
Proteasome-Associated Proteins, PA200 and ECPAS, Are Essential for Murine Spermatogenesis
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
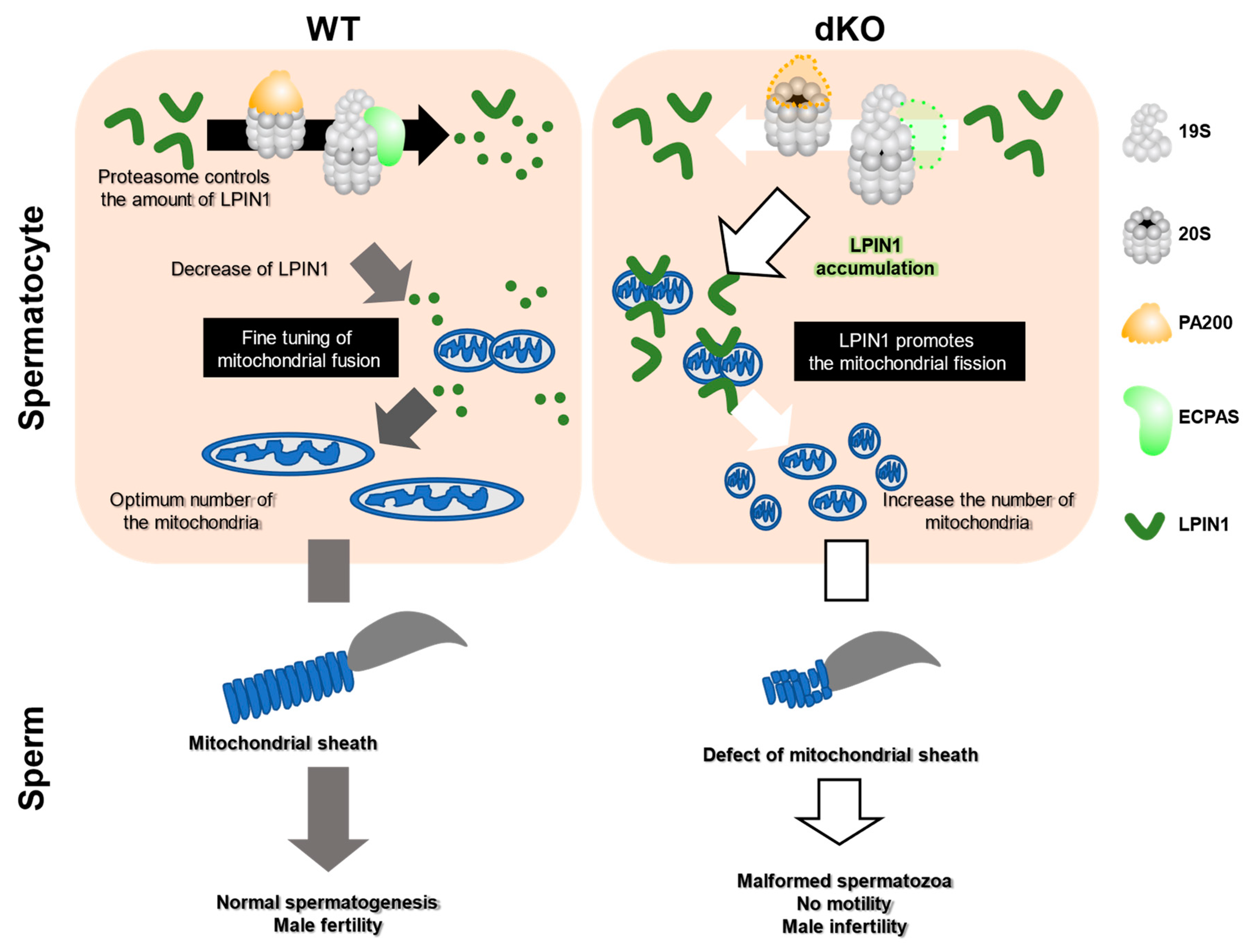
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Antibodies (Abs)
2.3. Production of dKO Mice
2.4. Proteasome Activity
2.5. Histochemistry and Immunohistochemistry
2.6. Mass Spectrometry
2.7. Transmission Electron Microscopy (TEM)
2.8. Analysis of Mitochondrial Morphology
2.9. Statistical Analysis
3. Results
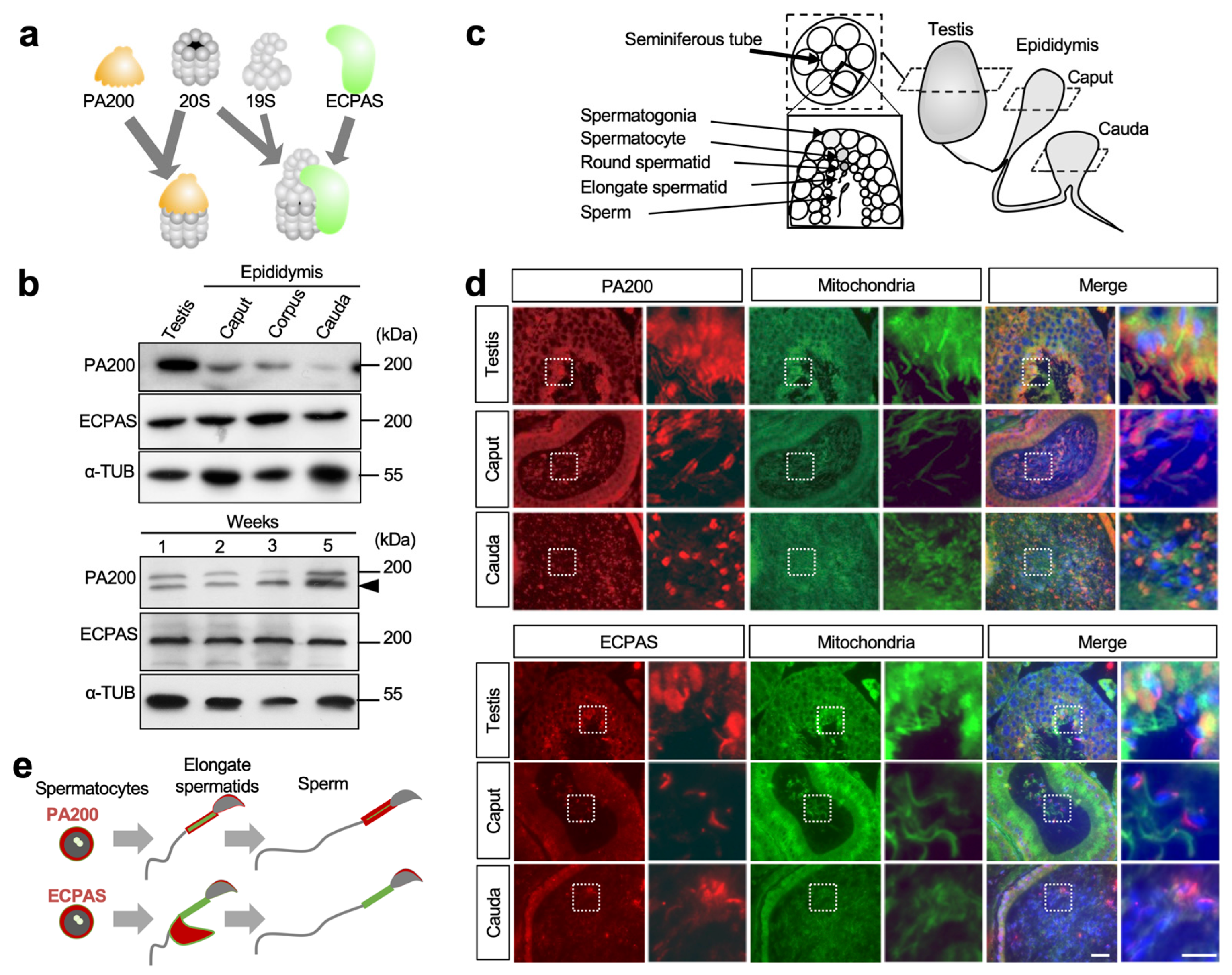
3.1. Expression Patterns of PA200 and Ecpas in Testes and Epididymides
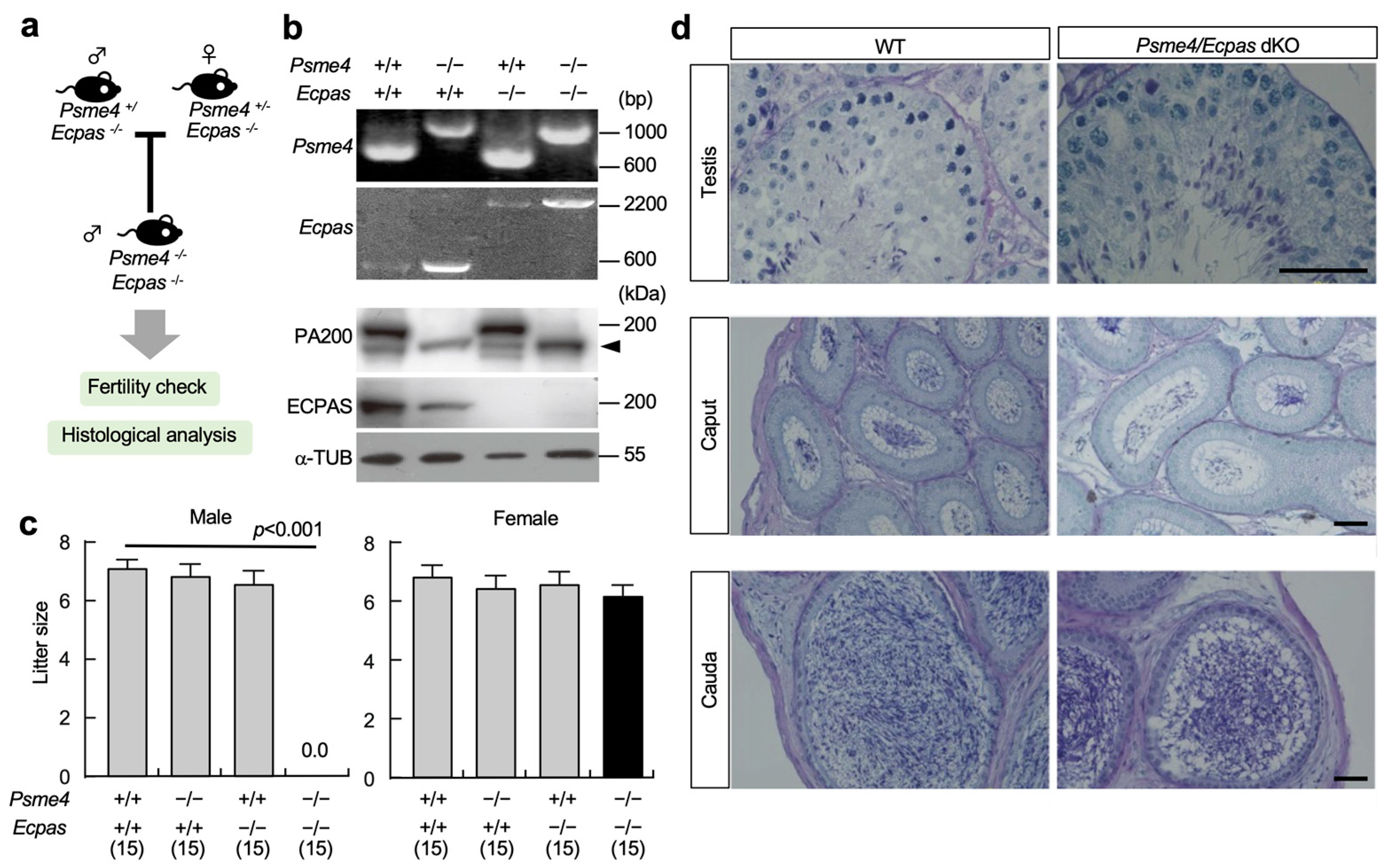
3.2. Fertility of dKO Male Mice
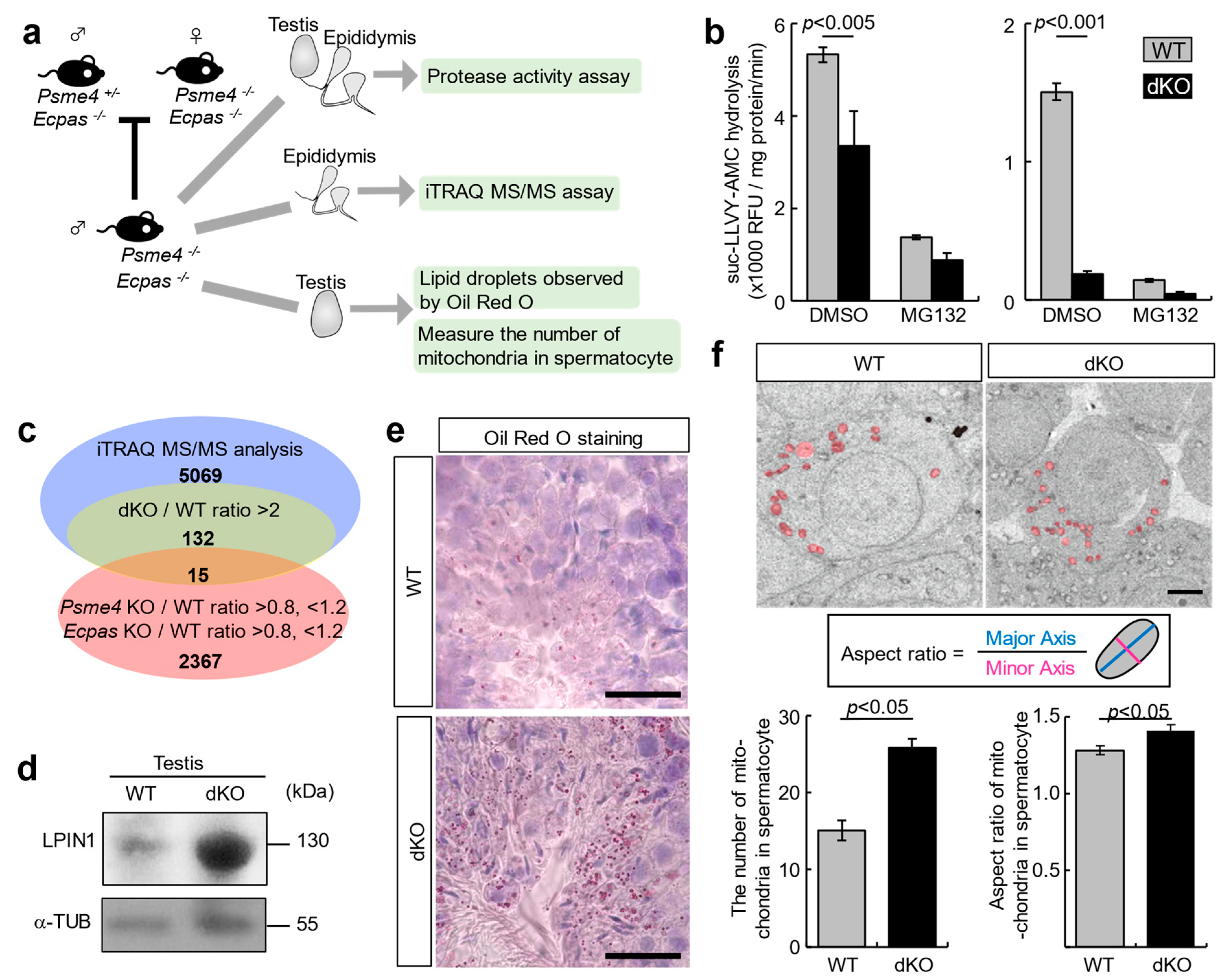
3.3. LPIN1 as a Target Protein of PA200 and ECPAS
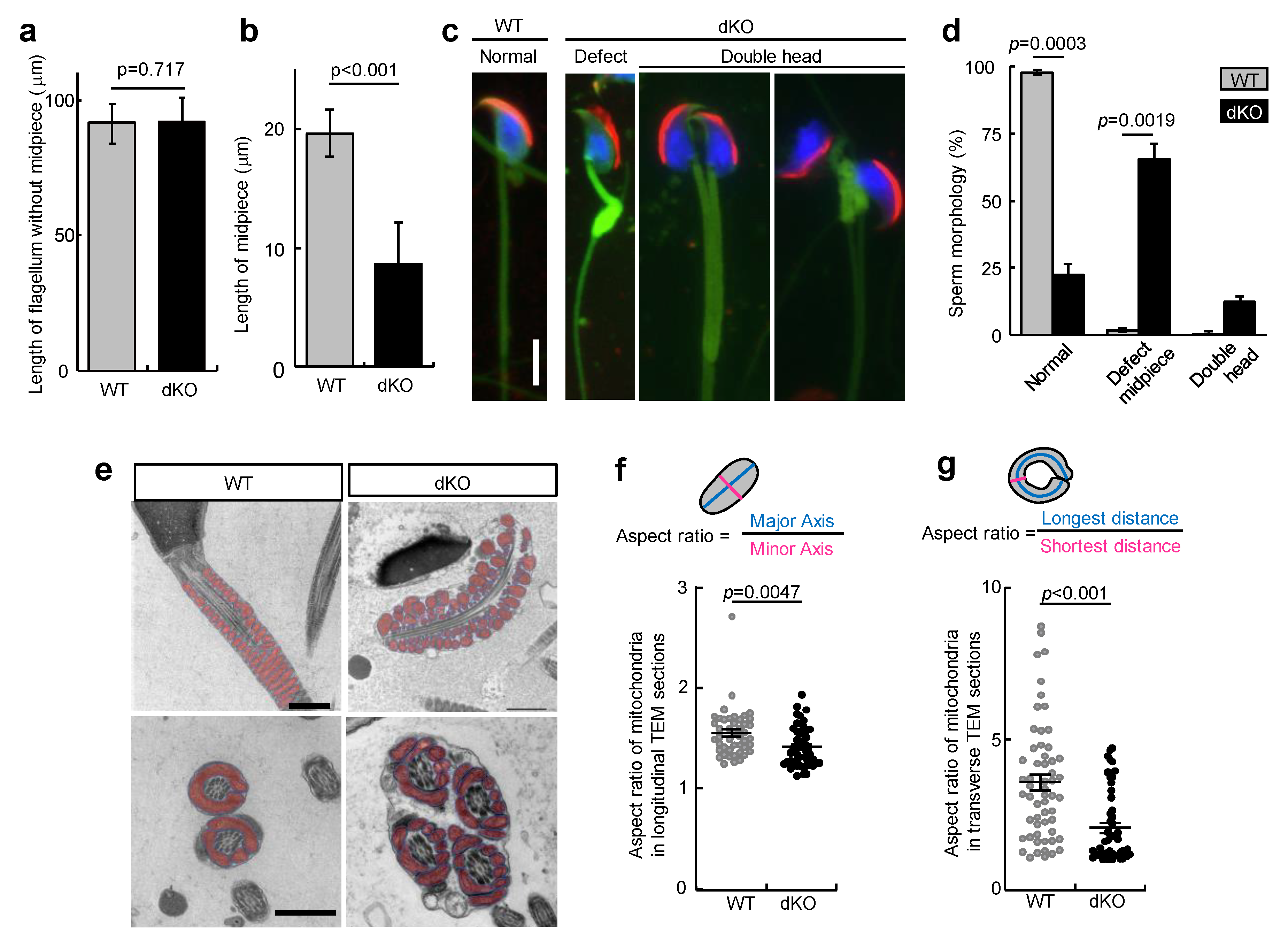
3.4. Loss of PA200 and ECPAS Causes Midpiece Defects in Sperm
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coux, O.; Tanaka, K.; Goldberg, A.L. Structure and Functions of the 20S and 26S Proteasomes. Annu. Rev. Biochem. 1996, 65, 801–847. [Google Scholar] [CrossRef]
- Baumeister, W.J.; Walz, J.; Zühl, F.; Seemüller, E. The Proteasome: Paradigm of a Self-Compartmentalizing Protease. Cell 1998, 92, 367–380. [Google Scholar] [CrossRef]
- Collins, G.A.; Goldberg, A.L. The Logic of the 26s Proteasome. Cell 2017, 169, 792–806. [Google Scholar] [CrossRef] [PubMed]
- Sawada, H.; Saito, T. Mechanisms of Sperm-Egg Interactions: What Ascidian Fertilization Research Has Taught Us. Cells 2022, 11, 2096. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, A.; Bertolotti, A. Regulation of Proteasome Assembly and Activity in Health and Disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 697–712. [Google Scholar] [CrossRef] [PubMed]
