.A. Toward maintaining the genome: DNA damage and replication checkpoints. Annu. Rev. Genet. 2002, 36, 617–656. [Google Scholar] [CrossRef]
- Branzei, D.; Foiani, M. Regulation of DNA repair throughout the cell cycle. Nat. Rev. Mol. Cell. Biol. 2008, 9, 297–308. [Google Scholar] [CrossRef]
- Symington, L.S. Mechanism and regulation of DNA end resection in Eukaryotes. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 195–212. [Google Scholar] [CrossRef]
- Moore, J.K.; Haber, J.E. Cell cycle and genetic requirements of two pathways of nonhomologous end-joining repair of double-strand breaks in S. cerevisiae. Mol. Cell. Biol. 1996, 16, 2164–2173. [Google Scholar] [CrossRef]
- Aylon, Y.; Liefshitz, B.; Kupiec, M. The CDK regulates repair of double-strand breaks by homologous recombination during the cell cycle. EMBO J. 2004, 23, 4868–4875. [Google Scholar] [CrossRef]
- Tosato, V.; Nicolini, C.; Bruschi, C.V. DNA bridging of yeast chromosomes VIII leads to near-reciprocal translocation and loss of heterozygosity with minor cellular defects. Chromosoma 2009, 118, 179–191. [Google Scholar] [CrossRef]
- Tosato, V.; Bruschi, C.V. Per aspera ad astra: When harmful chromosomal translocation become a plus value in genetic evolution. Lessons from S. cerevisiae. Microb. Cell. 2015, 2, 363–375. [Google Scholar] [CrossRef]
- Liu, H.; Liang, F.; Jin, F.; Wang, Y. The coordination of centromere replication, spindle formation and kinetochore microtubule interaction in budding yeast. PLoS Genet. 2008, 4, e1000262. [Google Scholar] [CrossRef]
- Klinner, U.; Schäfer, B. Genetic aspects of targeted insertion mutagenesis in yeast. FEMS Microbiol. Rev. 2004, 28, 201–223. [Google Scholar] [CrossRef][Green Version]
- Gerton, J.L.; DeRisi, J.; Shroff, R.; Lichten, M.; Brown, P.O.; Petes, T.D. Global mapping of meiotic recombination hotspots and coldspots in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2000, 97, 11383–11390. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Beach, A.; Haber, J.E. Homology requirements and competition between Gene Conversion and Break-Induced Replication during Double-Strand Break Repair. Mol. Cell. 2017, 65, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Deem, A.; Keszthelyi, A.; Blackgrove, T.; Vayl, A.; Coffey, B.; Mathur, R.; Chabes, A.; Malkova, A. Break-induced replication is highly inaccurate. PLoS Biol. 2011, 9, e1000594. [Google Scholar] [CrossRef] [PubMed]
- Donnianni, R.A.; Symington, L.S. Break-induced replication occurs by conservative DNA synthesis. Proc. Natl. Acad. Sci. USA 2013, 110, 13475–13480. [Google Scholar] [CrossRef]
- Ruiz, J.F.; Gómez-González, B.; Aguilera, A. Chromosomal translocations caused by either pol32-dependent or pol32-independent triparental break-induced replication. Mol. Cell. Biol. 2009, 29, 5441–5454. [Google Scholar] [CrossRef] [PubMed]
- Malkova, A.; Naylor, M.L.; Yamaguchi, M.; Ira, G.; Haber, J.E. RAD51-dependent break-induced replication differs in kinetics and checkpoint responses from RAD51-mediated gene conversion. Mol. Cell. Biol. 2005, 25, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Karras, G.I.; Jentsch, S. The RAD6 DNA damage tolerance pathway operates uncoupled from the replication fork and is functional behyond S phase. Cell 2010, 141, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Burgers, P.M.; Gerik, K.J. Structure and processivity of two forms of Saccharomyces cerevisiae DNA polymerase delta. J. Biol. Chem. 1998, 273, 19756–19762. [Google Scholar] [CrossRef]
- Tumini, E.; Barroso, S.; Perez-Calero, C.; Aguilera, A. Roles of human POLD1 and POLD3 in genome stability. Sci. Rep 2016, 6, 38873. [Google Scholar] [CrossRef]
- Dunlop, M.G.; Dobbins, S.E.; Farrington, S.M.; Jones, A.M.; Palles, C.; Whiffin, N.; Tenesa, A.; Spain, S.; Broderick, P.; Ooi, L.Y.; et al. Common variation near CDKN1A, POLD3 and SHROOM2 influences colorectal cancer risk. Nat. Genet. 2012, 44, 770–776. [Google Scholar] [CrossRef]
- Huang, M.E.; Le Douarin, B.; Henry, C.; Galibert, F. The Saccharomyces cerevisiae protein YJR043C (Pol32) interacts with the catalytic subunit of DNA polymerase alpha and is required for cell cycle progression in G2/M. Mol. Gen. Genet. 1999, 260, 541–550. [Google Scholar] [CrossRef]
- Jain, S.; Sugawara, N.; Lydeard, J.; Vaze, M.; Le Gac, N.T.; Haber, J.E. A recombination execution checkpoint regulates the choice of homologous recombination pathway during DNA double-strand break repair. Genes Dev. 2009, 23, 291–303. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tosato, V.; Rossi, B.; Sims, J.; Bruschi, C.V. Timing of Chromosome DNA Integration throughout the Yeast Cell Cycle. Biomolecules 2023, 13, 614. https://doi.org/10.3390/biom13040614
Tosato V, Rossi B, Sims J, Bruschi CV. Timing of Chromosome DNA Integration throughout the Yeast Cell Cycle. Biomolecules. 2023; 13(4):614. https://doi.org/10.3390/biom13040614
Chicago/Turabian StyleTosato, Valentina, Beatrice Rossi, Jason Sims, and Carlo V. Bruschi. 2023. "Timing of Chromosome DNA Integration throughout the Yeast Cell Cycle" Biomolecules 13, no. 4: 614. https://doi.org/10.3390/biom13040614
APA StyleTosato, V., Rossi, B., Sims, J., & Bruschi, C. V. (2023). Timing of Chromosome DNA Integration throughout the Yeast Cell Cycle. Biomolecules, 13(4), 614. https://doi.org/10.3390/biom13040614