- Murata, S.; Sasaki, K.; Kishimoto, T.; Niwa, S.-i.; Hayashi, H.; Takahama, Y.; Tanaka, K. Regulation of CD8+ T Cell Development by Thymus-Specific Proteasomes. Science 2007, 316, 1349–1353. [Google Scholar] [CrossRef]
- Kniepert, A.; Groettrup, M. The Unique Functions of Tissue-Specific Proteasomes. Trends Biochem. Sci. 2014, 39, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Gómez-H, L.; Felipe-Medina, N.; Condezo, Y.B.; Garcia-Valiente, R.; Ramos, I.; Suja, J.A.; Barbero, J.L.; Roig, I.; Sánchez-Martín, M.; de Rooij, D.G.; et al. The PSMA8 Subunit of the Spermatoproteasome is Essential for Proper Meiotic Exit and Mouse Fertility. PLoS Genet. 2019, 15, e1008316. [Google Scholar] [CrossRef]
- Zhang, Q.; Ji, S.Y.; Busayavalasa, K.; Shao, J.; Yu, C. Meiosis I Progression in Spermatogenesis Requires a Type of Testis-Specific 20S Core Proteasome. Nat. Commun. 2019, 10, 3387. [Google Scholar] [CrossRef]
- Schmidt, M.; Hanna, J.; Elsasser, S.; Finley, D. Proteasome-Associated Proteins: Regulation of a Proteolytic Machine. Biol. Chem. 2005, 386, 725–737. [Google Scholar] [CrossRef]
- Schmidt, M.; Haas, W.; Crosas, B.; Santamaria, P.G.; Gygi, S.P.; Walz, T.; Finley, D. The HEAT Repeat Protein Blm10 Regulates the Yeast Proteasome by Capping the Core Particle. Nat. Struct. Mol. Biol. 2005, 12, 294–303. [Google Scholar] [CrossRef]
- Dange, T.; Smith, D.; Noy, T.; Rommel, P.C.; Jurzitza, L.; Cordero, R.J.; Legendre, A.; Finley, D.; Goldberg, A.L.; Schmidt, M. Blm10 Protein Promotes Proteasomal Substrate Turnover by an Active Gating Mechanism. J. Biol. Chem. 2011, 286, 42830–42839. [Google Scholar] [CrossRef]
- Qian, M.X.; Pang, Y.; Liu, C.H.; Haratake, K.; Du, B.Y.; Ji, D.Y.; Wang, G.F.; Zhu, Q.Q.; Song, W.; Yu, Y.; et al. Acetylation-Mediated Proteasomal Degradation of Core Histones during DNA Repair and Spermatogenesis. Cell 2013, 153, 1012–1024. [Google Scholar] [CrossRef]
- Gorbea, C.; Goellner, G.M.; Teter, K.; Holmes, R.K.; Rechsteiner, M. Characterization of Mammalian Ecm29, a 26 S Proteasome-Associated Protein That Localizes to the Nucleus and Membrane Vesicles. J. Biol. Chem. 2004, 279, 54849–54861. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, A.; Niewienda, A.; Jechow, K.; Janek, K.; Enenkel, C. Ecm29 Fulfils Quality Control Functions in Proteasome Assembly. Mol. Cell 2010, 38, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Jones, S.; Minis, A.; Rodriguez, J.; Molina, H.; Steller, H. PI31 is an Adaptor Protein for Proteasome Transport in Axons and Required for Synaptic Development. Dev. Cell 2019, 50, 509–524.e510. [Google Scholar] [CrossRef] [PubMed]
- Živković, D.; Sanchez Dafun, A.; Menneteau, T.; Schahl, A.; Lise, S.; Kervarrec, C.; Toste Rêgo, A.; da Fonseca, P.C.A.; Chavent, M.; Pineau, C.; et al. Proteasome Complexes Experience Profound Structural and Functional Rearrangements throughout Mammalian Spermatogenesis. Proc. Natl. Acad. Sci. USA 2022, 119, e2116826119. [Google Scholar] [CrossRef]
- Khor, B.; Bredemeyer, A.L.; Huang, C.Y.; Turnbull, I.R.; Evans, R.; Maggi, L.B., Jr.; White, J.M.; Walker, L.M.; Carnes, K.; Hess, R.A.; et al. Proteasome Activator PA200 Is Required for Normal Spermatogenesis. Mol. Cell. Biol. 2006, 26, 2999–3007. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Haratake, K.; Miyahara, H.; Chiba, T. Proteasome Activators, PA28γ and PA200, Play Indispensable Roles in Male Fertility. Sci. Rep. 2016, 6, 23171. [Google Scholar] [CrossRef]
- Kondo, H.; Matsumura, T.; Kaneko, M.; Inoue, K.; Kosako, H.; Ikawa, M.; Takahama, Y.; Ohigashi, I. PITHD1 Is a Proteasome-Interacting Protein Essential for Male Fertilization. J. Biol. Chem. 2020, 295, 1658–1672. [Google Scholar] [CrossRef]
- Kajava, A.V.; Gorbea, C.; Ortega, J.n.; Rechsteiner, M.; Steven, A.C. New HEAT-like Repeat Motifs in Proteins Regulating Proteasome Structure and Function. J. Struct. Biol. 2004, 146, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Haratake, K.; Sato, A.; Tsuruta, F.; Chiba, T. KIAA0368-Deficiency Affects Disassembly of 26S Proteasome under Oxidative Stress Condition. J. Biochem. 2016, 159, 609–618. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, Z.; Tao, Y.; Zhu, J.; Yang, H.; Liang, W.; Ding, G. Increased Mitochondrial Fission of Glomerular Podocytes in Diabetic Nephropathy. Endocr. Connect. 2019, 8, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Bellve, A.R.; Cavicchia, J.; Millette, C.F.; O’brien, D.A.; Bhatnagar, Y.; Dym, M. Spermatogenic Cells of the Prepuberal Mouse: Isolation and Morphological Characterization. J. Cell Biol. 1977, 74, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Langner, C.; Birkenmeier, E.; Ben-Zeev, O.; Schotz, M.; Sweet, H.; Davisson, M.; Gordon, J. The Fatty Liver Dystrophy (fld) Mutation: A New Mutant Mouse with a Developmental Abnormality in Triglyceride Metabolism and Associated Tissue-Specific Defects in Lipoprotein Lipase and Hepatic Lipase Activities. J. Biol. Chem. 1989, 264, 7994–8003. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Fan, X.; Miao, Y. LPIN1 Promotes Triglycerides Synthesis and Is Transcriptionally Regulated by PPARG in Buffalo Mammary Epithelial Cells. Sci. Rep. 2022, 12, 2390. [Google Scholar] [CrossRef]
- Zhang, W.; Zhong, W.; Sun, Q.; Sun, X.; Zhou, Z. Adipose-Specific Lipin1 Overexpression in Mice Protects against Alcohol-Induced Liver Injury. Sci. Rep. 2018, 8, 408. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Kashiwagi, Y.; Arimitsu, N.; Kogure, T.; Edo, A.; Maruyama, T.; Nakao, K.; Nakanishi, H.; Kinoshita, M.; Frohman, M.A.; et al. Phosphatidic Acid (PA)-Preferring Phospholipase A1 Regulates Mitochondrial Dynamics*. J. Biol. Chem. 2014, 289, 11497–11511. [Google Scholar] [CrossRef]
- Huang, H.; Gao, Q.; Peng, X.; Choi, S.Y.; Sarma, K.; Ren, H.; Morris, A.J.; Frohman, M.A. piRNA-Associated Germline Nuage Formation and Spermatogenesis Require MitoPLD Profusogenic Mitochondrial-Surface Lipid Signaling. Dev. Cell 2011, 20, 376–387. [Google Scholar] [CrossRef]
- Shimada, K.; Kato, H.; Miyata, H.; Ikawa, M. Glycerol Kinase 2 Is Essential for Proper Arrangement of Crescent-like Mitochondria to Form the Mitochondrial Sheath during Mouse Spermatogenesis. J. Reprod. Dev. 2019, 65, 155–162. [Google Scholar] [CrossRef]
- Zhang, Z.; Miao, J.; Wang, Y. Mitochondrial Regulation in Spermatogenesis. Reproduction 2022, 163, R55–R69. [Google Scholar] [CrossRef]
- Nixon, B.; Bernstein, I.R.; Cafe, S.L.; Delehedde, M.; Sergeant, N.; Anderson, A.L.; Trigg, N.A.; Eamens, A.L.; Lord, T.; Dun, M.D.; et al. A Kinase Anchor Protein 4 Is Vulnerable to Oxidative Adduction in Male Germ Cells. Front. Cell Dev. Biol. 2019, 7, 319. [Google Scholar] [CrossRef]
- Jumeau, F.; Sigala, J.; Dossou-Gbete, F.; Frimat, K.; Barbotin, A.L.; Buée, L.; Béhal, H.; Sergeant, N.; Mitchell, V. A-Kinase Anchor Protein 4 Precursor (pro-AKAP4) in Human Spermatozoa. Andrology 2018, 6, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Scovell, J.M.; Bournat, J.C.; Szafran, A.T.; Solis, M.; Moore, J.; Rivera, A.; Chen, C.H.; Zhang, J.; Wilken, N.; Seth, A.; et al. PRSS50 Is a Testis Protease Responsible for Proper Sperm Tail Formation and Function. Development 2021, 148, dev197558. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Jiang, T.X.; Chen, L.B.; Zhou, W.; Liu, Y.; Gao, F.; Qiu, X.B. Proteasome Subunit α4s Is Essential for Formation of Spermatoproteasomes and Histone Degradation during Meiotic DNA Repair in Spermatocytes. J. Biol. Chem. 2021, 296, 100130. [Google Scholar] [CrossRef]
- Ryu, K.Y.; Sinnar, S.A.; Reinholdt, L.G.; Vaccari, S.; Hall, S.; Garcia, M.A.; Zaitseva, T.S.; Bouley, D.M.; Boekelheide, K.; Handel, M.A.; et al. The Mouse Polyubiquitin Gene Ubb Is Essential for Meiotic Progression. Mol. Cell. Biol. 2008, 28, 1136–1146. [Google Scholar] [CrossRef]
- Han, B.; Jung, B.-K.; Park, S.-H.; Song, K.J.; Anwar, M.A.; Ryu, K.-Y.; Kim, K.P. Polyubiquitin Gene Ubb Is Required for Upregulation of Piwi Protein Level during Mouse Testis Development. Cell Death Discov. 2021, 7, 194. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Kaufman, A.J.; Ryan, R.J.H.; Romin, Y.; Huryn, L.; Bains, S.; Manova-Todorova, K.; Morris, P.L.; Hunnicutt, G.R.; Adelman, C.A.; et al. Mouse DCUN1D1 (SCCRO) Is Required for Spermatogenetic Individualization. PLoS ONE 2019, 14, e0209995. [Google Scholar] [CrossRef] [PubMed]
- Nagata, O.; Nakamura, M.; Sakimoto, H.; Urata, Y.; Sasaki, N.; Shiokawa, N.; Sano, A. Mouse Model of Chorea-Acanthocytosis Exhibits Male Infertility Caused by Impaired Sperm Motility as a Result of Ultrastructural Morphological Abnormalities in the Mitochondrial Sheath in the Sperm Midpiece. Biochem. Biophys. Res. Commun. 2018, 503, 915–920. [Google Scholar] [CrossRef]
- Jiang, M.; Gao, M.; Wu, C.; He, H.; Guo, X.; Zhou, Z.; Yang, H.; Xiao, X.; Liu, G.; Sha, J. Lack of Testicular Seipin Causes Teratozoospermia Syndrome in Men. Proc. Natl. Acad. Sci. USA 2014, 111, 7054–7059. [Google Scholar] [CrossRef]
- Varuzhanyan, G.; Rojansky, R.; Sweredoski, M.J.; Graham, R.L.; Hess, S.; Ladinsky, M.S.; Chan, D.C. Mitochondrial Fusion Is Required for Spermatogonial Differentiation and Meiosis. eLife 2019, 8, e51601. [Google Scholar] [CrossRef] [PubMed]
- Varuzhanyan, G.; Chen, H.; Rojansky, R.; Ladinsky, M.S.; McCaffery, J.M.; Chan, D.C. Mitochondrial Fission Factor (Mff) Is Required for Organization of the Mitochondrial Sheath in Spermatids. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129845. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liang, P.; Huang, Y.; Li, M.; Zhang, X.; Ding, C.; Feng, J.; Zhang, Z.; Zhang, X.; Gao, Y.; et al. Glycerol Kinase-like Proteins Cooperate with Pld6 in Regulating Sperm Mitochondrial Sheath Formation and Male Fertility. Cell Discov. 2017, 3, 17030. [Google Scholar] [CrossRef] [PubMed]
- Ben Khelifa, M.; Coutton, C.; Zouari, R.; Karaouzène, T.; Rendu, J.; Bidart, M.; Yassine, S.; Pierre, V.; Delaroche, J.; Hennebicq, S.; et al. Mutations in DNAH1, Which Encodes an Inner Arm Heavy Chain Dynein, Lead to Male Infertility from Multiple Morphological Abnormalities of the Sperm Flagella. Am. J. Hum. Genet. 2014, 94, 95–104. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, B.; Kim, J.; Morohoshi, K.; Kang, W.; Miyado, K.; Tsuruta, F.; Kawano, N.; Chiba, T. Proteasome-Associated Proteins, PA200 and ECPAS, Are Essential for Murine Spermatogenesis. Biomolecules 2023, 13, 586. https://doi.org/10.3390/biom13040586
Sato B, Kim J, Morohoshi K, Kang W, Miyado K, Tsuruta F, Kawano N, Chiba T. Proteasome-Associated Proteins, PA200 and ECPAS, Are Essential for Murine Spermatogenesis. Biomolecules. 2023; 13(4):586. https://doi.org/10.3390/biom13040586
Chicago/Turabian StyleSato, Ban, Jiwoo Kim, Kazunori Morohoshi, Woojin Kang, Kenji Miyado, Fuminori Tsuruta, Natsuko Kawano, and Tomoki Chiba. 2023. "Proteasome-Associated Proteins, PA200 and ECPAS, Are Essential for Murine Spermatogenesis" Biomolecules 13, no. 4: 586. https://doi.org/10.3390/biom13040586
APA StyleSato, B., Kim, J., Morohoshi, K., Kang, W., Miyado, K., Tsuruta, F., Kawano, N., & Chiba, T. (2023). Proteasome-Associated Proteins, PA200 and ECPAS, Are Essential for Murine Spermatogenesis. Biomolecules, 13(4), 586. https://doi.org/10.3390/biom13040586